Inhibition of Ihh Reverses Temporomandibular Joint Osteoarthritis via a PTH1R Signaling Dependent Mechanism

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
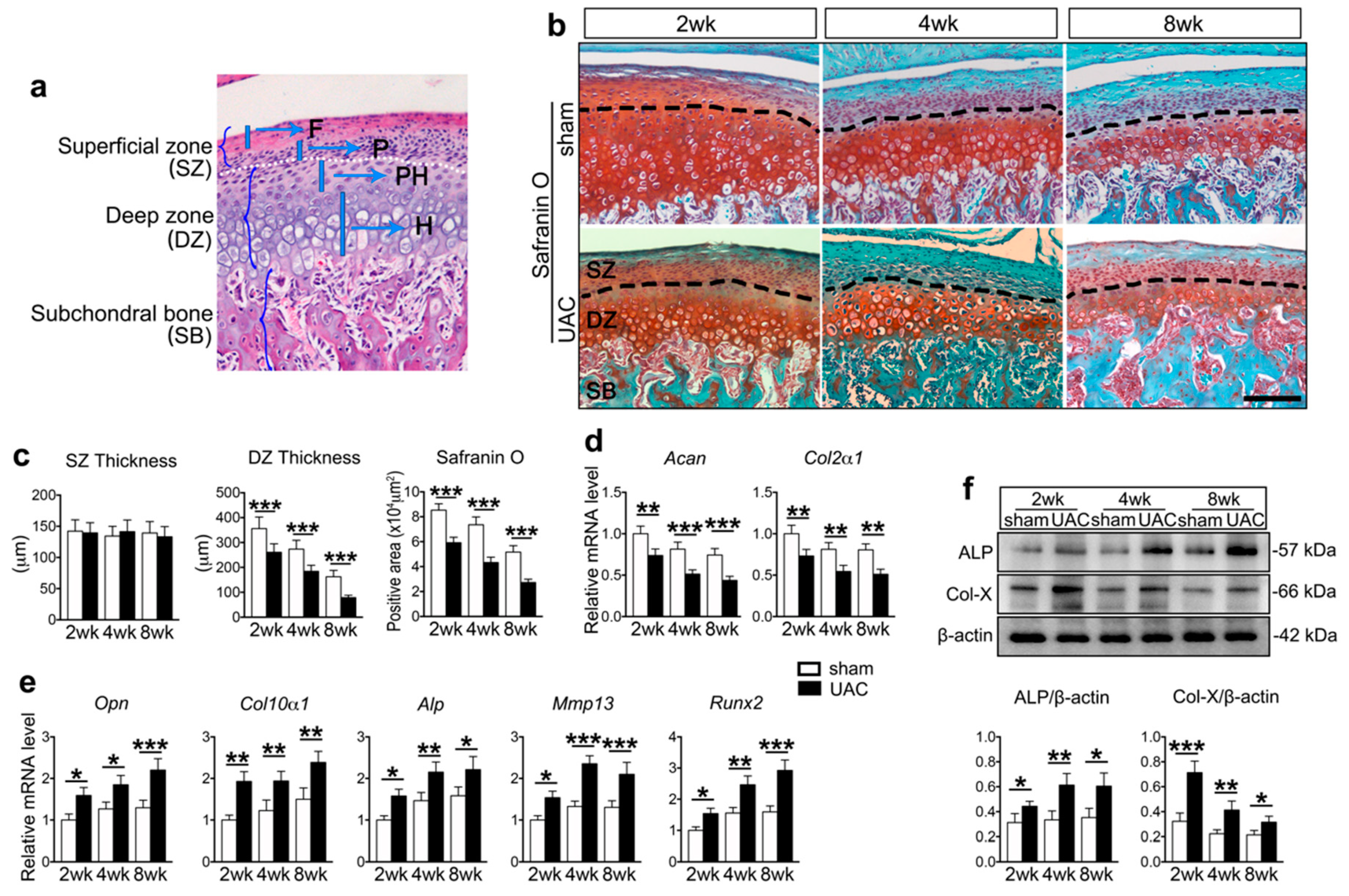
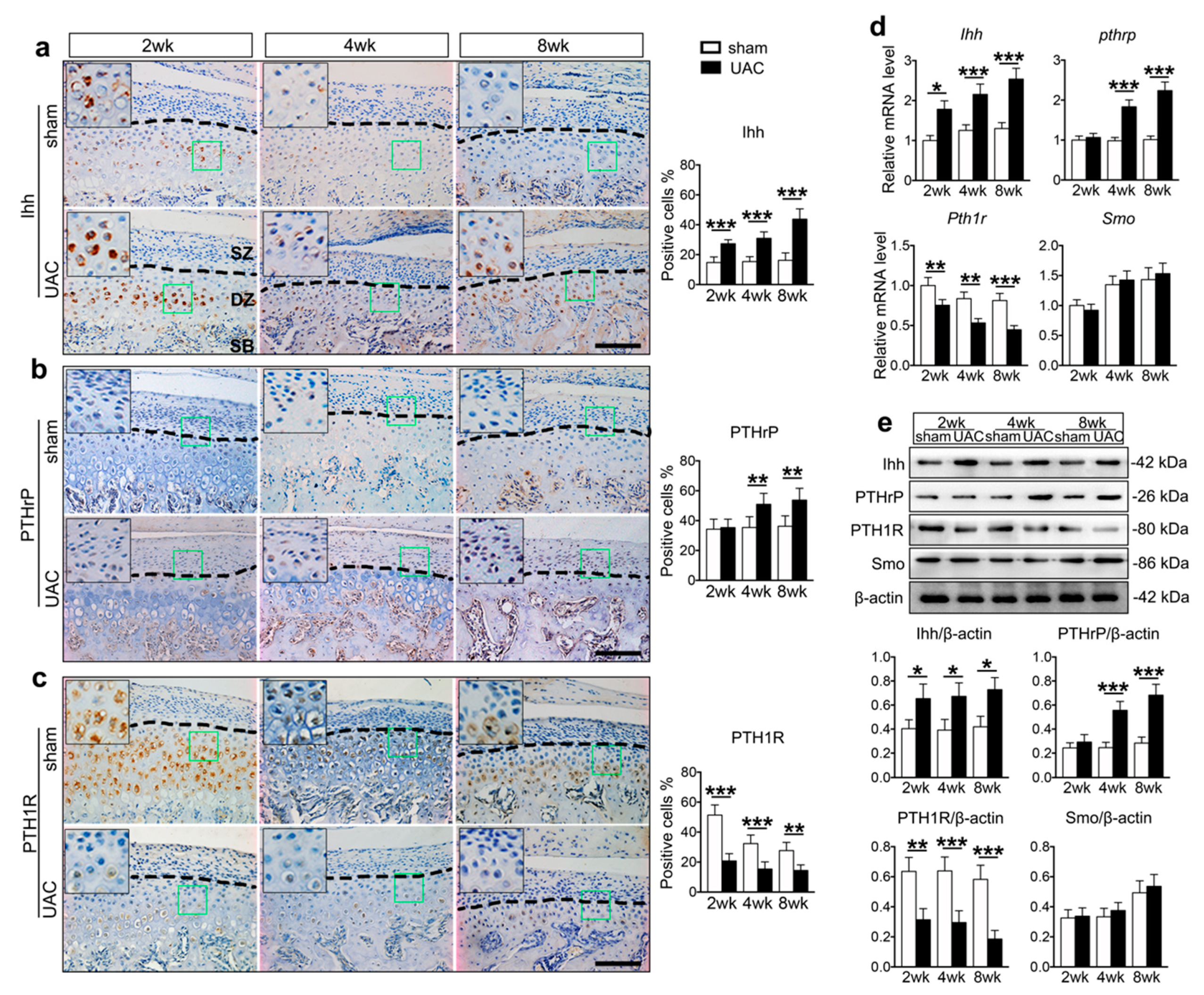
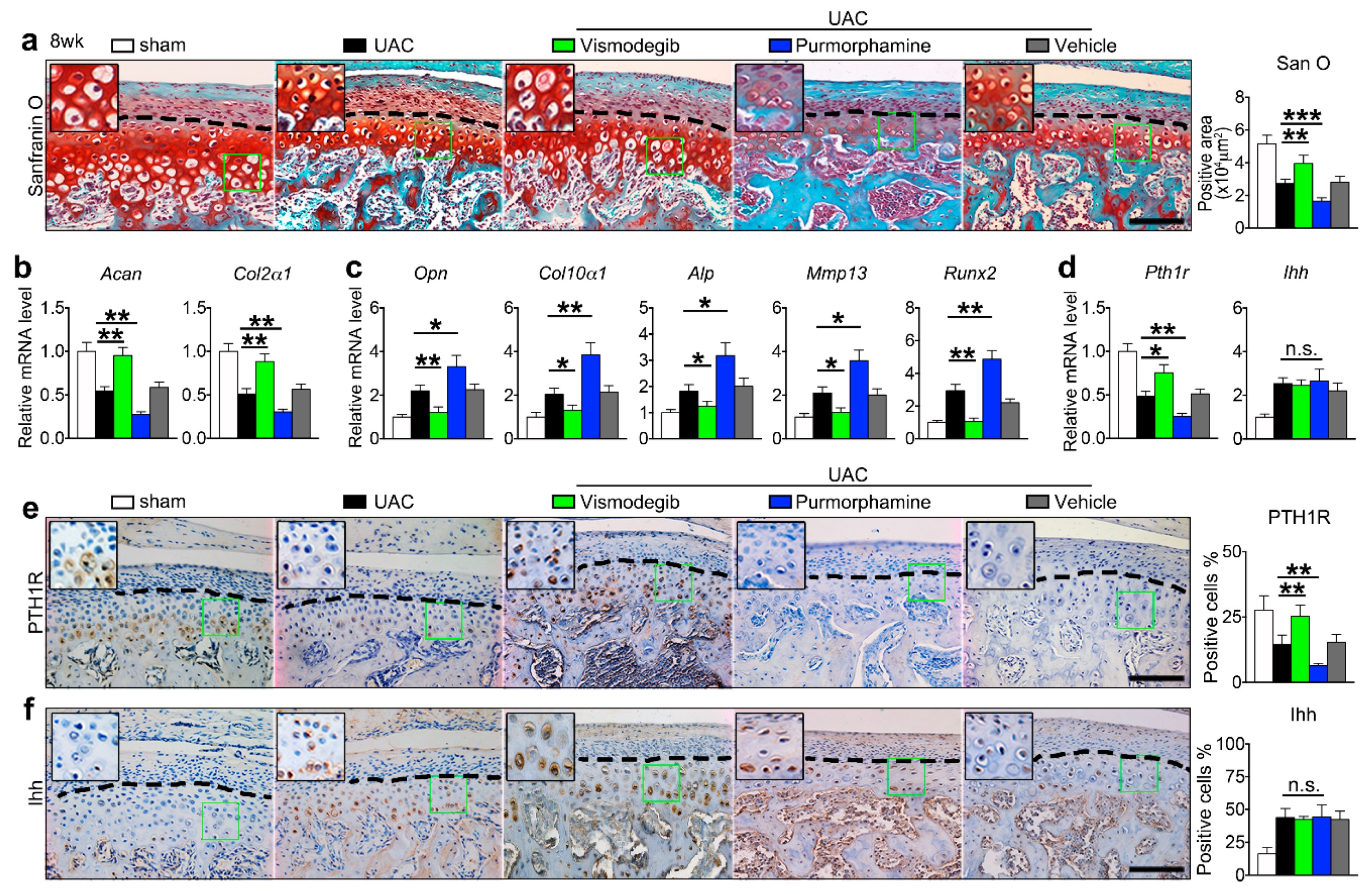
2.1. UAC Stimulated Chondrocyte Terminal Differentiation with the Modulation of Ihh and PTH1R Signaling
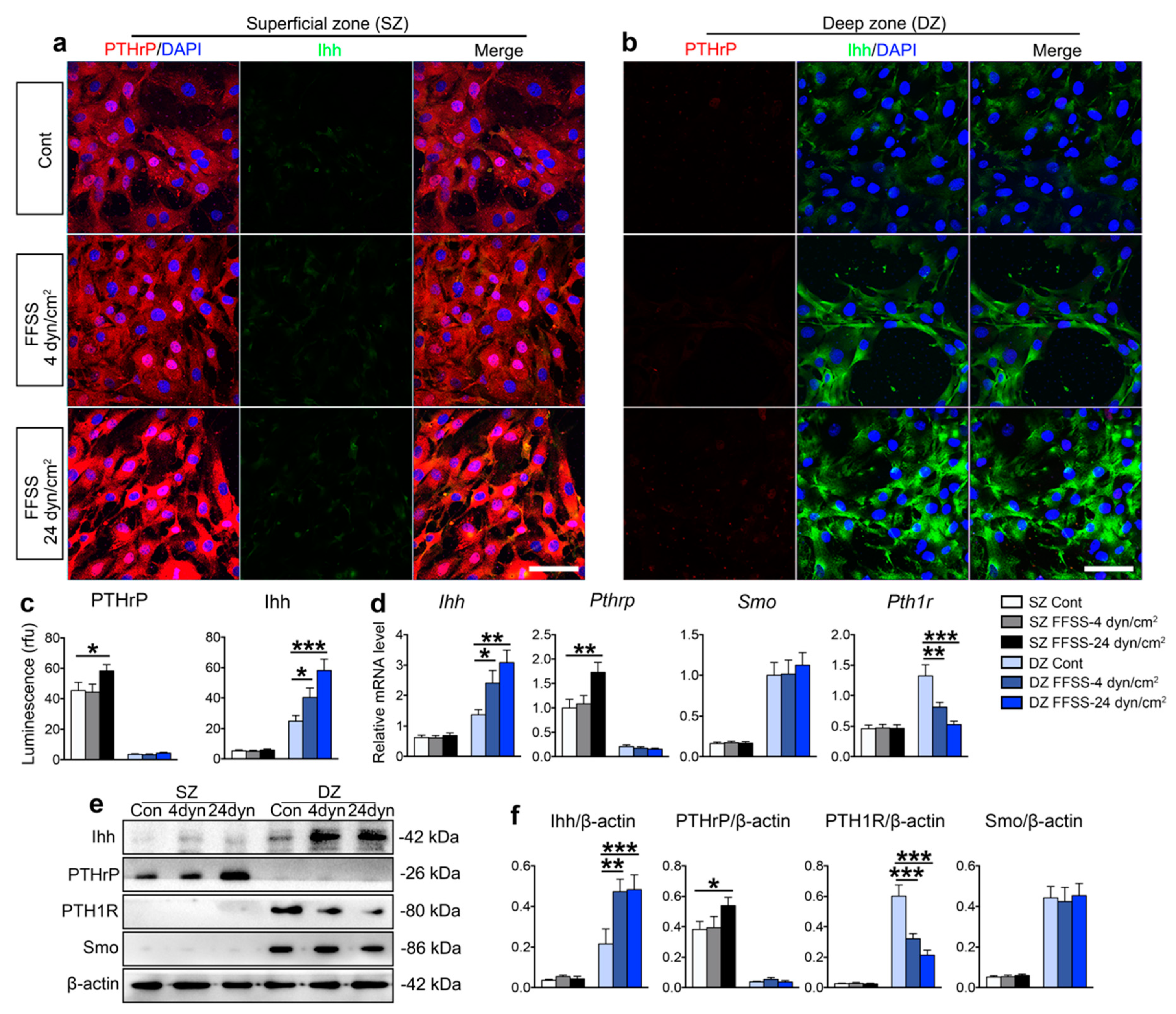
2.2. FFSS Stimulated Deep Zone Chondrocytes Terminal Differentiation with Activated Ihh Signaling but Suppressed PTH1R Expression
2.3. Suppressing Ihh Signaling Rescued FFSS and UAC Stimulated Chondrocytes Terminal Differentiation
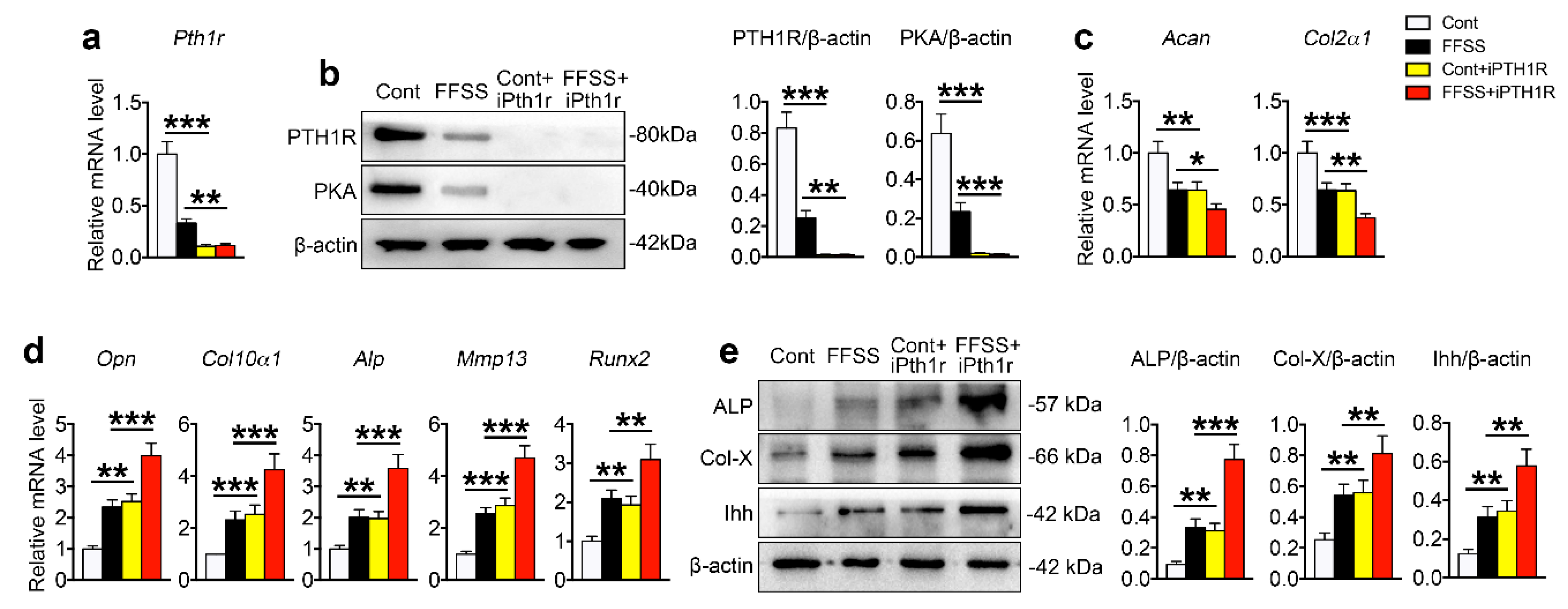
2.4. Reduction of PTH1R directly Promoted FFSS and UAC Stimulated Chondrocyte Terminal Differentiation
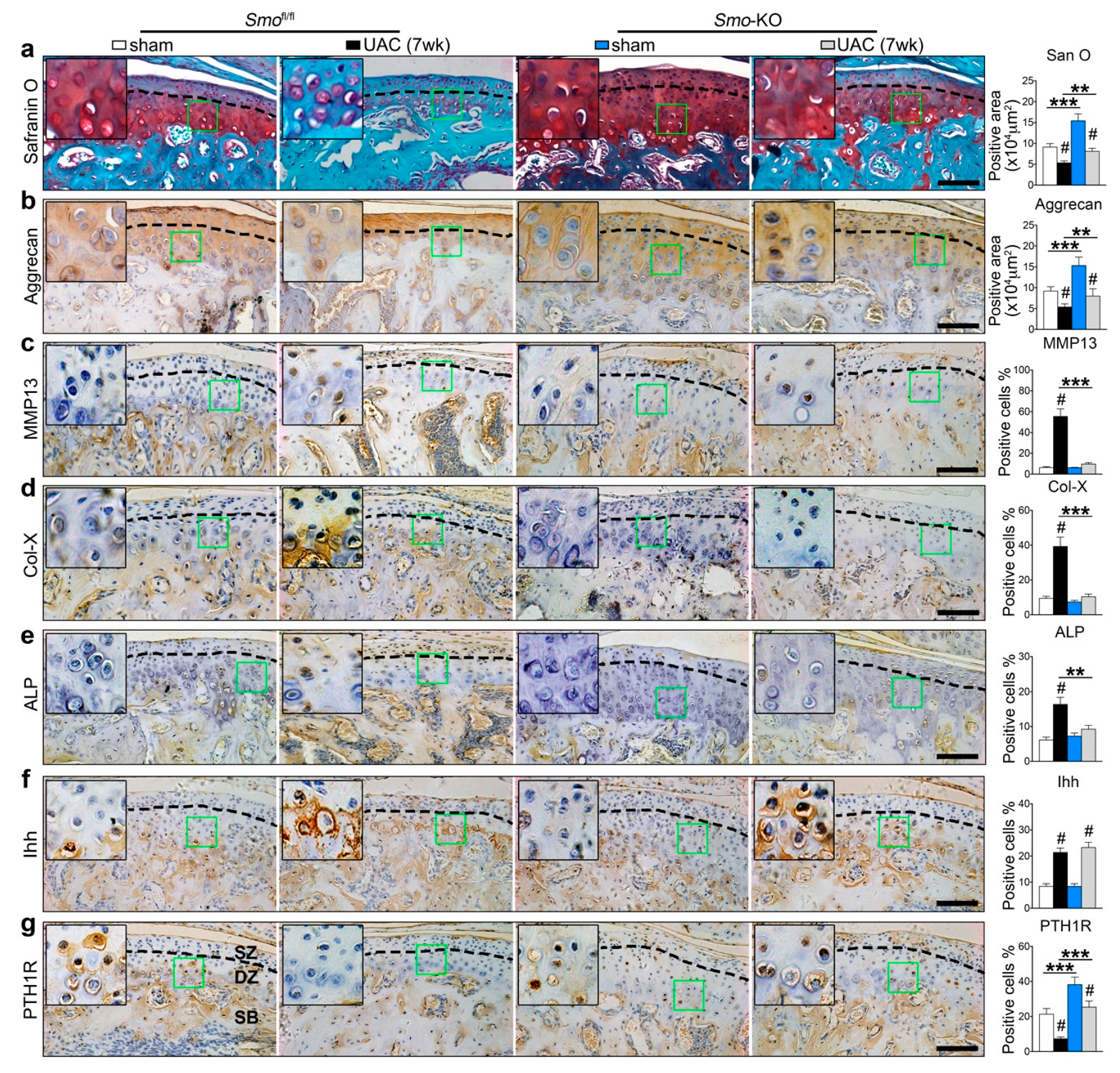
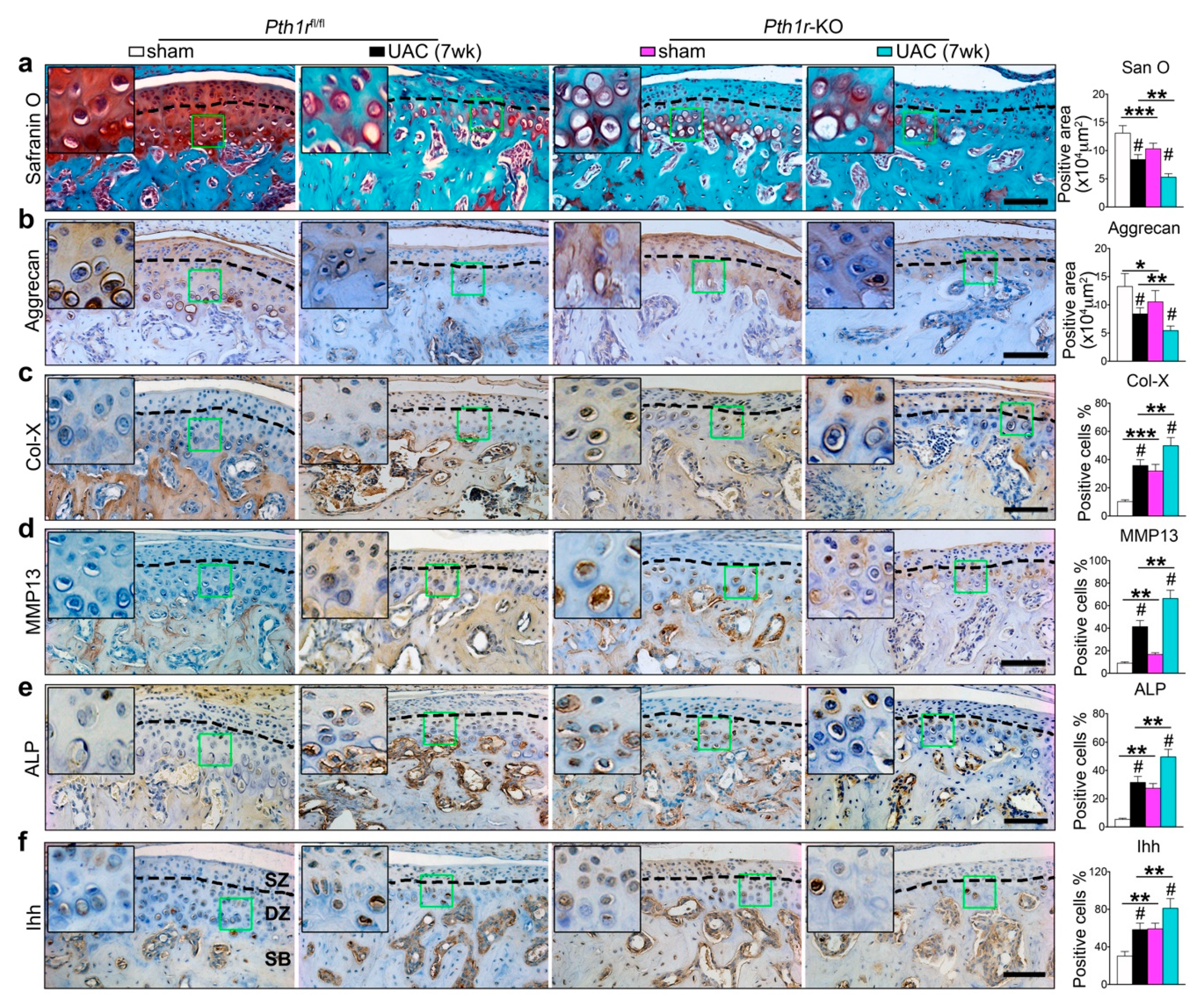
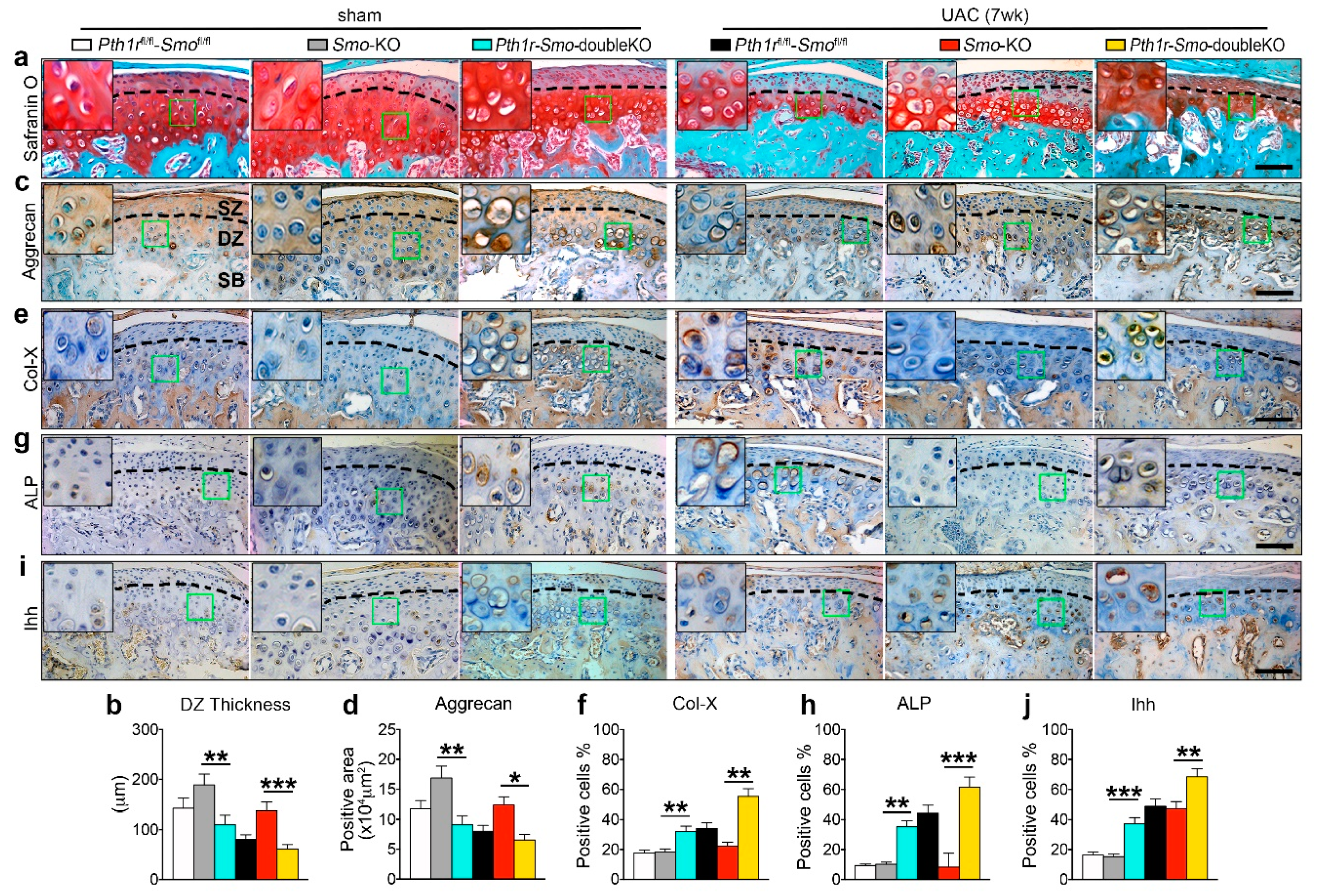
2.5. Inhibition of Ihh Prevented UAC Induced Chondrocyte Terminal Differentiation through a PTH1R Dependent Mechanism
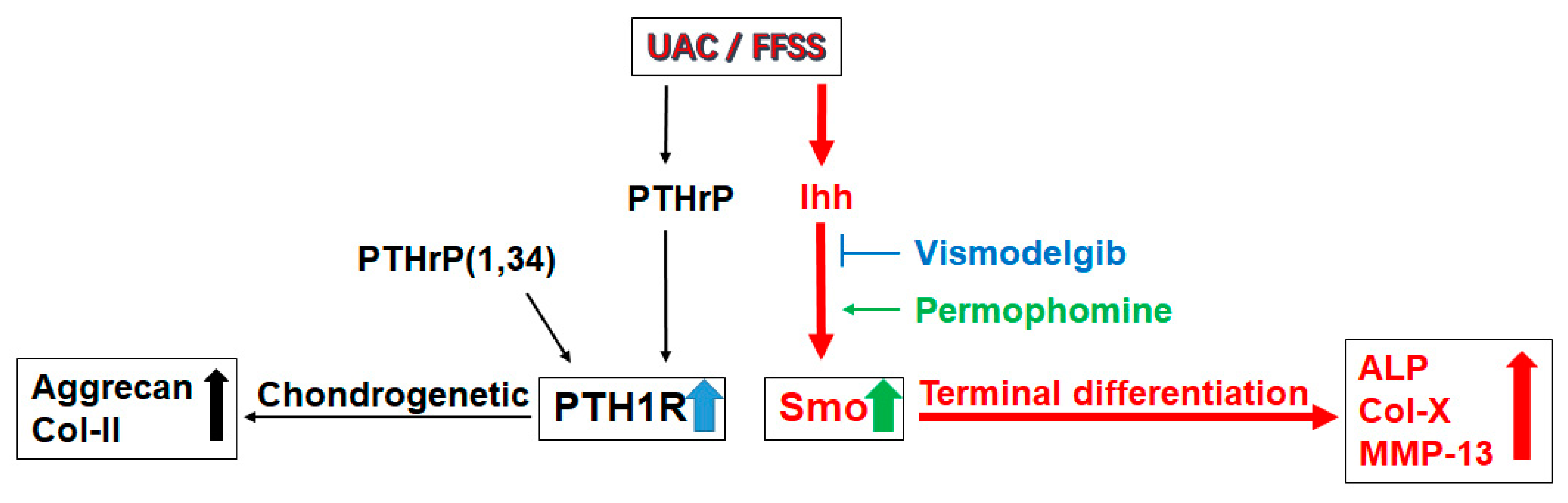
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Tamoxifen Administration and Genotyping
4.3. UAC Model and Tissue Preparation
4.4. Primary Chondrocyte Culture and Fluid Flow Shear Stress
4.5. siRNA Transfection for Pth1r Knockdown
4.6. Histomorphology and Immunohistochemical Staining
4.7. Immunoblotting, PCR and qRT-PCR
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhen, G.; Wen, C.; Jia, X. Inhibition of TGF-β signaling in subchondral bone mesenchymal stem cells attenuates osteoarthritis. Nat. Med. 2013, 19, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Kamper, M.; Paulsson, M.; Zaucke, F. Absence of collagen IX accelerates hypertrophic differentiation in the embryonic mouse spine through a disturbance of the Ihh-PTHrP feedback loop. Cell. Tissue. Res. 2017, 367, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dong, S. The signaling pathways involved in chondrocyte differentiation and hypertrophic differentiation. Stem. Cells. Int. 2016, 2016, 2470351. [Google Scholar] [CrossRef] [PubMed]
- Rhee, J.; Park, S.H.; Kim, S.K.; Kim, J.H.; Ha, C.W.; Chun, C.H.; Chun, J.S. Inhibition of BATF/JUN transcriptional activity protects against osteoarthritic cartilage destruction. Ann. Rheum. Dis. 2017, 76, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Van der Kraan, P.M.; van den Berg, W.B. Chondrocyte hypertrophy and osteoarthritis: Role in initiation and progression of cartilage degeneration? Osteoarthr. Cartil. 2012, 20, 223–232. [Google Scholar] [CrossRef]
- Kozhemyakina, E.; Lassar, A.B.; Zelzer, E. A pathway to bone: Signaling molecules and transcription factors involved in chondrocyte development and maturation. Development 2015, 142, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Zhou, J.; Wei, X.; Zhang, J.; Fleming, B.C.; Terek, R.; Pei, M.; Chen, Q.; Liu, T.; Wei, L. Activation of Indian hedgehog promotes chondrocyte hypertrophy and upregulation of MMP-13 in human osteoarthritic cartilage. Osteoarthr. Cartil. 2012, 20, 755–763. [Google Scholar] [CrossRef]
- Lum, L.; Beachy, P.A. The hedgehog response network: Sensors, switches, and routers. Science 2004, 304, 1755–1759. [Google Scholar] [CrossRef]
- Strewler, G.J. The physiology of parathyroid hormone-related protein. N. Engl. J. Med. 2000, 342, 177–185. [Google Scholar] [CrossRef]
- Becher, C.; Szuwart, T.; Ronstedt, P.; Ostermeier, S.; Skwara, A.; Fuchs-Winkelmann, S.; Tibesku, C.O. Decrease in the expression of the type 1 PTH/PTHrP receptor (PTH1R) on chondrocytes in animals with osteoarthritis. J. Orthop. Surg. Res. 2010, 5, 28. [Google Scholar] [CrossRef]
- Buckland, J. Osteoarthritis: Blocking hedgehog signaling might have therapeutic potential in OA. Nat. Rev. Rheumatol. 2010, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- Alman, B.A. The role of hedgehog signalling in skeletal health and disease. Nat. Rev. Rheumatol. 2015, 11, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Dentice, M.; Bandyopadhyay, A.; Gereben, B.; Callebaut, I.; Christoffolete, M.A.; Kim, B.W.; Nissim, S.; Mornon, J.P.; Zavacki, A.M.; Zeöld, A.; et al. The Hedgehog-inducible ubiquitin ligase subunit WSB-1 modulates thyroid hormone activation and PTHrP secretion in the developing growth plate. Nat. Cell. Biol. 2005, 7, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Taboas, J.M.; Tuan, R.S. PTHrP overexpression partially inhibits a mechanical strain-induced arthritic phenotype in chondrocytes. Osteoarthr. Cartil. 2011, 19, 213–221. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chang, J.K.; Chang, L.H.; Hung, S.H.; Wu, S.C.; Lee, H.Y.; Lin, Y.S.; Chen, C.H.; Fu, Y.C.; Wang, G.J.; Ho, M.L. Parathyroid Hormone 1-34 Inhibits Terminal Differentiation of Human Articular Chondrocytes and Osteoarthritis Progression in Rats. Arthritis. Rheum. 2009, 60, 3049–3060. [Google Scholar] [CrossRef] [PubMed]
- Luder, H.U.; Leblond, C.P.; von der Mark, K. Cellular stages in cartilage formation as revealed by morphometry, radioautography and type II collagen immunostaining of the mandibular condyle from weanling rats. Am. J. Anat. 1988, 182, 197–214. [Google Scholar] [CrossRef]
- Hasegawa, S.; Kitoh., H.; Ohkawara, B.; Mishima, K.; Matsushita, M.; Masuda, A.; Ishiguro, N.; Ohno, K. Tranilast stimulates endochondral ossification by upregulating SOX9 and RUNX2 promoters. Biochem. Biophys. Res. Commun. 2016, 470, 356–361. [Google Scholar] [CrossRef]
- Mak, K.K.; Kronenberg, H.M.; Chuang, P.T.; Mackem, S.; Yang, Y. Indian hedgehog signals independently of PTHrP to promote chondrocyte hypertrophy. Development 2008, 135, 1947–1956. [Google Scholar] [CrossRef]
- Qiu, T.; Xian, L.; Crane, J.; Wen, C.; Hilton, M.; Lu, W.; Newman, P.; Cao, X. PTH receptor signaling in osteoblasts regulates endochondral vascularization in maintenance of postnatal growth plate. J. Bone Miner. Res. 2015, 30, 309–317. [Google Scholar] [CrossRef]
- Tsutsui, T.W.; Riminucci, M.; Holmbeck, K.; Bianco, P.; Robey, P.G. Development of craniofacial structures in transgenic mice with constitutively active PTH/PTHrP receptor. Bone 2008, 42, 321–331. [Google Scholar] [CrossRef]
- Scrivani, S.J.; Keith, D.A.; Kaban, L.B. Temporomandibular Disorders. N. Engl. J. Med. 2008, 359, 2693–2705. [Google Scholar] [CrossRef] [PubMed]
- Shibukawa, Y. Temporomandibular joint formation and condyle growth require Indian hedgehog signaling. Temporomandibular joint formation and condyle growth require Indian hedgehog signaling. Dev. Dyn. 2007, 236, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.X.; Zhang, M.; Wang, X.; Zhang, H.; Zhang, J.; Jing, L.; Liao, L.; Wang, M. TNF Accelerates Death of Mandibular Condyle Chondrocytes in Rats with Biomechanical Stimulation-Induced Temporomandibular Joint Disease. PLoS ONE 2015, 10, e0141774. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, H.; Zhang, J.; Zhang, H.; Yang, H.; Wan, X.; Jing, L.; Lu, L.; Liu, X.; Yu, S.; et al. Unilateral anterior crossbite induces aberrant mineral deposition in degenerative temporomandibular cartilage in rats. Osteoarthr. Cartil. 2016, 24, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yang, H.X.; Wan, X.H.; Zhang, M.; Zhang, J.; Lu, L.; Xie, M.; Ren, H.T.; Yu, S.B.; Liu, X.D.; et al. Calcium-/calmodulin-dependent protein kinase II in occlusion-induced degenerative cartilage of rat mandibular condyle. J. Oral Rehabil. 2018, 45, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, H.; Lu, L.; Wan, X.; Zhang, J.; Zhang, H.; Liu, X.; Huang, X.; Xiao, G.; Wang, M. Matrix replenishing by BMSCs is beneficial for osteoarthritic temporomandibular joint cartilage. Osteoarthr. Cartil. 2017, 25, 1551–1562. [Google Scholar] [CrossRef]
- Liu, Y.D.; Yang, H.X.; Liao, L.F.; Jiao, K.; Zhang, H.Y.; Lu, L.; Zhang, M.; Zhang, J.; He, J.J.; Wu, Y.P.; et al. Systemic administration of strontium or NBD peptide ameliorates early stage cartilage degradation of mouse mandibular condyles. Osteoarthr. Cartil. 2016, 24, 178–187. [Google Scholar] [CrossRef]
- Zhang, J.; Liao, L.; Zhu, J.; Wan, X.; Xie, M.; Zhang, H.; Zhang, M.; Lu, L.; Yang, H.; Jing, D.; et al. Osteochondral Interface Stiffening in Mandibular Condylar Osteoarthritis. J. Dent. Res. 2018, 97, 563–570. [Google Scholar] [CrossRef]
- Jiao, K.; Niu, L.; Xu, X.; Liu, Y.; Li, X.; Tay, F.R.; Wang, M. Norepinephrine Regulates Condylar Bone Loss via Comorbid Factors. J. Dent. Res. 2015, 94, 813–820. [Google Scholar] [CrossRef]
- Lu, L.; Zhang, X.; Zhang, M.; Zhang, H.; Liao, L.; Yang, T.; Zhang, J.; Xian, L.; Chen, D.; Wang, M. RANTES and SDF-1 Are Keys in Cell-based Therapy of Osteoarthritis of the Temporomandibular Joint. J. Dent. Res. 2015, 94, 1601–1609. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.Y.; Zhang, M.; Qiu, Z.Y.; Wu, Y.P.; Callaway, D.A.; Jiang, J.X.; Lu, L.; Jing, L.; Yang, T.; et al. Connexin43 hemichannels mediate small molecule exchange between chondrocytes and matrix in biomechanically-stimulated temporomandibular joint cartilage. Osteoarthr. Cartil. 2014, 22, 822–830. [Google Scholar] [CrossRef]
- Yang, H.X.; Wen, Y.; Zhang, M.; Liu, Q.; Zhang, H.Y.; Zhang, J.; Lu, L.; Ye, T.; Bai, X.C.; Xiao, G.Z.; et al. MTORC1 coordinates the autophagy and apoptosis signaling in articular chondrocytes in osteoarthritic temporomandibular joint. Autophagy 2019, 21, 1–18. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, J.; Lu, L.; Qiu, Z.Y.; Zhang, X.; Yu, S.B.; Wu, Y.P.; Wang, M.Q. Enhancement of chondrocyte autophagy is an early response in the degenerative cartilage of the temporomandibular joint to biomechanical dental stimulation. Apoptosis 2013, 18, 423–434. [Google Scholar] [CrossRef]
- Long, F.; Zhang, X.M.; Karp, S.; Yang, Y.; McMahon, A.P. Genetic manipulation of hedgehog signaling in the endochondral skeleton reveals a direct role in the regulation of chondrocyte proliferation. Development 2001, 128, 5099–5108. [Google Scholar]
- Hirai, T.; Chagin, A.S.; Kobayashi, T.; Mackem, S.; Kronenberg, H.M. Parathyroid hormone/parathyroid hormone-related protein receptor signaling is required for maintenance of the growth plate in postnatal life. Proc. Natl. Acad. Sci. USA 2011, 108, 191–196. [Google Scholar] [CrossRef]
- Pucci, R.; Vellone, V.; Ramieri, V.; Cascone, P.; Della Rocca, C. Histological findings in TMJ treated with high condilectomy for internal derangement. J. Craniomaxillofac. Surg. 2018, 5182, 30223. [Google Scholar] [CrossRef]
- Dutra, E.H.; O’Brien, M.H.; Lima, A.; Nanda, R.; Yadav, S. A morphometric and cellular analysis method for the murine mandibular condyle. J. Vis. Exp. 2018, 11, 131. [Google Scholar] [CrossRef]
- Day, T.F.; Yang, Y. Wnt and hedgehog signaling pathways in bone development. J. Bone Jt. Surg. Am. 2008, 90, 19–24. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, H.; Zeng, C.; Yan, B.; Ouyang, J.; Liu, X.; Sun, Q.; Zhao, C.; Fang, H.; Pan, J.; et al. mTORC1 activation downregulates FGFR3 and PTH/PTHrP receptor in articular chondrocytes to initiate osteoarthritis. Osteoarthr. Cartil. 2017, 25, 952–963. [Google Scholar] [CrossRef]
- Vortkamp, A.; Lee, K.; Lanske, B.; Segre, G.V.; Kronenberg, H.M.; Tabin, C.J. Regulation of rate of cartilage differentiation by Indian hedgehog and PTH-related protein. Science 1996, 273, 613–622. [Google Scholar] [CrossRef]
- Hilton, M.J.; Tu, X.; Long, F. Tamoxifen-inducible gene deletion reveals a distinct cell type associated with trabecular bone, and direct regulation of PTHrP expression and chondrocyte morphology by Ihh in growth region cartilage. Dev. Biol. 2007, 308, 93–105. [Google Scholar] [CrossRef]
- Bravo, B.; Argüello, J.M.; Gortazar, A.R.; Forriol, F.; Vaquero, J. Modulation of Gene Expression in Infrapatellar Fat Pad-Derived Mesenchymal Stem Cells in Osteoarthritis. Cartilage 2018, 9, 55–62. [Google Scholar] [CrossRef]
- Aicher, W.K.; Rolauffs, B. The spatial organisation of joint surface chondrocytes: Review of its potential roles in tissue functioning, disease and early, preclinical diagnosis of osteoarthritis. Ann. Rheum. Dis. 2014, 73, 645–653. [Google Scholar] [CrossRef]
- Chen, D.; Shen, J.; Zhao, W.; Wang, T.; Han, L.; Hamilton, J.L.; Im, H.J. Osteoarthritis: Toward a comprehensive understanding of pathological mechanism. Bone Res. 2017, 5, 16044. [Google Scholar] [CrossRef]
- Thomas, C.M.; Fuller, C.J.; Whittles, C.E.; Sharif, M. Chondrocyte death by apoptosis is associated with cartilage matrix degradation. Osteoarthr. Cartil. 2007, 15, 27–34. [Google Scholar] [CrossRef]
- Singh, M.; Detamore, M.S. Biomechanical properties of the mandibular condylar cartilage and their relevance to the TMJ disc. J. Biomech. 2009, 42, 405–417. [Google Scholar] [CrossRef]
- Embree, M.C.; Chen, M.; Pylawka, S.; Kong, D.; Iwaoka, G.M.; Kalajzic, I.; Yao, H.; Shi, C.; Sun, D.; Sheu, T.J.; et al. Exploiting endogenous fibrocartilage stem cells to regenerate cartilage and repair joint injury. Nat. Commun. 2016, 7, 13073. [Google Scholar] [CrossRef]
- Maeda, Y.; Schipani, E.; Densmore, M.; Lanske, B. Partial rescue of postnatal growth plate abnormalities in Ihh mutants by expression of a constitutively active PTH/PTHrP receptor. Bone 2010, 46, 472–478. [Google Scholar] [CrossRef]
- Chen, J.; Kamiya, Y.; Polur, I.; Xu, M.; Choi, T.; Kalajzic, Z.; Drissi, H.; Wadhwa, S. Estrogen via Estrogen Receptor Beta Partially Inhibits Mandibular Condylar Cartilage Growth. Osteoarthr. Cartil. 2014, 22, 1861–1868. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Zhang, M.; Liu, Q.; Zhang, H.; Zhang, J.; Lu, L.; Xie, M.; Chen, D.; Wang, M. Inhibition of Ihh Reverses Temporomandibular Joint Osteoarthritis via a PTH1R Signaling Dependent Mechanism. Int. J. Mol. Sci. 2019, 20, 3797. https://doi.org/10.3390/ijms20153797
Yang H, Zhang M, Liu Q, Zhang H, Zhang J, Lu L, Xie M, Chen D, Wang M. Inhibition of Ihh Reverses Temporomandibular Joint Osteoarthritis via a PTH1R Signaling Dependent Mechanism. International Journal of Molecular Sciences. 2019; 20(15):3797. https://doi.org/10.3390/ijms20153797
Chicago/Turabian StyleYang, Hongxu, Mian Zhang, Qian Liu, Hongyun Zhang, Jing Zhang, Lei Lu, Mianjiao Xie, Di Chen, and Meiqing Wang. 2019. "Inhibition of Ihh Reverses Temporomandibular Joint Osteoarthritis via a PTH1R Signaling Dependent Mechanism" International Journal of Molecular Sciences 20, no. 15: 3797. https://doi.org/10.3390/ijms20153797
APA StyleYang, H., Zhang, M., Liu, Q., Zhang, H., Zhang, J., Lu, L., Xie, M., Chen, D., & Wang, M. (2019). Inhibition of Ihh Reverses Temporomandibular Joint Osteoarthritis via a PTH1R Signaling Dependent Mechanism. International Journal of Molecular Sciences, 20(15), 3797. https://doi.org/10.3390/ijms20153797