Overexpression of Notch Signaling Induces Hyperosteogeny in Zebrafish




Abstract
1. Introduction
2. Result
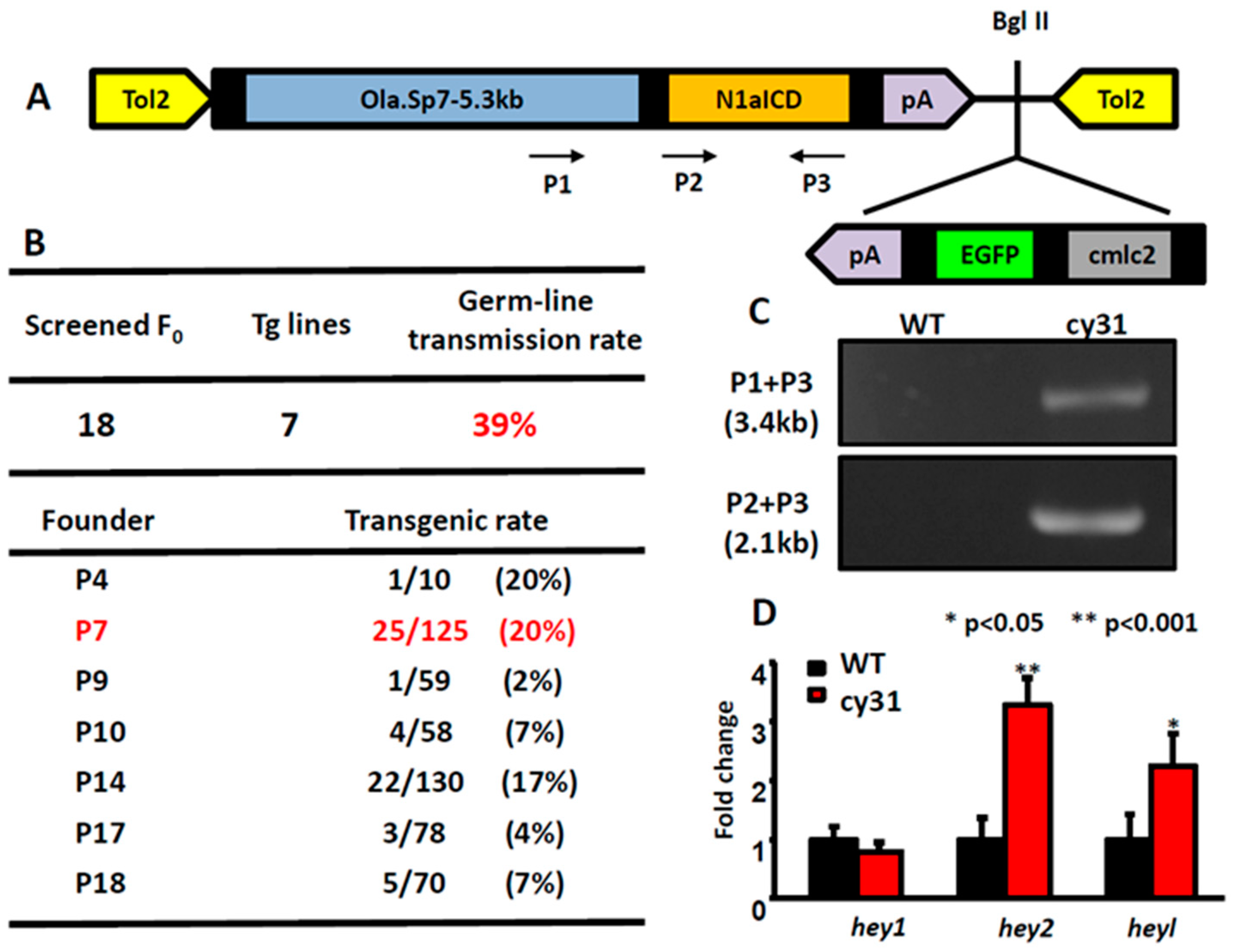
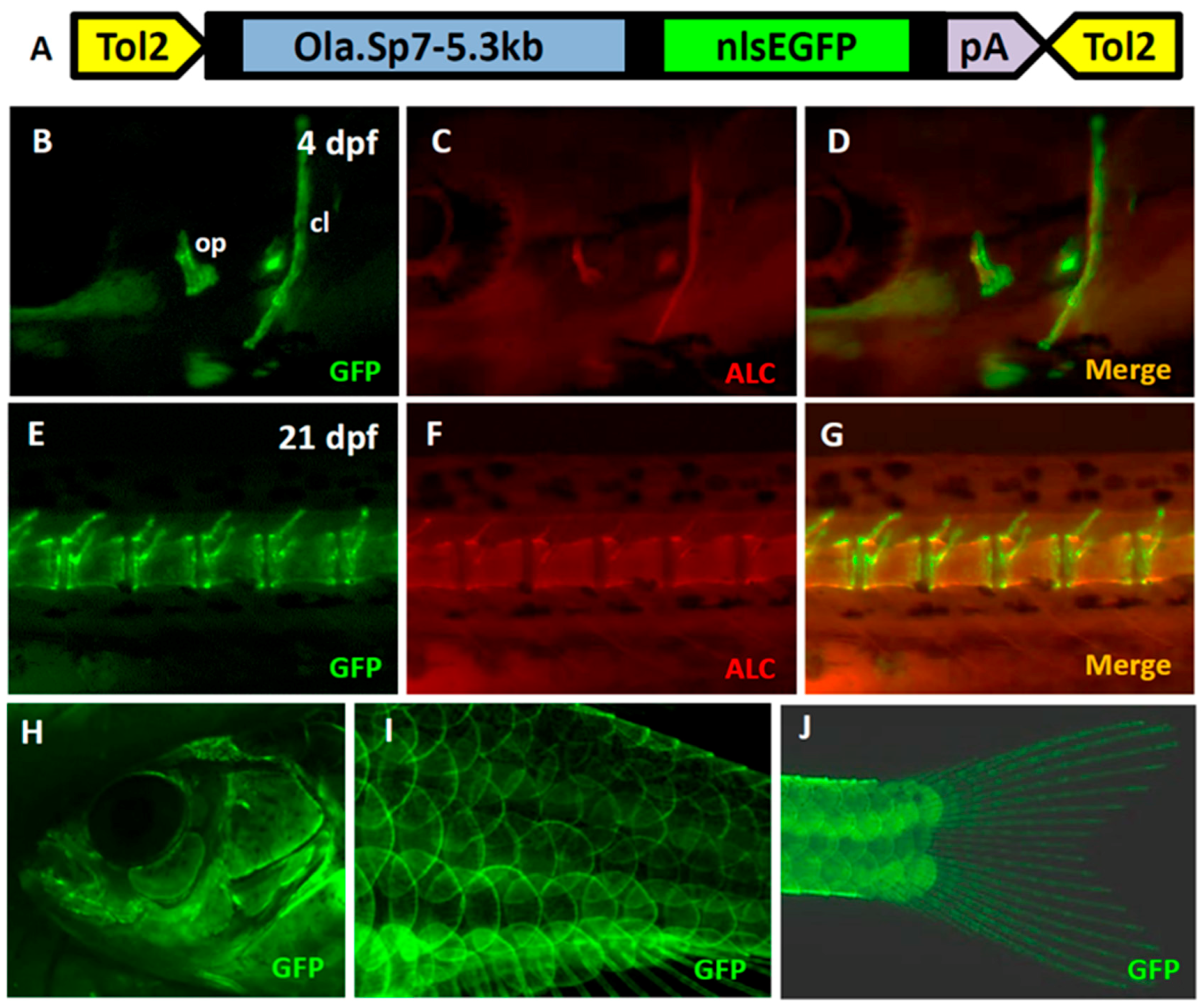
2.1. Generation of the Transgenic Fish Line Tg(Ola.Sp7:N1aICD)cy31
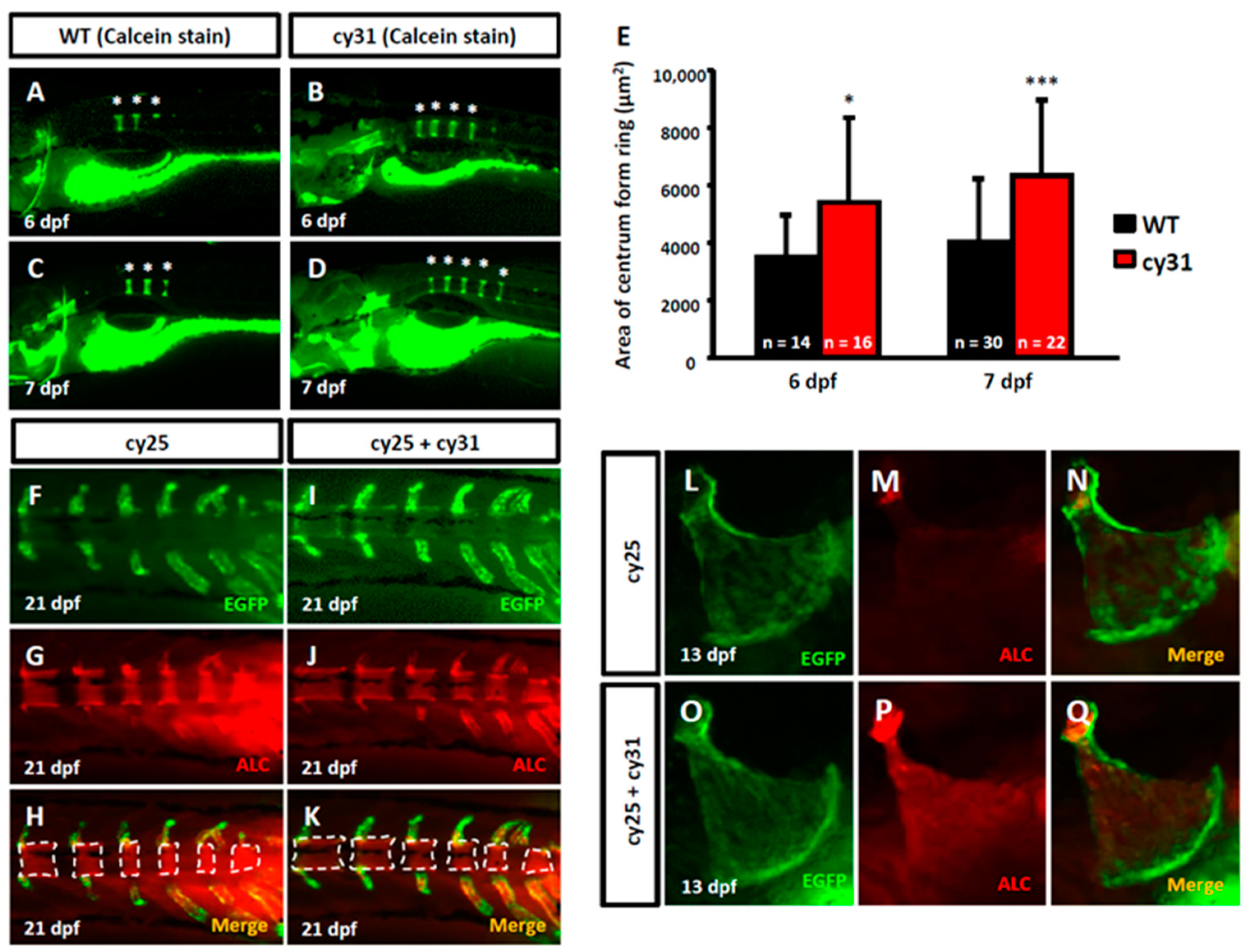
2.2. Calcification Level Increases in Early Embryonic Stage When N1aICD Overexpressed
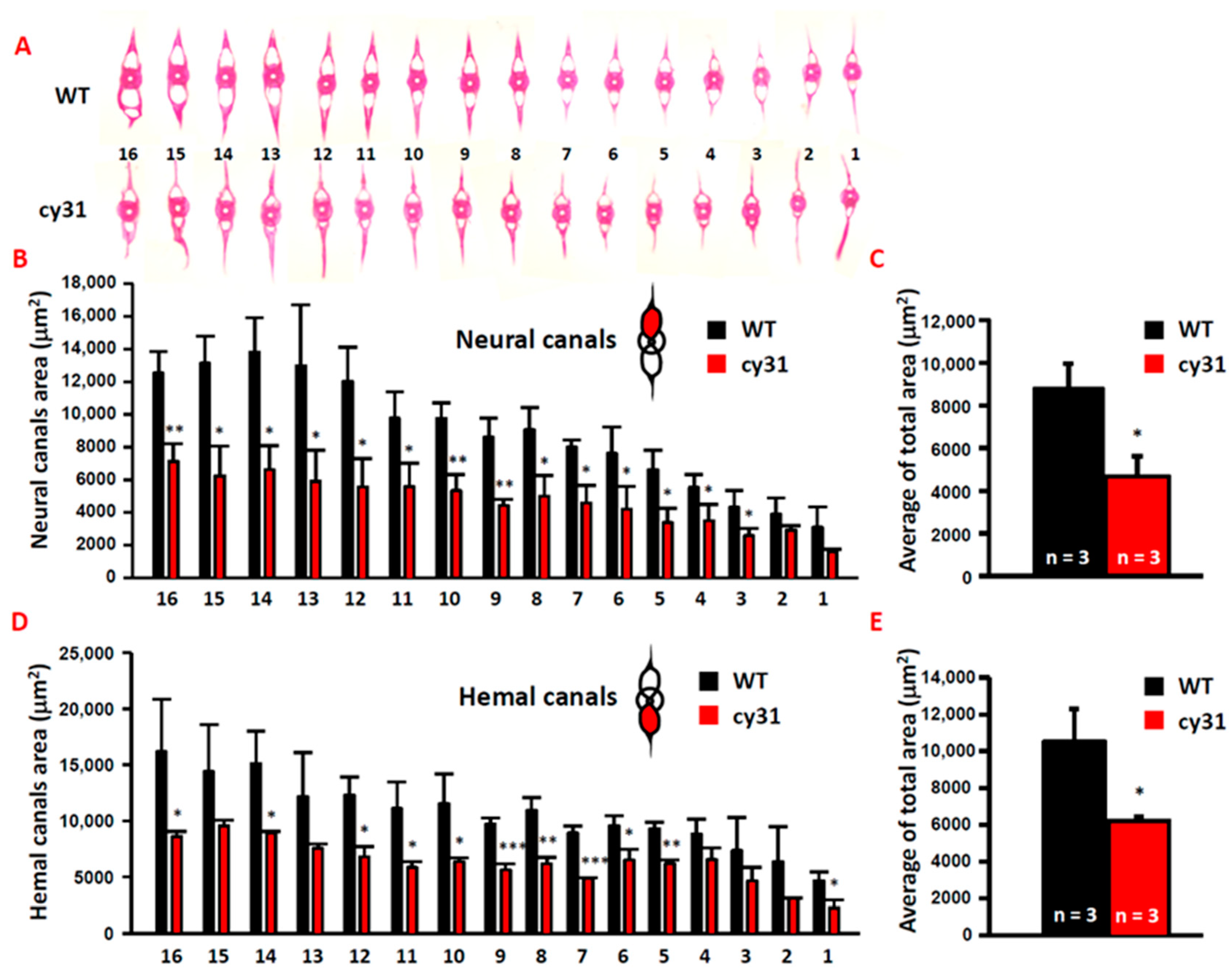
2.3. Hyperosteogeny Occurs in Late Adult Stage of N1aICD Overexpressed Zebrafish
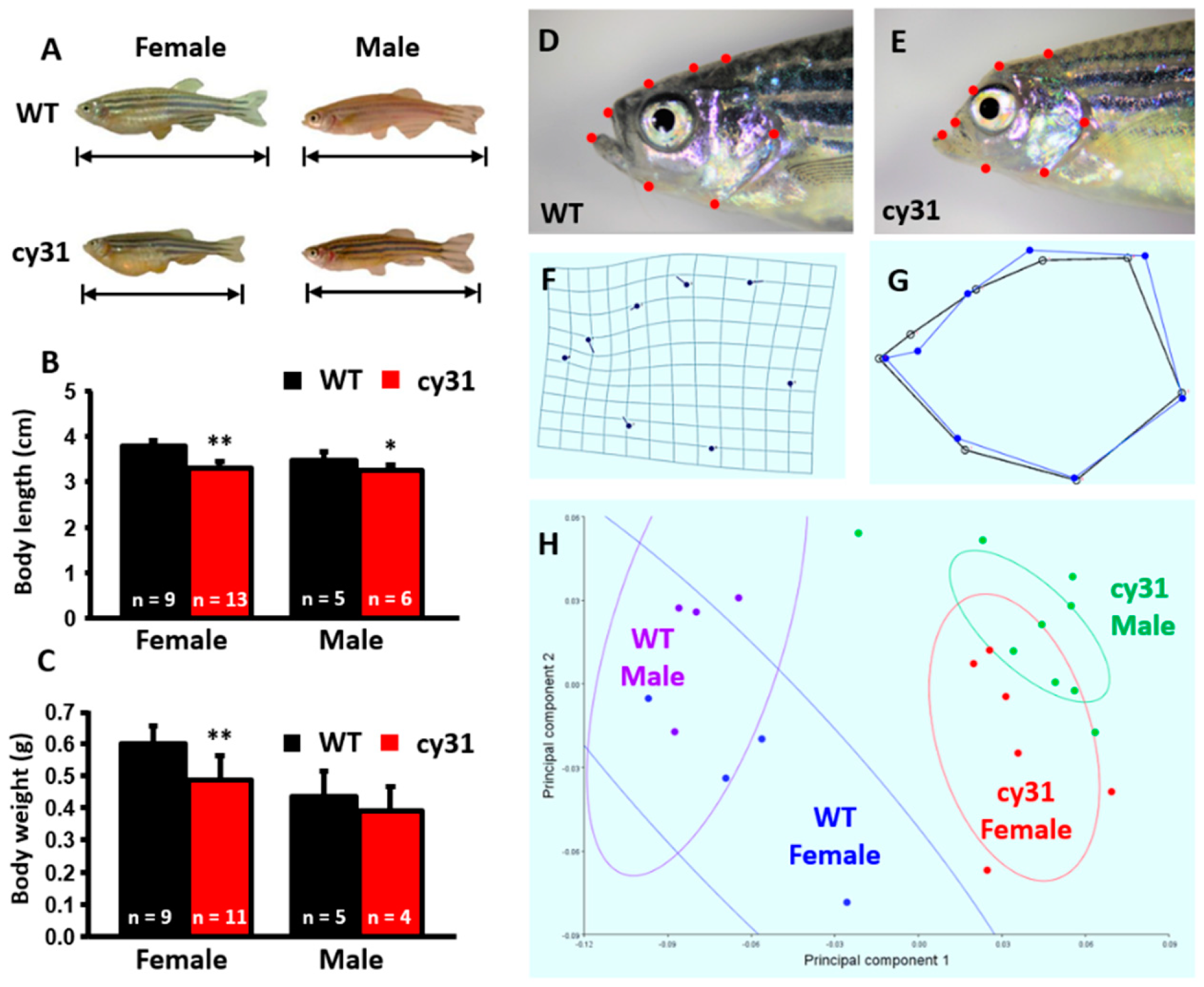
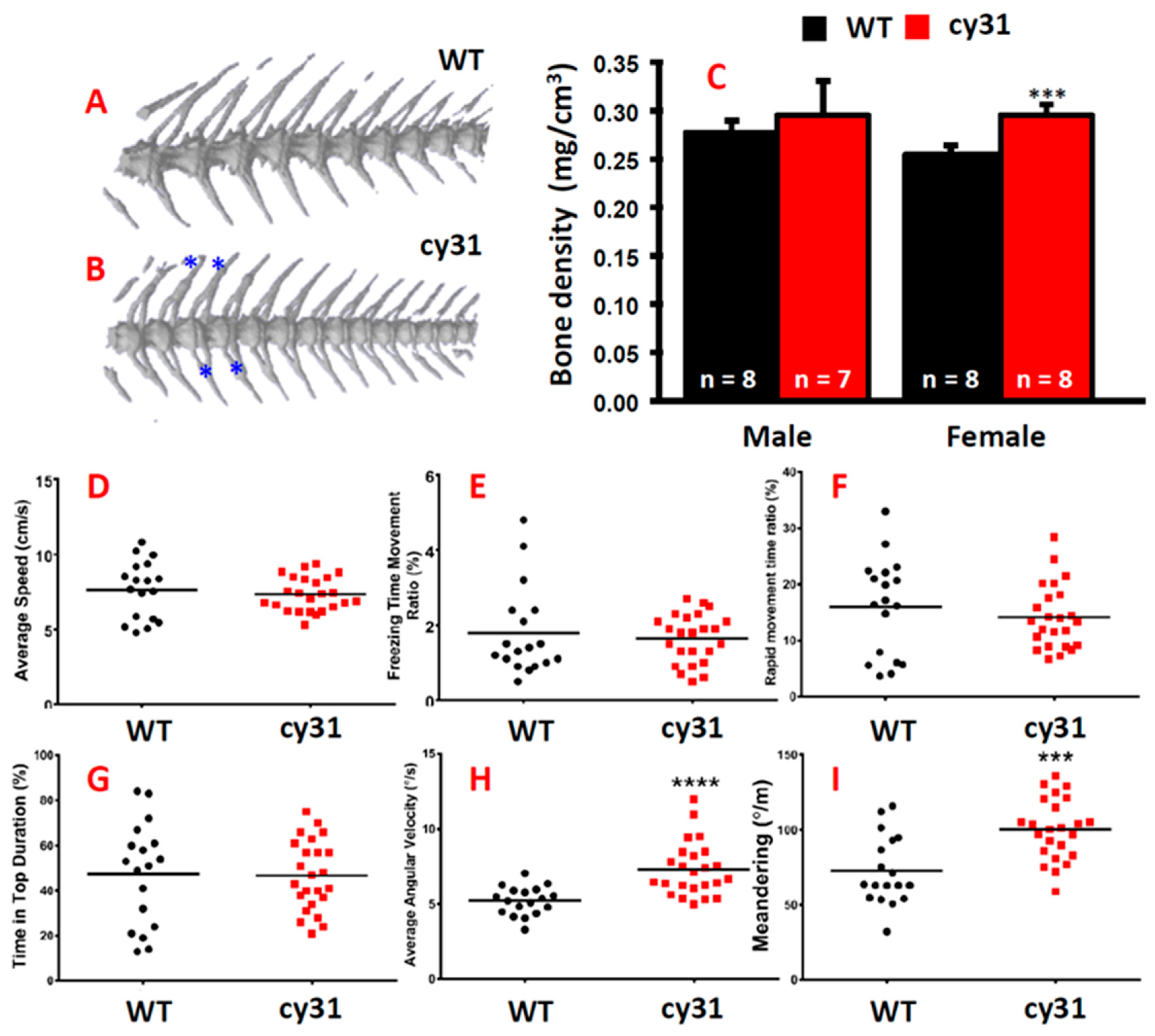
2.4. Notch Overexpression Causes High Bone Density and Swimming Behavior Alteration in Zebrafish
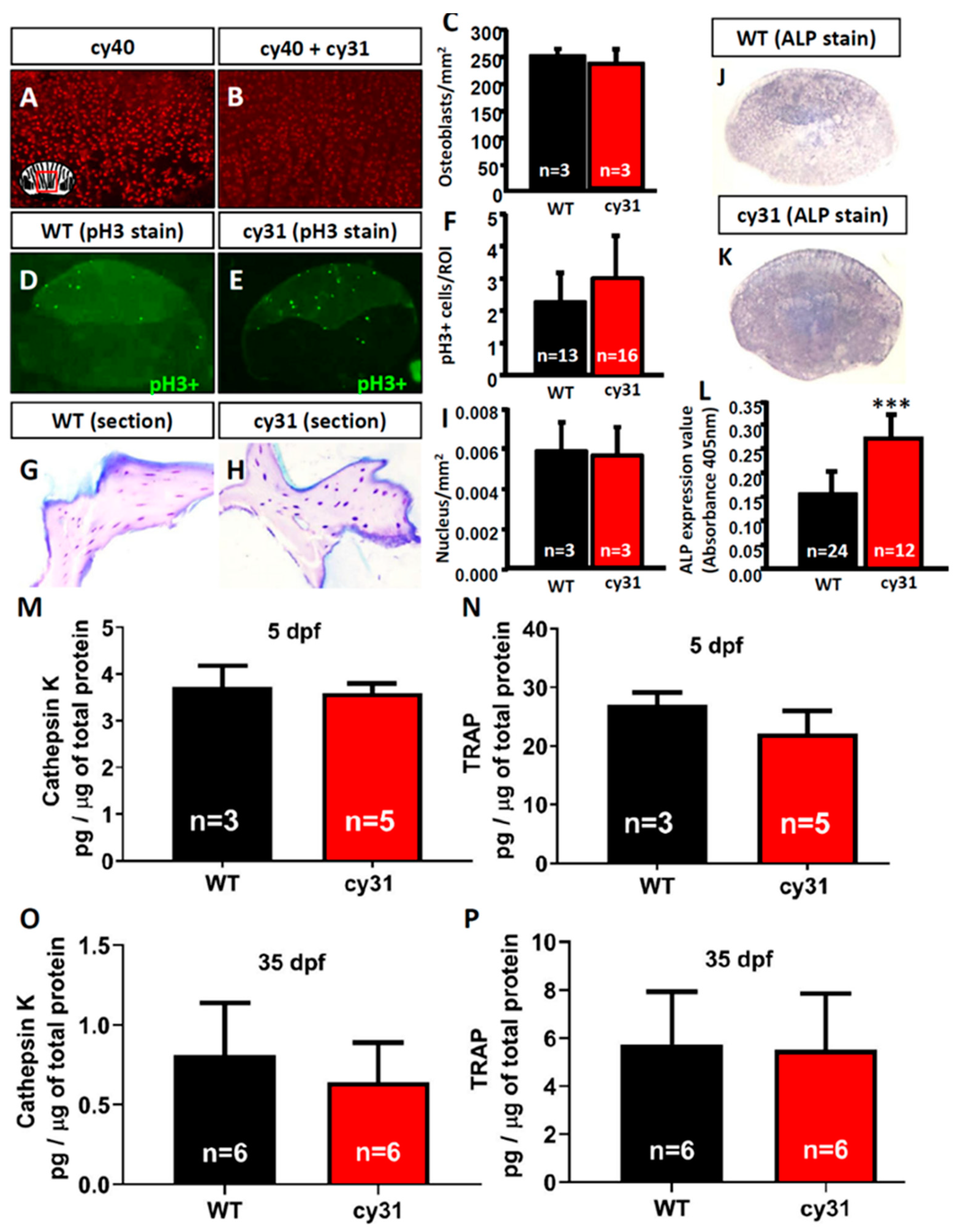
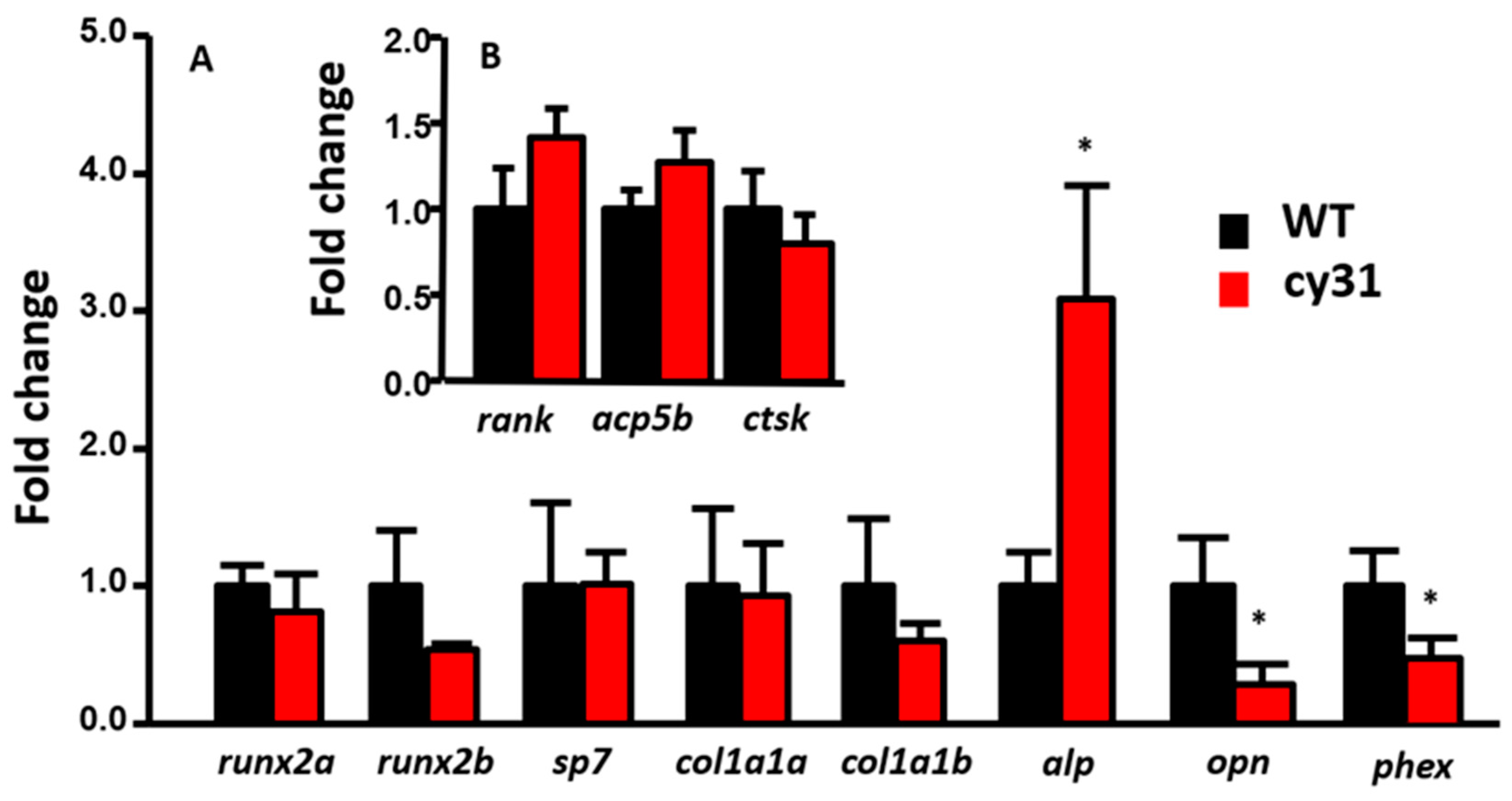
2.5. Increase of Osteoblast Activity Causes Hyperosteogeny in N1aICD-Overexpressed Zebrafish
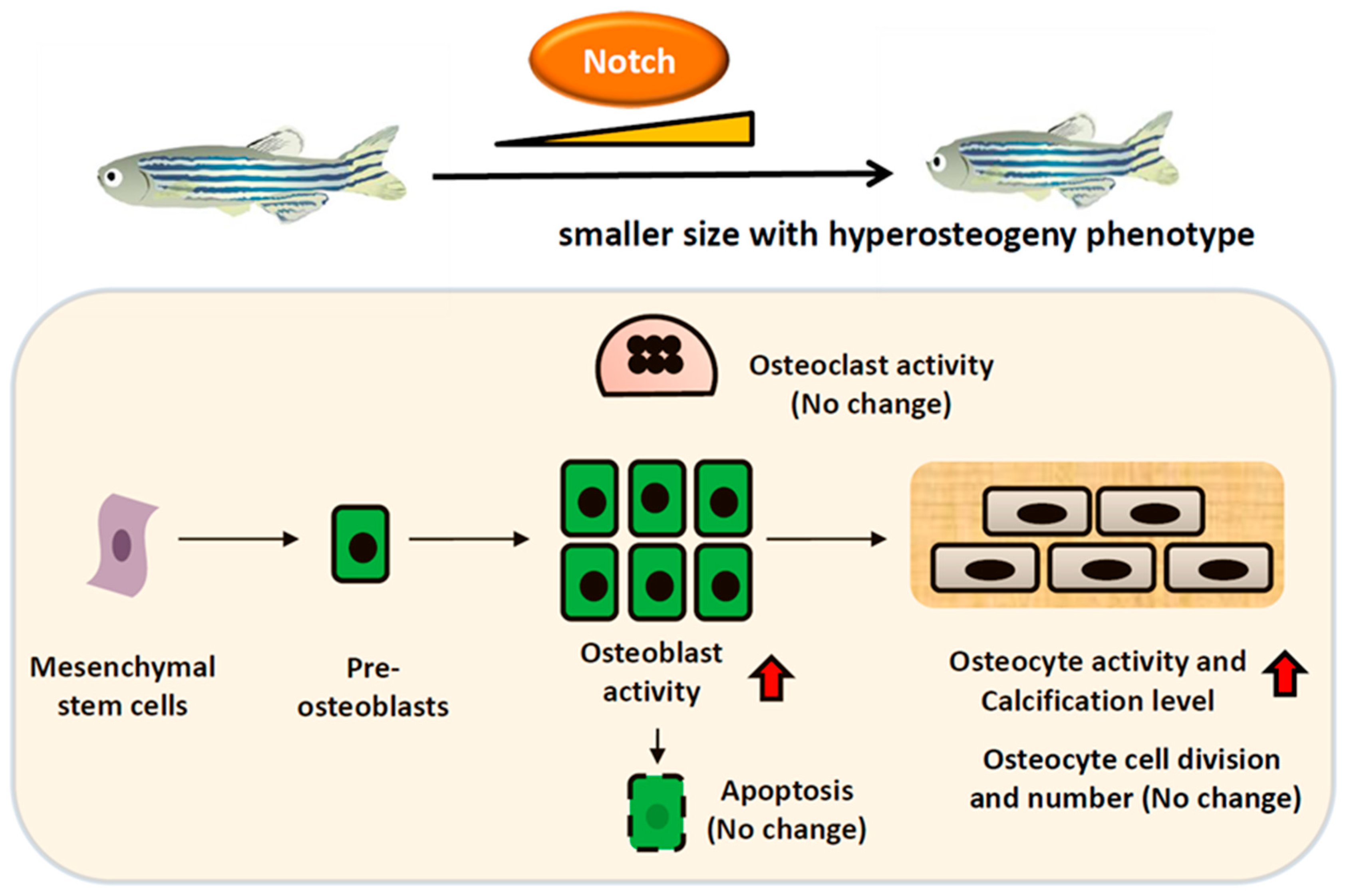
3. Discussion
4. Materials and Methods
4.1. Animal Ethics and Maintenance
4.2. Plasmid Construction
4.3. Genomic DNA Extraction
4.4. Calcein Labeling for Bone Formation Quantification
4.5. Bone and Cartilage Dual Staining
4.6. Whole-Mount Immunostaining
4.7. Histology
4.8. ALP Staining and ALP Activity Detection
4.9. Detection of Cathepsin K and TRAP Expression by ELISA
4.10. Micro CT
4.11. Morphometric Analysis
4.12. Locomotion Activity Assay
4.13. Quantitative Real-Time PCR
4.14. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A

References
- Lindsell, C.E.; Boulter, J.; DiSibio, G.; Gossler, A.; Weinmaster, G. Expression Patterns of Jagged, Delta1, Notch1, Notch2, and Notch3 Genes Identify Ligand–Receptor Pairs That May Function in Neural Development. Mol. Cell. Neurosci. 1996, 8, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Eiraku, M.; Hirata, Y.; Takeshima, H.; Hirano, T.; Kengaku, M. Delta/Notch-like Epidermal Growth Factor (EGF)-related Receptor, a Novel EGF-like Repeat-containing Protein Targeted to Dendrites of Developing and Adult Central Nervous System Neurons. J. Boil. Chem. 2002, 277, 25400–25407. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.-D.; Ang, B.-T.; Karsak, M.; Hu, W.-P.; Cui, X.-Y.; Duka, T.; Takeda, Y.; Chia, W.; Sankar, N.; Ng, Y.-K.; et al. F3/Contactin Acts as a Functional Ligand for Notch during Oligodendrocyte Maturation. Cell 2003, 115, 163–175. [Google Scholar] [CrossRef]
- Kopan, R.; Ilagan, M.X.G. The Canonical Notch Signaling Pathway: Unfolding the Activation Mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [PubMed]
- Groot, A.J.; Vooijs, M.A. The Role of Adams in Notch Signaling. Results Probl. Cell Differ. 2012, 727, 15–36. [Google Scholar]
- D’Souza, B.; Meloty-Kapella, L.; Weinmaster, G. Canonical and Non-Canonical Notch Ligands; Elsevier: Amsterdam, The Netherlands, 2010; Volume 92, pp. 73–129. [Google Scholar]
- Zhang, H.; Hilton, M.J.; Anolik, J.H.; Welle, S.L.; Zhao, C.; Yao, Z.; Li, X.; Wang, Z.; Boyce, B.F.; Xing, L. Notch inhibits osteoblast formation in inflammatory arthritis via noncanonical nf-kappab. J. Clin. Investig. 2014, 124, 3200–3214. [Google Scholar] [CrossRef] [PubMed]
- Engin, F.; Yao, Z.; Yang, T.; Zhou, G.; Bertin, T.; Jiang, M.M.; Chen, Y.; Wang, L.; Zheng, H.; Sutton, R.E.; et al. Dimorphic effects of Notch signaling in bone homeostasis. Nat. Med. 2008, 14, 299–305. [Google Scholar] [CrossRef]
- Zanotti, S.; Smerdel-Ramoya, A.; Stadmeyer, L.; Durant, D.; Radtke, F.; Canalis, E. Notch Inhibits Osteoblast Differentiation and Causes Osteopenia. Endocrinology 2008, 149, 3890–3899. [Google Scholar] [CrossRef] [PubMed]
- Ashley, J.W.; Ahn, J.; Hankenson, K.D. Notch signaling promotes osteoclast maturation and resorptive activity. J. Cell. Biochem. 2015, 116, 2598–2609. [Google Scholar] [CrossRef]
- Bai, S.; Kopan, R.; Zou, W.; Hilton, M.J.; Ong, C.T.; Long, F.; Ross, F.P.; Teitelbaum, S.L. Notch1 regulates osteoclastogenesis directly in osteoclast precursors and indirectly via osteoblast lineage cells. J. Biol. Chem. 2008, 283, 6509–6518. [Google Scholar] [CrossRef]
- Zanotti, S.; Canalis, E. Notch and the skeleton. Mol. Cell. Biol. 2010, 30, 886–896. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, H.; Nakao, A.; Okamoto, F.; Shin, M.; Kajiya, H.; Sakano, S.; Bigas, A.; Jimi, E.; Okabe, K. The association of notch2 and nf-kappab accelerates rankl-induced osteoclastogenesis. Mol. Cell. Biol. 2008, 28, 6402–6412. [Google Scholar] [CrossRef] [PubMed]
- DeLaurier, A.; Eames, B.F.; Blanco-Sánchez, B.; Peng, G.; He, X.; Swartz, M.E.; Ullmann, B.; Westerfield, M.; Kimmel, C.B.; Blanco-Sánchez, B. Zebrafish sp.7:EGFP: A transgenic for studying otic vesicle formation, skeletogenesis, and bone regeneration. Genesis 2010, 48, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A Simple Setup to Perform 3D Locomotion Tracking in Zebrafish by Using a Single Camera. Inventions 2018, 3, 11. [Google Scholar] [CrossRef]
- Pasqualetti, S.; Banfi, G.; Mariotti, M. Osteoblast and osteoclast behavior in Zebrafish cultured scales. Cell Tissue Res. 2012, 350, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Suzuki, N.; Takagi, C.; Ikegame, M.; Yamamoto, T.; Takahashi, A.; Moriyama, S.; Hattori, A.; Sakamoto, T. Prolactin Inhibits Osteoclastic Activity in the Goldfish Scale: A Novel Direct Action of Prolactin in Teleosts. Zoölog. Sci. 2008, 25, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Sato, M.; Nassar, H.F.; Abdel-Gawad, F.K.; Bassem, S.M.; Yachiguchi, K.; Tabuchi, Y.; Endo, M.; Sekiguchi, T.; Urata, M.; et al. Seawater Polluted with Highly Concentrated Polycyclic Aromatic Hydrocarbons Suppresses Osteoblastic Activity in the Scales of Goldfish, Carassius auratus. Zoölog. Sci. 2016, 33, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Bulman, M.P.; Kusumi, K.; Frayling, T.M.; McKeown, C.; Garrett, C.; Lander, E.S.; Krumlauf, R.; Hattersley, A.T.; Ellard, S.; Turnpenny, P.D. Mutations in the human Delta homologue, DLL3, cause axial skeletal defects in spondylocostal dysostosis. Nat. Genet. 2000, 24, 438–441. [Google Scholar] [CrossRef]
- Dunwoodie, S.L.; Clements, M.; Sparrow, D.B.; Sa, X.; Conlon, R.A.; Beddington, R.S.P. Axial skeletal defects caused by mutation in the spondylocostal dysplasia/pudgy gene Dll3 are associated with disruption of the segmentation clock within the presomitic mesoderm. Development 2002, 129, 1795–1806. [Google Scholar]
- Hilton, M.J.; Tu, X.; Wu, X.; Bai, S.; Zhao, H.; Kobayashi, T.; Kronenberg, H.M.; Teitelbaum, S.L.; Ross, F.P.; Kopan, R.; et al. Notch signaling maintains bone marrow mesenchymal progenitors by suppressing osteoblast differentiation. Nat. Med. 2008, 14, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Tezuka, K.-I.; Yasuda, M.; Watanabe, N.; Morimura, N.; Kuroda, K.; Miyatani, S.; Hozumi, N. Stimulation of Osteoblastic Cell Differentiation by Notch. J. Bone Miner. Res. 2002, 17, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Engin, F.; Lee, B. Notching the bone: Insights into multi-functionality. Bone 2010, 46, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z. Notch signaling stimulates osteogenic differentiation of human bone marrow-derived mesenchymal stem cells. Chin. Sci. Bull. 2004, 49, 815. [Google Scholar] [CrossRef]
- Hoo, J.Y.; Kumari, Y.; Shaikh, M.F.; Hue, S.M.; Goh, B.H. Zebrafish: A Versatile Animal Model for Fertility Research. BioMed Res. Int. 2016, 2016, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Dooley, K.; Zon, L.I. Zebrafish: A model system for the study of human disease. Curr. Opin. Genet. Dev. 2000, 10, 252–256. [Google Scholar] [CrossRef]
- Kari, G.; Rodeck, U.; Dicker, A.P.; Dicker, A. Zebrafish: An Emerging Model System for Human Disease and Drug Discovery. Clin. Pharmacol. Ther. 2007, 82, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Santoriello, C.; Zon, L.I. Hooked! Modeling human disease in Zebrafish. J. Clin. Investig. 2012, 122, 2337. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Barut, B.A.; Zon, L.I. Realizing the potential of Zebrafish as a model for human disease. Physiol. Genom. 2000, 2, 49–51. [Google Scholar] [CrossRef]
- Phillips, J.B.; Westerfield, M. Zebrafish models in translational research: Tipping the scales toward advancements in human health. Dis. Model. Mech. 2014, 7, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Friedman, M.S.; Luo, W.; Woolf, P.; Hankenson, K.D. The transcription factor osterix (sp7) regulates bmp6-induced human osteoblast differentiation. J. Cell. Physiol. 2012, 227, 2677–2685. [Google Scholar] [CrossRef] [PubMed]
- Zamurovic, N.; Cappellen, D.; Rohner, D.; Susa, M. Coordinated Activation of Notch, Wnt, and Transforming Growth Factor-β Signaling Pathways in Bone Morphogenic Protein 2-induced Osteogenesis. J. Boil. Chem. 2004, 279, 37704–37715. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Chen, S.; Yang, T.; Dawson, B.; Munivez, E.; Bertin, T.; Lee, B. Osteosclerosis owing to notch gain of function is solely rbpj-dependent. J. Bone Miner. Res. 2010, 25, 2175–2183. [Google Scholar] [CrossRef] [PubMed]
- Tu, X.; Chen, J.; Lim, J.; Karner, C.M.; Lee, S.Y.; Heisig, J.; Wiese, C.; Surendran, K.; Kopan, R.; Gessler, M.; et al. Physiological notch signaling maintains bone homeostasis via rbpjk and hey upstream of nfatc1. PLoS Genet. 2012, 8, e1002577. [Google Scholar] [CrossRef] [PubMed]
- Canalis, E.; Bridgewater, D.; Schilling, L.; Zanotti, S. Canonical notch activation in osteocytes causes osteopetrosis. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E171–E182. [Google Scholar] [CrossRef] [PubMed]
- Sciaudone, M.; Gazzerro, E.; Priest, L.; Delany, A.M.; Canalis, E. Notch 1 Impairs Osteoblastic Cell Differentiation. Endocrinology 2003, 144, 5631–5639. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Zhou, Y.; Lin, J.; Nguyen, T.D.; Huang, R.; Gu, Y.; Friis, T.; Crawford, R.; Xiao, Y. Notch expressed by osteocytes plays a critical role in mineralisation. J. Mol. Med. 2018, 96, 333–347. [Google Scholar] [CrossRef]
- Ji, Y.; Ke, Y.; Gao, S. Intermittent activation of notch signaling promotes bone formation. Am. J. Transl. Res. 2017, 9, 2933–2944. [Google Scholar]
- Stein, G.S.; Lian, J.B. Molecular Mechanisms Mediating Proliferation/Differentiation Interrelationships during Progressive Development of the Osteoblast Phenotype. Endocr. Rev. 1993, 14, 424–442. [Google Scholar] [CrossRef]
- Golub, E.; Harrison, G.; Taylor, A.; Camper, S.; Shapiro, I. The role of alkaline phosphatase in cartilage mineralization. Bone Miner. 1992, 17, 273–278. [Google Scholar] [CrossRef]
- Yao, W.; Lv, Y.; Gong, X.; Wu, J.; Bao, B. Different ossification patterns of intermuscular bones in fish with different swimming modes. Boil. Open 2015, 4, 1727–1732. [Google Scholar] [CrossRef] [PubMed]
- Crockett, J.C.; Rogers, M.J.; Coxon, F.P.; Hocking, L.J.; Helfrich, M.H. Bone remodelling at a glance. J. Cell Sci. 2011, 124, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; McDonald, J.M. Disorders of bone remodeling. Annu. Rev. Pathol. 2011, 6, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Gowen, M.; Pavasovic, D.; Lazner, F.; Kola, I. Osteopetrosis and osteoporosis: Two sides of the same coin. Hum. Mol. Genet. 1999, 8, 1839–1846. [Google Scholar]
- Wiener, R.C.; Sambamoorthi, U. Dental fluorosis and lumbar spine bone mineral density in adults, ages 20 to 49 years: Results from the 2003 to 2004 National Health and Nutrition Examination Survey. J. Dent. Hyg. JDH 2013, 87, 370–377. [Google Scholar] [PubMed]
- Jochum, W.; David, J.-P.; Elliott, C.; Wutz, A.; Plenk, H.; Matsuo, K.; Wagner, E.F. Increased bone formation and osteosclerosis in mice overexpressing the transcription factor Fra-1. Nat. Med. 2000, 6, 980–984. [Google Scholar] [CrossRef]
- Ihde, L.L.; Forrester, D.M.; Gottsegen, C.; Masih, S.; Patel, D.B.; Vachon, L.A.; White, E.A.; Matcuk, G.R. Sclerosing Bone Dysplasias: Review and Differentiation from Other Causes of Osteosclerosis. Radiographics 2011, 31, 1865–1882. [Google Scholar] [CrossRef]
- De Vernejoul, M.C. Sclerosing bone disorders. Best Pract. Res. Clin. Rheumatol. 2008, 22, 71–83. [Google Scholar] [CrossRef]
- Hoemann, C.; EI-Gabalawy, H.; McKee, M.; Hoemann, C. In vitro osteogenesis assays: Influence of the primary cell source on alkaline phosphatase activity and mineralization. Pathol. Boil. 2009, 57, 318–323. [Google Scholar] [CrossRef]
- Lin, C.H.; Su, C.H.; Tseng, D.Y.; Ding, F.C.; Hwang, P.P. Action of vitamin d and the receptor, vdra, in calcium handling in Zebrafish (Danio rerio). PLoS ONE 2012, 7, e45650. [Google Scholar] [CrossRef] [PubMed]
- Narisawa, S.; Yadav, M.C.; Millán, J.L. In Vivo Overexpression of Tissue-Nonspecific Alkaline Phosphatase Increases Skeletal Mineralization and Affects the Phosphorylation Status of Osteopontin. J. Bone Miner. Res. 2013, 28, 1587–1598. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Brachydanio Rerio); University of Oregon Press: Corvallis, OR, USA, 1995. [Google Scholar]
- Kwan, K.M.; Fujimoto, E.; Grabher, C.; Mangum, B.D.; Hardy, M.E.; Campbell, D.S.; Parant, J.M.; Yost, H.J.; Kanki, J.P.; Chien, C.-B.; et al. The Tol2kit: A multisite gateway-based construction kit for Tol2 transposon transgenesis constructs. Dev. Dyn. 2007, 236, 3088–3099. [Google Scholar] [CrossRef] [PubMed]
- Spoorendonk, K.M.; Peterson-Maduro, J.; Renn, J.; Trowe, T.; Kranenbarg, S.; Winkler, C.; Schulte-Merker, S. Retinoic acid and Cyp26b1 are critical regulators of osteogenesis in the axial skeleton. Development 2008, 135, 3765–3774. [Google Scholar] [CrossRef] [PubMed]
- Du, S.J.; Frenkel, V.; Kindschi, G.; Zohar, Y. Visualizing Normal and Defective Bone Development in Zebrafish Embryos Using the Fluorescent Chromophore Calcein. Dev. Boil. 2001, 238, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-R.; Lai, Y.-H.; Tsai, J.-J.; Hsiao, C.-D. Live Florescent Staining Platform Drug-Screening and Mechanism-Analysis in Zebrafish for Bone Mineralization. Life Sci. 2017, 22, 2068. [Google Scholar]
- Kelly, W.L.; Bryden, M.M. A Modified Differential Stain for Cartilage and Bone in Whole Mount Preparations of Mammalian Fetuses and Small Vertebrates. Stain. Technol. 1983, 58, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P. Morphoj: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′ to 3′) | Reverse (5′ to 3′) |
|---|---|---|
| Osteoblast Differentiation Makers | ||
| runx2a | GACGGTGGTGACGGTAATGG | TGCGGTGGGTTCGTGAATA |
| runx2b | CGGCTCCTACCAGTTCTCCA | CCATCTCCCTCCACTCCTCC |
| sp7 (osterix) | GGCTATGCTAACTGCGACCTG | GCTTTCATTGCGTCCGTTTT |
| col1a1a | GAATAAGCAGGTGGAGTCT | GAAGCAACTGAACGACATT |
| col1a1b | GGTGCTATGCTGTGATTCT | GATTCTCGCTAAGTGTCCAT |
| alp | CAAGAACTCAACAAGAAC | TGAGCATTGGTGTTATAC |
| osteopontin | GCCTCCATCATCATCGTA | AATCACCAAGCACCAGTA |
| phex | GAGAATGAATGGATGGATGA | TTGATGTCTTCGTTAATATAGGT |
| Osteoclast Differentiation Markers | ||
| ctsk | GGACTCAATCACTATCACT | AGAACAAGACATCTAAGACA |
| rank | GCACGGTTATTGTTGTTA | TATTCAGAGGTGGTGTTAT |
| acp5b | GCTGCTGCTAACAAACAAT | GACCAACCACGATGACAA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, S.-T.; Chen, J.-R.; Tsai, J.-J.; Lai, Y.-H.; Hsiao, C.-D. Overexpression of Notch Signaling Induces Hyperosteogeny in Zebrafish. Int. J. Mol. Sci. 2019, 20, 3613. https://doi.org/10.3390/ijms20153613
Liang S-T, Chen J-R, Tsai J-J, Lai Y-H, Hsiao C-D. Overexpression of Notch Signaling Induces Hyperosteogeny in Zebrafish. International Journal of Molecular Sciences. 2019; 20(15):3613. https://doi.org/10.3390/ijms20153613
Chicago/Turabian StyleLiang, Sung-Tzu, Jung-Ren Chen, Jhih-Jie Tsai, Yu-Heng Lai, and Chung-Der Hsiao. 2019. "Overexpression of Notch Signaling Induces Hyperosteogeny in Zebrafish" International Journal of Molecular Sciences 20, no. 15: 3613. https://doi.org/10.3390/ijms20153613
APA StyleLiang, S.-T., Chen, J.-R., Tsai, J.-J., Lai, Y.-H., & Hsiao, C.-D. (2019). Overexpression of Notch Signaling Induces Hyperosteogeny in Zebrafish. International Journal of Molecular Sciences, 20(15), 3613. https://doi.org/10.3390/ijms20153613