Intragenic Antimicrobial Peptide Hs02 Hampers the Proliferation of Single- and Dual-Species Biofilms of P. aeruginosa and S. aureus: A Promising Agent for Mitigation of Biofilm-Associated Infections

 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Antibacterial Activity of Peptide Hs02
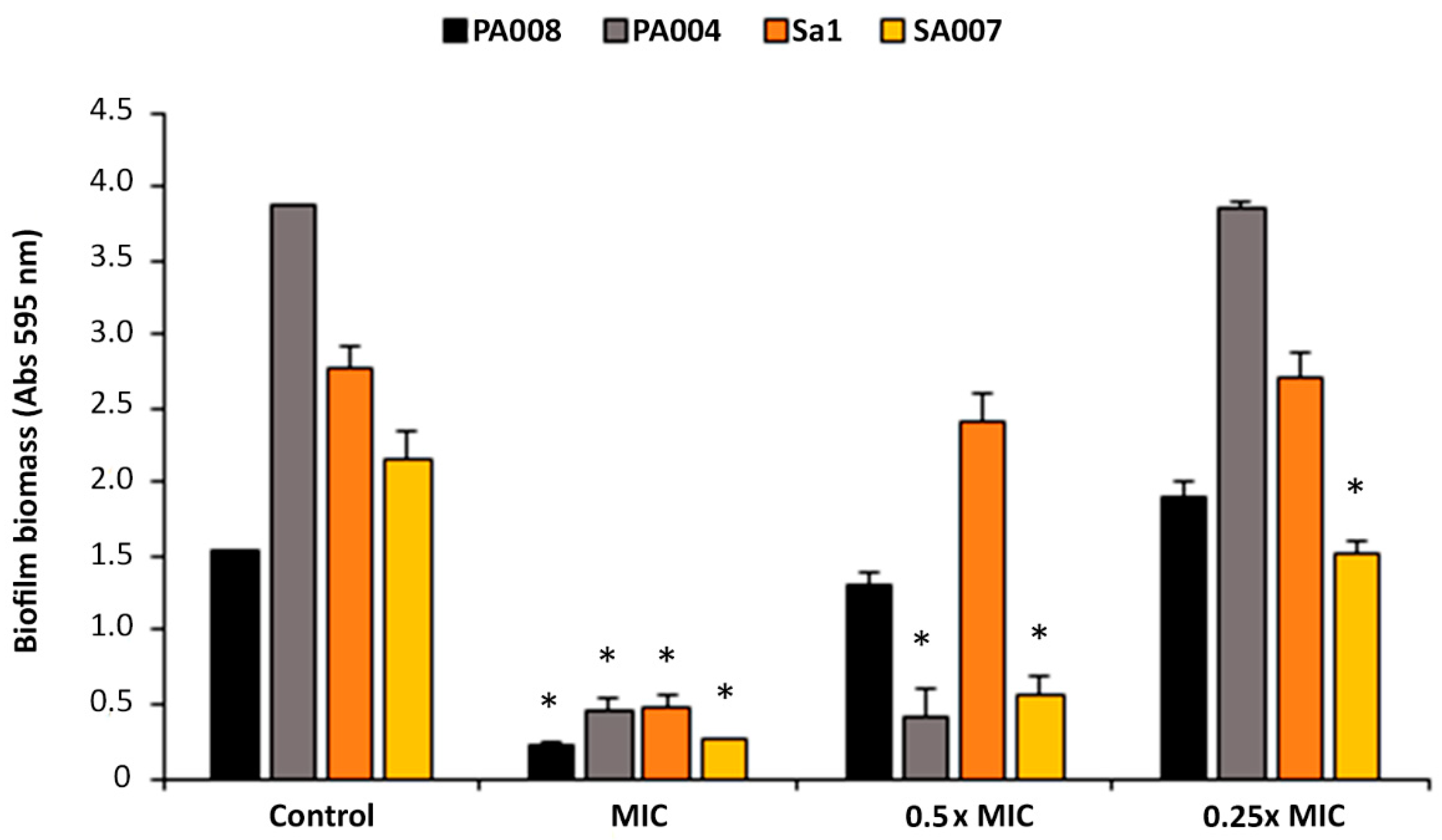
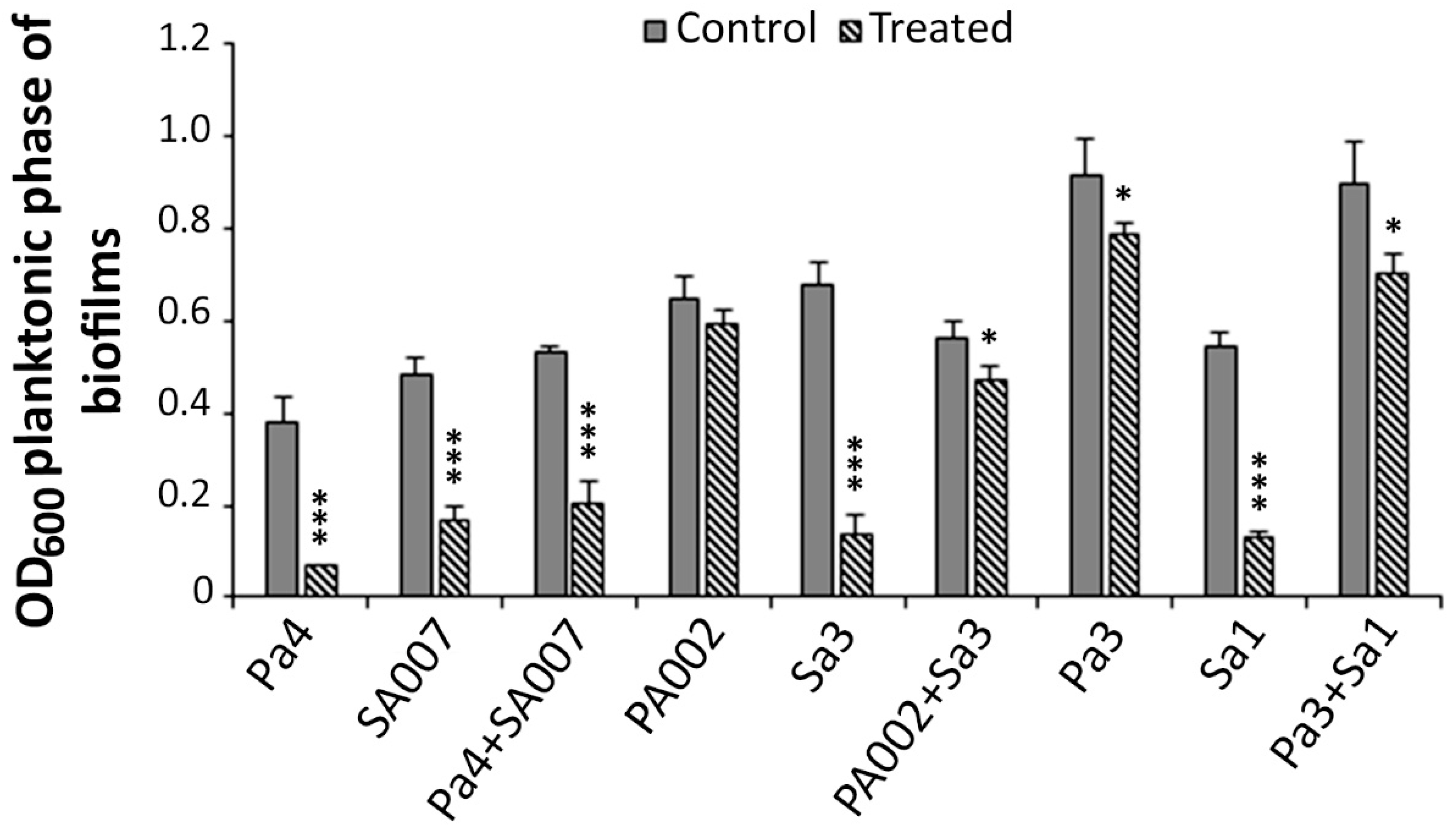
2.2. Antibiofilm Activity of Peptide Hs02
2.2.1. Peptide Hs02 Did Not Inhibit Biofilm Formation
2.2.2. Peptide Hs02 Hampered the Proliferation of 24 h Biofilms
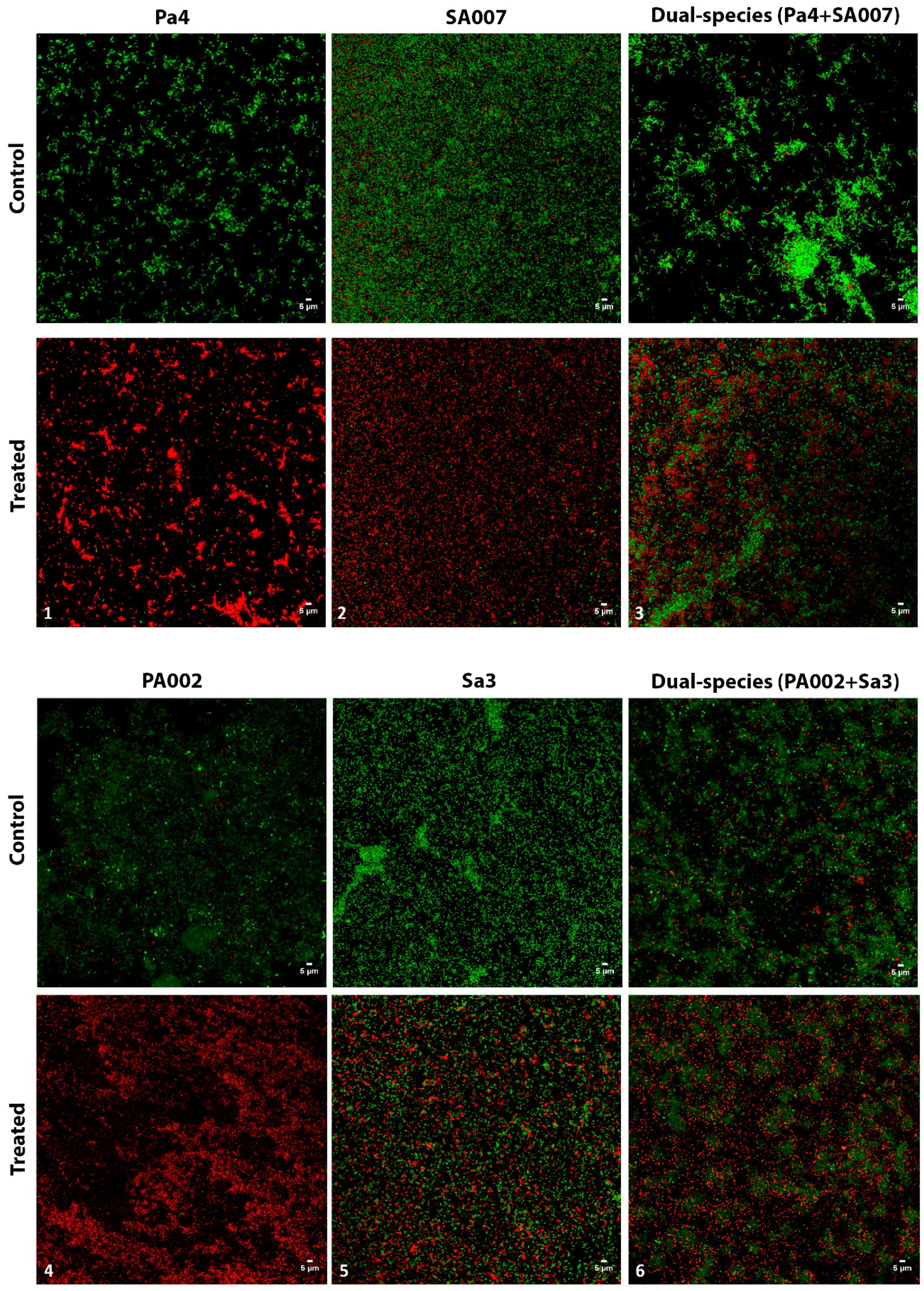
2.2.3. Peptide Hs02-Treated Biofilms Presented Reduced Viability
2.3. Peptide Hs02 Decreased Bacterial Membrane Fluidity
3. Materials and Methods
3.1. Hs02 Peptide
3.2. Bacterial Strains and Growth Conditions
3.3. MIC and MBC Determination
3.4. Biofilm Formation Inhibition Assay
3.5. Performed Biofilm Treatment Assay
3.6. Visualization of Biofilms by CLSM
3.7. Visualization of Biofilms by AFM
3.8. Membrane Fluidity Assessment by Laurdan Generalized Polarization (GP)
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AFM | Atomic force microscopy |
| AMP | Antimicrobial peptide |
| CFU | Colony-forming units |
| CLSM | Confocal laser scanning microscopy |
| GP | Generalized polarization |
| IAP | Intragenic antimicrobial peptide |
| MBC | Minimum bactericidal concentration |
| MDR | Multidrug-resistant |
| MIC | Minimum inhibitory concentration |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| PBS | Phosphate-buffered saline |
| TSB | Tryptic soy broth |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Antimicrobial resistance pattern |
|---|---|
| Pa3 | ATM, CIP, FEP, GEN |
| Pa4 | ATM, CAZ, CIP, FEP, IPM |
| PA002 | AMK, CIP, COL, GEN, TOB |
| PA004 | CIP, GEN, IPM, TOB, TZP |
| PA006 | AMK, CAZ, CIP, COL, FEP, GEN, IPM, TOB |
| Sa1 | AMC, AMP, CIP, FOX, TET |
| Sa3 | AMC, AMP, CIP, FOX, IPM |
| SA007 | CIP, CLI, ERI, GEN, LEV, MOX, OXA |
References
- Stacy, A.; McNally, L.; Darch, S.E.; Brown, S.P.; Whiteley, M. The biogeography of polymicrobial infection. Nat. Rev. Microbiol. 2016, 14, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Kong, E.F.; Tsui, C.; Kucharíková, S.; Andes, D.; Van Dijck, P.; Jabra-Rizk, M.A. Commensal Protection of Staphylococcus aureus against Antimicrobials by Candida albicans Biofilm Matrix. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, A.N.; De Jong, A.; Junker, S.; Becher, D.; Chlebowicz, M.A.; Duipmans, J.C.; Jonkman, M.F.; Van Dijl, J.M. From the wound to the bench: Exoproteome interplay between wound-colonizing Staphylococcus aureus strains and co-existing bacteria. Virulence 2018, 9, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Hotterbeekx, A.; Kumar-Singh, S.; Goossens, H.; Malhotra-Kumar, S. In vivo and In vitro Interactions between Pseudomonas aeruginosa and Staphylococcus spp. Front. Cell. Infect. Microbiol. 2017, 7, 106. [Google Scholar] [CrossRef] [PubMed]
- Bowler, P.G. Antibiotic resistance and biofilm tolerance: A combined threat in the treatment of chronic infections. J. Wound Care 2018, 27, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Burmølle, M.; Ren, D.; Bjarnsholt, T.; Sørensen, S.J. Interactions in multispecies biofilms: Do they actually matter? Trends Microbiol. 2014, 22, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Wolcott, R.; Costerton, J.; Raoult, D.; Cutler, S. The polymicrobial nature of biofilm infection. Clin. Microbiol. Infect. 2013, 19, 107–112. [Google Scholar] [CrossRef]
- Bessa, L.J.; Fazii, P.; Di Giulio, M.; Cellini, L. Bacterial isolates from infected wounds and their antibiotic susceptibility pattern: Some remarks about wound infection. Int. Wound J. 2015, 12, 47–52. [Google Scholar] [CrossRef]
- Radlinski, L.; Rowe, S.E.; Kartchner, L.B.; Maile, R.; Cairns, B.A.; Vitko, N.P.; Gode, C.J.; Lachiewicz, A.M.; Wolfgang, M.C.; Conlon, B.P. Pseudomonas aeruginosa exoproducts determine antibiotic efficacy against Staphylococcus aureus. PLoS Boil. 2017, 15, e2003981. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Sagel, S.D.; Gibson, R.L.; Emerson, J.; McNamara, S.; Burns, J.L.; Wagener, J.S.; Ramsey, B.W.; Inhaled Tobramycin in Young Children Study Group; Cystic Fibrosis Foundation Therapeutics Development Network. Impact of Pseudomonas and Staphylococcus infection on inflammation and clinical status in young children with cystic fibrosis. J. Pediatr. 2009, 154, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Hubert, D.; Reglier-Poupet, H.; Sermet-Gaudelus, I.; Ferroni, A.; Le Bourgeois, M.; Burgel, P.-R.; Serreau, R.; Dusser, D.; Poyart, C.; Coste, J. Association between Staphylococcus aureus alone or combined with Pseudomonas aeruginosa and the clinical condition of patients with cystic fibrosis. J. Cyst. Fibros. 2013, 12, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Dalton, T.; Dowd, S.E.; Wolcott, R.D.; Sun, Y.; Watters, C.; Griswold, J.A.; Rumbaugh, K.P. An in vivo polymicrobial biofilm wound infection model to study interspecies interactions. PLoS ONE 2011, 6, e27317. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.; Fothergill, J.L. The role of multispecies social interactions in shaping Pseudomonas aeruginosa pathogenicity in the cystic fibrosis lung. FEMS Microbiol. Lett. 2017, 364, 128. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Genet. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: Current and prospective therapeutic strategies. Nat. Rev. Genet. 2017, 15, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin, T.; Yau, Y.C.W.; Stapleton, P.J.; Gong, Y.; Wang, P.W.; Guttman, D.S.; Waters, V. Staphylococcus aureus interaction with Pseudomonas aeruginosa biofilm enhances tobramycin resistance. NPJ Biofilms Microbiomes 2017, 3, 25. [Google Scholar] [CrossRef]
- Kim, M.K.; Kang, H.K.; Ko, S.J.; Hong, M.J.; Bang, J.K.; Seo, C.H.; Park, Y. Mechanisms driving the antibacterial and antibiofilm properties of Hp1404 and its analogue peptides against multidrug-resistant Pseudomonas aeruginosa. Sci. Rep. 2018, 8, 1763. [Google Scholar] [CrossRef]
- Chung, P.Y.; Khanum, R. Antimicrobial peptides as potential anti-biofilm agents against multidrug-resistant bacteria. J. Microbiol. Immunol. Infect. 2017, 50, 405–410. [Google Scholar] [CrossRef]
- Bechinger, B.; Gorr, S.U. Antimicrobial peptides: Mechanisms of action and resistance. J. Dent. Res. 2017, 96, 254–260. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility In Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Pletzer, D.; Coleman, S.R.; Hancock, R.E. Anti-biofilm peptides as a new weapon in antimicrobial warfare. Curr. Opin. Microbiol. 2016, 33, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Di Luca, M.; Maccari, G.; Maisetta, G.; Batoni, G. BaAMPs: The database of biofilm-active antimicrobial peptides. Biofouling 2015, 31, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Brand, G.D.; Magalhães, M.T.Q.; Tinoco, M.L.P.; Aragão, F.J.L.; Nicoli, J.; Kelly, S.M.; Cooper, A.; Bloch, C. Probing Protein Sequences as Sources for Encrypted Antimicrobial Peptides. PLoS ONE 2012, 7, e45848. [Google Scholar] [CrossRef] [PubMed]
- Ramada, M.H.S.; Brand, G.D.; Abrão, F.Y.; Oliveira, M.; Filho, J.L.C.; Galbieri, R.; Gramacho, K.P.; Prates, M.V.; Bloch, C. Encrypted Antimicrobial Peptides from Plant Proteins. Sci. Rep. 2017, 7, 13263. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.; Pirri, G.; Rinaldi, A.C. Antimicrobial peptides: The LPS connection. Methods Mol. Biol. 2010, 618, 137–154. [Google Scholar]
- Brand, G.D.; Ramada, M.H.S.; Manickchand, J.R.; Correa, R.; Ribeiro, D.J.S.; Michele, A.; Santos, M.A.; Vasconcelos, A.G.; Abrão, F.Y.; Prates, M.V.; et al. Intragenic antimicrobial peptides (IAPs) from human proteins with potent antimicrobial and anti-inflammatory activity. PLoS ONE. in press.
- Monserrat-Martinez, A.; Gambin, Y.; Sierecki, E. Thinking Outside the Bug: Molecular Targets and Strategies to Overcome Antibiotic Resistance. Int. J. Mol. Sci. 2019, 20, 1255. [Google Scholar] [CrossRef]
- Travkova, O.G.; Moehwald, H.; Brezesinski, G. The interaction of antimicrobial peptides with membranes. Adv. Colloid Interface Sci. 2017, 247, 521–532. [Google Scholar] [CrossRef]
- Bessa, L.J.; Ferreira, M.; Gameiro, P. Evaluation of membrane fluidity of multidrug-resistant isolates of Escherichia coli and Staphylococcus aureus in presence and absence of antibiotics. J. Photochem. Photobiol. B Boil. 2018, 181, 150–156. [Google Scholar] [CrossRef]
- Bessa, L.J.; Ferreira, M.; Gameiro, P. Data on Laurdan spectroscopic analyses to compare membrane fluidity between susceptible and multidrug-resistant bacteria. Data Brief 2018, 21, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Scheinpflug, K.; Krylova, O.; Strahl, H. Measurement of cell membrane fluidity by Laurdan GP: Fluorescence spectroscopy and microscopy. Methods Mol. Biol. 2017, 1520, 159–174. [Google Scholar] [PubMed]
- Müller, A.; Wenzel, M.; Strahl, H.; Grein, F.; Saaki, T.N.V.; Kohl, B.; Siersma, T.; Bandow, J.E.; Sahl, H.G.; Schneider, T.; et al. Daptomycin inhibits cell envelope synthesis by interfering with fluid membrane microdomains. Proc. Natl. Acad. Sci. USA 2016, 113, E7077–E7086. [Google Scholar] [CrossRef] [PubMed]
- Baltz, R.H.; Miao, V.; Wrigley, S.K. Natural products to drugs: Daptomycin and related lipopeptide antibiotics. Nat. Prod. Rep. 2005, 22, 717–741. [Google Scholar] [CrossRef] [PubMed]
- Shireen, T.; Singh, M.; Das, T.; Mukhopadhyay, K. Differential Adaptive Responses of Staphylococcus aureus to In Vitro Selection with Different Antimicrobial Peptides. Antimicrob. Agents Chemother. 2013, 57, 5134–5137. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.; Lee, H.; Lee, S.; Kim, S.; Choi, K.-H. Membrane fluidity-related adaptive response mechanisms of foodborne bacterial pathogens under environmental stresses. Food Res. Int. 2015, 72, 25–36. [Google Scholar] [CrossRef]
- Wenzel, M.; Vischer, N.; Strahl, H.; Hamoen, L. Assessing Membrane Fluidity and Visualizing Fluid Membrane Domains in Bacteria Using Fluorescent Membrane Dyes. Bio-Protoc. 2018, 8, 1–26. [Google Scholar] [CrossRef]
- Chan, W.C.; White, P. Basic procedures. In Fmoc Solid Phase Peptide Synthesis: A Practical Approach; Chan, W.C., White, P., Eds.; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; CLSI Standard M07: Wayne, PA, USA, 2012. [Google Scholar]
- Bessa, L.J.; Eaton, P.; Dematei, A.; Plácido, A.; Vale, N.; Gomes, P.; Delerue-Matos, C.; Leite, J.R.S.; Gameiro, P. Synergistic and antibiofilm properties of ocellatin peptides against multidrug-resistant Pseudomonas aeruginosa. Futur. Microbiol. 2018, 13, 151–163. [Google Scholar] [CrossRef]




| Strains | MIC µg/mL (µM) | MBC µg/mL (µM) | |
|---|---|---|---|
| Reference strains | E. coli ATCC 25922 | 4 (2.0) | 4 (2.0) |
| P. aeruginosa ATCC 27853 | 8 (4.1) | 8 (4.1) | |
| S. aureus ATCC 25923 | 8 (4.1) | 8 (4.1) | |
| E. faecalis ATCC 29212 | 16 (8.2) | 16 (8.2) | |
| E. coli strains | E. coli TBX1/1(S) | 4 (2.0) | 4 (2.0) |
| E. coli TBX2/3(S) | 2 (1.0) | 2 (1.0) | |
| Ec1-SA1(R) | 4 (2.0) | 4 (2.0) | |
| EC001(R) | 4 (2.0) | 4 (2.0) | |
| P. aeruginosa strains | PAO1(S) | 8 (4.1) | 8 (4.1) |
| PA007(S) | 8 (4.1) | 8 (4.1) | |
| PA008(S) | 8 (4.1) | 8 (4.1) | |
| PA006(R) | 4 (2.0) | 4 (2.0) | |
| Pa4(R) | 4 (2.0) | 4 (2.0) | |
| PA002(R) | 16 (8.2) | 16 (8.2) | |
| PA004(R) | 8 (4.1) | 8 (4.1) | |
| Pa3(R) | 4 (2.0) | 4 (2.0) | |
| S. aureus strains | Sa1(R) | 8 (4.1) | 8 (4.1) |
| SA007(R) | 4 (2.0) | 4 (2.0) | |
| Sa3(R) | 8 (4.1) | 8 (4.1) | |
| E. faecalis strain | Ef1(R) | 4 (2.0) | 4 (2.0) |
| Control | 0.5× MIC | MIC | 2× MIC | |
|---|---|---|---|---|
| P. aeruginosa ATCC 27853 | 0.068 ± 0.002 | 0.077 ± 0.003 * | 0.123 ± 0.004 * | 0.155 ± 0.003 * |
| S. aureus ATCC 25923 | 0.007 ± 0.000 | 0.008 ± 0.000 * | 0.014 ± 0.002 * | 0.038 ± 0.002 * |
| PA002 | 0.015 ± 0.001 | 0.060 ± 0.003 * | 0.118 ± 0.003 * | 0.151 ± 0.002 * |
| PA004 | 0.065 ± 0.002 | 0.122 ± 0.002 * | 0.165 ± 0.001 * | 0.205 ± 0.002 * |
| Sa1 | 0.088 ± 0.003 | 0.057 ± 0.030 | 0.115 ± 0.002 * | 0.148 ± 0.002 * |
| Sa3 | 0.117 ± 0.001 | 0.110 ± 0.004 * | 0.093 ± 0.003 * | 0.123 ± 0.001 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bessa, L.J.; Manickchand, J.R.; Eaton, P.; Leite, J.R.S.A.; Brand, G.D.; Gameiro, P. Intragenic Antimicrobial Peptide Hs02 Hampers the Proliferation of Single- and Dual-Species Biofilms of P. aeruginosa and S. aureus: A Promising Agent for Mitigation of Biofilm-Associated Infections. Int. J. Mol. Sci. 2019, 20, 3604. https://doi.org/10.3390/ijms20143604
Bessa LJ, Manickchand JR, Eaton P, Leite JRSA, Brand GD, Gameiro P. Intragenic Antimicrobial Peptide Hs02 Hampers the Proliferation of Single- and Dual-Species Biofilms of P. aeruginosa and S. aureus: A Promising Agent for Mitigation of Biofilm-Associated Infections. International Journal of Molecular Sciences. 2019; 20(14):3604. https://doi.org/10.3390/ijms20143604
Chicago/Turabian StyleBessa, Lucinda J., Julia R. Manickchand, Peter Eaton, José Roberto S. A. Leite, Guilherme D. Brand, and Paula Gameiro. 2019. "Intragenic Antimicrobial Peptide Hs02 Hampers the Proliferation of Single- and Dual-Species Biofilms of P. aeruginosa and S. aureus: A Promising Agent for Mitigation of Biofilm-Associated Infections" International Journal of Molecular Sciences 20, no. 14: 3604. https://doi.org/10.3390/ijms20143604
APA StyleBessa, L. J., Manickchand, J. R., Eaton, P., Leite, J. R. S. A., Brand, G. D., & Gameiro, P. (2019). Intragenic Antimicrobial Peptide Hs02 Hampers the Proliferation of Single- and Dual-Species Biofilms of P. aeruginosa and S. aureus: A Promising Agent for Mitigation of Biofilm-Associated Infections. International Journal of Molecular Sciences, 20(14), 3604. https://doi.org/10.3390/ijms20143604