Premature Vascular Aging in Guinea Pigs Affected by Fetal Growth Restriction


Abstract

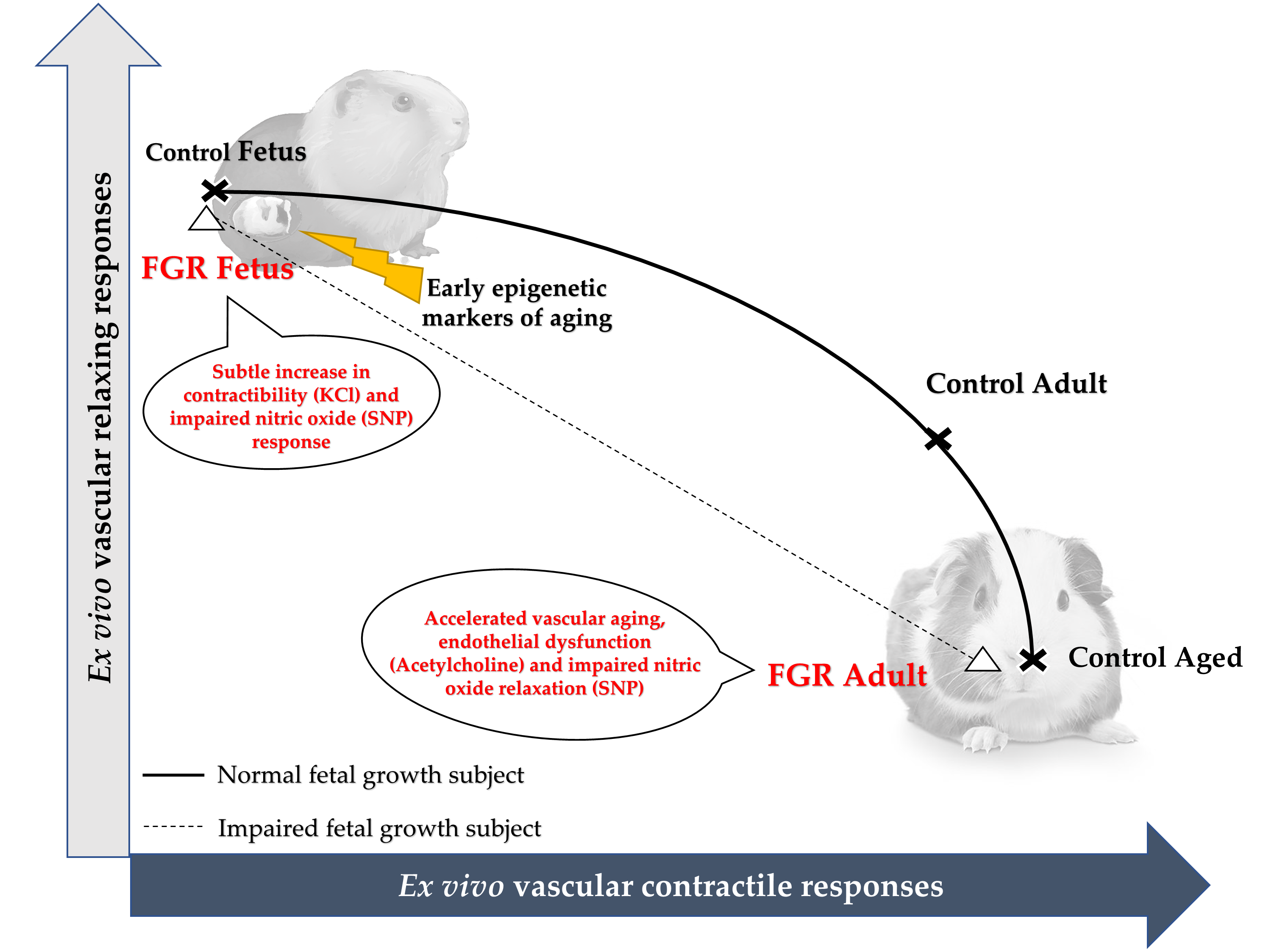
1. Introduction
2. Results
2.1. Life-Course Changes in the Ex Vivo Vascular Responses Control Guinea Pigs
2.2. Effect of FGR on Offspring Morphometry
2.3. Effect of Early Growth Restriction on the Vascular Responses of Fetal and Adult Guinea Pigs
2.4. LINE-1 DNA Methylation in HUAEC
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Human Umbilical Cord and Placental Samples
4.3. Animals & Experimental Design
4.4. Euthanasia
4.5. Carotid and Femoral Arteries Vascular Function
4.6. Human Umbilical Artery Endothelial Cell
4.7. LINE-1 DNA Methylation
4.8. Data and Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Ach | Acetylcholine |
| CpG | Cytosine-guanine dinucleotide |
| CVD | Cardiovascular Diseases |
| EVA | Early Vascular Aging |
| EDR | Endothelial Relaxation |
| FGR | Fetal Growth Restriction |
| HUAEC | Human Umbilical Artery Endothelial Cells |
| KCl | Potassium Chloride |
| LINE-1 | Long interspersed nuclear element-1 |
| NO | Nitric Oxide |
| SNP | Sodium Nitroprusside |
References
- Writing Group, M.; Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; Das, S.R.; de Ferranti, S.; Despres, J.P.; et al. Executive Summary: Heart Disease and Stroke Statistics—2016 Update: A Report From the American Heart Association. Circulation 2016, 133, 447–454. [Google Scholar]
- Barker, D.J.; Larsen, G.; Osmond, C.; Thornburg, K.L.; Kajantie, E.; Eriksson, J.G. The placental origins of sudden cardiac death. Int. J. Epidemiol. 2012, 41, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Figueras, F.; Gardosi, J. Intrauterine growth restriction: New concepts in antenatal surveillance, diagnosis, and management. Am. J. Obs. Gynecol. 2011, 204, 288–300. [Google Scholar] [CrossRef]
- Swanson, A.M.; David, A.L. Animal models of fetal growth restriction: Considerations for translational medicine. Placenta 2015, 36, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, S.; Kyle, P.M. Aetiology and pathogenesis of IUGR. Best Pract. Res. Clin. Obs. Gynaecol. 2009, 23, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Visentin, S.; Grumolato, F.; Nardelli, G.B.; Di Camillo, B.; Grisan, E.; Cosmi, E. Early origins of adult disease: Low birth weight and vascular remodeling. Atherosclerosis 2014, 237, 391–399. [Google Scholar] [CrossRef]
- Risnes, K.R.; Vatten, L.J.; Baker, J.L.; Jameson, K.; Sovio, U.; Kajantie, E.; Osler, M.; Morley, R.; Jokela, M.; Painter, R.C.; et al. Birthweight and mortality in adulthood: A systematic review and meta-analysis. Int. J. Epidemiol. 2011, 40, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Polglase, G.R.; Allison, B.J.; Coia, E.; Li, A.; Jenkin, G.; Malhotra, A.; Sehgal, A.; Kluckow, M.; Gill, A.W.; Hooper, S.B.; et al. Altered cardiovascular function at birth in growth-restricted preterm lambs. Pediatr. Res. 2016, 80, 538–546. [Google Scholar] [CrossRef]
- Sehgal, A.; Allison, B.J.; Gwini, S.M.; Menahem, S.; Miller, S.L.; Polglase, G.R. Vascular aging and cardiac maladaptation in growth-restricted preterm infants. J. Perinatol. 2018, 38, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Canas, D.; Herrera, E.A.; Garcia-Herrera, C.; Celentano, D.; Krause, B.J. Fetal Growth Restriction Induces Heterogeneous Effects on Vascular Biomechanical and Functional Properties in Guinea Pigs (Cavia porcellus). Front. Physiol. 2017, 8, 144. [Google Scholar] [CrossRef]
- Dodson, R.B.; Rozance, P.J.; Petrash, C.C.; Hunter, K.S.; Ferguson, V.L. Thoracic and abdominal aortas stiffen through unique extracellular matrix changes in intrauterine growth restricted fetal sheep. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H429–H437. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E.A.; Alegria, R.; Farias, M.; Diaz-Lopez, F.; Hernandez, C.; Uauy, R.; Regnault, T.R.; Casanello, P.; Krause, B.J. Assessment of in vivo fetal growth and placental vascular function in a novel intrauterine growth restriction model of progressive uterine artery occlusion in guinea pigs. J. Physiol. 2016, 594, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, P.M.; Lurbe, E.; Laurent, S. The early life origins of vascular ageing and cardiovascular risk: The EVA syndrome. J. Hypertens. 2008, 26, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Voors, A.W.; Webber, L.S.; Berenson, G.S. Time course studies of blood pressure in children—The Bogalusa Heart Study. Am. J. Epidemiol. 1979, 109, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y. Tracking of blood pressure from childhood to adulthood: A systematic review and meta-regression analysis. Circulation 2008, 117, 3171–3180. [Google Scholar] [CrossRef] [PubMed]
- Miles, K.L.; McDonnell, B.J.; Maki-Petaja, K.M.; Cockcroft, J.R.; Wilkinson, I.B.; McEniery, C.M. Enigma Study Investigators. The impact of birth weight on blood pressure and arterial stiffness in later life: The Enigma Study. J. Hypertens. 2011, 29, 2324–2331. [Google Scholar] [CrossRef] [PubMed]
- Crispi, F.; Figueras, F.; Cruz-Lemini, M.; Bartrons, J.; Bijnens, B.; Gratacos, E. Cardiovascular programming in children born small for gestational age and relationship with prenatal signs of severity. Am. J. Obs. Gynecol. 2012, 207, 121.e1–121.e9. [Google Scholar] [CrossRef] [PubMed]
- Bruno, R.M.; Duranti, E.; Ippolito, C.; Segnani, C.; Bernardini, N.; Di Candio, G.; Chiarugi, M.; Taddei, S.; Virdis, A. Different Impact of Essential Hypertension on Structural and Functional Age-Related Vascular Changes. Hypertension 2017, 69, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Vamos, Z.; Ivic, I.; Cseplo, P.; Toth, G.; Tamas, A.; Reglodi, D.; Koller, A. Pituitary adenylate cyclase-activating polypeptide (PACAP) induces relaxations of peripheral and cerebral arteries, which are differentially impaired by aging. J. Mol. Neurosci. 2014, 54, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Liskova, S.; Petrova, M.; Karen, P.; Behuliak, M.; Zicha, J. Contribution of Ca(2)(+)-dependent Cl(-) channels to norepinephrine-induced contraction of femoral artery is replaced by increasing EDCF contribution during ageing. Biomed. Res. Int. 2014, 2014, 289361. [Google Scholar] [CrossRef] [PubMed]
- Liskova, S.; Petrova, M.; Karen, P.; Kunes, J.; Zicha, J. Effects of aging and hypertension on the participation of endothelium-derived constricting factor (EDCF) in norepinephrine-induced contraction of rat femoral artery. Eur. J. Pharm. 2011, 667, 265–270. [Google Scholar]
- Hausman, N.; Martin, J.; Taggart, M.J.; Austin, C. Age-related changes in the contractile and passive arterial properties of murine mesenteric small arteries are altered by caveolin-1 knockout. J. Cell. Mol. Med. 2012, 16, 1720–1730. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.S.; Rueda-Clausen, C.F.; Davidge, S.T. Mechanisms of endothelium-dependent vasodilation in male and female, young and aged offspring born growth restricted. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R930–R938. [Google Scholar] [CrossRef] [PubMed]
- Lang, M.G.; Noll, G.; Luscher, T.F. Effect of aging and hypertension on contractility of resistance arteries: Modulation by endothelial factors. Am. J. Physiol. 1995, 269, H837–H844. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.C.; Loomis, E.D.; Collins, M.; Imig, J.D.; Inscho, E.W.; Pollock, J.S. Age-related alterations in NOS and oxidative stress in mesenteric arteries from male and female rats. J. Appl. Physiol. 2004, 97, 1268–1274. [Google Scholar] [CrossRef] [PubMed]
- Green, D. Point: Flow-mediated dilation does reflect nitric oxide-mediated endothelial function. J. Appl. Physiol. 2005, 99, 1233–1234. [Google Scholar] [CrossRef]
- Pyke, K.E.; Tschakovsky, M.E. The relationship between shear stress and flow-mediated dilatation: Implications for the assessment of endothelial function. J. Physiol. 2005, 568, 357–369. [Google Scholar] [CrossRef]
- Singh, N.; Prasad, S.; Singer, D.R.; MacAllister, R.J. Ageing is associated with impairment of nitric oxide and prostanoid dilator pathways in the human forearm. Clin. Sci. 2002, 102, 595–600. [Google Scholar] [CrossRef]
- Eskurza, I.; Monahan, K.D.; Robinson, J.A.; Seals, D.R. Effect of acute and chronic ascorbic acid on flow-mediated dilatation with sedentary and physically active human ageing. J. Physiol. 2004, 556, 315–324. [Google Scholar] [CrossRef]
- Gates, P.E.; Boucher, M.L.; Silver, A.E.; Monahan, K.D.; Seals, D.R. Impaired flow-mediated dilation with age is not explained by L-arginine bioavailability or endothelial asymmetric dimethylarginine protein expression. J. Appl. Physiol. 2007, 102, 63–71. [Google Scholar] [CrossRef]
- Black, M.A.; Cable, N.T.; Thijssen, D.H.; Green, D.J. Impact of age, sex, and exercise on brachial artery flow-mediated dilatation. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1109–H1116. [Google Scholar] [CrossRef] [PubMed]
- Albarwani, S.A.; Mansour, F.; Khan, A.A.; Al-Lawati, I.; Al-Kaabi, A.; Al-Busaidi, A.M.; Al-Hadhrami, S.; Al-Husseini, I.; Al-Siyabi, S.; Tanira, M.O. Aging Reduces L-Type Calcium Channel Current and the Vasodilatory Response of Small Mesenteric Arteries to Calcium Channel Blockers. Front. Physiol. 2016, 7, 171. [Google Scholar] [CrossRef] [PubMed]
- Stewart, K.G.; Zhang, Y.; Davidge, S.T. Aging increases PGHS-2-dependent vasoconstriction in rat mesenteric arteries. Hypertension 2000, 35, 1242–1247. [Google Scholar] [CrossRef] [PubMed]
- El Assar, M.; Fernandez, A.; Sanchez-Ferrer, A.; Angulo, J.; Rodriguez-Manas, L. Multivessel analysis of progressive vascular aging in the rat: Asynchronous vulnerability among vascular territories. Mech. Ageing Dev. 2018, 173, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.S.; Rueda-Clausen, C.F.; Davidge, S.T. Flow-mediated vasodilation is impaired in adult rat offspring exposed to prenatal hypoxia. J. Appl. Physiol. 2011, 110, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.; Pierce, G.L.; Stehouwer, C.D.; Padilla, J.; Thijssen, D.H. The impact of age on vascular smooth muscle function in humans. J. Hypertens. 2015, 33, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Yzydorczyk, C.; Armengaud, J.B.; Peyter, A.C.; Chehade, H.; Cachat, F.; Juvet, C.; Siddeek, B.; Simoncini, S.; Sabatier, F.; Dignat-George, F.; et al. Endothelial dysfunction in individuals born after fetal growth restriction: Cardiovascular and renal consequences and preventive approaches. J. Dev. Orig Health Dis. 2017, 8, 448–464. [Google Scholar] [CrossRef]
- Takeuchi, F.; Yamamoto, K.; Katsuya, T.; Sugiyama, T.; Nabika, T.; Ohnaka, K.; Yamaguchi, S.; Takayanagi, R.; Ogihara, T.; Kato, N. Reevaluation of the association of seven candidate genes with blood pressure and hypertension: A replication study and meta-analysis with a larger sample size. Hypertens. Res. 2012, 35, 825–831. [Google Scholar] [CrossRef]
- de Jong, F.; Monuteaux, M.C.; van Elburg, R.M.; Gillman, M.W.; Belfort, M.B. Systematic review and meta-analysis of preterm birth and later systolic blood pressure. Hypertension 2012, 59, 226–234. [Google Scholar] [CrossRef]
- Herrera, E.A.; Cifuentes-Zúñiga, F.; Figueroa, E.; Villanueva, C.; Hernández, C.; Alegría, R.; Arroyo-Jousse, V.; Peñaloza, E.; Farías, M.; Uauy, R.; et al. N-Acetylcysteine, a glutathione precursor, reverts vascular dysfunction and endothelial epigenetic programming in intrauterine growth restricted guinea pigs. J. Physiol. 2017, 595, 1077–1092. [Google Scholar] [CrossRef]
- Krause, B.J.; Peñaloza, E.; Candia, A.; Cañas, D.; Hernández, C.; Arenas, G.A.; Peralta-Scholz, M.J.; Valenzuela, R.; García-Herrera, C.; Herrera, E.A. Adult vascular dysfunction in fetal growth restricted guinea pigs is associated with a neonate-adult switching in Nos3 DNA methylation. Acta Physiol. 2019, e13328. [Google Scholar] [CrossRef]
- Morrison, J.L.; Botting, K.J.; Darby, J.R.T.; David, A.L.; Dyson, R.M.; Gatford, K.L.; Gray, C.; Herrera, E.A.; Hirst, J.J.; Kim, B.; et al. Guinea pig models for translation of the developmental origins of health and disease hypothesis into the clinic. J. Physiol. 2018, 596, 5535–5569. [Google Scholar] [CrossRef] [PubMed]
- Krause, B.J.; Costello, P.M.; Munoz-Urrutia, E.; Lillycrop, K.A.; Hanson, M.A.; Casanello, P. Role of DNA methyltransferase 1 on the altered eNOS expression in human umbilical endothelium from intrauterine growth restricted fetuses. Epigenetics 2013, 8, 944–952. [Google Scholar] [CrossRef]
- Postberg, J.; Kanders, M.; Forcob, S.; Willems, R.; Orth, V.; Hensel, K.O.; Weil, P.P.; Wirth, S.; Jenke, A.C. CpG signalling, H2A.Z/H3 acetylation and microRNA-mediated deferred self-attenuation orchestrate foetal NOS3 expression. Clin. Epigenetics 2015, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.F.; Ma, X.L.; Shen, Z.; Wu, X.L.; Cheng, F.; Du, L.Z. Epigenetic regulation of the endothelial nitric oxide synthase gene in persistent pulmonary hypertension of the newborn rat. J. Hypertens. 2010, 28, 2227–2235. [Google Scholar] [CrossRef] [PubMed]
- Castellano-Castillo, D.; Moreno-Indias, I.; Sanchez-Alcoholado, L.; Ramos-Molina, B.; Alcaide-Torres, J.; Morcillo, S.; Ocana-Wilhelmi, L.; Tinahones, F.; Queipo-Ortuno, M.I.; Cardona, F. Altered Adipose Tissue DNA Methylation Status in Metabolic Syndrome: Relationships Between Global DNA Methylation and Specific Methylation at Adipogenic, Lipid Metabolism and Inflammatory Candidate Genes and Metabolic Variables. J. Clin. Med. 2019, 8, 87. [Google Scholar] [CrossRef]
- Baccarelli, A.; Wright, R.; Bollati, V.; Litonjua, A.; Zanobetti, A.; Tarantini, L.; Sparrow, D.; Vokonas, P.; Schwartz, J. Ischemic heart disease and stroke in relation to blood DNA methylation. Epidemiology 2010, 21, 819–828. [Google Scholar] [CrossRef]
- Kim, M.; Long, T.I.; Arakawa, K.; Wang, R.; Yu, M.C.; Laird, P.W. DNA methylation as a biomarker for cardiovascular disease risk. PLoS ONE 2010, 5, e9692. [Google Scholar] [CrossRef]
- Cash, H.L.; McGarvey, S.T.; Houseman, E.A.; Marsit, C.J.; Hawley, N.L.; Lambert-Messerlian, G.M.; Viali, S.; Tuitele, J.; Kelsey, K.T. Cardiovascular disease risk factors and DNA methylation at the LINE-1 repeat region in peripheral blood from Samoan Islanders. Epigenetics 2011, 6, 1257–1264. [Google Scholar] [CrossRef]
- Erichsen, L.; Beermann, A.; Arauzo-Bravo, M.J.; Hassan, M.; Dkhil, M.A.; Al-Quraishy, S.; Hafiz, T.A.; Fischer, J.C.; Santourlidis, S. Genome-wide hypomethylation of LINE-1 and Alu retroelements in cell-free DNA of blood is an epigenetic biomarker of human aging. Saudi J. Biol. Sci. 2018, 25, 1220–1226. [Google Scholar] [CrossRef]
- Forristal, C.E.; Wright, K.L.; Hanley, N.A.; Oreffo, R.O.; Houghton, F.D. Hypoxia inducible factors regulate pluripotency and proliferation in human embryonic stem cells cultured at reduced oxygen tensions. Reproduction 2010, 139, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Bourque, D.K.; Avila, L.; Penaherrera, M.; von Dadelszen, P.; Robinson, W.P. Decreased placental methylation at the H19/IGF2 imprinting control region is associated with normotensive intrauterine growth restriction but not preeclampsia. Placenta 2010, 31, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Tzschoppe, A.; Doerr, H.; Rascher, W.; Goecke, T.; Beckmann, M.; Schild, R.; Struwe, E.; Geisel, J.; Jung, H.; Dotsch, J. DNA methylation of the p66Shc promoter is decreased in placental tissue from women delivering intrauterine growth restricted neonates. Prenat. Diagn. 2013, 33, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Michels, K.B.; Harris, H.R.; Barault, L. Birthweight, maternal weight trajectories and global DNA methylation of LINE-1 repetitive elements. PLoS ONE 2011, 6, e25254. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Chang, S.; Liu, C.; Zhang, M.; Zhang, L.; Liang, L.; Li, R.; Wang, X.; Qin, C.; Zhang, T.; et al. Low Maternal Dietary Folate Alters Retrotranspose by Methylation Regulation in Intrauterine Growth Retardation (IUGR) Fetuses in a Mouse Model. Med. Sci. Monit. 2019, 25, 3354–3365. [Google Scholar] [CrossRef] [PubMed]
- Krause, B.J.; Carrasco-Wong, I.; Caniuguir, A.; Carvajal, J.; Farias, M.; Casanello, P. Endothelial eNOS/arginase imbalance contributes to vascular dysfunction in IUGR umbilical and placental vessels. Placenta 2013, 34, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Delaey, C.; Van de Voorde, J. Pressure-induced myogenic responses in isolated bovine retinal arteries. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1871–1875. [Google Scholar]
- Mulvany, M.J.; Aalkjaer, C. Structure and function of small arteries. Physiol. Rev. 1990, 70, 921–961. [Google Scholar] [CrossRef]
- Outzen, E.M.; Zaki, M.; Abdolalizadeh, B.; Sams, A.; Boonen, H.C.; Sheykhzade, M. Translational value of mechanical and vasomotor properties of mouse isolated mesenteric resistance-sized arteries. Pharm. Res. Perspect. 2015, 3, e00200. [Google Scholar] [CrossRef]
- Benitez-Trinidad, A.B.; Medina-Diaz, I.M.; Bernal-Hernandez, Y.Y.; Barron-Vivanco, B.S.; Gonzalez-Arias, C.A.; Herrera-Moreno, J.F.; Alvarado-Cruz, I.; Quintanilla-Vega, B.; Rojas-Garcia, A.E. Relationship between LINE-1 methylation pattern and pesticide exposure in urban sprayers. Food Chem. Toxicol. 2018, 113, 125–133. [Google Scholar] [CrossRef]
- Tsuboi, Y.; Yamada, H.; Munetsuna, E.; Yamazaki, M.; Mizuno, G.; Murase, Y.; Ohashi, K.; Ishikawa, H.; Kondo, M.; Inoue, T.; et al. Relationship between Long Interspersed Nuclear Element-1 DNA Methylation in Leukocytes and Dyslipidemia in the Japanese General Population. J. Atheroscler. Thromb. 2018, 25, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.H.; Jin, E.H.; Kim, S.; Song, K.S.; Sung, J.K. LINE-1 hypomethylation is inversely correlated with UHRF1 overexpression in gastric cancer. Oncol. Lett. 2018, 15, 6666–6670. [Google Scholar] [CrossRef] [PubMed]
- Schneider, D.; Hernandez, C.; Farias, M.; Uauy, R.; Krause, B.J.; Casanello, P. Oxidative stress as common trait of endothelial dysfunction in chorionic arteries from fetuses with IUGR and LGA. Placenta 2015, 36, 552–558. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fetuses | Control (n = 12) | FGR (n = 12) | |
|---|---|---|---|
| Body (g) | 82.5 ± 3.0 | 53.8 ± 7.1** | |
| Heart (g) | 0.58 ± 0.02 | 0.46 ± 0.04* | |
| Kidney (g) | 0.76 ± 0.02 | 0.61 ± 0.05** | |
| Rel. heart weight | 0.71 ± 0.02 | 0.71 ± 0.03 | |
| Adults | Control (n = 6) | FGR (n = 6) | Aged (n = 6) |
| Body (g) | 660.3 ± 21.0 | 634.0 ± 36.1 | 0.65 ± 0.04* |
| Heart (g) | 2.11 ± 0.11 | 2.11 ± 0.09 | 2.83 ± 0.30* |
| Kidney (g) | 5.25 ± 0.20 | 4.27 ± 0.26* | 4.82 ± 0.25 |
| Rel. heart weight | 0.32 ± 0.02 | 0.34 ± 0.01 | 0.44 ± 0.05* |
| Relative kidney weight | 0.80 ± 0.03 | 0.65 ± 0.04* | 0.75 ± 0.05 |
| Carotid Artery | Control Fetus | FGR Fetus | Control Adult | FGR Adult | Control Aged |
| Diameter (μm) | 761 ± 16 | 675 ± 14*# | 1323 ± 33* | 1308 ± 43* | 1250 ± 69* |
| KClMax (Nm-2) | 1.41 ± 0.20 | 1.07 ± 0.10# | 6.10 ± 1.26* | 7.23 ± 1.06* | 5.64 ± 1.10* |
| AchMax (%KCl) | 72.9 ± 5.0 | 74.8 ± 7.8# | 41.4 ± 5.4* | 10.1 ± 11.0*# | 15.6 ± 3.9*# |
| SNPMax (%KCl) | 94.6 ± 3.8 | 100.0 ± 4.7# | 65.2 ± 5.3* | 40.0 ± 5.2*# | 22.7 ± 2.3*# |
| SNP pD2 | 6.99 ± 0.10 | 7.00 ± 0.10 | 6.98 ± 0.18 | 6.43 ± 0.20 | 6.25 ± 0.19*# |
| Femoral Artery | Control Fetus | FGR Fetus | Control Adult | FGR Adult | Control Aged |
| Diameter (μm) | 497 ± 10 | 396 ± 17*# | 648 ± 22* | 631 ± 21* | 697 ± 24* |
| KClMax (Nm-2) | 2.57 ± 0.29 | 5.43 ± 0.72*# | 8.86 ± 0.81* | 6.91 ± 0.92* | 6.97 ± 0.75* |
| AchMax (%KCl) | 57.9 ± 6.2 | 56.1 ± 5.7# | 87.6 ± 6.1* | 44.4 ± 7.5# | 51.7 ± 4.0# |
| SNPMax (%KCl) | 94.5 ± 3.2 | 95.4 ± 4.8 | 96.9 ± 4.4 | 89.7 ± 4.6 | 77.1 ± 3.9* |
| SNP pD2 | 7.78 ± 0.11 | 7.11 ± 0.10* | 7.06 0.12* | 6.44 ± 0.11*# | 6.40 ± 0.11*# |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paz, A.A.; Arenas, G.A.; Castillo-Galán, S.; Peñaloza, E.; Cáceres-Rojas, G.; Suazo, J.; Herrera, E.A.; Krause, B.J. Premature Vascular Aging in Guinea Pigs Affected by Fetal Growth Restriction. Int. J. Mol. Sci. 2019, 20, 3474. https://doi.org/10.3390/ijms20143474
Paz AA, Arenas GA, Castillo-Galán S, Peñaloza E, Cáceres-Rojas G, Suazo J, Herrera EA, Krause BJ. Premature Vascular Aging in Guinea Pigs Affected by Fetal Growth Restriction. International Journal of Molecular Sciences. 2019; 20(14):3474. https://doi.org/10.3390/ijms20143474
Chicago/Turabian StylePaz, Adolfo A., German A. Arenas, Sebastián Castillo-Galán, Estefanía Peñaloza, Gabriela Cáceres-Rojas, José Suazo, Emilio A. Herrera, and Bernardo J. Krause. 2019. "Premature Vascular Aging in Guinea Pigs Affected by Fetal Growth Restriction" International Journal of Molecular Sciences 20, no. 14: 3474. https://doi.org/10.3390/ijms20143474
APA StylePaz, A. A., Arenas, G. A., Castillo-Galán, S., Peñaloza, E., Cáceres-Rojas, G., Suazo, J., Herrera, E. A., & Krause, B. J. (2019). Premature Vascular Aging in Guinea Pigs Affected by Fetal Growth Restriction. International Journal of Molecular Sciences, 20(14), 3474. https://doi.org/10.3390/ijms20143474