Biocontrol and Action Mechanism of Bacillus amyloliquefaciens and Bacillus subtilis in Soybean Phytophthora Blight

Abstract
1. Introduction
2. Results and Discussion
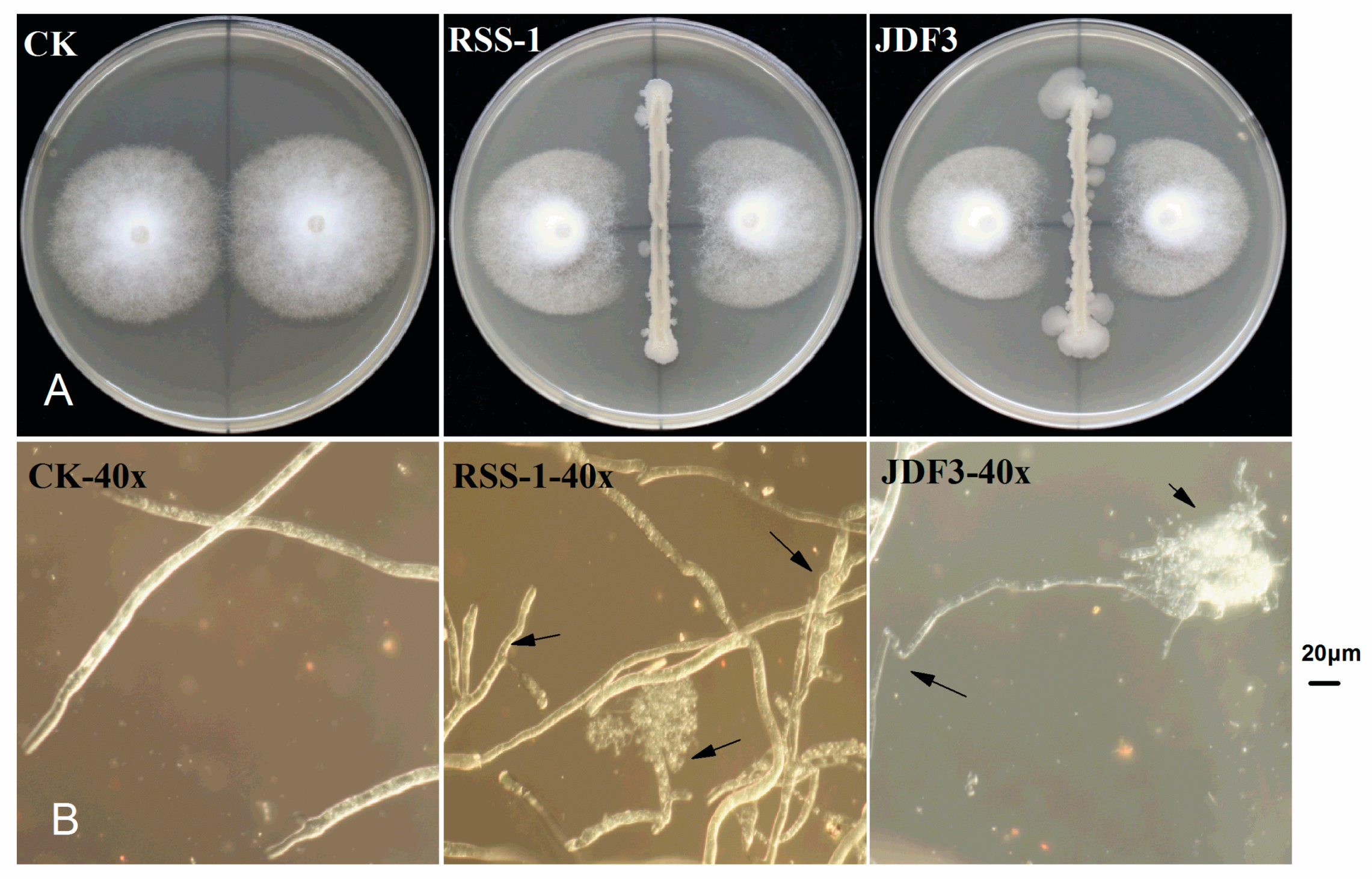
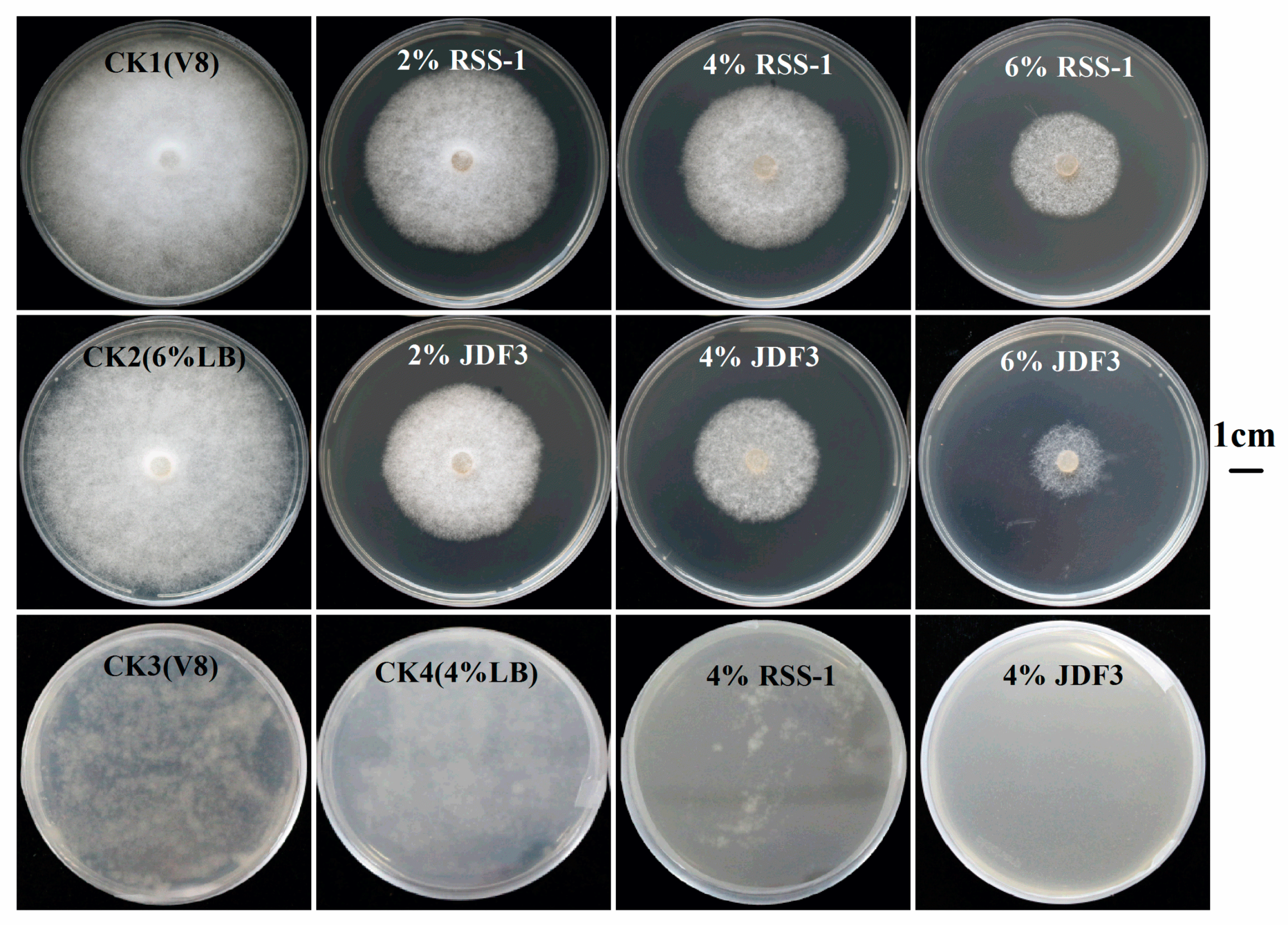
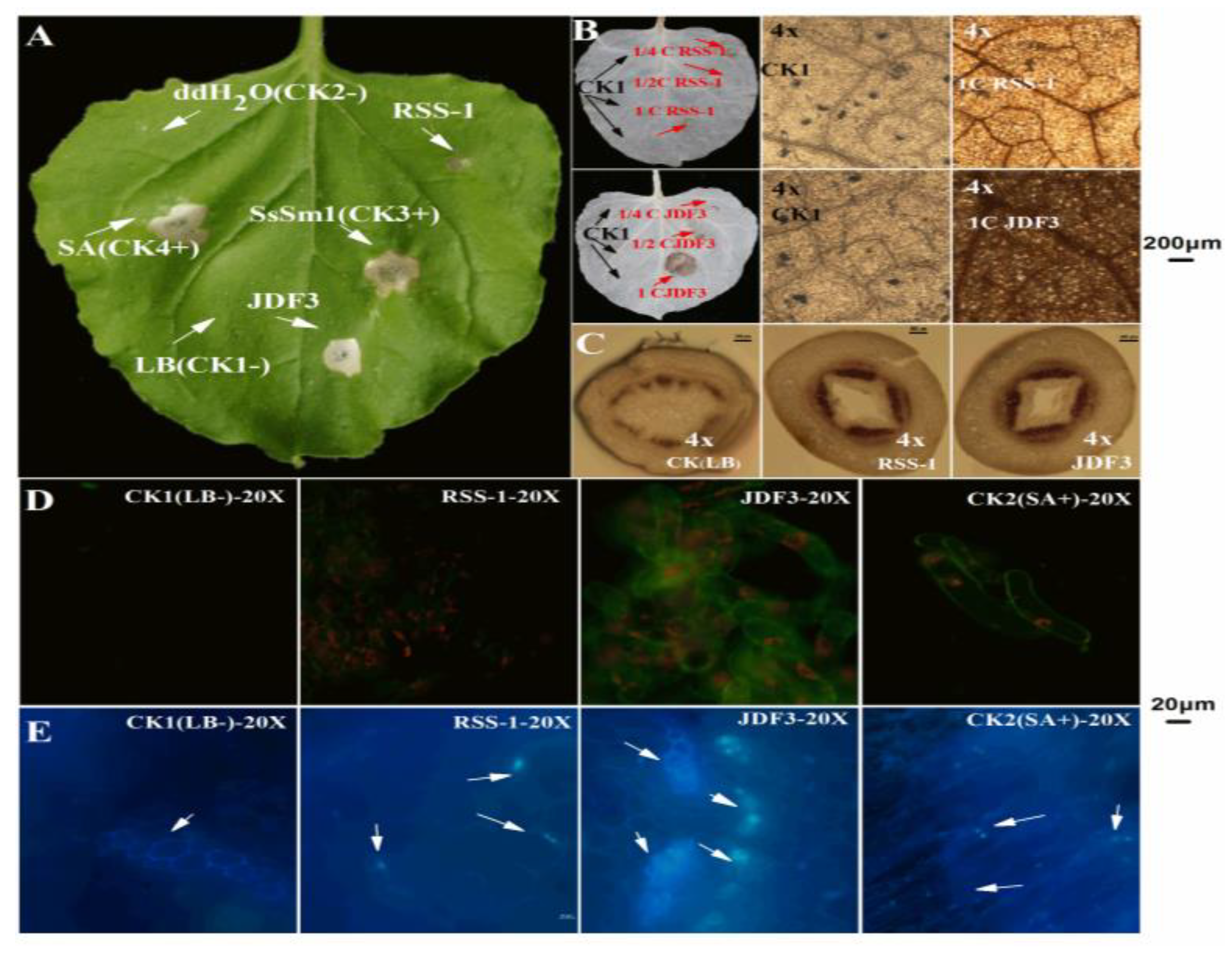
2.1. Inhibition of P. sojae by B. amyloliquefaciens and B. subtilis
2.2. Transcriptome Sequencing Quality Analysis
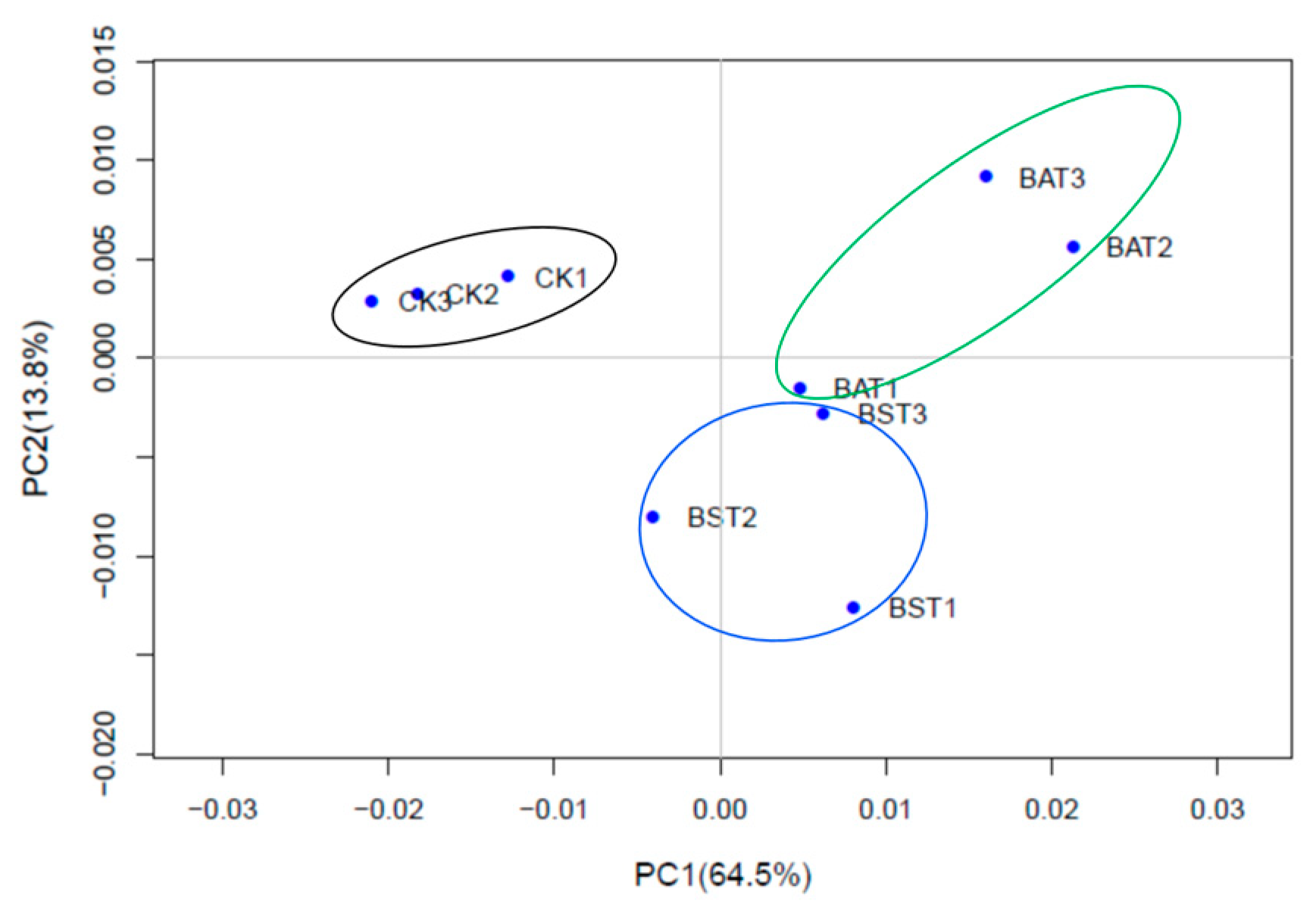
2.3. Analysis of Principal Components
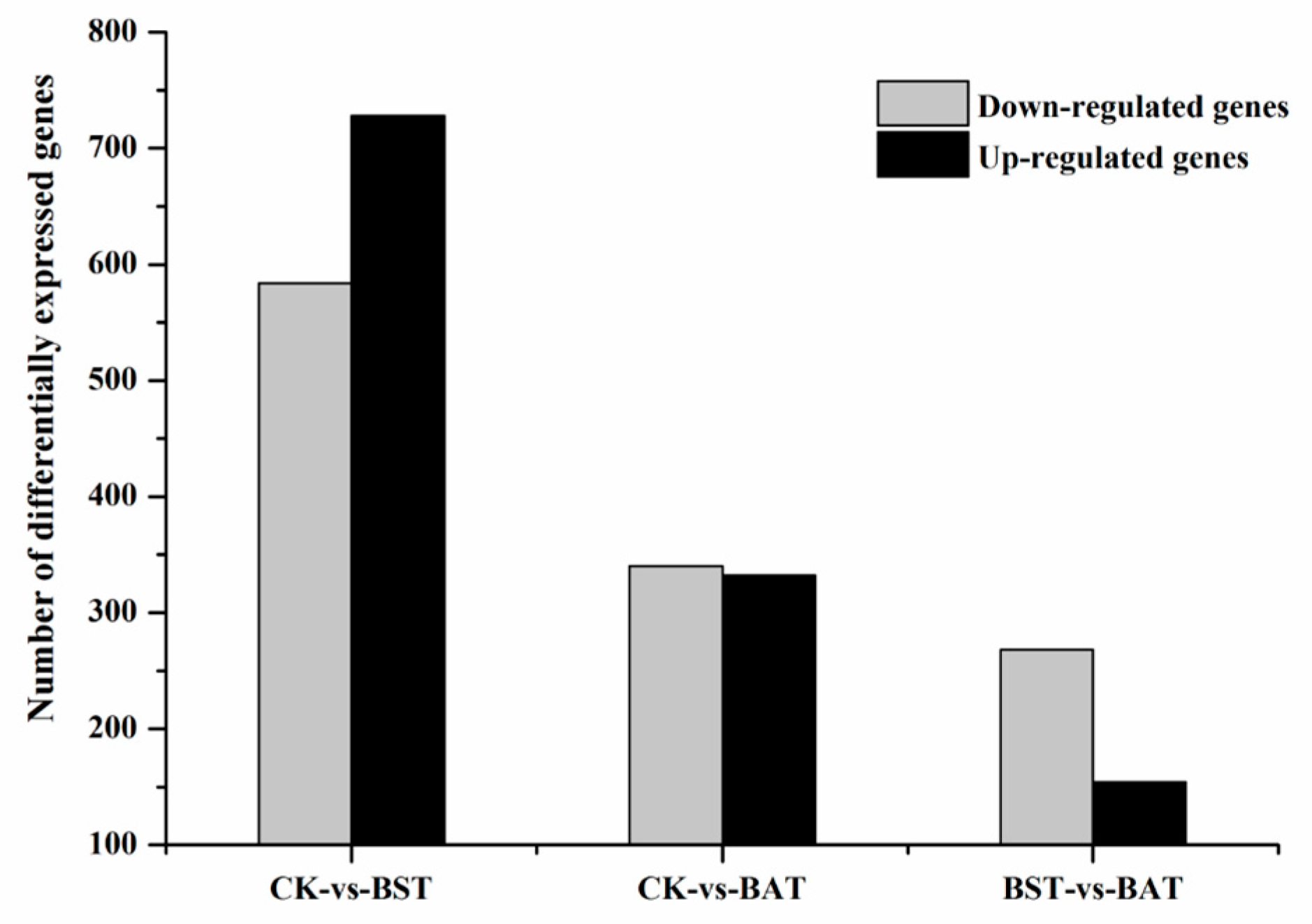
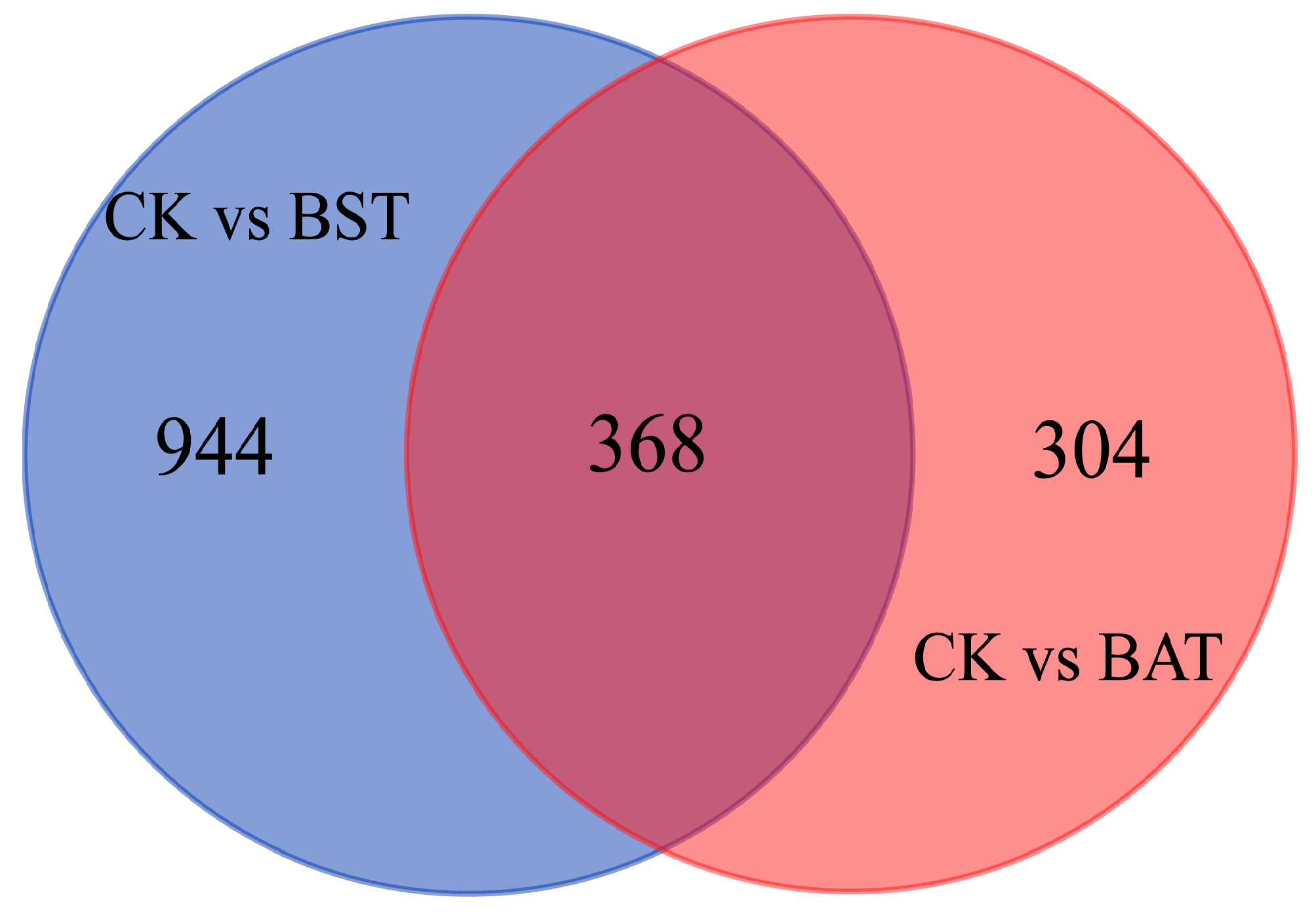
2.4. Effect of B. amyloliquefaciens and B. subtilis on the Gene Expression of P. sojae and KEGG Enrichment Analysis
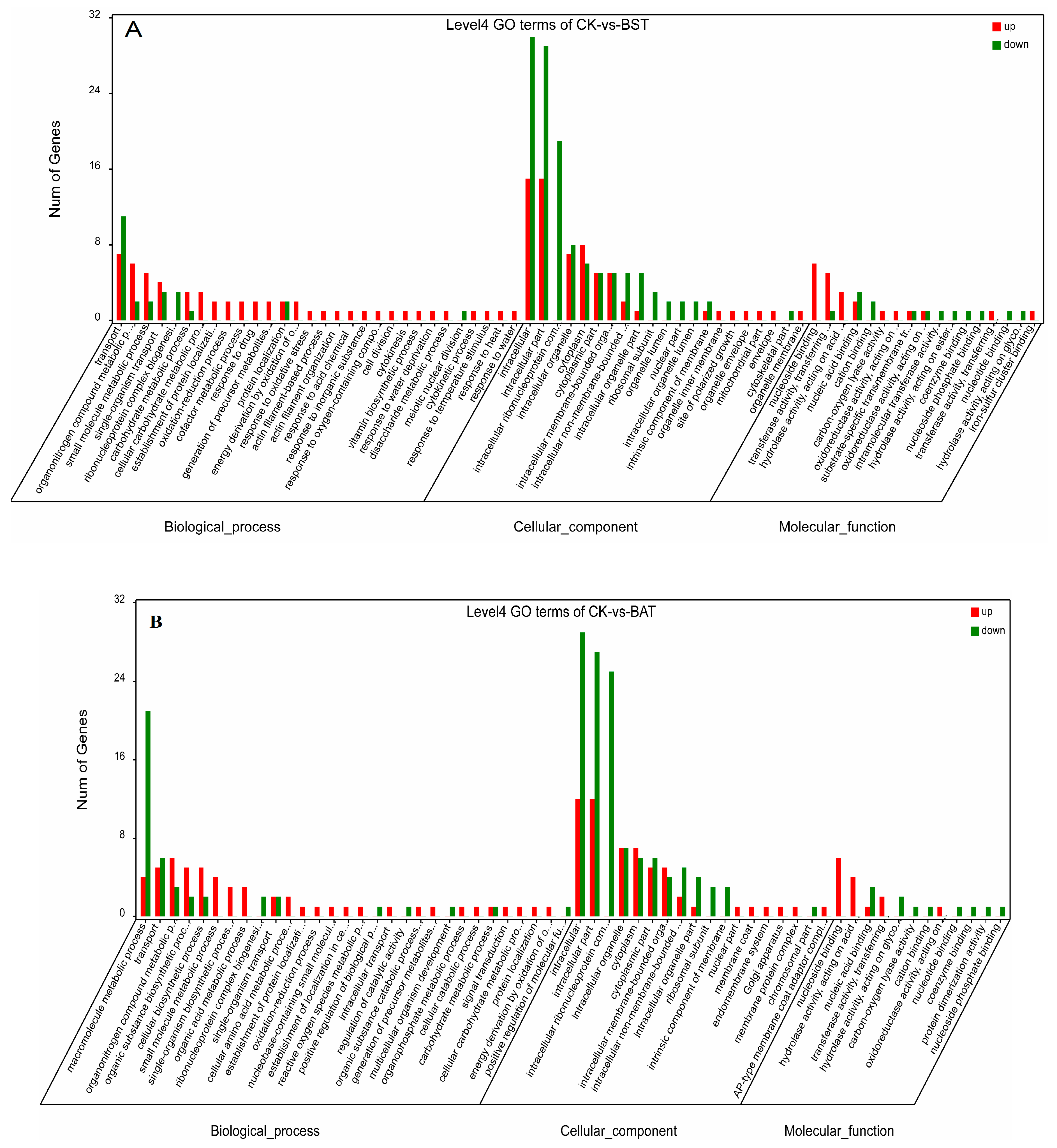
2.5. GO Enrichment Analysis
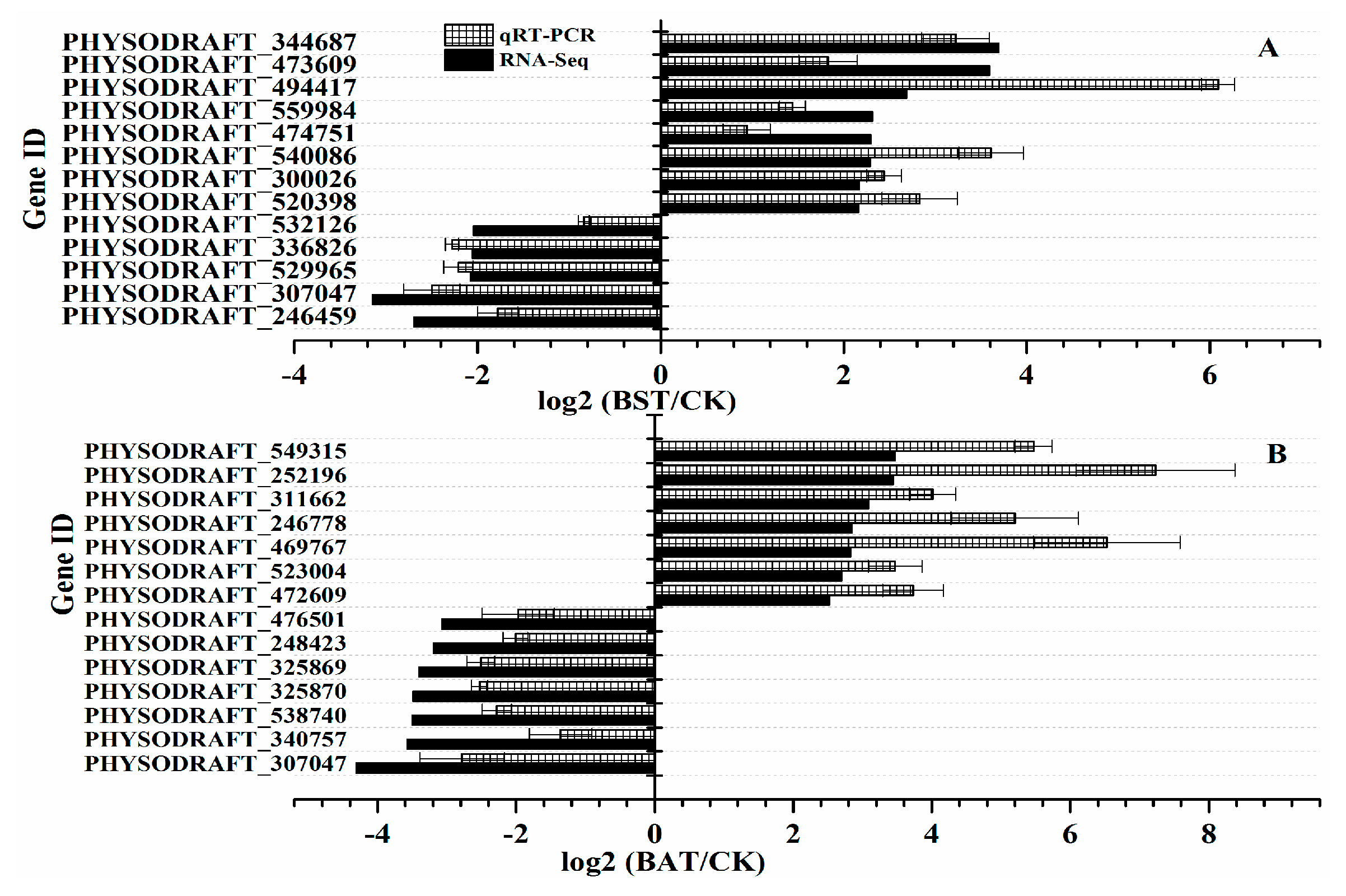
2.6. Validation of RNA-Seq Sequencing
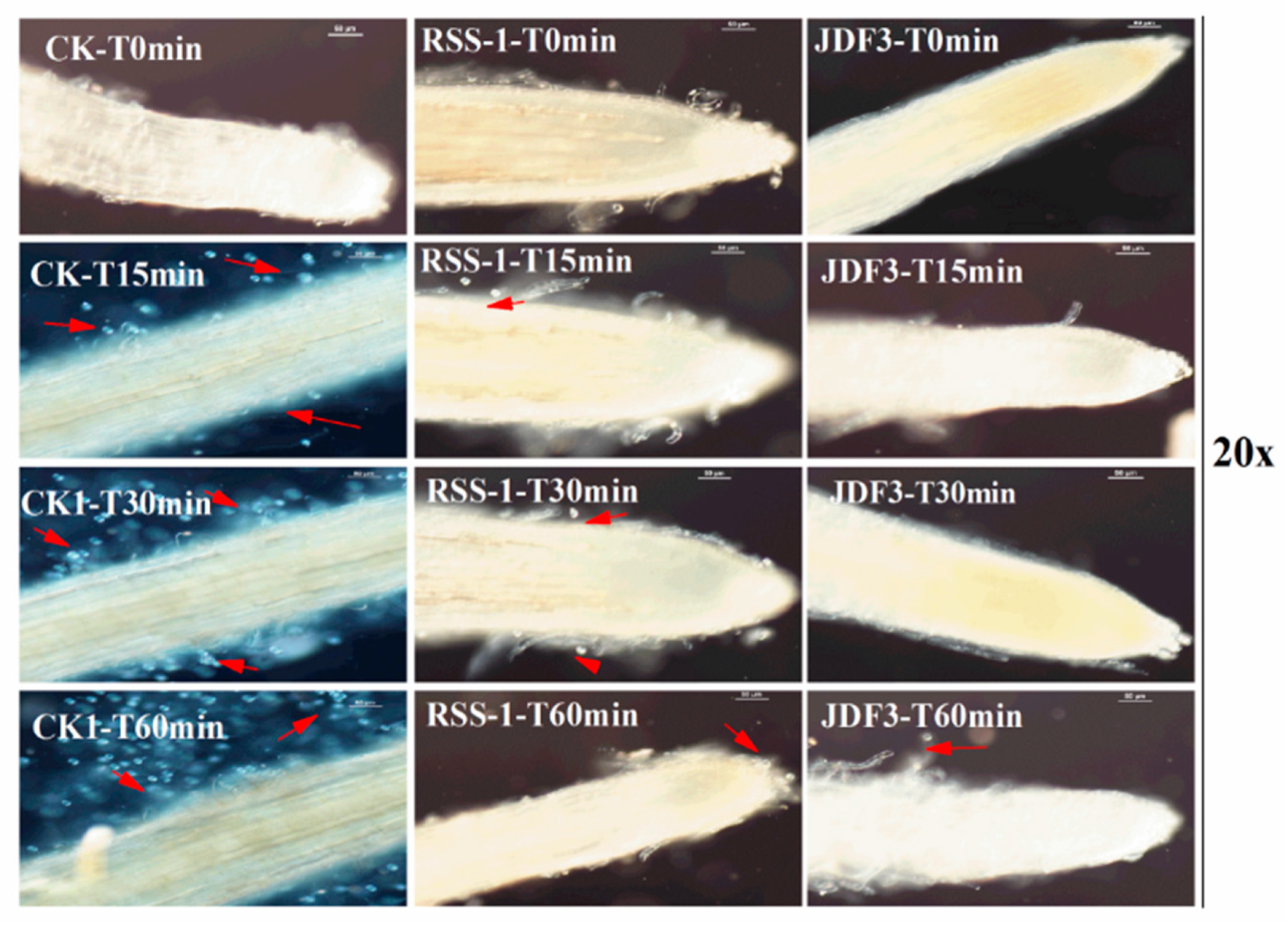
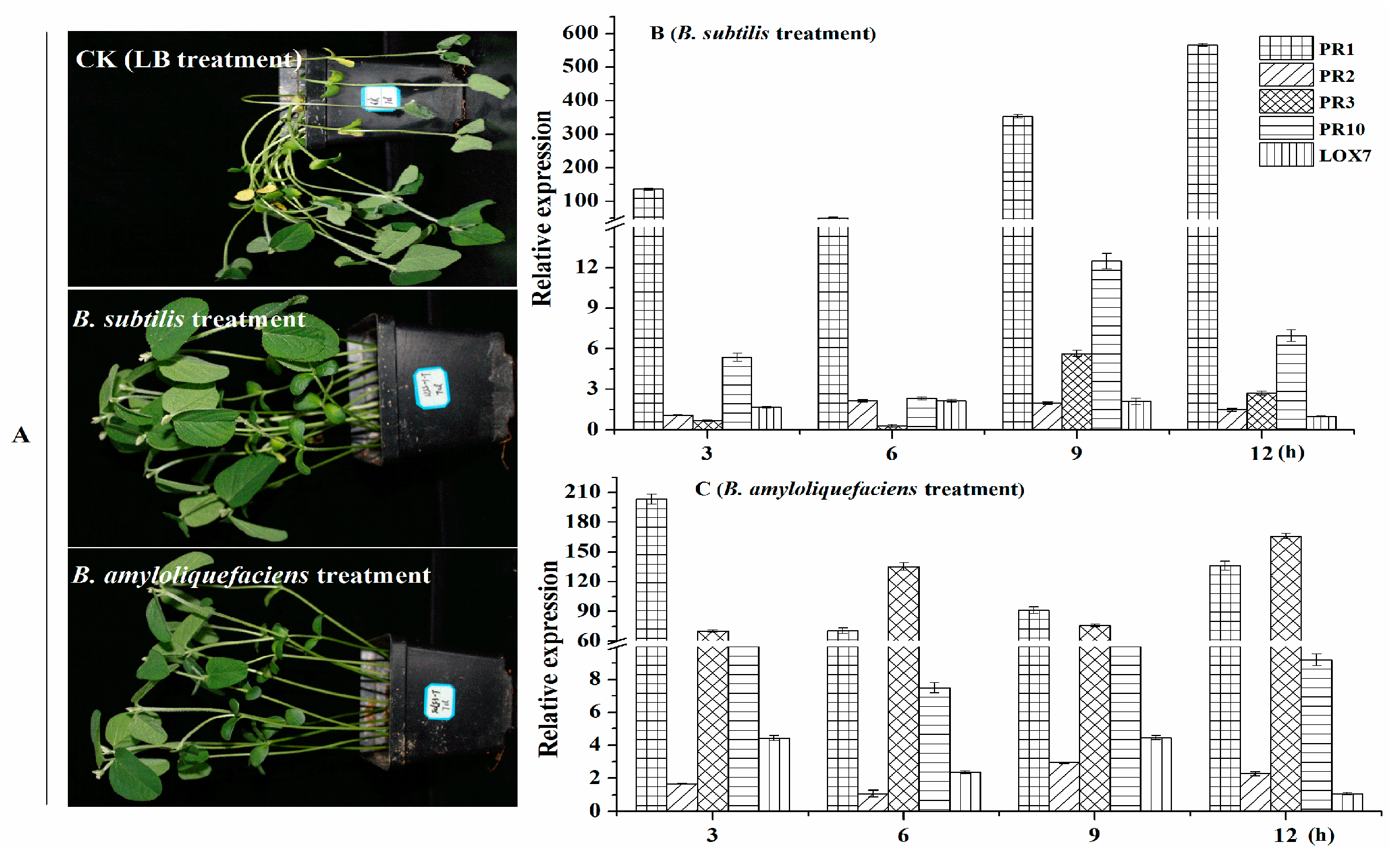
2.7. Effect of B. amyloliquefaciens and B. subtilis on the Control of the Phytophthora Root Rot of Soybean
2.8. B. amyloliquefaciens Fermentation Broth (BAFB) Activates Soybean Plant Defense Responses
3. Materials and Methods
3.1. Strains and Culture Conditions
3.2. RNA Extraction, cDNA Library Preparation. and Transcriptome Sequencing
3.3. RNA-Seq Data Analysis
3.4. Gene Annotation
3.5. Quantitative Real-Time PCR (qRT-PCR)
3.6. Effect of B. amyloliquefaciens and B. subtilis on the Control of Phytophthora Root Rot of Soybean
3.7. BAFB Activates Soybean Defense Responses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, D.; Li, P.; Hu, J.; Li, K.; Zhao, Z.; Wang, W.; Zhang, J.Y.; Ding, X.; Gao, Z. Genetic Diversity among Isolates of Phytophthora sojae in Anhui Province of China based on ISSR-PCR Markers. J. Am. Soc. Hortic. Sci. 2018, 143, 304–309. [Google Scholar] [CrossRef]
- Lu, X.X.; Zhou, D.M.; Chen, X.; Zhang, J.F.; Huang, H.W.; Wei, L.H. Isolation and characterization of Bacillus altitudinis JSCX-1 as a new potential biocontrol agent against Phytophthora sojae in soybean. Plant Soil 2017, 416, 53–66. [Google Scholar] [CrossRef]
- Bhat, R.G.; McBlain, B.A.; Schmitthenner, A.F. The inheritance of resistance to metalaxyl and to fluorophenylalanine in matings of homothallic phytophthora sojae. Mycol. Res. 1993, 97, 865–870. [Google Scholar] [CrossRef]
- Cai, M.; Miao, J.; Song, X.; Lin, D.; Bi, Y.; Chen, L.; Liu, X.L.; Tyler, B.M. C239S Mutation in the β-Tubulin of Phytophthora sojae confers resistance to zoxamide. Front. Microbiol. 2016, 7, 762. [Google Scholar] [CrossRef] [PubMed]
- Dorrance, A.E. Management of Phytophthora sojae of soybean: A review and future perspectives. Can. J. Plant Pathol. 2018, 40, 210–219. [Google Scholar] [CrossRef]
- Han, Q.; Zhao, H.; Huang, L.; Buchenauer, H.; Zuo, Y.; Kang, Z. Light and electron microscopy studies on the infection of a wild-type and metalaxyl-resistant isolate of phytophthora sojae in soybean hypocotyls. J. Phytopathol. 2011, 159, 368–376. [Google Scholar] [CrossRef]
- Miao, J.; Chi, Y.; Lin, D.; Tyler, B.M.; Liu, X.L. Mutations in ORP1 conferring oxathiapiprolin resistance confirmed by genome editing using CRISPR/Cas9 in Phytophthora capsici and Phytophthora sojae. Phytopathology 2018, 108, 1412–1419. [Google Scholar] [CrossRef] [PubMed]
- Randall, E.; Young, V.; Sierotzki, H.; Scalliet, G.; Birch, P.R.; Cooke, D.E.; Csukai, M.; Whisson, S.C. Sequence diversity in the large subunit of RNA polymerase I contributes to M efenoxam insensitivity in Phytophthora infestans. Mol. Plant Pathol. 2014, 15, 664–676. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42–a review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef]
- Maksimov, I.V.; Abizgil’Dina, R.R.; Pusenkova, L.I. Plant growth promoting rhizobacteria as alternative to chemical crop protectors from pathogens. Appl. Biochem. Microbiol. 2011, 47, 333–345. [Google Scholar] [CrossRef]
- Wagner, A.; Norris, S.; Chatterjee, P.; Morris, P.F.; Wildschutte, H. Aquatic Pseudomonads Inhibit Oomycete Plant Pathogens of Glycine max. Front. Microbiol. 2018, 9, 1007. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, S.A.; Silby, M.W.; Falloon, P.G.; Mahanty, H.K. Biological control of Phytophthora megasperma var. sojae, causal agent of Phytophthora rot of asparagus, by Pseudomonas aureofaciens PA147-2: A preliminary field trial. N. Z. J. Crop Hortic. Sci. 2000, 28, 97–103. [Google Scholar] [CrossRef][Green Version]
- Hou, J.; Bi, S.; Yan, L.; Zuo, Y.; Wang, Y.; Liu, T.; Zhu, J. Biological potential of Pseudomonas sp. BS1 in the control of Phytophthora root rot of soybean. Afr. J. Microbiol. Res. 2012, 6, 3589–3593. [Google Scholar]
- Sowanpreecha, R.; Rerngsamran, P. Biocontrol of Orchid-pathogenic Mold, Phytophthora palmivora, by Antifungal Proteins from Pseudomonas aeruginosa RS1. Mycobiology 2018, 46, 129–135. [Google Scholar] [CrossRef]
- Du, Q.; Yang, X.; Zhang, J.; Zhong, X.; Kim, K.S.; Yang, J.; Xing, G.J.; Li, X.Y.; Jiang, Z.Y.; Li, Q.Y.; et al. Over-expression of the Pseudomonas syringae harpin-encoding gene hrpZm confers enhanced tolerance to Phytophthora root and stem rot in transgenic soybean. Transgenic Res. 2018, 27, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xu, Y.; Lai, X. Antagonistic endophytic bacteria associated with nodules of soybean (Glycine max L.) and plant growth-promoting properties. Braz. J. Microbiol. 2018, 49, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Arfaoui, A.; Adam, L.R.; Bezzahou, A.; Daayf, F. Isolation and identification of cultivated bacteria associated with soybeans and their biocontrol activity against Phytophthora sojae. BioControl 2018, 63, 607–617. [Google Scholar] [CrossRef]
- Gao, Y.M.; Wang, X.J.; Zhang, J.; Li, M.; Liu, C.X.; An, J.; Jiang, L.; Xiang, W.S. Borrelidin, a potent antifungal agent: Insight into the antifungal mechanism against Phytophthora sojae. J. Agric. Food Chem. 2012, 60, 9874–9881. [Google Scholar] [CrossRef]
- Syed-Ab-Rahman, S.F.; Carvalhais, L.C.; Chua, E.; Xiao, Y.; Wass, T.J.; Schenk, P.M. Identification of soil bacterial isolates suppressing different Phytophthora spp. and promoting plant growth. Front. Plant Sci. 2018, 9, 1502. [Google Scholar] [CrossRef]
- Metzker, M.L. Sequencing technologies-the next generation. Nat. Rev. Genet. 2010, 11, 31–46. [Google Scholar] [CrossRef]
- Tripathy, S.; Deo, T.; Tyler, B.M. Oomycete Transcriptomics Database: A resource for oomycete transcriptomes. BMC Genom. 2012, 13, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Wang, X.; Tao, K.; Lu, Y.; Dai, T.; Dong, S.; Gijzen, M.; Wang, Y. Digital gene expression profiling of the Phytophthora sojae transcriptome. Mol. Plant Microbe Interact. 2011, 24, 1530–1539. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, X.; Rui, H.; Liu, T.; Hou, J. Cold temperature regulation of zoospore release in Phytophthora sojae: The genes that differentially expressed by cold temperature. Russ. J. Genet. 2018, 54, 618–628. [Google Scholar] [CrossRef]
- Chen, X.R.; Xing, Y.P.; Li, Y.P.; Tong, Y.H.; Xu, J.Y. RNA-Seq reveals infection-related gene expression changes in Phytophthora capsici. PLoS ONE 2013, 8, e74588. [Google Scholar] [CrossRef] [PubMed]
- Zuluaga, A.P.; Vega-Arreguín, J.C.; Fei, Z.; Ponnala, L.; Lee, S.J.; Matas, A.J.; Patev, S.; Fry, W.E.; Rose, J.K. Transcriptional dynamics of Phytophthora infestans during sequential stages of hemibiotrophic infection of tomato. Mol. Plant Pathol. 2016, 17, 29–41. [Google Scholar] [CrossRef]
- Gkarmiri, K.; Finlay, R.D.; Alström, S.; Thomas, E.; Cubeta, M.A.; Högberg, N. Transcriptomic changes in the plant pathogenic fungus Rhizoctonia solani AG-3 in response to the antagonistic bacteria Serratia proteamaculans and Serratia plymuthica. BMC Genom. 2015, 16, 630. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Fan, D.; Li, Y.; Chen, Y.; Huang, L.; Yan, X. Transcriptome analysis of valsa mali reveals its response mechanism to the biocontrol actinomycete saccharothrix yanglingensis hhs.015. BMC Microbiol. 2018, 18, 2–13. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, X.; Reiter, R.J.; Feng, S.; Wang, Y.; Liu, S.; Jin, L.; Li, Z.G.; Ren, M. Melatonin attenuates potato late blight by disrupting cell growth, stress tolerance, fungicide susceptibility and homeostasis of gene expression in Phytophthora infestans. Front. Plant Sci. 2017, 8, 1993. [Google Scholar] [CrossRef]
- Mei, X.Y.; Liu, Y.X.; Huang, H.C.; Du, F.; Huang, L.L.; Wu, J.Q.; Li, Y.W.; Zhu, S.S.; Yang, M. Benzothiazole inhibits the growth of Phytophthora capsici through inducing apoptosis and suppressing stress responses and metabolic detoxification. Pestic. Biochem. Phys. 2019, 154, 7–16. [Google Scholar] [CrossRef]
- Fromont-Racine, M.; Senger, B.; Saveanu, C.; Fasiolo, F. Ribosome assembly in eukaryotes. Gene 2003, 313, 17–42. [Google Scholar] [CrossRef]
- Yusupova, G.; Yusupov, M. High-resolution structure of the eukaryotic 80S ribosome. Annu. Rev. Biochem. 2014, 83, 467–486. [Google Scholar] [CrossRef] [PubMed]
- Dibble, C.C.; Manning, B.D. Signal integration by mTORC1 coordinates nutrient input with biosynthetic output. Nat. Cell. Biol. 2013, 15, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Donati, G.; Montanaro, L.; Derenzini, M. Ribosome biogenesis and control of cell proliferation: p53 is not alone. Cancer Res. 2012, 72, 1602–1607. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, S.; Merrill, A.H., Jr. Sphingolipid metabolism and cell growth regulation. FASEB J. 1996, 10, 1388–1397. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Ye, T.; Gao, Z. Cloning and functional analysis of succinate dehydrogenase gene PsSDHA in Phytophthora sojae. Microb. Pathog. 2017, 108, 40–48. [Google Scholar] [CrossRef]
- Pan, Y.; Ye, T.; Gao, Z. The succinate dehydrogenase PsSDHB is involved in hyphal morphology, chemical stress response and pathogenicity of Phytophthora sojae. Physiol. Mol. Plant Pathol. 2018, 102, 8–16. [Google Scholar] [CrossRef]
- Zhao, W.; Dong, S.; Ye, W.; Hua, C.C.; Meijer, H.J.G.; Dou, D.L.; Govers, F.; Wang, Y.C. Genome-wide identification of Phytophthora sojae SNARE genes and functional characterization of the conserved SNARE PsYKT6. Fungal Genet. Biol. 2011, 48, 241–251. [Google Scholar] [CrossRef]
- Ayers, A.R.; Valent, B.; Ebel, J.; Albersheim, P. Host-Pathogen Interactions: XI. Composition and structure of wall-released elicitor fractions. Plant Physiol. 1976, 57, 766–774. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, X.; Wang, W.; Geng, X.; Shi, Y.; Na, R.; Dou, D.L.; Li, H.L. Network and role analysis of autophagy in phytophthora sojae. Sci. Rep. 2017, 7, 1879. [Google Scholar] [CrossRef]
- Luo, Q.; Wang, F.X.; Zhong, N.Q.; Wang, H.Y.; Xia, G.X. The role of autophagy during development of the oomycete pathogen Phytophthora infestans. J. Genet. Genom. 2014, 41, 225–228. [Google Scholar] [CrossRef]
- Pollack, J.K.; Harris, S.D.; Marten, M.R. Autophagy in filamentous fungi. Fungal Genet. Biol. 2009, 46, 1–8. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Emr, S.D. Autophagy as a regulated pathway of cellular degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Wang, Y.; Tao, K.; Dong, S.; Huang, Q.; Dai, T.; Wang, Y. PsSAK1, a stress-activated MAP kinase of Phytophthora sojae, is required for zoospore viability and infection of soybean. Mol. Plant Microbe Interact. 2010, 23, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Cao, M.N.; Ye, W.W.; Li, H.Y.; Kong, L.; Zheng, X.; Wang, Y.C. PsMPK7, a stress-associated mitogen-activated protein kinase (MAPK) in phytophthora sojae, is required for stress tolerance, reactive oxygenated species detoxification, cyst germination, sexual reproduction and infection of soybean. Mol. Plant Pathol. 2015, 16, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Morrison, C.K.; Arseneault, T.; Novinscak, A.; Filion, M. Phenazine-1-carboxylic acid production by Pseudomonas fluorescens LBUM636 alters Phytophthora infestans growth and late blight development. Phytopathology 2016, 107, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Roquigny, R.; Novinscak, A.; Arseneault, T.; Joly, D.L.; Filion, M. Transcriptome alteration in Phytophthora infestans in response to phenazine-1-carboxylic acid production by Pseudomonas fluorescens strain LBUM223. BMC Genom. 2018, 19, 474–489. [Google Scholar] [CrossRef] [PubMed]
- Upchurch, R.G.; Ramirez, M.E. Defense-related gene expression in soybean leaves and seeds inoculated with Cercospora kikuchii and Diaporthe phaseolorum var. meridionalis. Physiol. Mol. Plant Pathol. 2010, 75, 64–70. [Google Scholar] [CrossRef]
- Upchurch, R.G.; Ramirez, M.E. Effects of temperature during soybean seed development on defense-related gene expression and fungal pathogen accumulation. Biotechnol. Lett. 2011, 33, 2397–2404. [Google Scholar] [CrossRef]
- Pawlowski, M.L.; Bowen, C.R.; Hill, C.B.; Hartman, G.L. Responses of soybean genotypes to pathogen infection after the application of elicitors. Crop Prot. 2016, 87, 78–84. [Google Scholar] [CrossRef]
- Wang, N.; Liu, M.; Guo, L.; Yang, X.; Qiu, D. A novel protein elicitor (PeBA1) from Bacillus amyloliquefaciens NC6 induces systemic resistance in tobacco. Int. J. Biol. Sci. 2016, 12, 757. [Google Scholar] [CrossRef]
- Yamamoto, S.; Shiraishi, S.; Suzuki, S. Are cyclic lipopeptides produced by Bacillus amyloliquefaciens S13-3 responsible for the plant defence response in strawberry against Colletotrichum gloeosporioides. Lett. Appl. Microbiol. 2015, 60, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Delledonne, M.; Xia, Y.; Dixon, R.A.; Lamb, C. Nitric oxide functions as a signal in plant disease resistance. Nature 1998, 394, 585. [Google Scholar] [CrossRef] [PubMed]
- Cui, T.B.; Chai, H.Y.; Jiang, L.X. Isolation and partial characterization of an antifungal protein produced by Bacillus licheniformis BS-3. Molecules 2012, 17, 7336–7347. [Google Scholar] [CrossRef] [PubMed]
- Suo, B.; Chen, Q.M.; Wu, W.X.; Wu, D.; Tian, M.; Jie, Y.; Zhang, B.; Wen, J.Z. Chemotactic responses of Phytophthora sojae zoospores to amino acids and sugars in root exudates. J. Gen. Plant Pathol. 2016, 82, 142–148. [Google Scholar] [CrossRef]
- Betsuyaku, S.; Katou, S.; Takebayashi, Y.; Sakakibara, H.; Nomura, N.; Fukuda, H. Salicylic acid and jasmonic acid pathways are activated in spatially different domains around the infection site during effector-triggered immunity in Arabidopsis thaliana. Plant Cell. Physiol. 2017, 59, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.M.; Wei, J.J.; Yao, C.C.; Reng, H.X.; Gao, Z.M. SsSm1, a Cerato-platanin family protein, is involved in the hyphal development and pathogenic process of Sclerotinia sclerotiorum. Plant Sci. 2018, 270, 37–46. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Raw Reads | Clean Reads | Q30 (%) | Total Reads | Mapped Reads | GC (%) |
|---|---|---|---|---|---|---|
| CK1 | 44778932 | 43841202 | 86.19 | 39868976 | 24694246 | 56.56 |
| CK2 | 47469548 | 45251026 | 85.26 | 44950158 | 25585591 | 53.88 |
| CK3 | 72226262 | 68948564 | 85.97 | 68250198 | 34299648 | 54.2 |
| BST1 | 47339402 | 46338742 | 84.74 | 46161358 | 24371232 | 56.87 |
| BST2 | 53546758 | 52564124 | 83.62 | 52223646 | 12675947 | 48.25 |
| BST3 | 44294010 | 43117708 | 84.12 | 42928882 | 14629394 | 48.64 |
| BAT1 | 51208004 | 50159286 | 85.89 | 49893596 | 29622070 | 57.73 |
| BAT2 | 51667698 | 50872290 | 85.20 | 50318686 | 10625344 | 55.25 |
| BAT3 | 46427178 | 45170256 | 85.48 | 44662734 | 15649105 | 56.96 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Li, K.; Hu, J.; Wang, W.; Liu, X.; Gao, Z. Biocontrol and Action Mechanism of Bacillus amyloliquefaciens and Bacillus subtilis in Soybean Phytophthora Blight. Int. J. Mol. Sci. 2019, 20, 2908. https://doi.org/10.3390/ijms20122908
Liu D, Li K, Hu J, Wang W, Liu X, Gao Z. Biocontrol and Action Mechanism of Bacillus amyloliquefaciens and Bacillus subtilis in Soybean Phytophthora Blight. International Journal of Molecular Sciences. 2019; 20(12):2908. https://doi.org/10.3390/ijms20122908
Chicago/Turabian StyleLiu, Dong, Kunyuan Li, Jiulong Hu, Weiyan Wang, Xiao Liu, and Zhimou Gao. 2019. "Biocontrol and Action Mechanism of Bacillus amyloliquefaciens and Bacillus subtilis in Soybean Phytophthora Blight" International Journal of Molecular Sciences 20, no. 12: 2908. https://doi.org/10.3390/ijms20122908
APA StyleLiu, D., Li, K., Hu, J., Wang, W., Liu, X., & Gao, Z. (2019). Biocontrol and Action Mechanism of Bacillus amyloliquefaciens and Bacillus subtilis in Soybean Phytophthora Blight. International Journal of Molecular Sciences, 20(12), 2908. https://doi.org/10.3390/ijms20122908