Detecting Brachypodium distachyon Chromosomes Bd4 and Bd5 in MH- and X-Ray-Induced Micronuclei Using mcFISH




Abstract
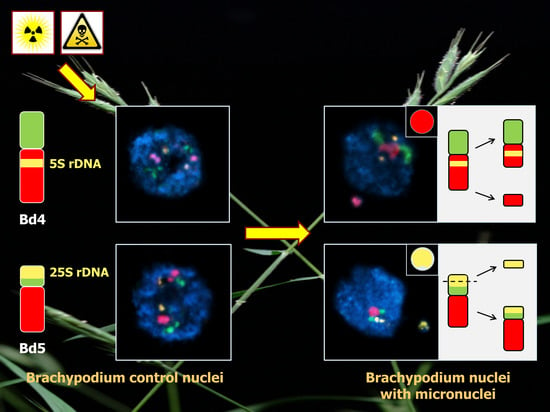
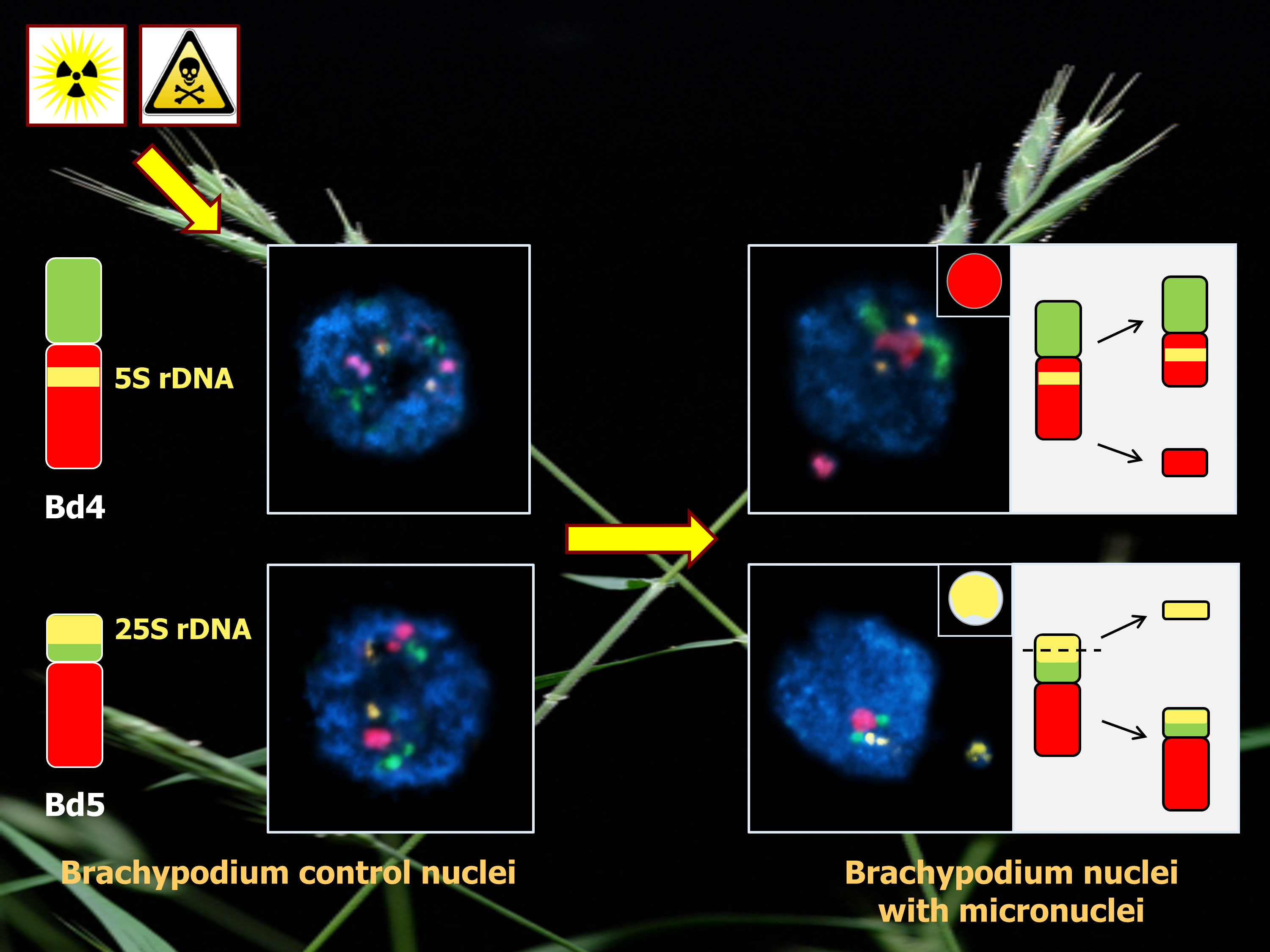{kind=link}
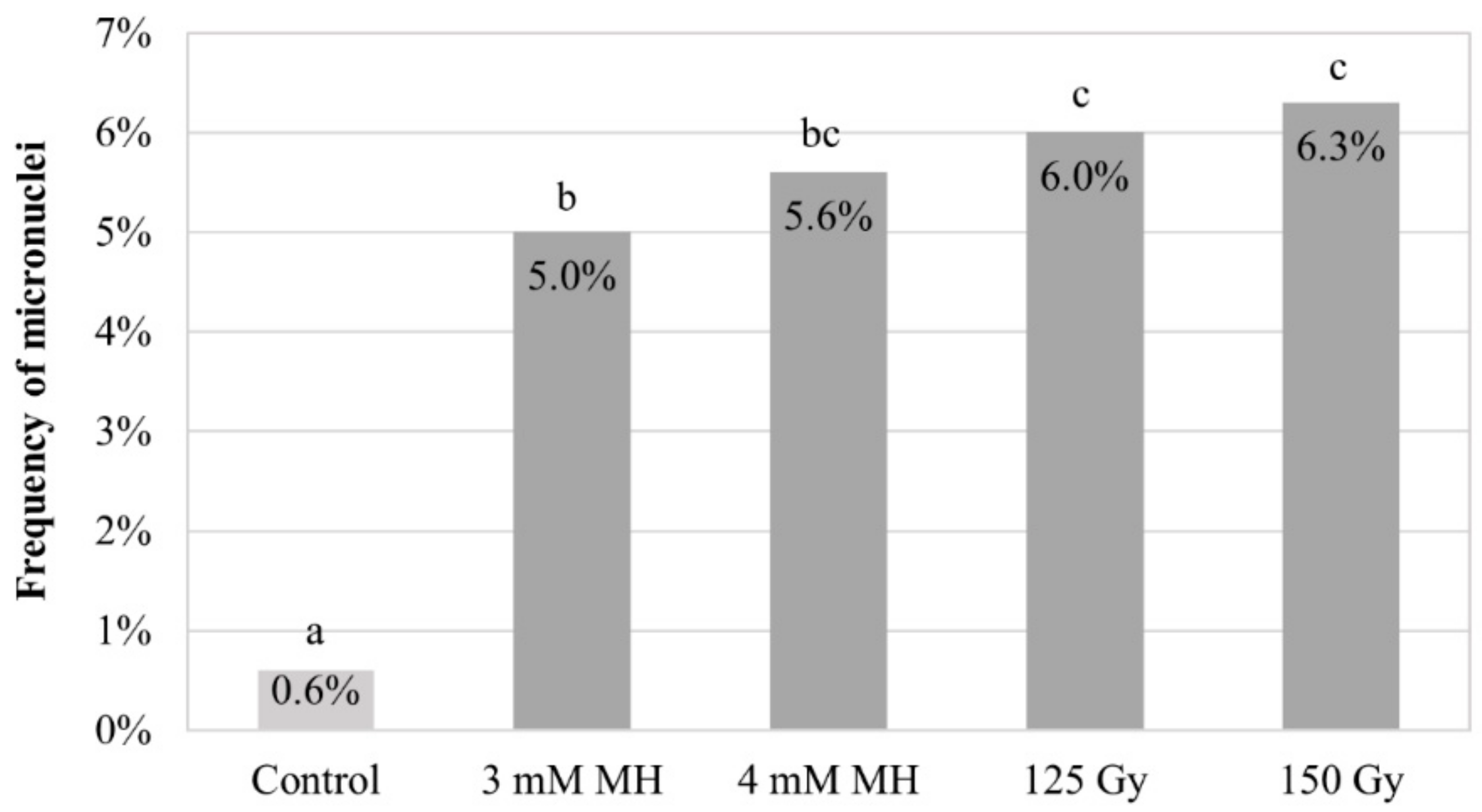{kind=link}
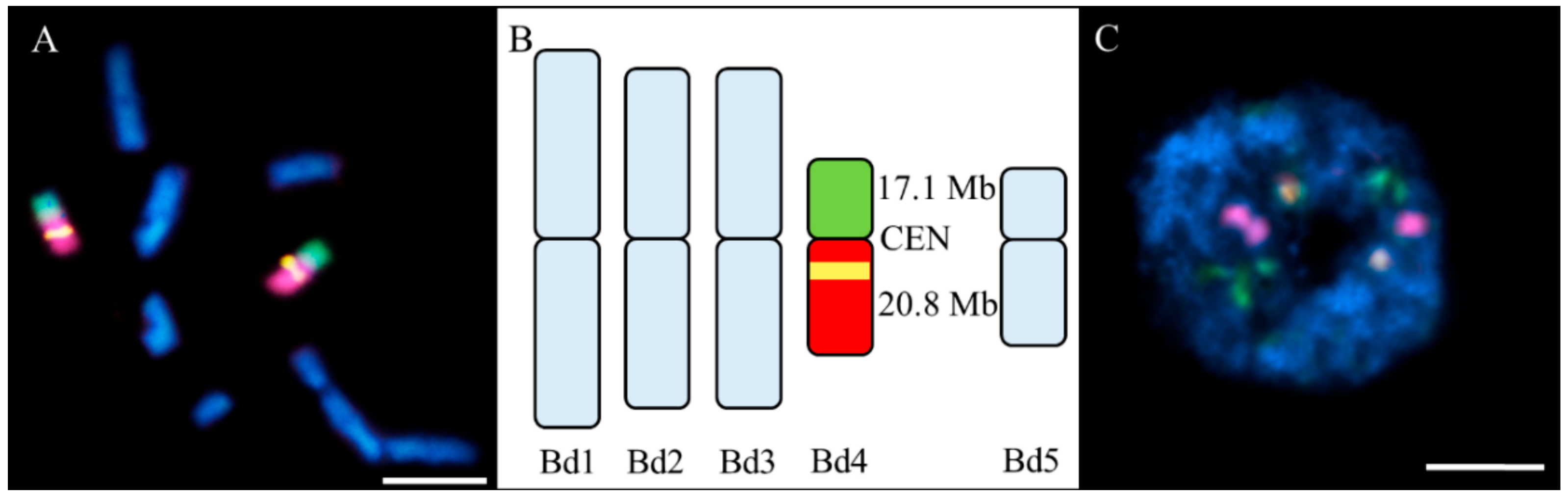{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
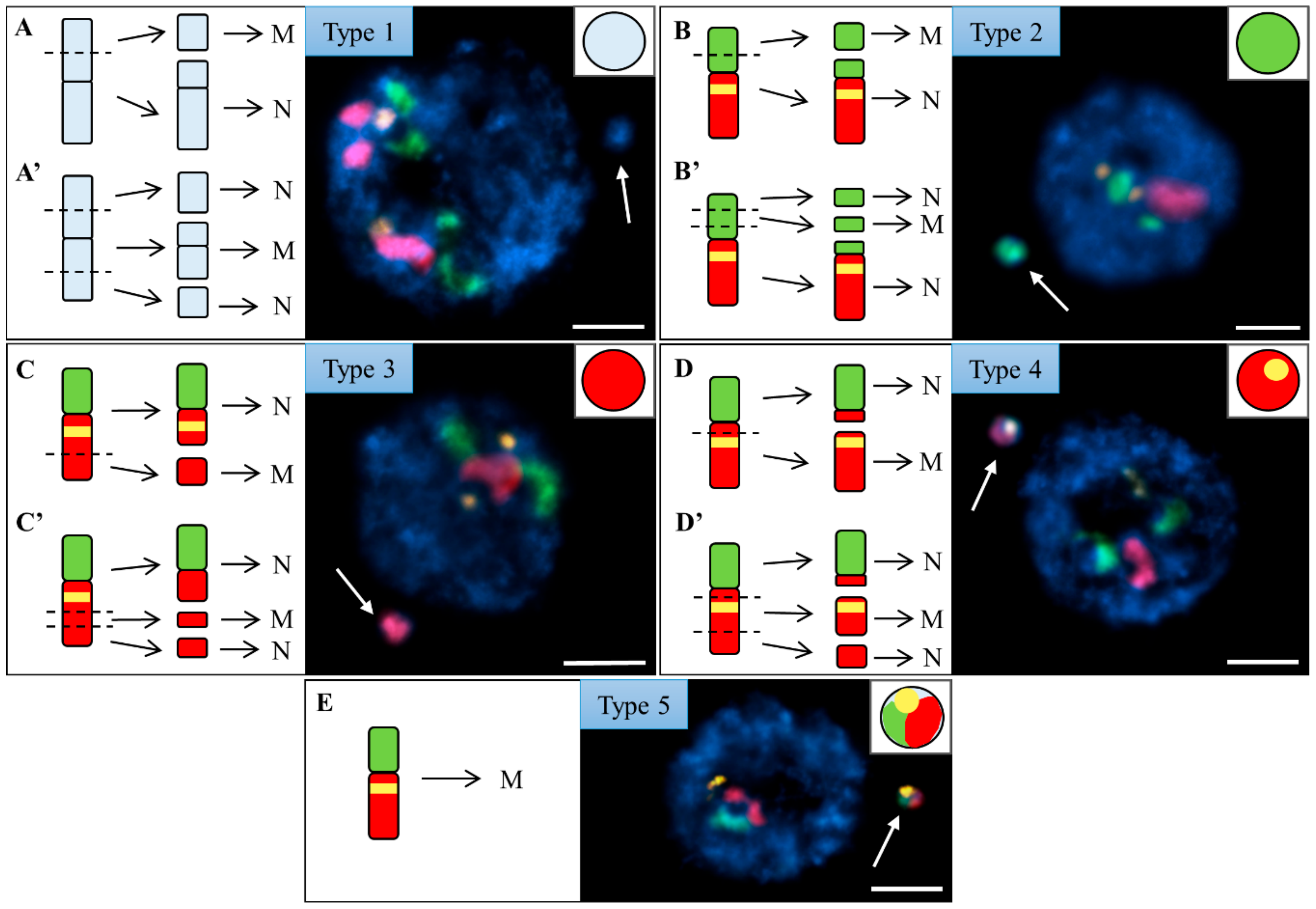
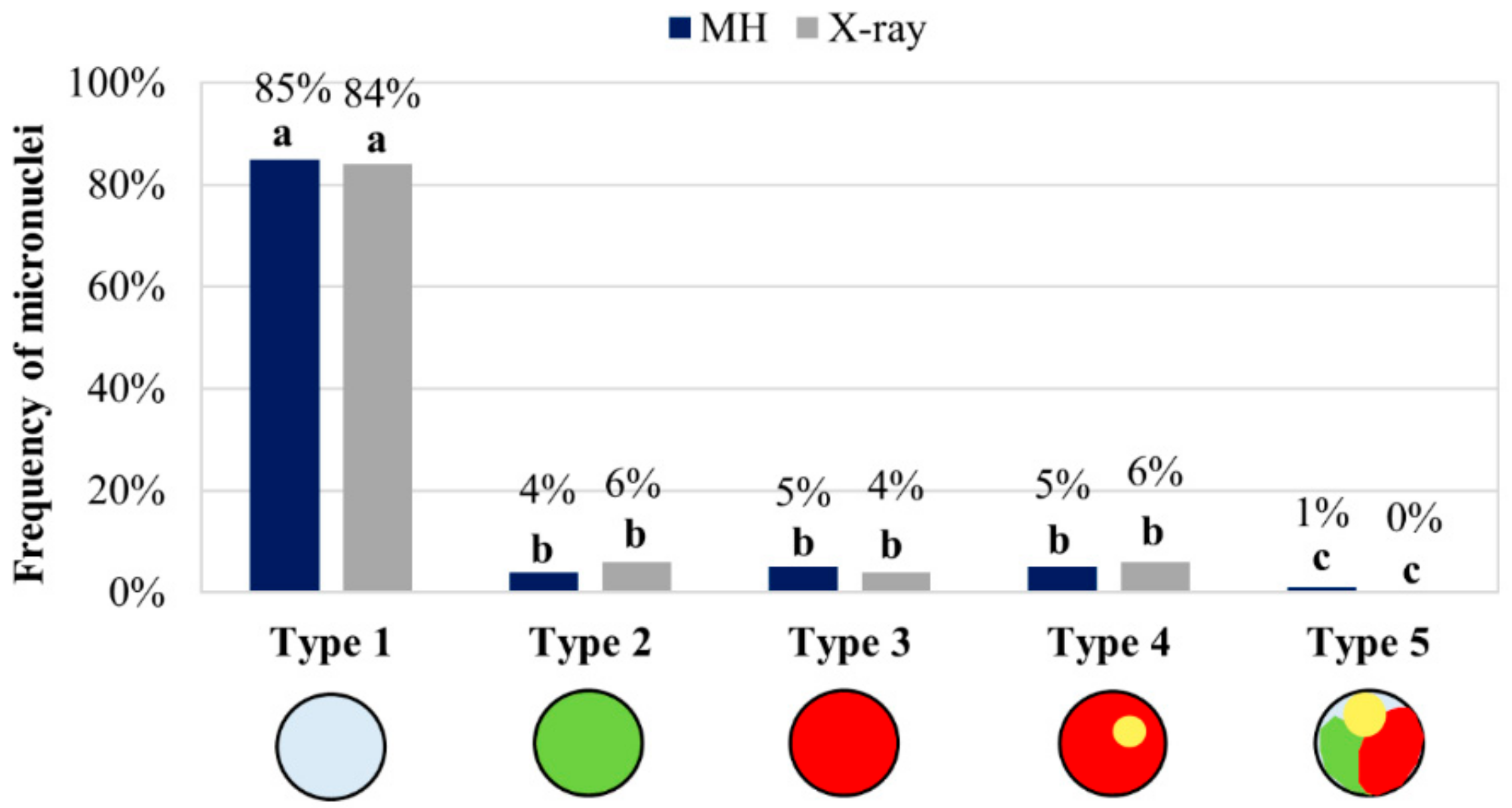
2.1. DSBs from MH Treatment and X-Radiation Are Non-Randomly Distributed along the Arms of Chromosome Bd4
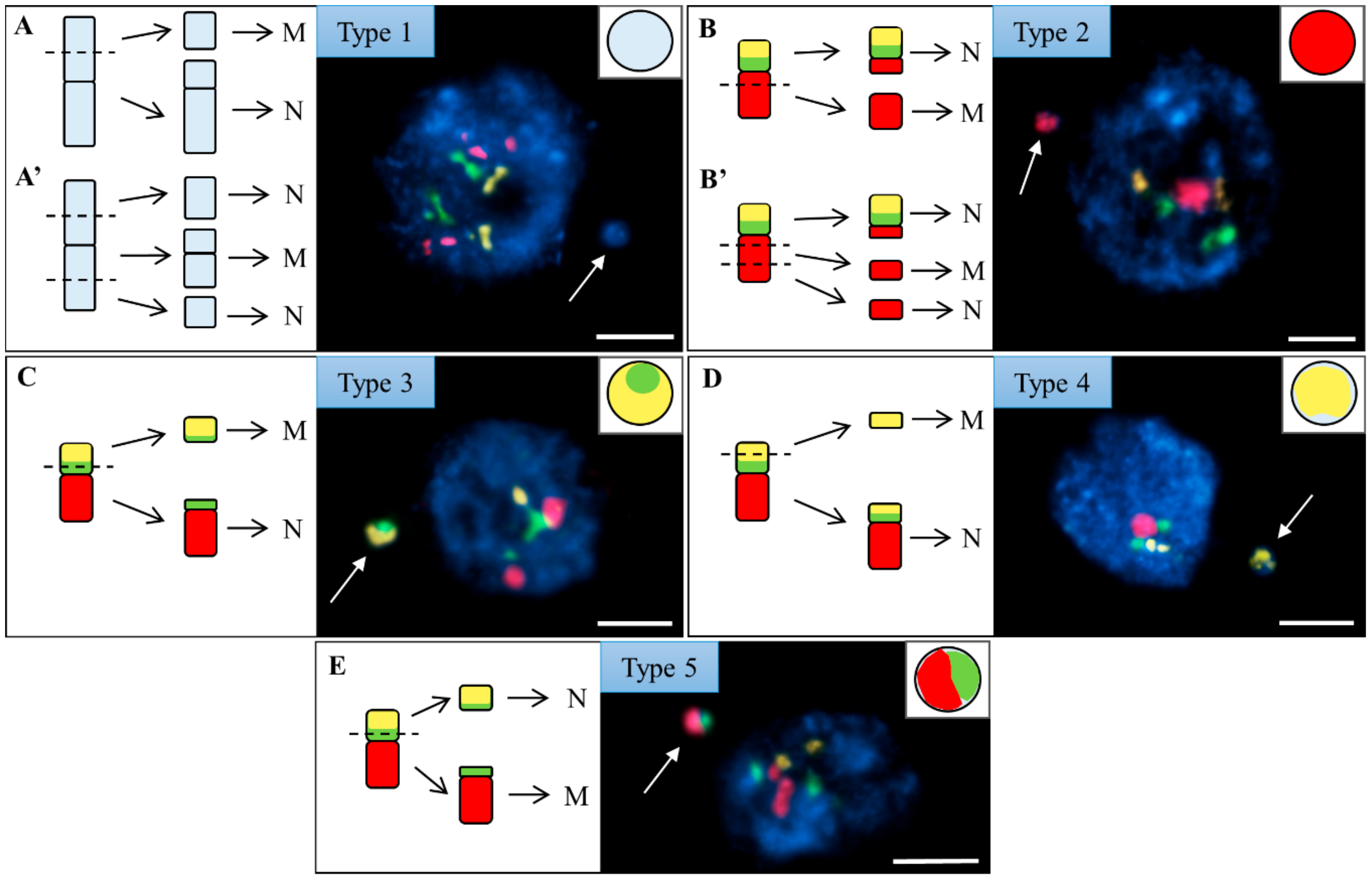
2.2. The Proximal Region of the Short Arm of Bd5 Is More Prone to DSBs Than Its Distal Part
3. Materials and Methods
3.1. Plant Material and Mutagenic Treatment
3.2. Root Meristem Preparation
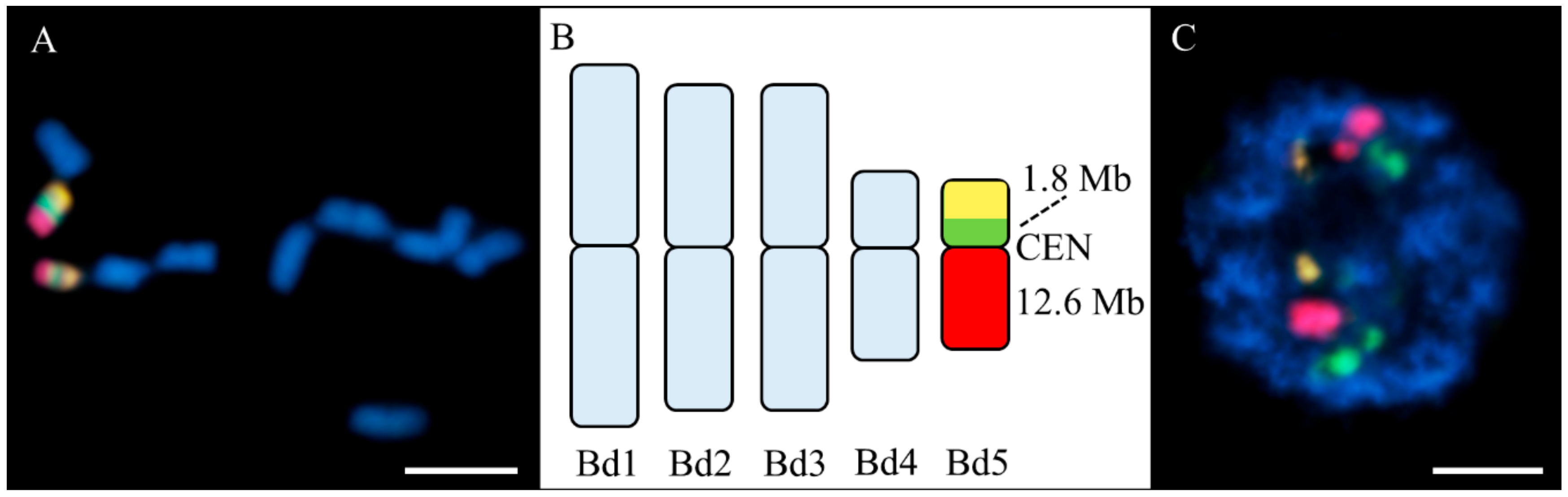
3.3. DNA Probes and FISH
3.4. Image Acquisition and Processing
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rocha, L.C.; Mittelmann, A.; Houben, A.; Techio, V.H. Fragile sites of 45S rDNA of Lolium multiflorum are not hotspots for chromosomal breakages induced by X-ray. Mol. Biol. Rep. 2016, 43, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Olasupo, F.O.; Ilori, C.O.; Forster, B.P.; Bado, S. Mutagenic effects of gamma radiation on eight accessions of cowpea (Vigna unguiculata [L.] Walp.). Am. J. Plant. Sci. 2016, 7, 339–351. [Google Scholar] [CrossRef]
- Kumar, G.; Pandey, A. Effect of UV-B radiation on chromosomal organisation and biochemical constituents of Coriandrum sativum L. Jordan J. Boil. Sci. 2017, 10, 85–93. [Google Scholar]
- Amin, R.; Wani, M.R.; Raina, A.; Khursheed, S.; Khan, S. Induced morphological and chromosomal diversity in the mutagenized population of black cumin (Nigella sativa l.) using single and combination treatments of gamma rays and ethyl methane sulfonate. Jordan J. Boil. Sci. 2019, 12, 23–30. [Google Scholar]
- Hsu, T.C. A possible function of constitutive heterochromatin: The bodyguard hypothesis. Genetics 1975, 137–150. [Google Scholar]
- Kwasniewska, J.; Mikolajczyk, A. Influence of the presence of B chromosomes on DNA damage in Crepis capillaris. PLoS ONE 2014, 9, e87337. [Google Scholar] [CrossRef] [PubMed]
- Schubert, I.; Rieger, R.; Fuchs, J.; Pich, U. Sequence organization and the mechanism of interstitial deletion clustering in a plant genome (Vicia faba). Mutat Res. 1994, 325, 1–5. [Google Scholar] [CrossRef]
- Schubert, I.; Pecinka, A.; Meister, A.; Schubert, V.; Klatte, M.; Jovtchev, G. DNA damage processing and aberration formation in plants. Cytogenet. Genome Res. 2004, 104, 104–108. [Google Scholar] [CrossRef]
- Larson, K.; Yan, S.J.; Tsurumi, A.; Liu, J.; Zhou, J.; Gaur, K.; Guo, D.; Eickbush, T.H.; Li, W.X. Heterochromatin formation promotes longevity and represses ribosomal RNA synthesis. PLoS Genet. 2012, 8, e1002473. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.J.; Lim, S.J.; Shi, S.; Dutta, P.; Li, W.X. Unphosphorylated stat and heterochromatin protect genome stability. FASEB J. 2011, 25, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Falk, M.; Lukasova, E.; Kozubek, S. Chromatin structure influences the sensitivity of DNA to gamma-radiation. Biochim. Biophys. Acta 2008, 1783, 2398–2414. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, A.T. Chromosome aberrations: Past, present and future. Mutat Res. 2002, 504, 3–16. [Google Scholar] [CrossRef]
- Hayashi, M. The micronucleus test-most widely used in vivo genotoxicity test. Genes Environ. 2016, 38, 18. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, L.B.; Bond, M.A.; Tobin, A.K.. Ultraviolet-B radiation reduces the rates of cell division and elongation in the primary leaf of wheat (Triticum aestivum L. cv Maris Huntsman). Plant Cell Environ. 2002, 25, 617–624. [Google Scholar] [CrossRef]
- Rank, J.; Lopez, L.C.; Nielsen, M.H.; Moretton, J. Genotoxicity of maleic hydrazide, acridine and DEHP in Allium cepa root cells performed by two different laboratories. Hereditas 2002, 136, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.; Morley, A.A. Measurement of micronuclei in lymphocytes. Mutat Res. 1985, 147, 29–36. [Google Scholar] [CrossRef]
- Fenech, M.; Kirsch-Volders, M.; Natarajan, A.T.; Surralles, J.; Crott, J.W.; Parry, J.; Norppa, H.; Eastmond, D.A.; Tucker, J.D.; Thomas, P. Molecular mechanisms of micronucleus, nucleoplasmic bridge and nuclear bud formation in mammalian and human cells. Mutagenesis 2011, 26, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Mateuca, R.; Lombaert, N.; Aka, P.V.; Decordier, I.; Kirsch-Volders, M. Chromosomal changes: Induction, detection methods and applicability in human biomonitoring. Biochimie 2006, 88, 1515–1531. [Google Scholar] [CrossRef]
- Luzhna, L.; Kathiria, P.; Kovalchuk, O. Micronuclei in genotoxicity assessment: From genetics to epigenetics and beyond. Front. Genet. 2013, 4, 131. [Google Scholar] [CrossRef]
- Evans, H.J.; Neary, G.J.; Williamson, F.S. The relative biological efficiency of single doses of fast neutrons and gamma-rays on Vicia faba roots and the effect of oxygen. Part ii. Chromosome damage: The production of micronuclei. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1959, 1, 216–229. [Google Scholar] [CrossRef]
- Jovtchev, G.; Stergios, M.; Schubert, I. A comparison of N-methyl-N-nitrosourea-induced chromatid aberrations and micronuclei in barley meristems using FISH techniques. Mutat. Res. 2002, 517, 47–51. [Google Scholar] [CrossRef]
- Juchimiuk, J.; Hering, B.; Maluszynska, J. Multicolour FISH in an analysis of chromosome aberrations induced by N-nitroso-N-methylurea and maleic hydrazide in barley cells. J. Appl. Genet. 2007, 48, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Juchimiuk-Kwasniewska, J.; Brodziak, L.; Maluszynska, J. FISH in analysis of gamma ray-induced micronuclei formation in barley. J. Appl. Genet. 2011, 52, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Maluszynska, J.; Juchimiuk, J.; Wolny, E. Chromosomal aberrations in Crepis capillaris cells detected by FISH. Folia Histochem. Cytobiol. 2003, 41, 101–104. [Google Scholar] [PubMed]
- Kus, A.; Kwasniewska, J.; Szymanowska-Pulka, J.; Hasterok, R. Dissecting the chromosomal composition of mutagen-induced micronuclei in Brachypodium distachyon using multicolour FISH. Ann. Bot. 2018, 122, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Fauth, E.; Zankl, H. Comparison of spontaneous and idoxuridine-induced micronuclei by chromosome painting. Mutat. Res. 1999, 440, 147–156. [Google Scholar] [CrossRef]
- Fimognari, C.; Sauer-Nehls, S.; Braselmann, H.; Nusse, M. Analysis of radiation-induced micronuclei by FISH using a combination of painting and centromeric DNA probes. Mutagenesis 1997, 12, 91–95. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Leach, N.T.; Jackson-Cook, C. The application of spectral karyotyping (SKY) and fluorescent in situ hybridization (FISH) technology to determine the chromosomal content(s) of micronuclei. Mutat. Res. 2001, 495, 11–19. [Google Scholar] [CrossRef]
- Chung, H.W.; Kang, S.J.; Kim, S.Y. A combination of the micronucleus assay and a FISH technique for evaluation of the genotoxicity of 1,2,4-benzenetriol. Mutat. Res. 2002, 516, 49–56. [Google Scholar] [CrossRef]
- Norppa, H.; Falck, G.C. What do human micronuclei contain? Mutagenesis 2003, 18, 221–233. [Google Scholar] [CrossRef]
- Hovhannisyan, G.; Aroutiounian, R.; Liehr, T. Chromosomal composition of micronuclei in human leukocytes exposed to mitomycin C. J. Histochem. Cytochem. 2012, 60, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Puerto, S.; Surralles, J.; Ramirez, M.J.; Creus, A.; Marcos, R. Equal induction and persistence of chromosome aberrations involving chromosomes with heterogeneous lengths and gene densities. Cytogenet. Cell. Genet. 1999, 87, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Surrallés, J.P.; Puerto, S.; Ramírez, M.J.; Creus, A.; Marcos, R.; Mullenders, L.H.; Natarajan, A.T. Links between chromatin structure, DNA repair and chromosome fragility. Mutat. Res. 1998, 404, 39–44. [Google Scholar] [CrossRef]
- Fauth, E.; Scherthan, H.; Zankl, H. Chromosome painting reveals specific patterns of chromosome occurrence in mitomycin C- and diethylstilboestrol-induced micronuclei. Mutagenesis 2000, 15, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Wuttke, K.; Streffer, C.; Muller, W.U. Detection of chromosome 2 and chromosome 7 within x-ray- or colchicine-induced micronuclei by fluorescence in situ hybridization. Mutagenesis 1997, 12, 55–59. [Google Scholar] [CrossRef] [PubMed]
- International Brachypodium Initiative. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Catalan, P.; Chalhoub, B.; Chochois, V.; Garvin, D.F.; Hasterok, R.; Manzaneda, A.J.; Mur, L.A.J.; Pecchioni, N.; Rasmussen, S.K.; Vogel, J.P.; et al. Update on the genomics and basic biology of Brachypodium. Trends Plant. Sci. 2014, 19, 414–418. [Google Scholar] [CrossRef]
- Mur, L.A.; Allainguillaume, J.; Catalan, P.; Hasterok, R.; Jenkins, G.; Lesniewska, K.; Thomas, I.; Vogel, J. Exploiting the Brachypodium tool box in cereal and grass research. New Phytol. 2011, 191, 334–347. [Google Scholar] [CrossRef]
- Kus, A.; Kwasniewska, J.; Hasterok, R. Brachypodium distachyon—A useful model in the qualification of mutagen-induced micronuclei using multicolor FISH. PLoS ONE 2017, 12, e0170618. [Google Scholar] [CrossRef]
- Lusinska, J.; Majka, J.; Betekhtin, A.; Susek, K.; Wolny, E.; Hasterok, R. Chromosome identification and reconstruction of evolutionary rearrangements in Brachypodium distachyon, B. stacei and B. hybridum. Ann. Bot. 2018, 122, 445–459. [Google Scholar] [CrossRef]
- Huang, J.; Ma, L.; Yang, F.; Fei, S.Z.; Li, L. 45s rDNA regions are chromosome fragile sites expressed as gaps in vitro on metaphase chromosomes of root-tip meristematic cells in Lolium spp. PLoS ONE 2008, 3, e2167. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Li, H.; Zhang, L.; Gao, F.; Wang, P.; Hu, Y.; Yan, S.; Zhao, L.; Zhang, Q.; Tan, J.; et al. Plant 45s rDNA clusters are fragile sites and their instability is associated with epigenetic alterations. PLoS ONE 2012, 7, e35139. [Google Scholar] [CrossRef] [PubMed]
- Waminal, N.E.; Ryu, K.H.; Choi, S.H.; Kim, H.H. Randomly detected genetically modified (GM) maize (Zea mays l.) near a transport route revealed a fragile 45s rDNA phenotype. PLoS ONE 2013, 8, e74060. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.C.; Bustamante Fde, O.; Silveira, R.A.; Torres, G.A.; Mittelmann, A.; Techio, V.H. Functional repetitive sequences and fragile sites in chromosomes of Lolium perenne L. Protoplasma 2015, 252, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.; Chen, C.L.; Miao, Y.; Yu, C.X.; Guo, W.W.; Xu, Q.; Deng, X.X. Fragile sites of ‘Valencia’ sweet orange (Citrus sinensis) chromosomes are related with active 45S rDNA. PLoS ONE 2016, 11, e0151512. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.I. Fragile and unstable chromosomes in cancer: Causes and consequences. Trends Genet. 2001, 17, 339–345. [Google Scholar] [CrossRef]
- Glover, T.W. Common fragile sites. Cancer Lett. 2006, 232, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Siroky, J.; Zluvova, J.; Riha, K.; Shippen, D.E.; Vyskot, B. Rearrangements of ribosomal DNA clusters in late generation telomerase-deficient Arabidopsis. Chromosoma 2003, 112, 116–123. [Google Scholar] [CrossRef]
- Ide, S.; Miyazaki, T.; Maki, H.; Kobayashi, T. Abundance of ribosomal RNA gene copies maintains genome integrity. Science 2010, 327, 693–696. [Google Scholar] [CrossRef]
- Kobayashi, T. Regulation of ribosomal RNA gene copy number and its role in modulating genome integrity and evolutionary adaptability in yeast. Cell. Mol. Life Sci. 2011, 68, 1395–1403. [Google Scholar] [CrossRef]
- Jenkins, G.; Hasterok, R. BAC ‘landing’ on chromosomes of Brachypodium distachyon for comparative genome alignment. Nat. Protoc. 2007, 2, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Febrer, M.; Goicoechea, J.L.; Wright, J.; McKenzie, N.; Song, X.; Lin, J.; Collura, K.; Wissotski, M.; Yu, Y.; Ammiraju, J.S.; et al. An integrated physical, genetic and cytogenetic map of Brachypodium distachyon, a model system for grass research. PLoS ONE 2010, 5, e13461. [Google Scholar] [CrossRef] [PubMed]
- Idziak, D.; Betekhtin, A.; Wolny, E.; Lesniewska, K.; Wright, J.; Febrer, M.; Bevan, M.W.; Jenkins, G.; Hasterok, R. Painting the chromosomes of Brachypodium: Current status and future prospects. Chromosoma 2011, 120, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Unfried, I.; Gruendler, P. Nucleotide sequence of the 5.8S and 25S rRNA genes and of the internal transcribed spacers from Arabidopsis thaliana. Nucleic Acids Res. 1990, 18, 4011. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, W.L.; Dyer, T.A. Sequence organization of the repeating units in the nucleus of wheat which contain 5S rRNA genes. Nucleic Acids Res. 1980, 8, 4851–4865. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kus, A.; Szymanowska-Pułka, J.; Kwasniewska, J.; Hasterok, R. Detecting Brachypodium distachyon Chromosomes Bd4 and Bd5 in MH- and X-Ray-Induced Micronuclei Using mcFISH. Int. J. Mol. Sci. 2019, 20, 2848. https://doi.org/10.3390/ijms20112848
Kus A, Szymanowska-Pułka J, Kwasniewska J, Hasterok R. Detecting Brachypodium distachyon Chromosomes Bd4 and Bd5 in MH- and X-Ray-Induced Micronuclei Using mcFISH. International Journal of Molecular Sciences. 2019; 20(11):2848. https://doi.org/10.3390/ijms20112848
Chicago/Turabian StyleKus, Arita, Joanna Szymanowska-Pułka, Jolanta Kwasniewska, and Robert Hasterok. 2019. "Detecting Brachypodium distachyon Chromosomes Bd4 and Bd5 in MH- and X-Ray-Induced Micronuclei Using mcFISH" International Journal of Molecular Sciences 20, no. 11: 2848. https://doi.org/10.3390/ijms20112848
APA StyleKus, A., Szymanowska-Pułka, J., Kwasniewska, J., & Hasterok, R. (2019). Detecting Brachypodium distachyon Chromosomes Bd4 and Bd5 in MH- and X-Ray-Induced Micronuclei Using mcFISH. International Journal of Molecular Sciences, 20(11), 2848. https://doi.org/10.3390/ijms20112848