Comprehensive Analysis of SnRK Gene Family and their Responses to Salt Stress in Eucalyptus grandis

Abstract
1. Introduction
2. Results
2.1. Identification of SnRK Genes in E. grandis
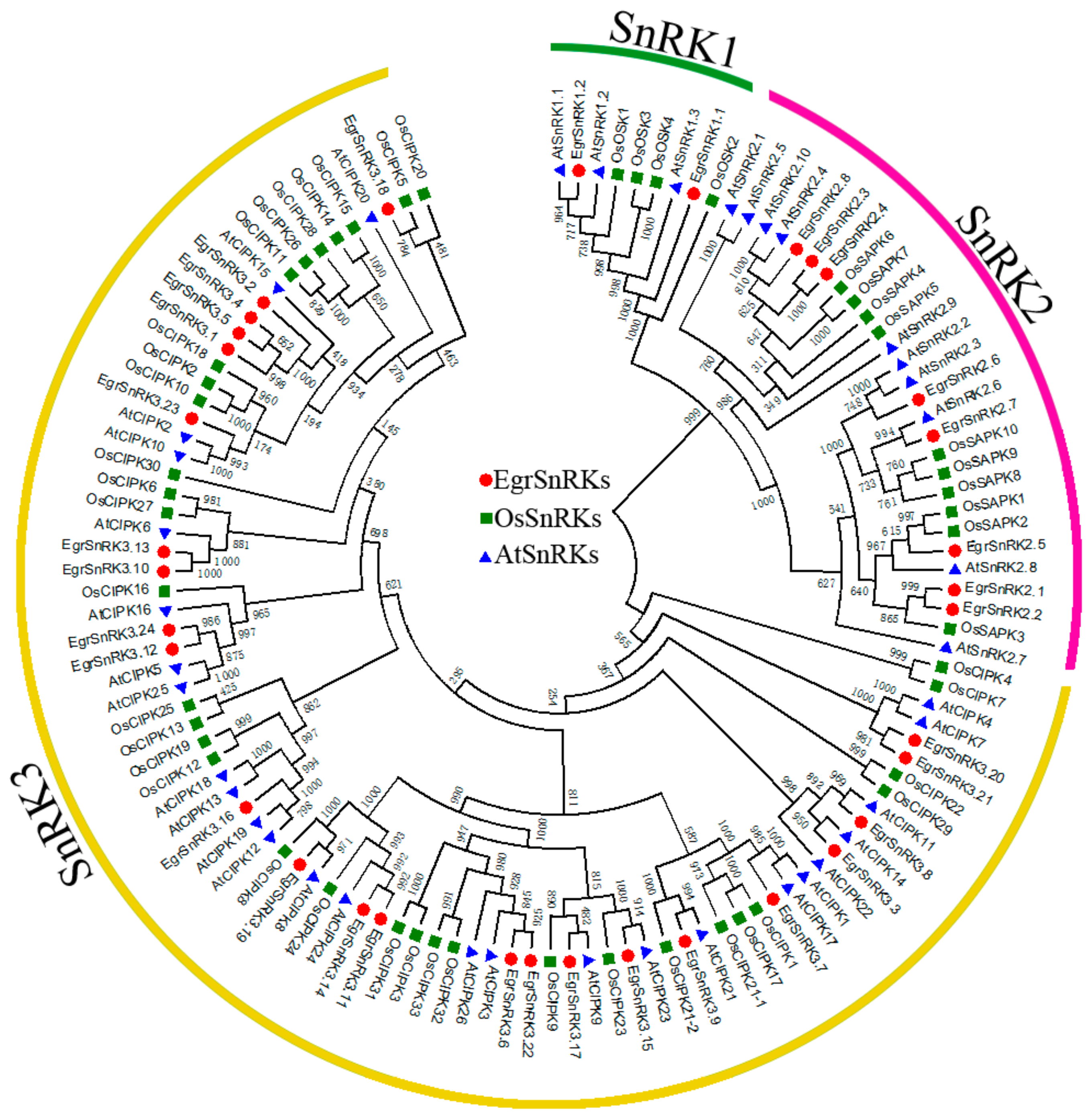
2.2. Phylogenetic Tree of SnRK Genes
2.3. Multiple Sequence Alignment of the SnRK Gene Family
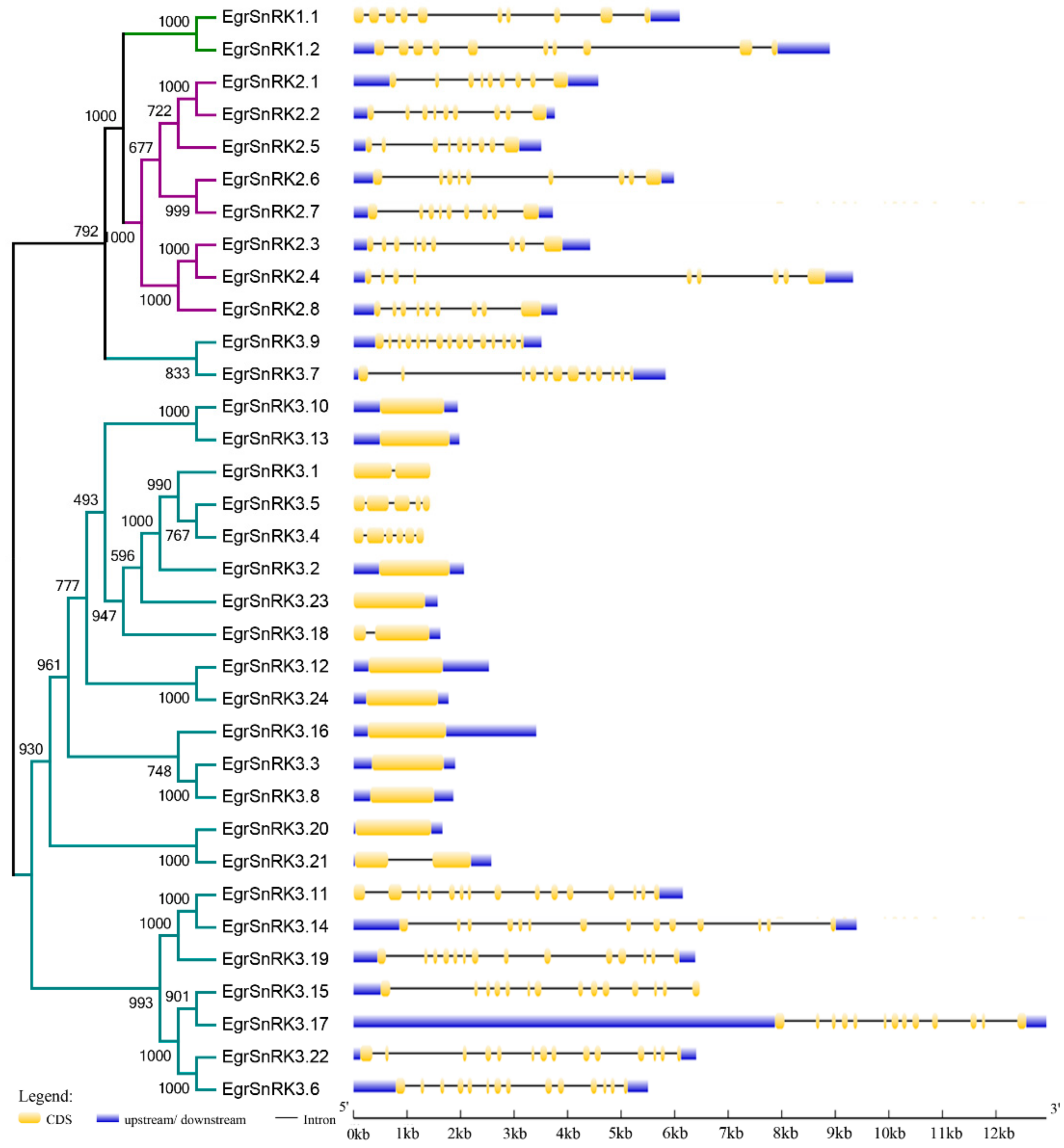
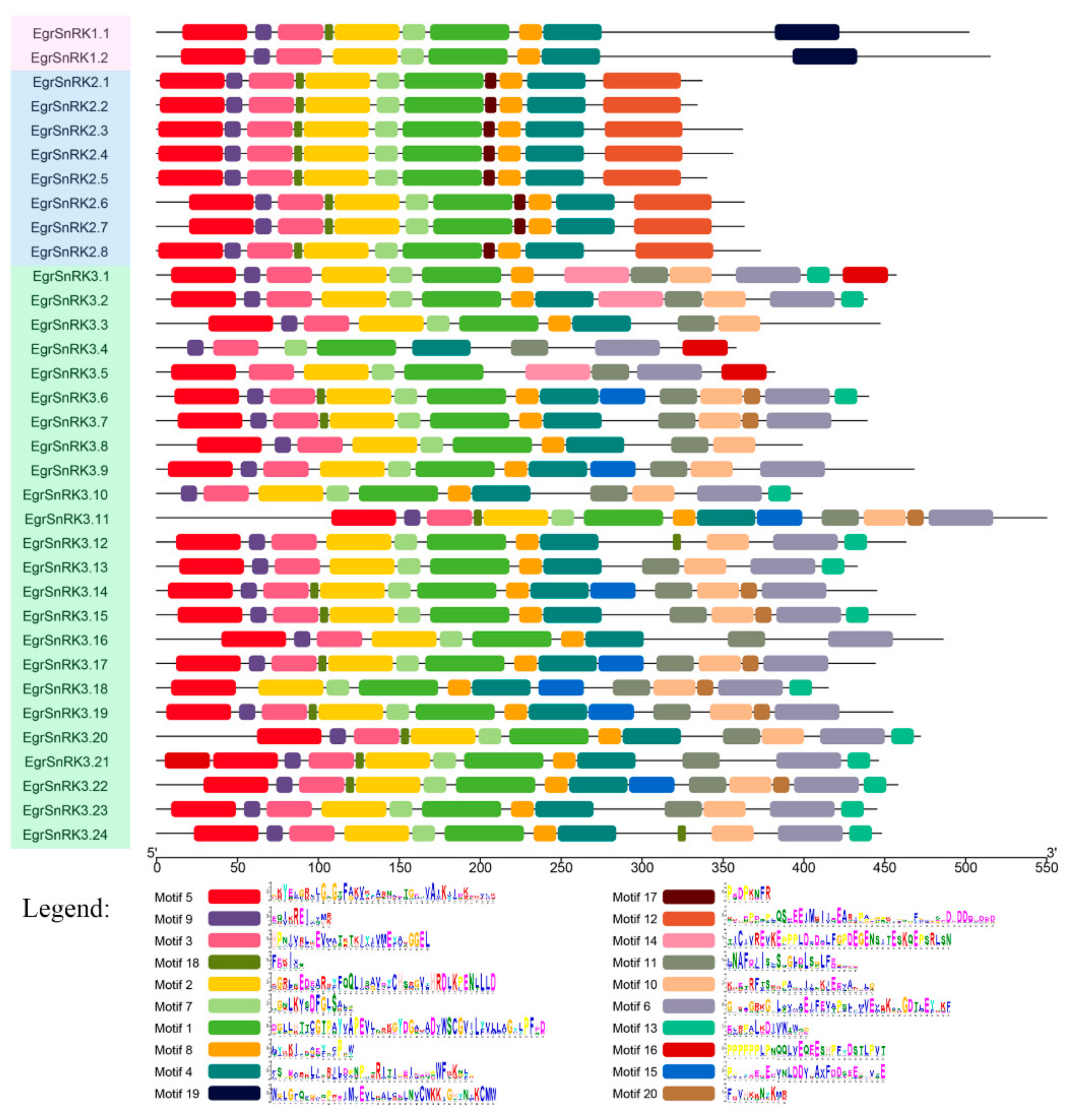
2.4. Motif Composition and Gene Structural Analysis of the SnRK Gene Family in E. grandis
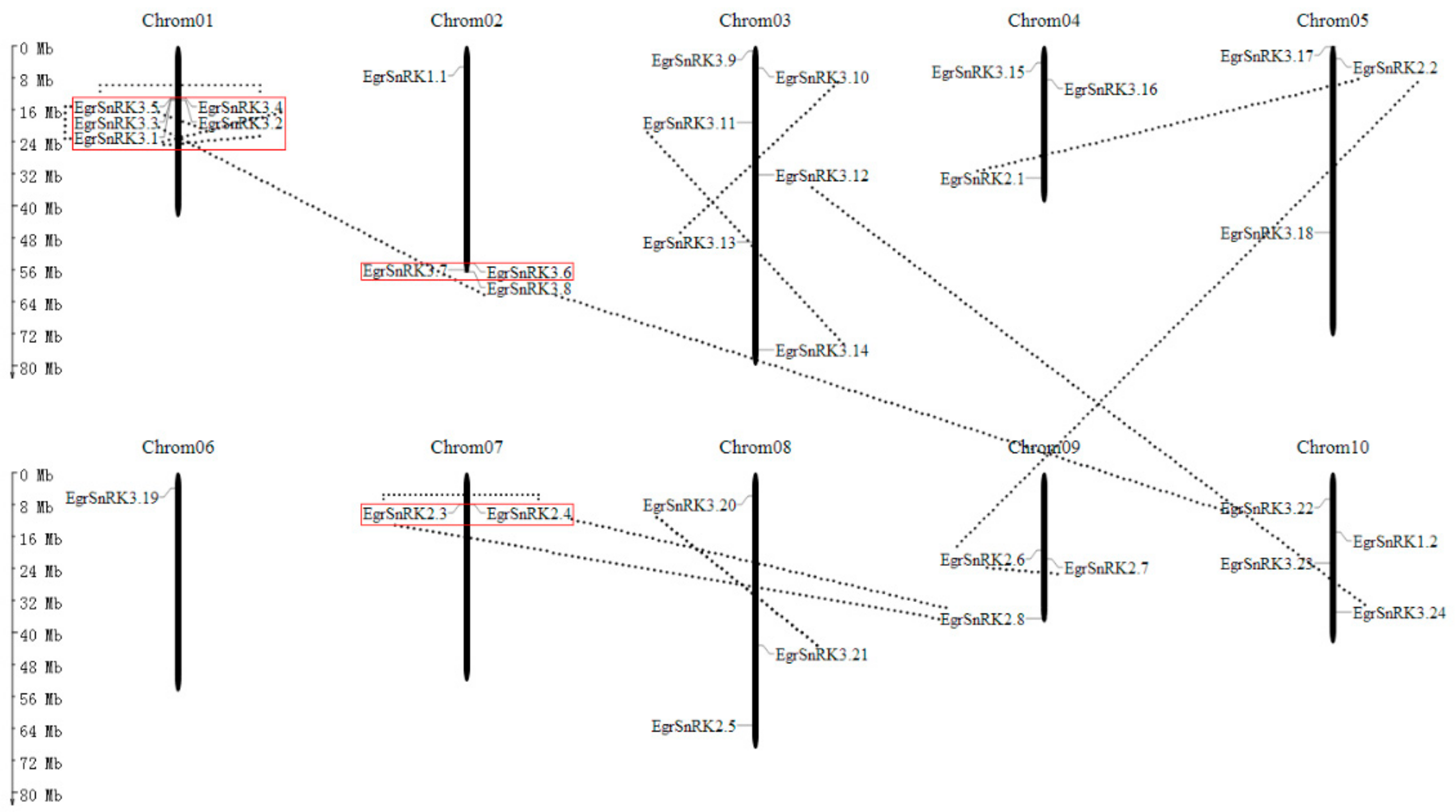
2.5. Chromosomal Location and Gene Pairs Analysis in E. grandis
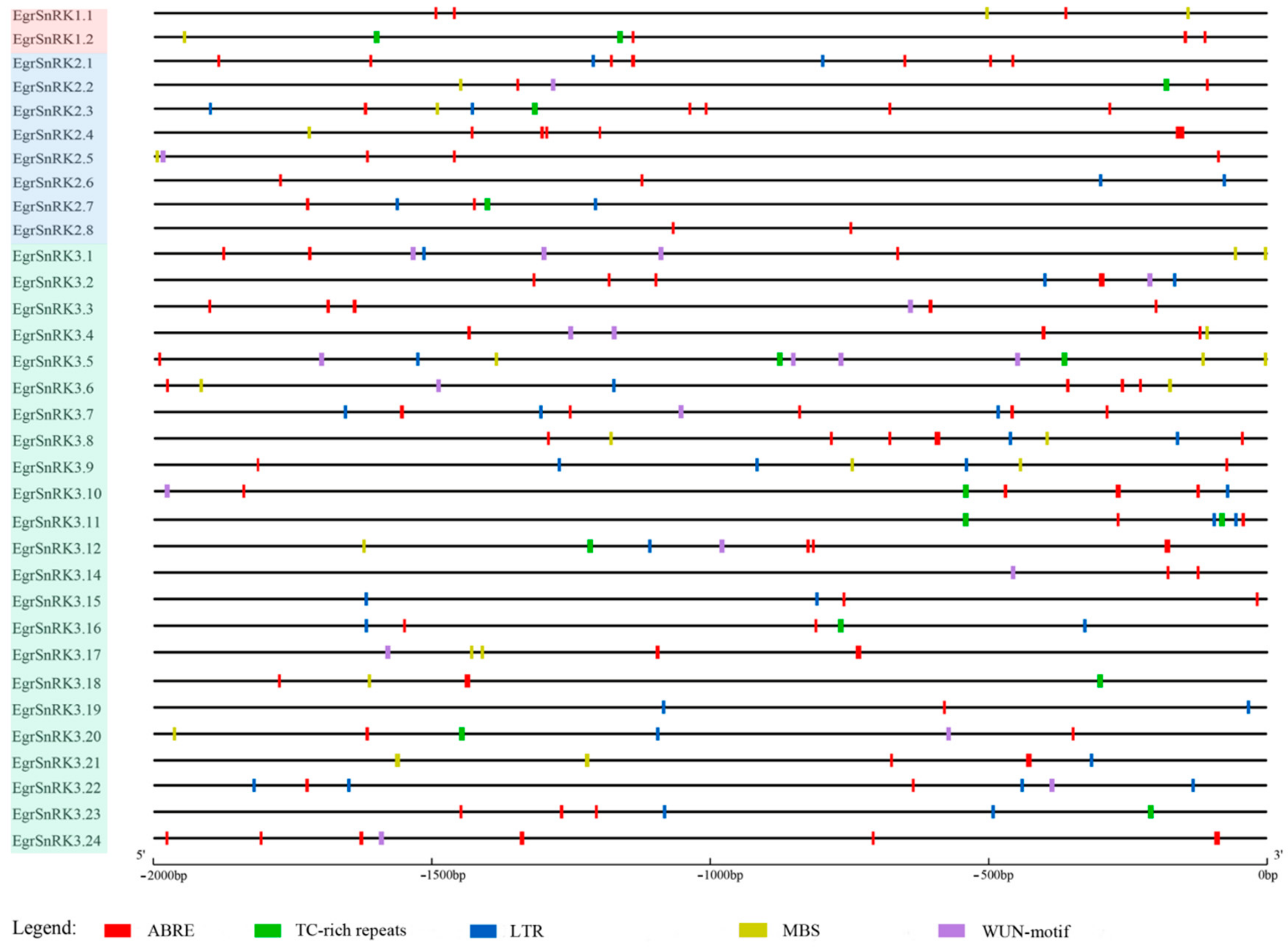
2.6. Promoter Analysis
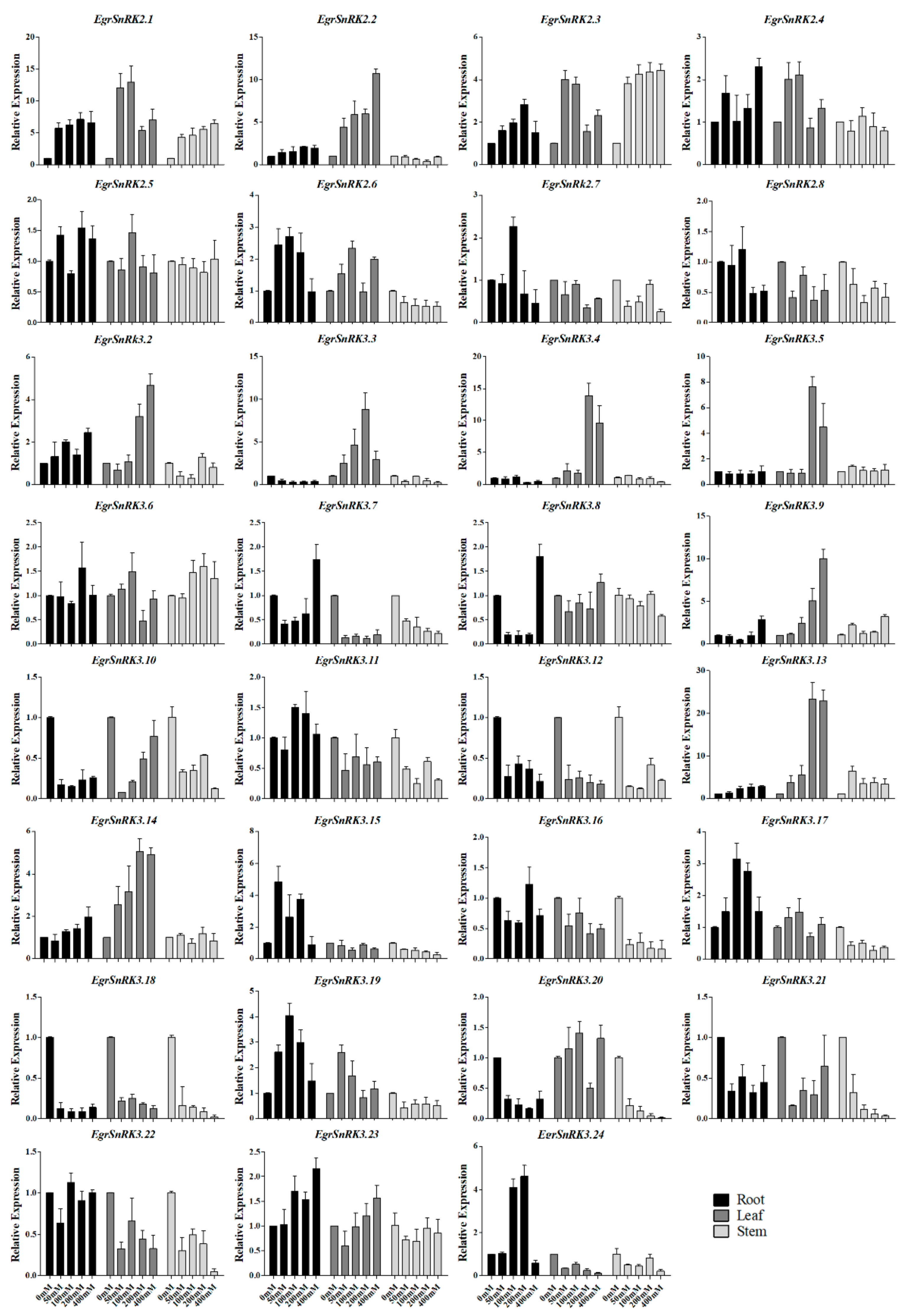
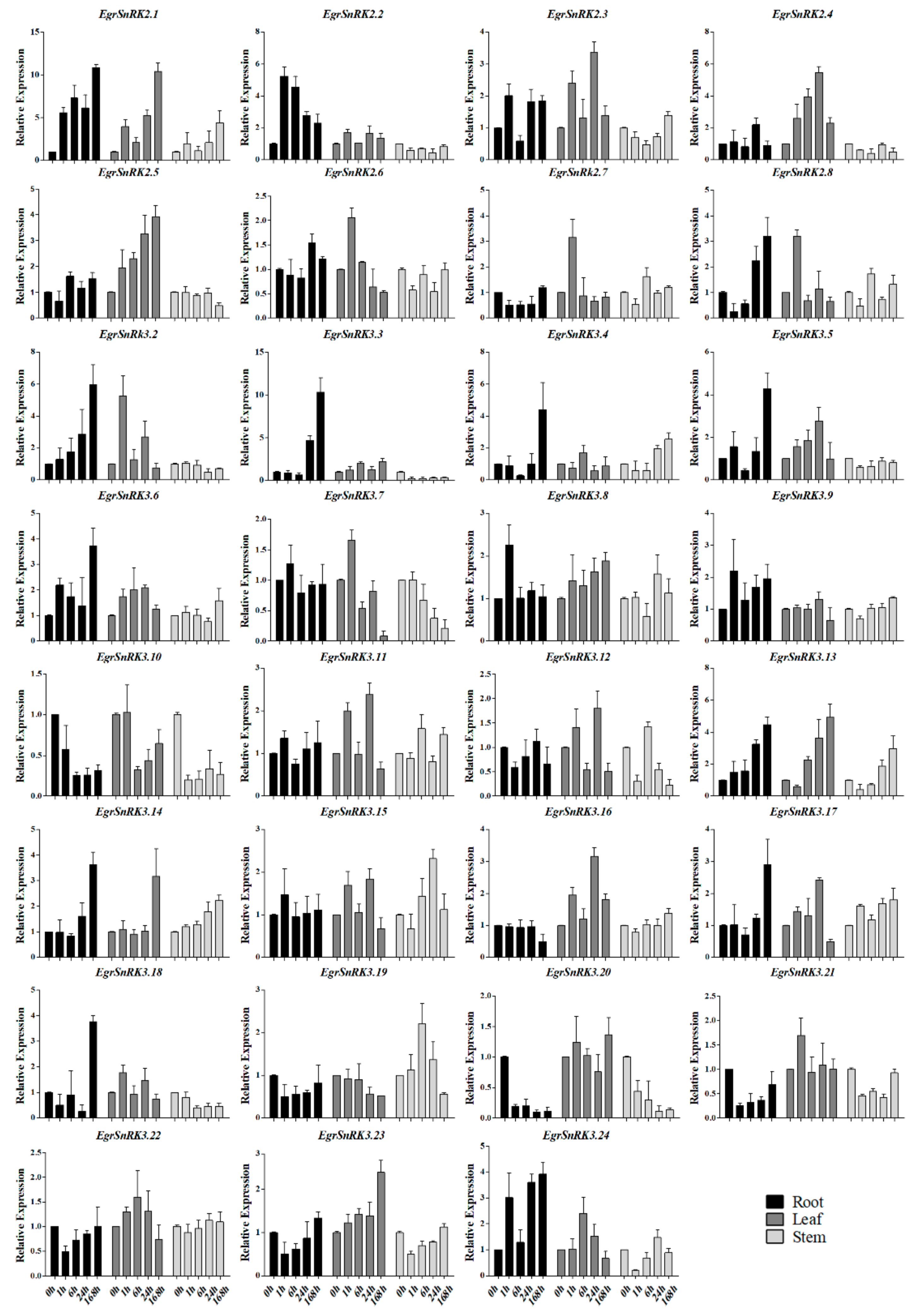
2.7. EgrSnRK Genes Expression under NaCl Treatment
3. Discussion
4. Materials and Methods
4.1. Sequence Retrieval and Gene Identification
4.2. Multiple Alignment and Phylogenetic Analysis
4.3. Identification of Conserved Motifs and Analysis of Gene Structure
4.4. Chromosomal Location
4.5. Ka and Ks Analysis of Homologous Pair
4.6. Cis-Elements in the Promoter Regions of EgrSnRK Genes
4.7. Plant Materials, Growth Conditions and Salt Treatments
4.8. RNA Extraction and qRT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Availability of Data and Materials
References
- Cohen, P. Review Lecture: Protein Phosphorylation and Hormone Action. Proc. R. Soc. Lond. 1988, 234, 115–144. [Google Scholar] [PubMed]
- Hunter, T.J.C. Protein kinases and phosphatases: The yin and yang of protein phosphorylation and signaling. Trend Biochem Sci. 1995, 80, 225. [Google Scholar] [CrossRef]
- Tör, M.; Lotze, M.T.; Holton, N. Receptor-mediated signalling in plants: Molecular patterns and programmes. J. Exp. Bot. 2009, 60, 3645–3654. [Google Scholar] [CrossRef] [PubMed]
- Wrzaczek, M.; Hirt, H.J.B. Plant MAP kinase pathways: How many and what for? Biol. Cell 2012, 93, 81–87. [Google Scholar] [CrossRef]
- Ludwig, A.A.; Romeis, T.; Jones, J.D. CDPK-mediated signalling pathways: Specificity and cross-talk. J. Exp. Bot. 2004, 55, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Hrabak, E.M.; Chan, C.W.M.; Michael, G.; Harper, J.F.; Choi, J.H.; Nigel, H.; Jorg, K.; Sheng, L.; Nimmo, H.G.; Sussman, M.R.; et al. The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol. 2003, 132, 666–680. [Google Scholar] [CrossRef] [PubMed]
- Halford, N.G.; Hardie, D.G. SNF1-related protein kinases: Global regulators of carbon metabolism in plants? Plant Mol. Biol. 1998, 37, 735–748. [Google Scholar] [CrossRef]
- Celenza, J.L.; Carlson, M. A yeast gene that is essential for release from glucose repression encodes a protein kinase. Science 1986, 233, 1175–1180. [Google Scholar] [CrossRef]
- Kulik, A.; Wawer, I.; Krzywińska, E.; Bucholc, M.; Dobrowolska, G. SnRK2 protein kinases--key regulators of plant response to abiotic stresses. Omics 2011, 15, 859. [Google Scholar] [CrossRef]
- Albrecht, V.; Ritz, O.; Linder, S.; Harter, K.; Kudla, J. The NAF domain defines a novel protein-protein interaction module conserved in Ca2+-regulated kinases. Embo J. 2014, 20, 1051–1063. [Google Scholar] [CrossRef]
- Masaru, O.; Yan, G.; Ursula, H.; Jian-Kang, Z. A novel domain in the protein kinase SOS2 mediates interaction with the protein phosphatase 2C ABI2. Proc. Natl. Acad. Sci. USA 2003, 100, 11771–11776. [Google Scholar]
- Baena-González, E.; Rolland, F.; Thevelein, J.M.; Sheen, J. A central integrator of transcription networks in plant stress and energy signalling. Nature 2007, 448, 938. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Shewry, P.H.; Barcelo, P.; Lazzeri, P.A.; Halford, N. Expression of antisense SnRK1 protein kinase sequence causes abnormalpollen development and male sterility in transgenic barley. Plant J. 2001, 28, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Ruslana, R.; Volodymyr, R.; Winfriede, W.; Ljudmilla, B.; Hans, W. Repressing the expression of the SUCROSE NONFERMENTING-1-RELATED PROTEIN KINASE gene in pea embryo causes pleiotropic defects of maturation similar to an abscisic acid-insensitive phenotype. Plant Physiol. 2006, 140, 263–278. [Google Scholar]
- Halford, N.G.; Hey, S.J. Snf1-related protein kinases (SnRKs) act within an intricate network that links metabolic and stress signalling in plants. Biochem. J. 2009, 419, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Marie, B.; Hélène, B.B.; Christiane, L.J. Identification of nine sucrose nonfermenting 1-related protein kinases 2 activated by hyperosmotic and saline stresses in Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 41758–41766. [Google Scholar]
- Anna-Chiara, M.; Sylvain, M.; Alain, V.; Francesca, F.; Giraudat, J. Arabidopsis OST1 protein kinase mediates the regulation of stomatal aperture by abscisic acid and acts upstream of reactive oxygen species production. Plant Cell 2002, 14, 3089–3099. [Google Scholar]
- Riichiro, Y.; Taishi, U.; Tsuyoshi, M.; Seiji, T.; Fuminori, T.; Kazuo, S. The regulatory domain of SRK2E/OST1/SnRK2.6 interacts with ABI1 and integrates abscisic acid (ABA) and osmotic stress signals controlling stomatal closure in Arabidopsis. J. Biol. Chem. 2006, 281, 5310–5318. [Google Scholar]
- Umezawa, T.; Yoshida, R.; Maruyama, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. SRK2C, a SNF1-related protein kinase 2, improves drought tolerance by controlling stress-responsive gene expression in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2004, 101, 17306–17311. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Yamamoto, S.; Minami, H.; Kagaya, Y.; Hattori, T. Differential activation of the rice sucrose nonfermenting1-related protein kinase2 family by hyperosmotic stress and abscisic acid. Plant Cell 2004, 16, 1163–1177. [Google Scholar] [CrossRef]
- Huai, J.; Wang, M.; He, J.; Zheng, J.; Dong, Z.; Lv, H.; Zhao, J.; Wang, G. Cloning and characterization of the SnRK2 gene family from Zea mays. Plant Cell Rep. 2008, 27, 1861–1868. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Cheng, Y.H.; Zhang, C.; Shen, X.J.; You, Q.B.; Guo, W.; Li, X.; Song, X.J.; Zhou, X.A.; Jiao, Y.Q. Genome-Wide Identification and Characterization of theGmSnRK2Family in Soybean. Int. J. Mol. Sci. 2017, 18, 1834. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ge, X.; Yang, Z.; Zhang, C.; Zhao, G.; Chen, E.; Liu, J.; Zhang, X.; Li, F. Genome-wide identification and characterization of SnRK2 gene family in cotton (Gossypium hirsutum L.). BMC Genet. 2017, 18, 54. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, W.; Sun, J.; Liang, X.; Yang, X.; Wei, S.; Wang, X.; Zhou, Y.; Xiao, Q.; Yang, G. Genome-wide analysis of SnRK gene family in Brachypodium distachyon and functional characterization of BdSnRK2.9. Plant Sci. 2015, 237, 33–45. [Google Scholar] [CrossRef]
- Guo, D.; Li, H.L.; Zhu, J.H.; Wang, Y.; An, F.; Xie, G.S.; Peng, S. Genomes, Genome-wide identification, characterization, and expression analysis of SnRK2 family in Hevea brasiliensis. Tree Genet. Genome 2017, 13, 86. [Google Scholar] [CrossRef]
- Diédhiou, C.J.; Popova, O.V.; Dietz, K.J.; Golldack, D. The SNF1-type serine-threonine protein kinase SAPK4regulates stress-responsive gene expression in rice. BMC Plant Biol. 2008, 8, 49. [Google Scholar] [CrossRef]
- Hongying, Z.; Hongfang, J.; Guoshun, L.; Shengnan, Y.; Songtao, Z.; Yongxia, Y.; Peipei, Y.; Hong, C. Cloning and characterization of SnRK2 subfamily II genes from Nicotiana tabacum. Mol. Biol. Res. 2014, 41, 5701. [Google Scholar]
- Song, X.; Yu, X.; Hori, C.; Demura, T.; Ohtani, M.; Zhuge, Q. Heterologous overexpression of poplar SnRK2 genes enhanced salt stress tolerance in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 612. [Google Scholar] [CrossRef]
- Zhang, H.; Mao, X.; Jing, R.; Chang, X.; Xie, H. Characterization of a common wheat (Triticum aestivum L.) TaSnRK2. 7 gene involved in abiotic stress responses. J. Exp. Bot. 2010, 62, 975–988. [Google Scholar] [CrossRef]
- Feng, J.; Wang, L.; Wu, Y.; Luo, Q.; Zhang, Y.; Qiu, D.; Han, J.; Su, P.; Xiong, Z.; Chang, J.; et al. TaSnRK2. 9, a sucrose non-fermenting 1-related protein kinase gene, positively regulates plant response to drought and salt stress in transgenic tobacco. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- He, L.; Yang, X.; Wang, L.; Zhu, L.; Zhou, T.; Deng, J.; Zhang, X. Molecular cloning and functional characterization of a novel cotton CBL-interacting protein kinase gene (GhCIPK6) reveals its involvement in multiple abiotic stress tolerance in transgenic plants. Biochem. Bioph. Res. 2013, 435, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Chen, X.-J.; Wang, J.-H.; Zheng, J. Overexpression of a maize SNF-related protein kinase gene, ZmSnRK2. 11, reduces salt and drought tolerance in Arabidopsis. J. Integr. Agric.. 2015, 14, 1229–1241. [Google Scholar] [CrossRef]
- Piao, H.-L.; Xuan, Y.-H.; Park, S.H.; Je, B.I.; Park, S.J.; Park, S.H.; Kim, C.M.; Huang, J.; Wang, G.K.; Kim, M. OsCIPK31, a CBL-interacting protein kinase is involved in germination and seedling growth under abiotic stress conditions in rice plants. Mol. Cell 2010, 30, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, V.; Parasuraman, B.; Laxmi, A.; Chattopadhyay, D. CIPK6, a CBL-interacting protein kinase is required for development and salt tolerance in plants. Plant J. 2009, 58, 778–790. [Google Scholar] [CrossRef] [PubMed]
- Huertas, R.; Olias, R.; Eljakaoui, Z.; Gálvez, F.J.; Li, J.; de Morales, P.A.; Belver, A.; Rodríguez-Rosales, M. Overexpression of SlSOS2 (SlCIPK24) confers salt tolerance to transgenic tomato. Plant Cell Environ. 2012, 35, 1467–1482. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-N.; Cheong, Y.H.; Grant, J.J.; Pandey, G.K.; Luan, S. CIPK3, a calcium sensor–associated protein kinase that regulates abscisic acid and cold signal transduction in Arabidopsis. Plant Cell Online 2003, 15, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.K.; Kanwar, P.; Singh, A.; Steinhorst, L.; Pandey, A.; Yadav, A.K.; Tokas, I.; Sanyal, S.K.; Kim, B.-G.; Lee, S.-C. Calcineurin B-like protein-interacting protein kinase CIPK21 regulates osmotic and salt stress responses in Arabidopsis. Plant Physiol. 2015, 169, 780–792. [Google Scholar] [CrossRef]
- Halfter, U.; Ishitani, M.; Zhu, J.K. The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium-binding protein SOS3. Proc. Natl. Acad. Sci. USA 2000, 97, 3735–3740. [Google Scholar] [CrossRef]
- Guo, Y.; Xiong, L.; Song, C.P.; Gong, D.; Halfter, U.; Zhu, J.K. A Calcium Sensor and Its Interacting Protein Kinase Are Global Regulators of Abscisic Acid Signaling in Arabidopsis. Dev. Cell 2002, 3, 233–244. [Google Scholar] [CrossRef]
- Liu, J.; Ishitani, M.; Halfter, U.; Kim, C.S.; Zhu, J.K. The Arabidopsis thaliana SOS2 gene encodes a protein kinase that is required for salt tolerance. Proc. Natl. Acad. Sci. USA 2000, 97, 3730–3734. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, J.; Bi, Y.; Wang, L.; Tang, L.; Yu, X.; Ohtani, M.; Demura, T.; Zhuge, Q. Overexpression of PtSOS2 enhances salt tolerance in transgenic poplars. Plant Mol. Biol. Rep. 2014, 32, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Myburg, A.A.; Dario, G.; Tuskan, G.A.; Uffe, H.; Hayes, R.D.; Jane, G.; Jerry, J.; Erika, L.; Hope, T.; Diane, B. The genome of Eucalyptus grandis. Nature 2014, 510, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Qin, Y.; Zou, Y.; Ma, F. Genome-wide identification and expression profiling of the SnRK2 gene family in Malus prunifolia. Gene 2014, 552, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Chen, N.N.; Cheng, Z.M.; Xiong, J.S. Genome-wide identification, annotation and expression profile analysis of SnRK2 gene family in grapevine. Aust. J. Grape Wine R. 2016, 22, 478–488. [Google Scholar] [CrossRef]
- Ali, G.M.; Komatsu, S. Proteomic analysis of rice leaf sheath during drought stress. J. Proteome Res. 2006, 5, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Sarda, X.; Tousch, D.; Ferrare, K.; Legrand, E.; Dupuis, J.; Casse-Delbart, F.; Lamaze, T. Two TIP-like genes encoding aquaporins are expressed in sunflower guard cells. Plant J. 1997, 12, 1103–1111. [Google Scholar] [CrossRef]
- Chen, X.; Gu, Z.; Xin, D.; Hao, L.; Liu, C.; Huang, J.; Ma, B.; Zhang, H. Genomics, Identification and characterization of putative CIPK genes in maize. J. Gent. Genom. 2011, 38, 77–87. [Google Scholar] [CrossRef]
- Su, Y.; Wang, Y.; Zhen, J.; Zhang, X.; Chen, Z.; Li, L.; Huang, Y.; Hua, J. SnRK2 Homologs in Gossypium and GhSnRK2.6 Improved Salt Tolerance in Transgenic Upland Cotton and Arabidopsis. Plant Mol. Biol. Rep. 2017, 35, 1–15. [Google Scholar] [CrossRef]
- Mao, X.G.; Zhang, H.Y.; Tian, S.J.; Chang, X.P.; Jing, R.L. TaSnRK2.4, an SNF1-type serine/threonine protein kinase of wheat (Triticum aestivum L.), confers enhanced multistress tolerance in Arabidopsis. J. Exp. Bot. 2010, 61, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangradorvegas, A. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, (Database issue), Database–D285. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2010, 39 (Suppl. 1), D225–D229. [Google Scholar] [CrossRef]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2011, 40, D302–D305. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37 (Suppl. 2), W202–W208. [Google Scholar] [CrossRef]
- Blanc, G.; Wolfe, K.H. Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes. Plant Cell 2004, 16, 1667–1678. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Rozas, J. DNA sequence polymorphism analysis using DnaSP. In Bioinformatics for DNA Sequence Analysis; Springer: Berlin/Heidelberg, Germany, 2009; pp. 337–350. [Google Scholar]
- Liu, Q.; Wang, H.; Zhang, Z.; Wu, J.; Feng, Y.; Zhu, Z. Divergence in function and expression of the NOD26-like intrinsic proteins in plants. BMC Genom. 2009, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Guo, G.; Yan, H.; Qiu, Z.; Liu, Q.; Zeng, B. Characterization of Brassinazole resistant (BZR) gene family and stress induced expression in Eucalyptus grandis. Physiol. Mol. Biol. Plants 2018, 24, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C T method. Nat. Protoc. 2008, 3, 1101. [Google Scholar] [CrossRef] [PubMed]
- Bryfczynski, S. GraphPad: A CS2/CS7 tool for graph creation. In Proceedings of the 47th Annual Southeast Regional Conference; ACM: New York, NY, USA, 2009; p. 53. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene Identifier | Chr | Location Coordinates (5’–3’) | ORF Length (bp) | Protein | |||
|---|---|---|---|---|---|---|---|---|
| Length (a.a.) | PI | Mol.Wt. (kDa) | Exons | |||||
| EgrSnRK1.1 | Eucgr.B00544.1 | 2 | 5601699–5607791 | 1509 | 502 | 8.6 | 57.22 | 10 |
| EgrSnRK1.2 | Eucgr.J01364.3 | 10 | 15761839–15770736 | 1548 | 515 | 8.51 | 59.06 | 10 |
| EgrSnRK2.1 | Eucgr.D02135.1 | 4 | 34783964–34788536 | 1014 | 337 | 6.18 | 38.09 | 9 |
| EgrSnRK2.2 | Eucgr.E00345.1 | 5 | 3283351–3287110 | 1005 | 334 | 5.63 | 38.07 | 9 |
| EgrSnRK2.3 | Eucgr.G00557.1 | 7 | 8288459–8292881 | 1089 | 362 | 5.31 | 41.25 | 9 |
| EgrSnRK2.4 | Eucgr.G00558.1 | 7 | 8300485–8309815 | 1071 | 356 | 6 | 40.66 | 9 |
| EgrSnRK2.5 | Eucgr.H04745.1 | 8 | 66440405–66443914 | 1023 | 340 | 5.36 | 38.47 | 9 |
| EgrSnRK2.6 | Eucgr.I00977.1 | 9 | 20388568–20394557 | 1092 | 363 | 4.73 | 41.19 | 9 |
| EgrSnRK2.7 | Eucgr.I01180.1 | 9 | 22740915–22744636 | 1092 | 363 | 4.87 | 41.44 | 9 |
| EgrSnRK2.8 | Eucgr.I02742.1 | 9 | 38306416–38310220 | 1122 | 373 | 6.18 | 42.90 | 9 |
| EgrSnRK3.1 | Eucgr.A00690.1 | 1 | 14305972–14307407 | 1374 | 457 | 8.51 | 51.41 | 2 |
| EgrSnRK3.2 | Eucgr.A00691.1 | 1 | 14298350–14300413 | 1320 | 439 | 9.06 | 49.90 | 1 |
| EgrSnRK3.3 | Eucgr.A00711.1 | 1 | 14006453–14008352 | 1344 | 447 | 5.52 | 49.76 | 1 |
| EgrSnRK3.4 | Eucgr.A00734.1 | 1 | 13706777–13708090 | 1077 | 358 | 9.52 | 40.60 | 6 |
| EgrSnRK3.5 | Eucgr.A00737.1 | 1 | 13619140–13620568 | 1149 | 382 | 5.92 | 43.19 | 5 |
| EgrSnRK3.6 | Eucgr.B03773.1 | 2 | 57202252–57207754 | 1323 | 440 | 6.66 | 49.91 | 14 |
| EgrSnRK3.7 | Eucgr.B03958.1 | 2 | 5872963–58735460 | 1320 | 439 | 8.15 | 49.29 | 12 |
| EgrSnRK3.8 | Eucgr.B04021.1 | 2 | 59343262–59345126 | 1200 | 399 | 7 | 45.36 | 1 |
| EgrSnRK3.9 | Eucgr.C00193.1 | 3 | 1473208–1476720 | 1407 | 468 | 6 | 52.51 | 15 |
| EgrSnRK3.10 | Eucgr.C00357.1 | 3 | 5833911–5835857 | 1200 | 399 | 8.81 | 44.92 | 1 |
| EgrSnRK3.11 | Eucgr.C01333.1 | 3 | 20087289–20093442 | 1656 | 550 | 9.17 | 61.62 | 15 |
| EgrSnRK3.12 | Eucgr.C01944.1 | 3 | 33758197–33760724 | 1392 | 463 | 7.97 | 52.37 | 1 |
| EgrSnRK3.13 | Eucgr.C02590.1 | 3 | 51530920–51532898 | 1302 | 433 | 9.05 | 48.55 | 1 |
| EgrSnRK3.14 | Eucgr.C04226.1 | 3 | 79695224–79704623 | 1338 | 445 | 9.1 | 50.09 | 14 |
| EgrSnRK3.15 | Eucgr.D00272.1 | 4 | 4577009–4583478 | 1410 | 469 | 8.94 | 52.69 | 14 |
| EgrSnRK3.16 | Eucgr.D00481.1 | 4 | 8897498–8900910 | 1461 | 486 | 8.46 | 53.90 | 1 |
| EgrSnRK3.17 | Eucgr.E00017.1 | 5 | 222650–235591 | 1335 | 444 | 8.65 | 50.85 | 13 |
| EgrSnRK3.18 | Eucgr.E02758.1 | 5 | 48871914–48873537 | 1248 | 415 | 8.68 | 46.78 | 2 |
| EgrSnRK3.19 | Eucgr.F00453.1 | 6 | 4226550–4232934 | 1368 | 455 | 6.48 | 51.46 | 14 |
| EgrSnRK3.20 | Eucgr.H00223.1 | 8 | 6265751–6267413 | 1419 | 472 | 9.26 | 50.70 | 1 |
| EgrSnRK3.21 | Eucgr.H03182.1 | 8 | 45402293–45404868 | 1341 | 446 | 8.59 | 49.24 | 2 |
| EgrSnRK3.22 | Eucgr.J00641.1 | 10 | 7028075–7034476 | 1377 | 458 | 6.5 | 52.77 | 14 |
| EgrSnRK3.23 | Eucgr.J01840.1 | 10 | 23702496–23704068 | 1338 | 445 | 9.39 | 50.84 | 1 |
| EgrSnRK3.24 | Eucgr.J03116.1 | 10 | 36664597–36666371 | 1347 | 448 | 8.72 | 50.34 | 1 |
| Paralogues | Ka (JC) | Ks (JC) | Ka/Ks | |
|---|---|---|---|---|
| Gene 1 | Gene 2 | |||
| EgrSnRK2.2 | EgrSnRK2.1 | 0.0992 | 1.3197 | 0.075 |
| EgrSnRK2.6 | 0.23388 | 1.64249 | 0.142 | |
| EgrSnRK2.3 | EgrSnRK2.4 | 0.03228 | 0.09785 | 0.33 |
| EgrSnRK2.8 | 0.17009 | 1.24632 | 0.136 | |
| EgrSnRK2.4 | EgrSnRK2.8 | 0.13842 | 1.32215 | 0.105 |
| EgrSnRK2.6 | EgrSnRK2.7 | 0.11743 | 2.19481 | 0.054 |
| EgrSnRK3.1 | EgrSnRK3.2 | 0.11413 | 0.14826 | 0.77 |
| EgrSnRK3.4 | 0.54087 | 0.52272 | 1.035 | |
| EgrSnRK3.5 | EgrSnRK3.1 | 0.08151 | 0.18191 | 0.448 |
| EgrSnRK3.2 | 0.06487 | 0.13926 | 0.466 | |
| EgrSnRK3.4 | 0.15223 | 0.15972 | 0.953 | |
| EgrSnRK3.3 | EgrSnRK3.8 | 0.0813 | 1.14857 | 0.071 |
| EgrSnRK3.6 | EgrSnRK3.22 | 0.35042 | 0.77327 | 0.453 |
| EgrSnRK3.10 | EgrSnRK3.13 | 0.10413 | 0.60606 | 0.172 |
| EgrSnRK3.11 | EgrSnRK3.14 | 0.13937 | 1.09741 | 0.127 |
| EgrSnRK3.12 | EgrSnRK3.24 | 0.19932 | 1.39897 | 0.142 |
| Gene Name | ABRE | TC-Rich Repeats | LTR | MBS | WUN-Motif |
|---|---|---|---|---|---|
| EgrSnRK1.1 | 3 | 2 | |||
| EgrSnRK1.2 | 4 | 2 | 1 | ||
| EgrSnRK2.1 | 7 | 2 | |||
| EgrSnRK2.2 | 2 | 1 | 1 | 1 | |
| EgrSnRK2.3 | 5 | 1 | 2 | 1 | |
| EgrSnRK2.4 | 10 | 1 | |||
| EgrSnRK2.5 | 3 | 1 | 1 | ||
| EgrSnRK2.6 | 2 | 2 | |||
| EgrSnRK2.7 | 3 | 1 | 2 | ||
| EgrSnRK2.8 | 2 | ||||
| EgrSnRK3.1 | 3 | 1 | 2 | 3 | |
| EgrSnRK3.2 | 5 | 2 | 1 | ||
| EgrSnRK3.3 | 6 | 1 | |||
| EgrSnRK3.4 | 4 | 1 | 3 | ||
| EgrSnRK3.5 | 1 | 2 | 1 | 3 | 4 |
| EgrSnRK3.6 | 6 | 1 | 2 | 1 | |
| EgrSnRK3.7 | 6 | 3 | 1 | ||
| EgrSnRK3.8 | 5 | 2 | 2 | ||
| EgrSnRK3.9 | 2 | 3 | 2 | ||
| EgrSnRK3.10 | 6 | 1 | 1 | 1 | |
| EgrSnRK3.11 | 3 | 2 | 2 | ||
| EgrSnRK3.12 | 5 | 1 | 1 | 1 | 1 |
| EgrSnRK3.13 | |||||
| EgrSnRK3.14 | 2 | 1 | |||
| EgrSnRK3.15 | 2 | 2 | |||
| EgrSnRK3.16 | 2 | 1 | 2 | ||
| EgrSnRK3.17 | 2 | 2 | 1 | ||
| EgrSnRK3.18 | 2 | 1 | 1 | ||
| EgrSnRK3.19 | 1 | 2 | |||
| EgrSnRK3.20 | 3 | 1 | 1 | 1 | 1 |
| EgrSnRK3.21 | 3 | 1 | 2 | 2 | |
| EgrSnRK3.22 | 3 | 5 | 2 | ||
| EgrSnRK3.23 | 4 | 1 | 2 | ||
| EgrSnRK3.24 | 7 | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Yan, H.; Qiu, Z.; Hu, B.; Zeng, B.; Zhong, C.; Fan, C. Comprehensive Analysis of SnRK Gene Family and their Responses to Salt Stress in Eucalyptus grandis. Int. J. Mol. Sci. 2019, 20, 2786. https://doi.org/10.3390/ijms20112786
Wang Y, Yan H, Qiu Z, Hu B, Zeng B, Zhong C, Fan C. Comprehensive Analysis of SnRK Gene Family and their Responses to Salt Stress in Eucalyptus grandis. International Journal of Molecular Sciences. 2019; 20(11):2786. https://doi.org/10.3390/ijms20112786
Chicago/Turabian StyleWang, Yujiao, Huifang Yan, Zhenfei Qiu, Bing Hu, Bingshan Zeng, Chonglu Zhong, and Chunjie Fan. 2019. "Comprehensive Analysis of SnRK Gene Family and their Responses to Salt Stress in Eucalyptus grandis" International Journal of Molecular Sciences 20, no. 11: 2786. https://doi.org/10.3390/ijms20112786
APA StyleWang, Y., Yan, H., Qiu, Z., Hu, B., Zeng, B., Zhong, C., & Fan, C. (2019). Comprehensive Analysis of SnRK Gene Family and their Responses to Salt Stress in Eucalyptus grandis. International Journal of Molecular Sciences, 20(11), 2786. https://doi.org/10.3390/ijms20112786