Identification of Proteins Differentially Expressed by Adipose-derived Mesenchymal Stem Cells Isolated from Immunodeficient Mice

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
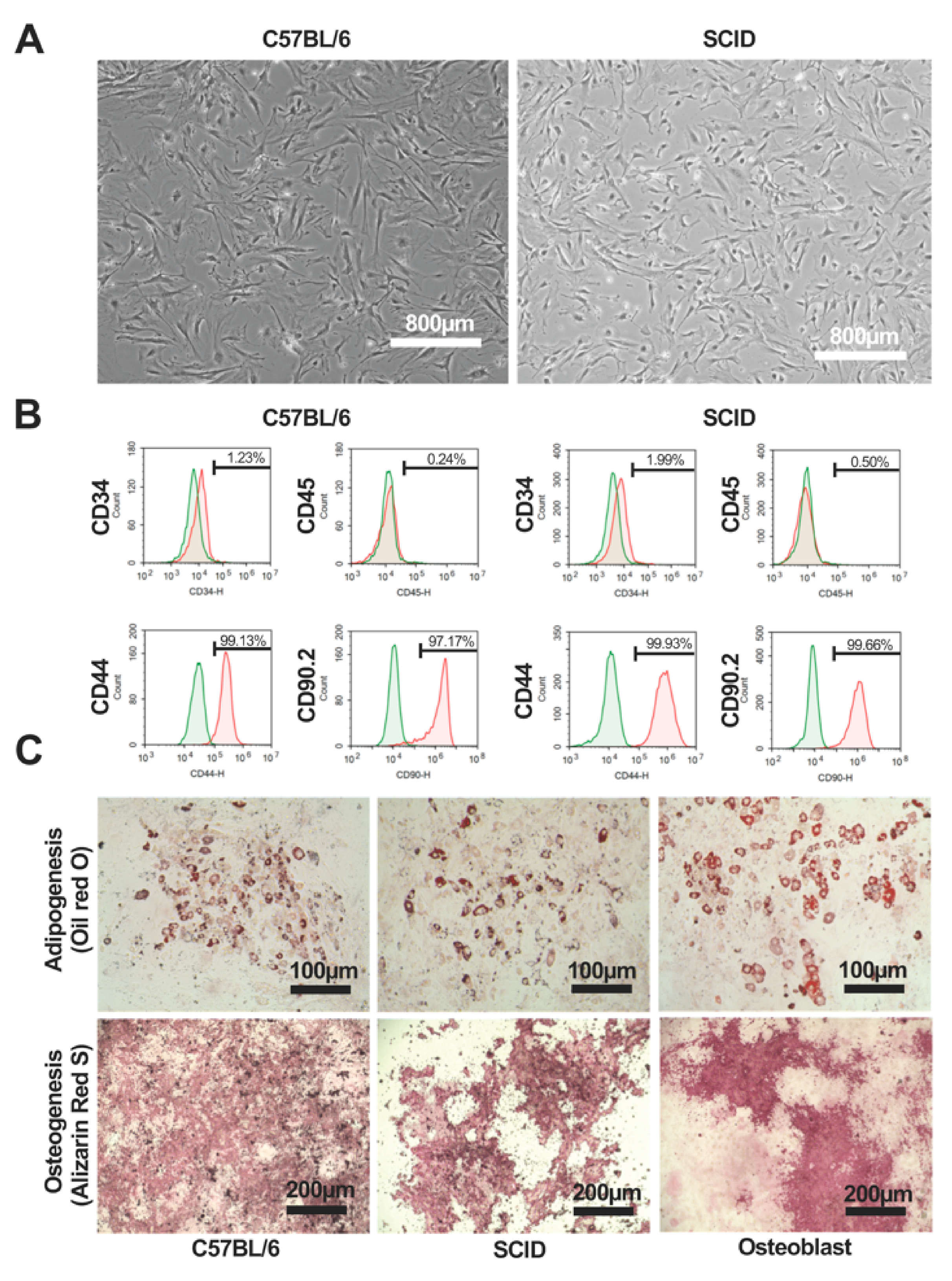
2.1. Characteristics and Quality of C57BL/6 and SCID mAdMSCs
2.2. Analysis of Protein Expression by C57BL/6 and SCID mAdMSCs
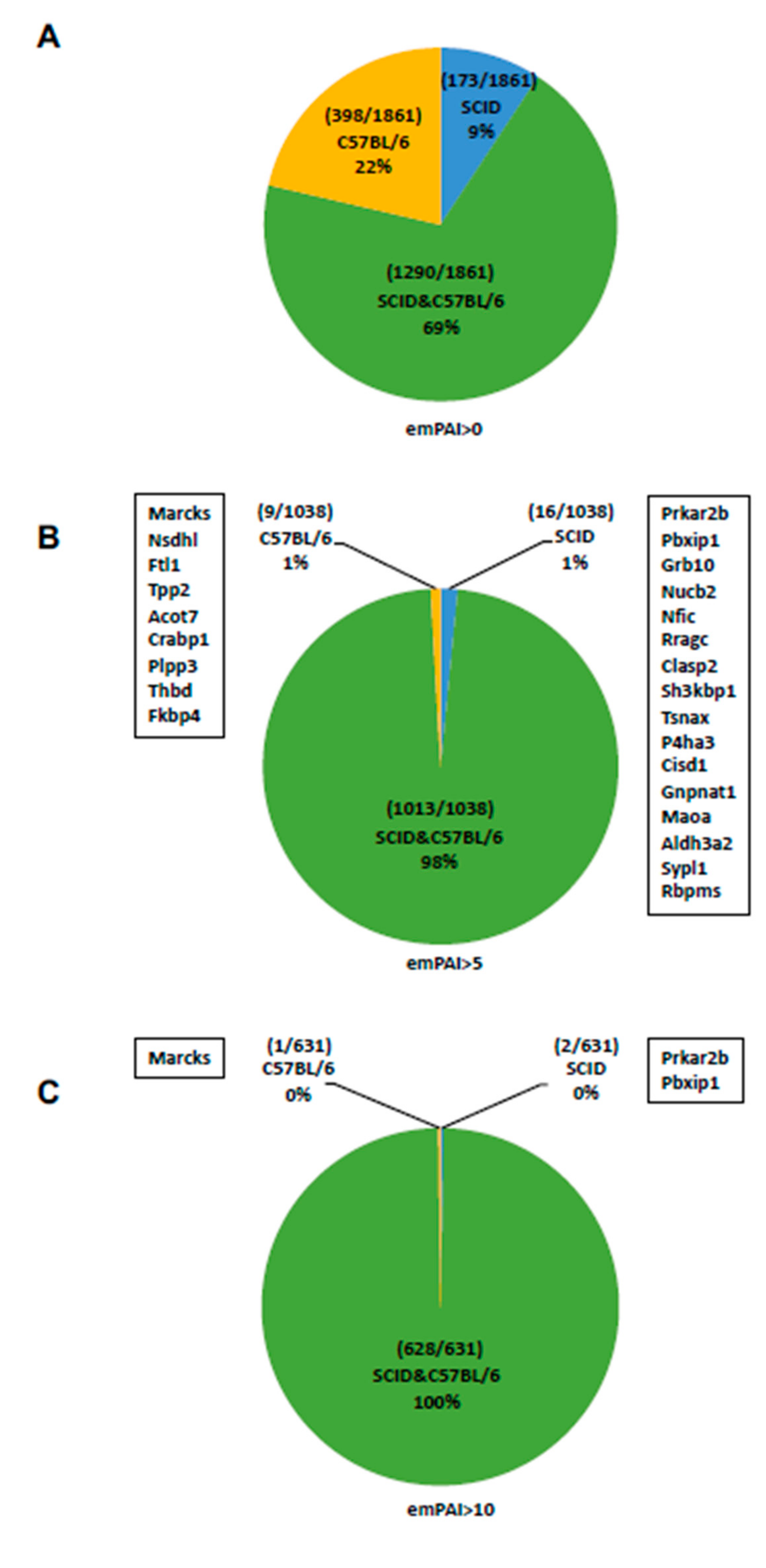
2.3. Analysis of Proteins Expressed by C57BL/6 mAdMSCs and SCID mAdMSCs (emPAI > 0)
2.4. Analysis of Proteins Expressed by C57BL/6 mAdMSCs and SCID mAdMSCs (emPAI > 5)
2.5. Analysis of Proteins Expressed by C57BL/6 mAdMSCs and SCID mAdMSCs (emPAI > 10)
2.6. Analysis of Proteins Expressed by C57BL/6 mAdMSCs and SCID mAdMSCs (emPAI > 5)
2.7. Analysis of Proteins Expressed by C57BL/6 mAdMSCs and SCID mAdMSCs (emPAI > 10)
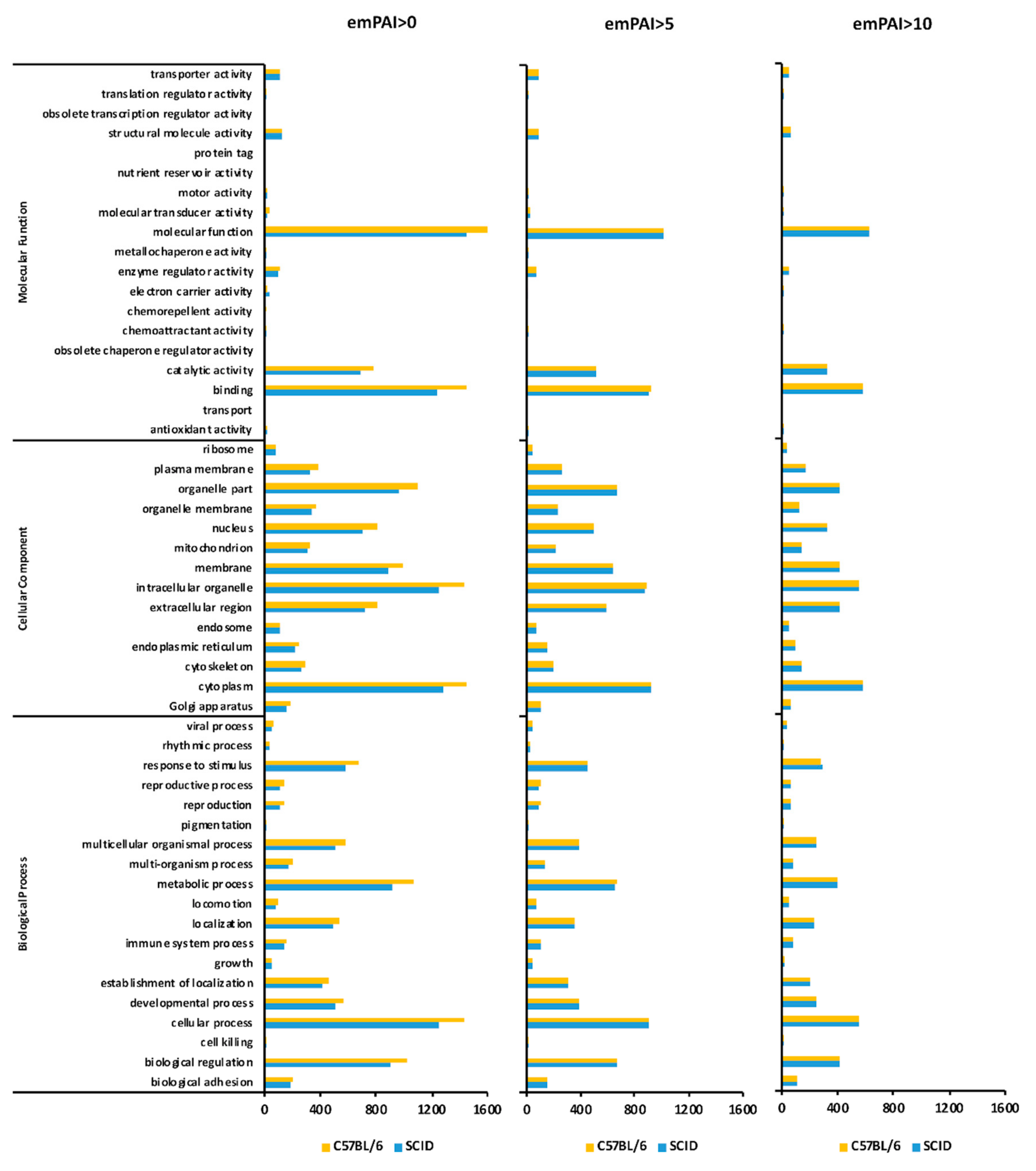
2.8. GO Classification (emPAI > 0)
2.9. GO Classification (emPAI > 5)
2.10. GO Classification (emPAI > 10 and emPAI > 0)
2.11. B Cell-Related Proteins Expressed by C57BL/6 mAdMSCs and SCID mAdMSCs
2.12. T Cell-Related Proteins Expressed by C57BL/6 mAdMSCs and SCID mAdMSCs
2.13. Viral Process-Related Proteins Expressed by C57BL/6 mAdMSCs and SCID mAdMSCs
2.14. Immune System Process-Related Proteins Expressed by C57BL/6 mAdMSCs and SCID mAdMSCs
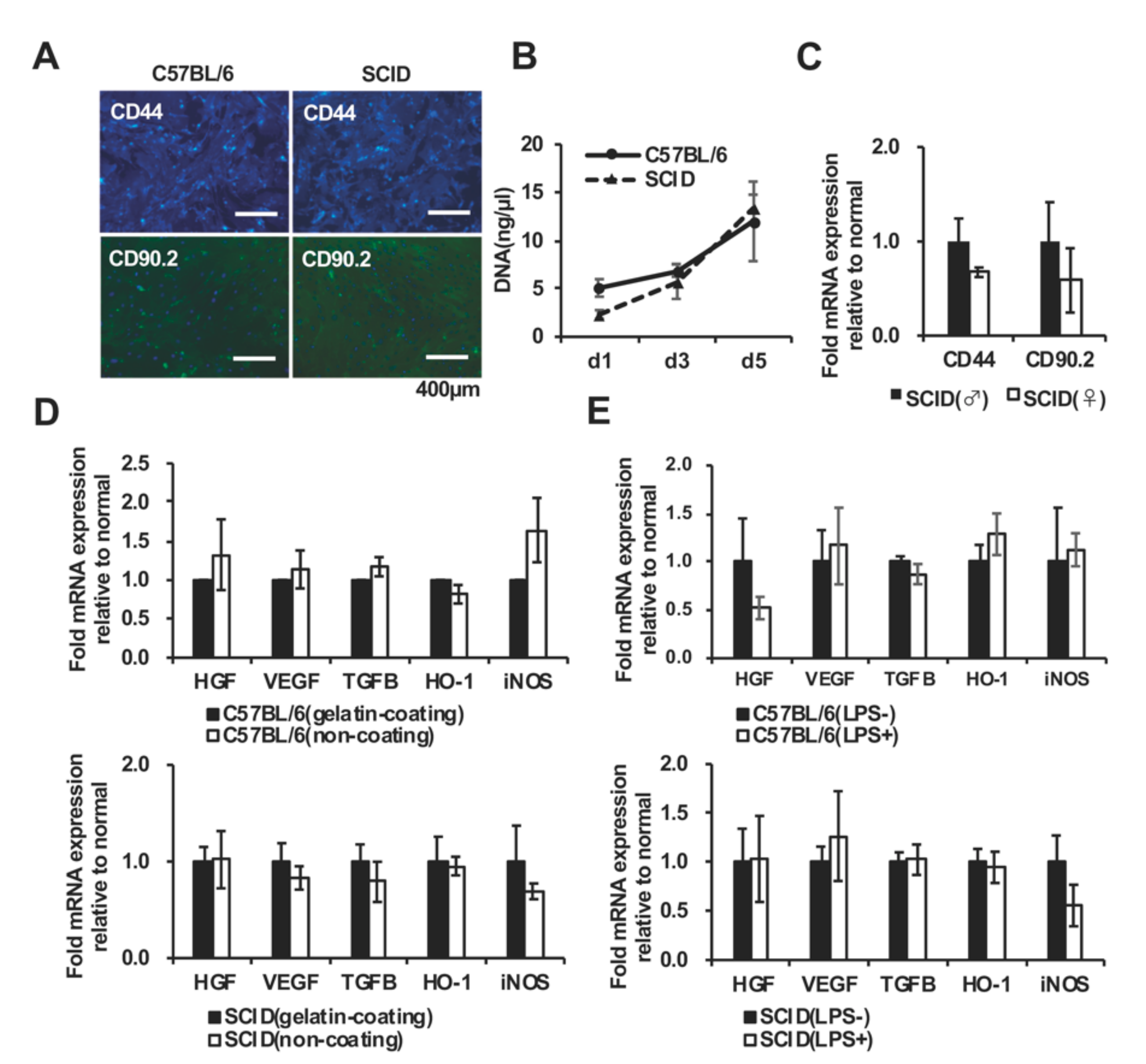
2.15. Effects of Culture Conditions on Protein Expression by AdMSCs
2.16. Quantitative Analysis of Proteins Expressed by C57BL/6 mAD MSCs and SCID mAdMSCs
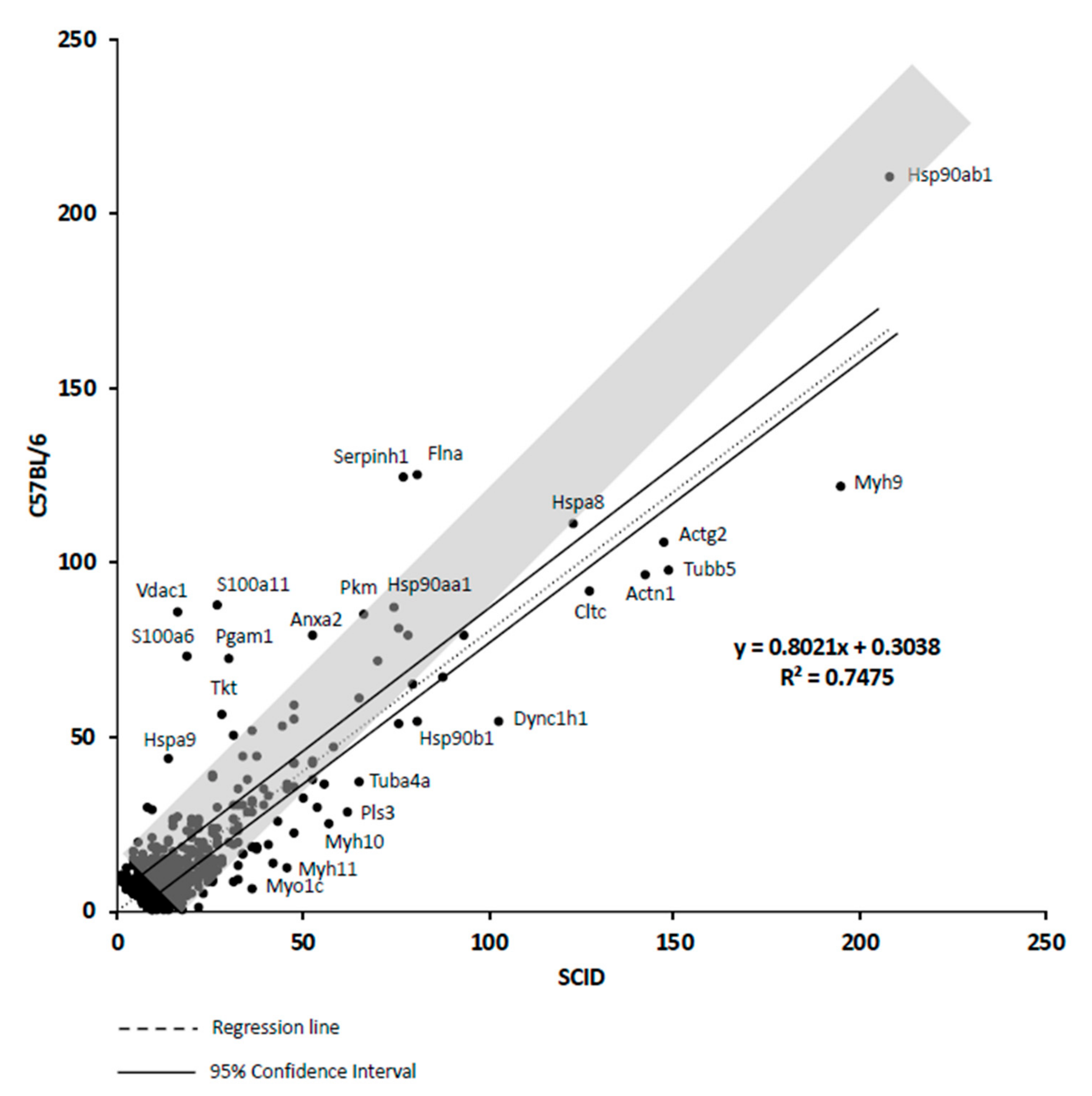
2.17. Correlation Analysis of Proteins Expressed by mAdMSCs
3. Discussion
3.1. Reliability of LC-MS/MS for Comparative Analysis of Protein Expression
3.2. T Cell/B Cell-Associated Proteins
3.3. Global Comparison of Proteins Expressed by mAdMSCs
3.4. Proteins Related to the Immune Response
3.5. Analysis of Proteins Associated with the Viral Response
3.6. Summary
4. Materials and Methods
4.1. Reagents and Materials
4.2. Animal Care
4.3. Isolation of AdMSCs from Mouse Adipose Tissue Via the Inguinal Pad Fat
4.4. Preparation of mAdMSCSs
4.5. Real-Time PCR (qPCR)
- mouse CD44 forward, TACTCATATTCTAGCCTCCCTCCTT,
- mouse CD44 reverse, GTGGAGAATAGCCAAGAATCATCTA,
- mouse CD90.2 forward, GTCCTTCAAATATCTCAGAACATGG,
- mouse CD90.2 reverse, GCCCTGGAATAAATACAGAGTACAA,
- mouse HGF forward, ACTCTTACCAAGGAAGACCCATTAC,
- mouse HGF reverse, ATACCAGTAGCATCGTTTTCTTGAC,
- mouse VEGF forward, TGTCTTCACTGGATATGTTTGACTG,
- mouse VEGF reverse, TTCTCTGTCATCATCTGTCTCTCTG,
- mouse TGF-β forward, AGTAGCTCCCCTATTTAAGAACACC,
- mouse TGF-β reverse, GGAAAGGTAGGTGATAGTCCTGAAT,
- mouse HO-1 forward, TCCAGACATTTCTGTCTCGTATTTC,
- mouse HO-1 reverse, CACACAGAAGTTAGAGACCAAGGTT,
- mouse iNOS forward, AGCTTCTGGCACTGAGTAAAGATAA,
- mouse iNOS reverse, GGGAGGAGAGGAGAGAGATTTAGTA,
- mouse GAPDH forward, GCCAAGTATGATGACATCAAGAAGG,
- and mouse GAPDH reverse, GTGCAGCGAACTTTATTGATGGTAT.
4.6. Flow Cytometry
4.7. Immunofluorescence Analysis
4.8. Cell Proliferation
4.9. Cell Differentiation
4.10. LC-MS/MS Analysis of Protein Expression
4.11. Data Analyses
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Wagers, A.J.; Weissman, I.L. Plasticity of adult stem cells. Cell 2004, 116, 639–648. [Google Scholar] [CrossRef]
- Wei, X.; Yang, X.; Han, Z.P.; Qu, F.F.; Shao, L.; Shi, Y.F. Mesenchymal stem cells: A new trend for cell therapy. Acta Pharmacol. Sin. 2013, 34, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Aoyama, T.; Fukiage, K.; Otsuka, S.; Furu, M.; Jin, Y.; Nasu, A.; Ueda, M.; Kasai, Y.; Ashihara, E.; et al. A novel method to isolate mesenchymal stem cells from bone marrow in a closed system using a device made by nonwoven fabric. Tissue Eng. Part C Methods 2010, 16, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Shohara, R.; Yamamoto, A.; Takikawa, S.; Iwase, A.; Hibi, H.; Kikkawa, F.; Ueda, M. Mesenchymal stromal cells of human umbilical cord Wharton’s jelly accelerate wound healing by paracrine mechanisms. Cytotherapy 2012, 14, 1171–1181. [Google Scholar] [CrossRef]
- Mak, J.; Jablonski, C.L.; Leonard, C.A.; Dunn, J.F.; Raharjo, E.; Matyas, J.R.; Biernaskie, J.; Krawetz, R.J. Intra-articular injection of synovial mesenchymal stem cells improves cartilage repair in a mouse injury model. Sci. Rep-Uk 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.B.; McIntosh, K.; Zvonic, S.; Garrett, S.; Floyd, Z.E.; Kloster, A.; di Halvorsen, Y.; Storms, R.W.; Goh, B.; Kilroy, G.; et al. Immunophenotype of human adipose-derived cells: Temporal changes in stromal-associated and stem cell-associated markers. Stem. Cells 2006, 24, 376–385. [Google Scholar] [CrossRef]
- Ben-Ami, E.; Berrih-Aknin, S.; Miller, A. Mesenchymal stem cells as an immunomodulatory therapeutic strategy for autoimmune diseases. Autoimmun. Rev. 2011, 10, 410–415. [Google Scholar] [CrossRef]
- Klinker, M.W.; Wei, C.H. Mesenchymal stem cells in the treatment of inflammatory and autoimmune diseases in experimental animal models. World J. Stem Cell. 2015, 7, 556–567. [Google Scholar] [CrossRef]
- Kay, A.G.; Long, G.; Tyler, G.; Stefan, A.; Broadfoot, S.J.; Piccinini, A.M.; Middleton, J.; Kehoe, O. Mesenchymal Stem Cell-Conditioned Medium Reduces Disease Severity and Immune Responses in Inflammatory Arthritis. Sci. Rep-Uk 2017, 7, 18019. [Google Scholar] [CrossRef]
- Tanaka, Y. Human mesenchymal stem cells as a tool for joint repair in rheumatoid arthritis. Clin. Exp. Rheumatol. 2015, 33, S58–S62. [Google Scholar]
- Seki, A.; Sakai, Y.; Komura, T.; Nasti, A.; Yoshida, K.; Higashimoto, M.; Honda, M.; Usui, S.; Takamura, M.; Takamura, T.; et al. Adipose tissue-derived stem cells as a regenerative therapy for a mouse steatohepatitis-induced cirrhosis model. Hepatology 2013, 58, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells - current trends and future prospective. Biosci. Rep. 2015, 35. [Google Scholar] [CrossRef]
- Miyagi-Shiohira, C.; Kobayashi, N.; Saitoh, I.; Watanabe, M.; Noguchi, Y.; Matsushita, M.; Noguchi, H. Evaluation of Serum-Free, Xeno-Free Cryopreservation Solutions for Human Adipose-Derived Mesenchymal Stem Cells. Cell Med. 2017, 9, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Nauta, A.J.; Fibbe, W.E. Immunomodulatory properties of mesenchymal stromal cells. Blood 2007, 110, 3499–3506. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.F.; Su, J.J.; Roberts, A.I.; Shou, P.S.; Rabson, A.B.; Ren, G.W. How mesenchymal stem cells interact with tissue immune responses. Trends Immunol. 2012, 33, 136–143. [Google Scholar] [CrossRef]
- Van Laar, J.M.; Tyndall, A. Adult stem cells in the treatment of autoimmune diseases. Rheumatology (Oxford) 2006, 45, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.; Hayashi, R.; Lafond-Walker, A.; Lowenstein, C.; Pardoll, D.; Levitsky, H. The central role of CD4(+) T cells in the antitumor immune response. J. Exp. Med. 1998, 188, 2357–2368. [Google Scholar] [CrossRef] [PubMed]
- LeBien, T.W.; Tedder, T.F. B lymphocytes: How they develop and function. Blood 2008, 112, 1570–1580. [Google Scholar] [CrossRef]
- Jacob, J.; Kelsoe, G.; Rajewsky, K.; Weiss, U. Intraclonal generation of antibody mutants in germinal centres. Nature 1991, 354, 389–392. [Google Scholar] [CrossRef]
- Jelicic, K.; Cimbro, R.; Nawaz, F.; Huang, D.W.; Zheng, X.; Yang, J.; Lempicki, R.A.; Pascuccio, M.; Van Ryk, D.; Schwing, C.; et al. The HIV-1 envelope protein gp120 impairs B cell proliferation by inducing TGF-beta 1 production and FcRL4 expression. Nat. Immunol. 2013, 14, 1256. [Google Scholar] [CrossRef] [PubMed]
- Stoecklein, V.M.; Osuka, A.; Ishikawa, S.; Lederer, M.R.; Wanke-Jellinek, L.; Lederer, J.A. Radiation exposure induces inflammasome pathway activation in immune cells. J. Immunol. 2015, 194, 1178–1189. [Google Scholar] [CrossRef] [PubMed]
- Bosma, G.C.; Custer, R.P.; Bosma, M.J. A severe combined immunodeficiency mutation in the mouse. Nature 1983, 301, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Denton, P.W.; Garcia, J.V. Humanized mouse models of HIV infection. AIDS Rev. 2011, 13, 135–148. [Google Scholar] [PubMed]
- Kyoizumi, S.; Suzuki, T.; Teraoka, S.; Seyama, T. Radiation sensitivity of human hair follicles in SCID-hu mice. Radiat Res. 1998, 149, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Huntley, R.P.; Sawford, T.; Martin, M.J.; O’Donovan, C. Understanding how and why the Gene Ontology and its annotations evolve: The GO within UniProt. Gigascience 2014, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, Y.; Nahar, S.; Miyagi-Shiohira, C.; Kinjo, T.; Kobayashi, N.; Saitoh, I.; Watanabe, M.; Fujita, J.; Noguchi, H. A Liquid Chromatography with Tandem Mass Spectrometry-Based Proteomic Analysis of Cells Cultured in DMEM 10% FBS and Chemically Defined Medium Using Human Adipose-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2018, 19, 2042. [Google Scholar] [CrossRef] [PubMed]
- Tudyk, J.; Forsthuber, T. ERK2 regulation of mitophagy is important for T cell activation. J. Immunol. 2016, 196, 204. [Google Scholar]
- D’Souza, W.N.; Chang, C.F.; Fischer, A.M.; Li, M.Q.; Hedrick, S.M. The Erk2 MAPK Regulates CD8 T Cell Proliferation and Survival. J.Immunol. 2008, 181, 7617–7629. [Google Scholar] [CrossRef]
- Veillette, A.; Davidson, D. IMMUNOLOGY When lymphocytes run out of steam. Nature 2014, 510, 222–223. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Palmic, N.; Sanquer, S.; Lenoir, C.; Hauck, F.; Mongellaz, D.; Fabrega, S.; Nitschke, P.; Esposti, M.D.; Schwartzentruber, J.; et al. CTP synthase 1 deficiency in humans reveals its central role in lymphocyte proliferation (vol 510, pg 288, 2014). Nature 2014, 511, 288–292. [Google Scholar] [CrossRef]
- Asari, S.; Itakura, S.; Ferreri, K.; Liu, C.P.; Kuroda, Y.; Kandeel, F.; Mullen, Y. Mesenchymal stem cells suppress B-cell terminal differentiation. Exp. Hematol. 2009, 37, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Wan, Q.; Huang, J.; Han, L.; Chen, X.; Chen, G.; Olsen, N.; Zheng, S.G.; Liang, D. Culture medium from TNF-alpha-stimulated mesenchymal stem cells attenuates allergic conjunctivitis through multiple antiallergic mechanisms. J. Allergy Clin. Immunol. 2015, 136, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Klinker, M.W.; Lundy, S.K. Multiple Mechanisms of Immune Suppression by B Lymphocytes. Mol. Med. 2012, 18, 123–137. [Google Scholar] [CrossRef]
- Akiyama, K.; Chen, C.; Wang, D.D.; Xu, X.T.; Qu, C.Y.; Yamaza, T.; Cai, T.; Chen, W.J.; Sun, L.Y.; Shi, S.T. Mesenchymal-Stem-Cell-Induced Immunoregulation Involves FAS-Ligand-/FAS-Mediated T Cell Apoptosis. Cell Stem. Cell. 2012, 10, 544–555. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qi, F.; Dai, X.C.; Tian, W.J.; Liu, T.; Han, H.Q.; Zhang, B.; Li, H.Y.; Zhang, Z.X.; Du, C.G. Requirement of B7-H1 in mesenchymal stem cells for immune tolerance to cardiac allografts in combination therapy with rapamycin. Transpl. Immunol. 2014, 31, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.X.; Hu, C.X.; Chen, J.J.; Cen, P.P.; Wang, J.; Li, L.J. Interaction between Mesenchymal Stem Cells and B-Cells. Int. J. Mol. Sci. 2016, 17, 650. [Google Scholar] [CrossRef]
- Crossley, M.P.; Cimprich, K.A. Faulty replication can sting. Nature 2018, 557, 34–35. [Google Scholar] [CrossRef]
- Luan, Z.G.; Zhang, H.; Yang, P.T.; Ma, X.C.; Zhang, C.; Guo, R.X. HMGB1 activates nuclear factor-kappa B signaling by RAGE and increases the production of TNF-alpha in human umbilical vein endothelial cells. Immunobiology 2010, 215, 956–962. [Google Scholar] [CrossRef]
- Bora, P.; Majumdar, A.S. Adipose tissue-derived stromal vascular fraction in regenerative medicine: A brief review on biology and translation. Stem. Cell Res. Ther. 2017, 8, 145. [Google Scholar] [CrossRef] [PubMed]
- Nahar, S.; Nakashima, Y.; Miyagi-Shiohira, C.; Kinjo, T.; Kobayashi, N.; Saitoh, I.; Watanabe, M.; Noguchi, H.; Fujita, J. A Comparison of the Preservation of Mouse Adipose Tissue-Derived Mesenchymal Stem Cells Using the University of Wisconsin Solution and Hank’s Balanced Salt Solution. Stem. Cells Int. 2018. [Google Scholar] [CrossRef] [PubMed]
- Nahar, S.; Nakashima, Y.; Miyagi-Shiohira, C.; Kinjo, T.; Kobayashi, N.; Saitoh, I.; Watanabe, M.; Noguchi, H.; Fujita, J. A Comparison of Proteins Expressed between Human and Mouse Adipose-Derived Mesenchymal Stem Cells by a Proteome Analysis through Liquid Chromatography with Tandem Mass Spectrometry. Int. J. Mol. Sci. 2018, 19, 3497. [Google Scholar] [CrossRef] [PubMed]
- Miyagi-Shiohira, C.; Nakashima, Y.; Kobayashi, N.; Saitoh, I.; Watanabe, M.; Noguchi, H. Characterization of induced tissue-specific stem cells from pancreas by a synthetic self-replicative RNA. Sci. Rep-Uk 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, K.; Tomita, M.; Ishihama, Y. emPAI Calc-for the estimation of protein abundance from large-scale identification data by liquid chromatography-tandem mass spectrometry. Bioinformatics 2010, 26, 576–577. [Google Scholar] [CrossRef] [PubMed]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially modified protein abundance index (emPAI) for estimation of absolute protein amount in proteomics by the number of sequenced peptides per protein. Mol. Cell. Proteomics. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.H.; Smith, J.W.; Huang, C.M. Mass Spectrometry-Based Label-Free Quantitative Proteomics. J. Biomed. Biotechnol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Rappsilber, J.; Ryder, U.; Lamond, A.I.; Mann, M. Large-scale proteomic analysis of the human spliceosome. Genome Res. 2002, 12, 1231–1245. [Google Scholar] [CrossRef]
- Nakashima, Y.; Nahar, S.; Miyagi-Shiohira, C.; Kinjo, T.; Toyoda, Z.; Kobayashi, N.; Saitoh, I.; Watanabe, M.; Fujita, J.; Noguchi, H. A Liquid Chromatography with Tandem Mass Spectrometry-Based Proteomic Analysis of the Proteins Secreted by Human Adipose-Derived Mesenchymal Stem Cells. Cell. Transplant. 2018, 27, 1469–1494. [Google Scholar] [CrossRef]
- Nakashima, Y.; Miyagi-Shiohira, C.; Kobayashi, N.; Saitoh, I.; Watanabe, M.; Noguchi, H. A proteome analysis of pig pancreatic islets and exocrine tissue by liquid chromatography with tandem mass spectrometry. Islets 2017, 9, 159–176. [Google Scholar] [CrossRef]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakashima, Y.; Nahar, S.; Miyagi-Shiohira, C.; Kinjo, T.; Kobayashi, N.; Kitamura, S.; Saitoh, I.; Watanabe, M.; Fujita, J.; Noguchi, H. Identification of Proteins Differentially Expressed by Adipose-derived Mesenchymal Stem Cells Isolated from Immunodeficient Mice. Int. J. Mol. Sci. 2019, 20, 2672. https://doi.org/10.3390/ijms20112672
Nakashima Y, Nahar S, Miyagi-Shiohira C, Kinjo T, Kobayashi N, Kitamura S, Saitoh I, Watanabe M, Fujita J, Noguchi H. Identification of Proteins Differentially Expressed by Adipose-derived Mesenchymal Stem Cells Isolated from Immunodeficient Mice. International Journal of Molecular Sciences. 2019; 20(11):2672. https://doi.org/10.3390/ijms20112672
Chicago/Turabian StyleNakashima, Yoshiki, Saifun Nahar, Chika Miyagi-Shiohira, Takao Kinjo, Naoya Kobayashi, Shinji Kitamura, Issei Saitoh, Masami Watanabe, Jiro Fujita, and Hirofumi Noguchi. 2019. "Identification of Proteins Differentially Expressed by Adipose-derived Mesenchymal Stem Cells Isolated from Immunodeficient Mice" International Journal of Molecular Sciences 20, no. 11: 2672. https://doi.org/10.3390/ijms20112672
APA StyleNakashima, Y., Nahar, S., Miyagi-Shiohira, C., Kinjo, T., Kobayashi, N., Kitamura, S., Saitoh, I., Watanabe, M., Fujita, J., & Noguchi, H. (2019). Identification of Proteins Differentially Expressed by Adipose-derived Mesenchymal Stem Cells Isolated from Immunodeficient Mice. International Journal of Molecular Sciences, 20(11), 2672. https://doi.org/10.3390/ijms20112672