Leptin and Immunological Profile in Obesity and Its Associated Diseases in Dogs



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Physiological Role of Leptin and Its Relationship with Obesity
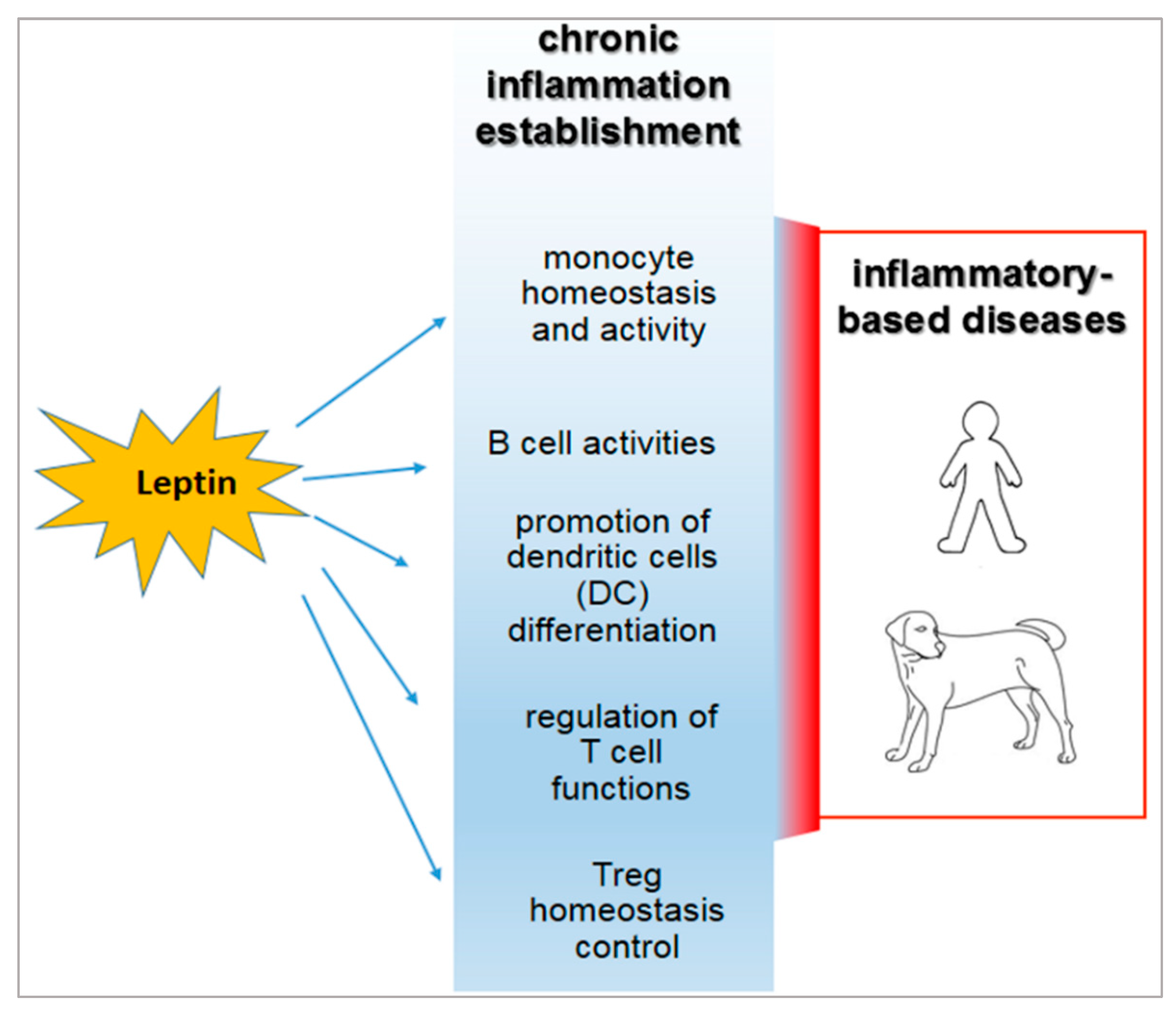
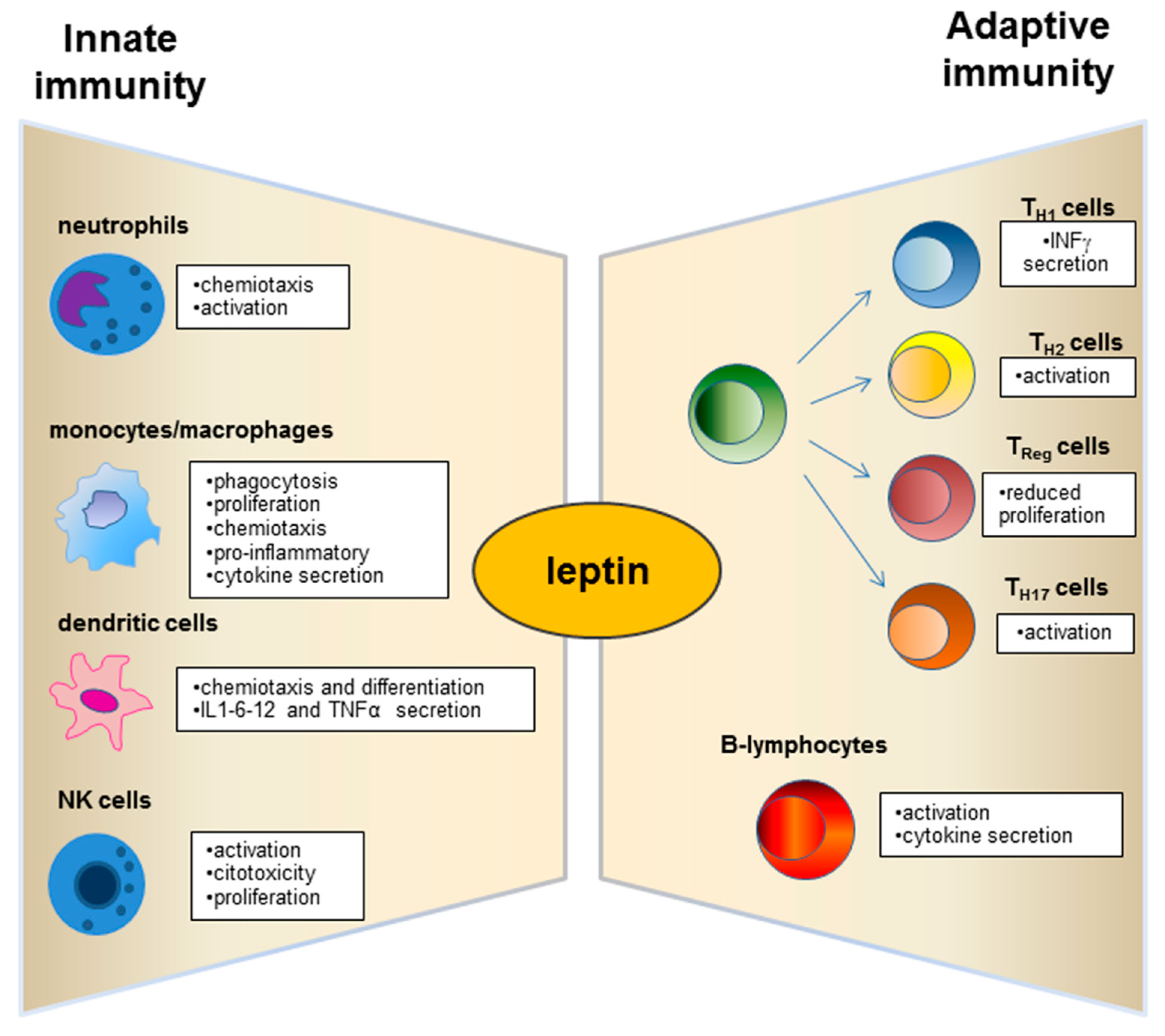
3. Role of Leptin in the Relationship between Obesity and Immune-Modulation.
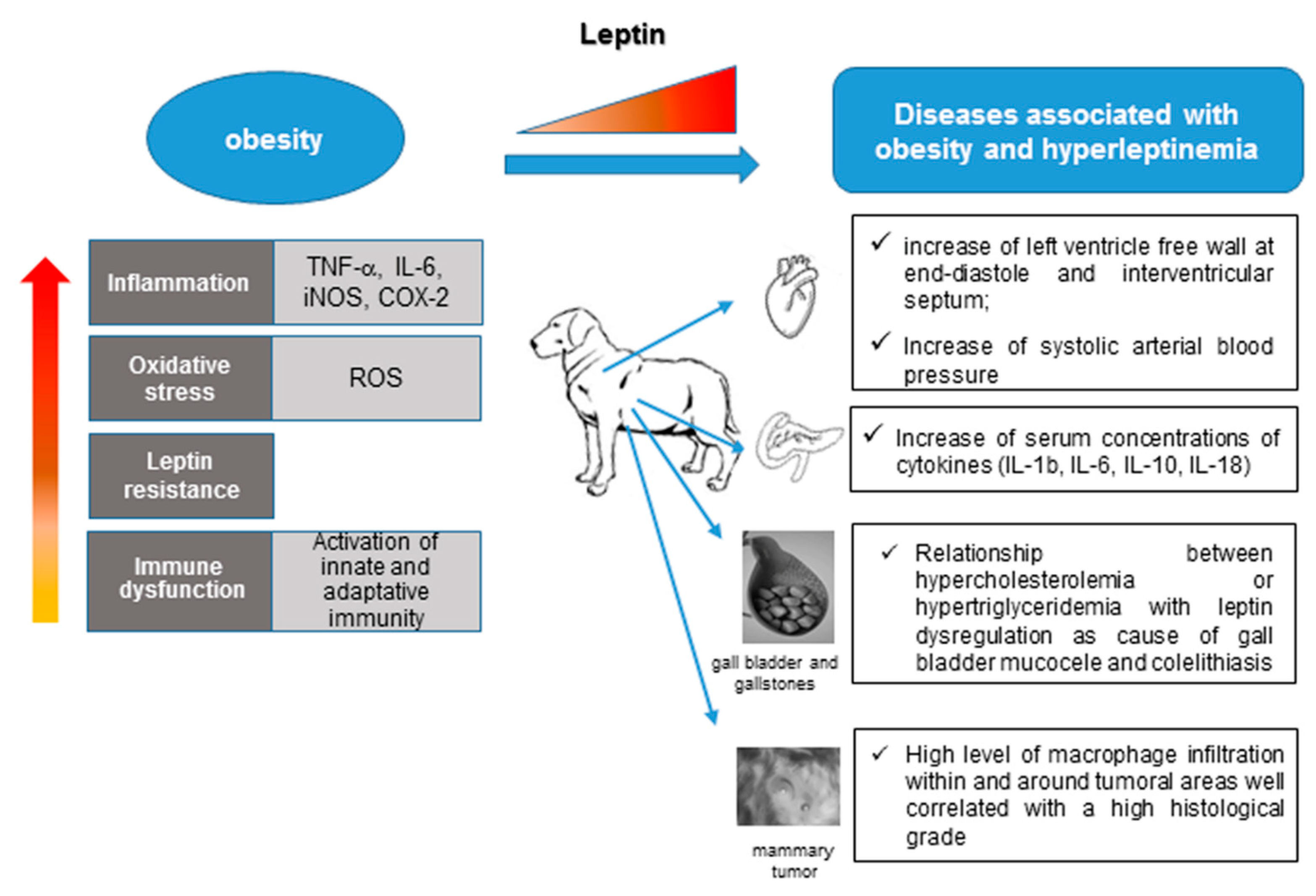
4. Leptin and Associated Diseases in Humans and Dogs
5. Conclusions
Funding
Conflicts of Interest
Abbreviations
| apoE | Apolipoprotein E |
| AP | Acute pancreatitis |
| BCS | Body condition score |
| CDi | Cardiac disease |
| CHF | Congestive heart failure |
| CNS | Central nervous system |
| CRP | C-reactive protein |
| DC | Dendritic cells |
| GBM | Gallbladder mucocele |
| HF | Heart failure |
| IFN | Interferon |
| IL | Interleukin |
| JAK | Janus kinase |
| LEPR | Leptin receptor |
| LVH | Left ventricle hypertrophy |
| MAPK | Mitogen-activated protein kinases |
| MC | Mammary carcinoma |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MS | Metabolic Syndrome |
| VEGF | Vascular growth factor |
| VL | Visceral Leishmaniasis |
| ROR | Retinoic acid-related orphan receptor |
| ROS | Reactive oxygen species |
| NK | Natural Killer |
| STAT | Signal transducer and activator of transcription |
| TNF | Tumour necrosis factor |
| Treg cells | Regulatory T cells |
| VSMCs | Vascular smooth muscle cells |
References
- Tremmel, M.; Gerdtham, U.G.; Nilsson, P.M.; Saha, S. Economic burden of obesity: A systematic literature review. Int. J. Environ. Res. Public Health 2017, 14, 435. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, A. Obesity, metabolic syndrome, and type 2 diabetes: Inflammatory basis of glucose metabolic disorders. Nutr. Rev. 2007, 65, S152–S156. [Google Scholar] [CrossRef] [PubMed]
- Mihalko, W.M.; Bergin, P.F.; Kelly, F.B.; Canale, S.T. Obesity, orthopaedics, and outcomes. J. Am. Acad. Orthop. Surg. 2014, 22, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Zammit, C.; Liddicoat, H.; Moonsie, I.; Makker, H. Obesity and respiratory diseases. Int. J. Gen. Med. 2010, 3, 335–343. [Google Scholar]
- Câmara, N.O.; Iseki, K.; Kramer, H.; Liu, Z.H.; Sharma, K. Kidney disease and obesity: Epidemiology, mechanisms and treatment. Nat. Rev. Nephrol. 2017, 13, 181–190. [Google Scholar] [CrossRef]
- German, A.J. The growing problem of obesity in dogs and cats. J. Nutr. 2006, 136, 1940S–1946S. [Google Scholar] [CrossRef]
- La Cava, A. Leptin in inflammation and autoimmunity. Cytokine 2017, 98, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Abella, V.; Scotece, M.; Conde, J.; Pino, J.; Gonzalez-Gay, M.A.; Gómez-Reino, J.J.; Mera, A.; Lago, F.; Gómez, R.; Gualillo, O. Leptin in the interplay of inflammation, metabolism and immune system disorders. Nat. Rev. Rheumatol. 2017, 13, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef]
- Lago, F.; Dieguez, C.; Gómez-Reino, J.; Gualillo, O. Adipokines as emerging mediators of immune response and inflammation. Nat. Clin. Pract. Rheumatol. 2007, 3, 716–724. [Google Scholar] [CrossRef]
- Lago, F.; Dieguez, C.; Gómez-Reino, J.; Gualillo, O. The emerging role of adipokines as mediators of inflammation and immune responses. Cytokine Growth Factor Rev. 2007, 18, 313–325. [Google Scholar] [CrossRef]
- Lago, F.; Gómez, R.; Gómez-Reino, J.J.; Dieguez, C.; Gualillo, O. Adipokines as novel modulators of lipid metabolism. Trends Biochem. Sci. 2009, 34, 500–510. [Google Scholar] [CrossRef]
- Ahima, R.S.; Flier, J.S. Leptin. Annu. Rev. Physiol. 2000, 62, 413–437. [Google Scholar] [CrossRef]
- Park, H.K.; Ahima, R.S. Physiology of leptin: Energy homeostasis, neuroendocrine function and metabolism. Metabolism 2015, 64, 24–34. [Google Scholar] [CrossRef]
- Friedman, J. The long road to leptin. J. Clin. Investig. 2016, 126, 4727–4734. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Havel, P.J. Role of adipose tissue in body-weight regulation: Mechanisms regulating leptin production and energy balance. Proc. Nutr. Soc. 2000, 59, 359–371. [Google Scholar] [CrossRef]
- Lee, G.H.; Proenca, R.; Montez, J.M.; Carroll, K.M.; Darvishzadeh, J.G.; Lee, J.I.; Friedman, J.M. Abnormal splicing of the leptin receptor in diabetic mice. Nature 1996, 379, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Armağan, C.; Yılmaz, C.; Koç, A.; Abac, A.; Ülgenalp, A.; Böber, E.; Erçal, D.; Demir, K. A toddler with a novel LEPR mutation. Hormones 2019. [Google Scholar] [CrossRef]
- Wasim, M.; Awan, F.R.; Najam, S.S.; Khan, A.R.; Khan, H.N. Role of Leptin Deficiency, Inefficiency, and Leptin Receptors in Obesity. Biochem. Genet. 2016, 54, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, L.A. The leptin receptor. J. Biol. Chem. 1997, 272, 6093–6096. [Google Scholar] [CrossRef]
- Gorska, E.; Popko, K.; Stelmaszczyk-Emmel, A.; Ciepiela, O.; Kucharska, A.; Wasik, M. Leptin receptors. Eur. J. Med. Res. 2010, 15, 50–54. [Google Scholar]
- Fei, H.; Okano, H.J.; Li, C.; Lee, G.H.; Zhao, C.; Darnell, R.; Friedman, J.M. Anatomic localization of alternatively spliced leptin receptors (Ob-R) in mouse brain and other tissues. Proc. Natl. Acad. Sci. USA 1997, 94, 7001–7005. [Google Scholar] [CrossRef]
- Yamashita, T.; Murakami, T.; Otani, S.; Kuwajima, M.; Shima, K. Leptin receptor signal transduction: OBRa and OBRb of fa type. Biochem. Biophys. Res. Commun. 1998, 246, 752. [Google Scholar] [CrossRef]
- Margetic, S.; Gazzola, C.; Pegg, G.G.; Hill, R.A. Leptin: A review of its peripheral actions and interactions. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 1407–1433. [Google Scholar] [CrossRef]
- Frühbeck, G. Intracellular signalling pathways activated by leptin. Biochem. J. 2006, 393, 7–20. [Google Scholar] [CrossRef]
- Satoh, N.; Ogawa, Y.; Katsuura, G.; Numata, Y.; Tsuji, T.; Hayase, M.; Ebihara, K.; Masuzaki, H.; Hosoda, K.; Yoshimasa, Y.; et al. Sympathetic activation of leptin via the ventromedial hypothalamus: Leptin-induced increase in catecholamine secretion. Diabetes 1999, 48, 1787–1793. [Google Scholar] [CrossRef] [PubMed]
- Dalamaga, M.; Chou, S.H.; Shields, K.; Papageorgiou, P.; Polyzos, S.A.; Mantzoros, C.S. Leptin at the intersection of neuroendocrinology and metabolism: Current evidence and therapeutic perspectives. Cell Metabol. 2013, 18, 29–42. [Google Scholar] [CrossRef]
- Fantuzzi, G.; Faggioni, R. Leptin in the regulation of immunity, inflammation, and hematopoiesis. J. Leukoc. Biol. 2000, 68, 437–446. [Google Scholar]
- Pérez-Pérez, A.; Vilariño-García, T.; Fernández-Riejos, P.; Martín-González, J.; Segura-Egea, J.J.; Sánchez-Margalet, V. Role of leptin as a link between metabolism and the immune system. Cytokine Growth Factor Rev. 2017, 35, 71–84. [Google Scholar] [CrossRef]
- Gainsford, T.; Willson, T.A.; Metcalf, D.; Handman, E.; McFarlane, C.; Nq, A.; Nicola, N.A.; Alexander, W.S.; Hilton, D.J. Leptin can induce proliferation, differentiation, and functional activation of hemopoietic cells. Proc. Natl. Acad. Sci. USA 1996, 93, 14564–14568. [Google Scholar] [CrossRef]
- Bennett, B.D.; Solar, G.P.; Yuan, J.Q.; Mathias, J.; Thomas, G.R.; Matthews, W. A role for leptin and its cognate receptor in hematopoiesis. Curr. Biol. 1996, 6, 1170–1180. [Google Scholar] [CrossRef]
- Claycombe, K.; King, L.E.; Fraker, P.J. A role for leptin in sustaining lymphopoiesis and myelopoiesis. Proc. Natl. Acad. Sci. USA 2008, 105, 2017–2021. [Google Scholar] [CrossRef] [PubMed]
- Han, T.J.; Wang, X. Leptin and its receptor in hematologic malignancies. Int. J. Clin. Exp. Med. 2015, 8, 19840–19849. [Google Scholar]
- Ahima, R.S.; Prabakaran, D.; Mantzoros, C.; Qu, D.; Lowell, B.; Maratos-Flier, E.; Flier, J.S. Role of leptin in the neuroendocrine response to fasting. Nature 1996, 382, 250–252. [Google Scholar] [CrossRef]
- Myers, M.G., Jr.; Heymsfield, S.B.; Haft, C.; Kahn, B.B.; Laughlin, M.; Leibel, R.L.; Tschöp, M.H.; Yanovski, J.A. Challenges and opportunities of defining clinical leptin resistance. Cell Metabol. 2012, 15, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Kim, M.S. Molecular mechanisms of central leptin resistance in obesity. Arch. Pharm. Res. 2013, 36, 201–207. [Google Scholar] [CrossRef]
- Zoran, D.L. Obesity in dogs and cats: A metabolic and endocrine disorder. Vet. Clin. North Am. Small Anim. Pract. 2010, 40, 221–239. [Google Scholar] [CrossRef]
- Packer, M. Epicardial adipose tissue may mediate deleterious effects of obesity and inflammation on the myocardium. J. Am. Coll. Cardiol. 2018, 71, 2360–2372. [Google Scholar] [CrossRef]
- Russo, I. The prothrombotic tendency in metabolic syndrome: Focus on the potential mechanisms involved in impaired haemostasis and fibrinolytic balance. Scientifica 2012, 2012, 525374. [Google Scholar] [CrossRef] [PubMed]
- Barbosa-Ferreira, J.M.; Fernandes, F.; Dabarian, A.; Mady, C. Leptin in heart failure. Expert. Opin. Med. Diagn. 2013, 7, 113–117. [Google Scholar] [CrossRef]
- Chiba, T.; Shinozaki, S.; Nakazawa, T.; Kawakami, A.; Ai, M.; Kaneko, E.; Kitagawa, M.; Kondo, K.; Chait, A.; Shimokado, K. Leptin deficiency suppresses progression of atherosclerosis in apoE-deficient mice. Atherosclerosis 2008, 196, 68–75. [Google Scholar] [CrossRef]
- Lopaschuk, G.D.; Folmes, C.D.; Stanley, W.C. Cardiac energy metabolism in obesity. Circ. Res. 2007, 101, 335–347. [Google Scholar] [CrossRef]
- Tune, J.D.; Considine, R.V. Effects of leptin on cardiovascular physiology. J. Am. Soc. Hypertens. 2007, 1, 231–241. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Balasubramanian, P.; Hall, D.; Subramanian, M. Sympathetic nervous system as a target for aging and obesity-related cardiovascular diseases. Geroscience 2019, 41, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Oda, A.; Taniguchi, T.; Yokoyama, M. Leptin stimulates rat aortic smooth muscle cell proliferation and migration. Kobe J. Med. Sci. 2001, 47, 141–150. [Google Scholar] [CrossRef]
- Fortuño, A.; Rodríguez, A.; Gómez-Ambrosi, J.; Muñiz, P.; Salvador, J.; Díez, J.; Frühbeck, G. Leptin inhibits angiotensin II-induced intracellular calcium increase and vasoconstriction in the rat aorta. Endocrinology 2002, 143, 3555–3560. [Google Scholar] [CrossRef] [PubMed]
- Suganami, T.; Mukoyama, M.; Mori, K.; Yokoi, H.; Koshikawa, M.; Sawai, K.; Hidaka, S.; Ebihara, K.; Tanaka, T.; Sugawara, A.; et al. Prevention and reversal of renal injury by leptin in a new mouse model of diabetic nephropathy. FASEB J. 2005, 19, 127–129. [Google Scholar] [CrossRef]
- Morales-Ruiz, M.; Fulton, D.; Sowa, G.; Languino, L.R.; Fujio, Y.; Walsh, K.; Sessa, W.C. Vascular endothelial growth factor-stimulated actin reorganization and migration of endothelial cells is regulated via the serine/threonine kinase Akt. Circ. Res. 2000, 86, 892–896. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Jaworek, J.; Maniatoglou, A.; Bonior, J.; Meixner, H.; Konturek, S.J.; Hahn, E.G. Leptin modulates the inflammatory response in acute pancreatitis. Digestion 2002, 65, 149–160. [Google Scholar] [CrossRef]
- Frossard, J.L.; Lescuyer, P.; Pastor, C.M. Experimental evidence of obesity as a risk factor for severe acute pancreatitis. World J. Gastroenterol. 2009, 15, 5260–5265. [Google Scholar] [CrossRef]
- Karpavicius, A.; Dambrauskas, Z.; Sileikis, A.; Vitkus, D.; Strupas, K. Value of adipokines in predicting the severity of acute pancreatitis: Comprehensive review. World J. Gastroenterol. 2012, 18, 6620–6627. [Google Scholar] [CrossRef]
- Konturek, P.C.; Konturek, S.J.; Brzozowski, T.; Jaworek, J.; Hahn, E.G. Role of leptin in the stomach and the pancreas. J. Physiol. 2001, 95, 345–354. [Google Scholar] [CrossRef]
- Warzecha, Z.; Dembiński, A.; Ceranowicz, P.; Jaworek, J.; Konturek, P.C.; Dembiński, M.; Bilskl, J.; Konturek, S.J. Influence of leptin administration on the course of acute ischemic pancreatitis. J. Physiol. Pharmacol. 2002, 53, 775–790. [Google Scholar] [PubMed]
- Gultekin, F.A.; Kerem, M.; Tatlicioglu, E.; Aricioglu, A.; Unsal, C.; Bukan, N. Leptin treatment ameliorates acute lung injury in rats with cerulein-induced acute pancreatitis. World J. Gastroenterol. 2007, 13, 2932–2938. [Google Scholar] [CrossRef]
- Carbone, F.; La Rocca, C.; De Candia, P.; Procaccini, C.; Colamatteo, A.; Micillo, T.; De Rosa, V.; Matarese, G. Metabolic control of immune tolerance in health and autoimmunity. Semin. Immunol. 2016, 28, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Medina, G.; Vera-Lastra, O.; Peralta-Amaro, A.L.; Jiménez-Arellano, M.P.; Saavedra, M.A.; Cruz-Domínguez, M.P.; Jara, L.J. Metabolic syndrome, autoimmunity and rheumatic diseases. Pharmacol. Res. 2018, 133, 277–288. [Google Scholar] [CrossRef]
- Katona, P.; Katona-Apte, J. The interaction between nutrition and infection. Clin. Infect. Dis. 2008, 46, 1582–1588. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, S.; Ovchinnikov, R.S. The relationship between nutrition and infectious diseases: A review. Biomed. Biotechnol. Res. J. 2018, 2, 168–172. [Google Scholar]
- Hennig, B.; Petriello, M.C.; Gamble, M.V.; Surh, Y.J.; Kresty, L.A.; Frank, N.; Rangkadilok, N.; Ruchirawat, M.; Suk, W.A. The role of nutrition in influencing mechanisms involved in environmentally mediated diseases. Rev. Environ. Health 2018, 33, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Maurya, R.; Bhattacharya, P.; Dey, R.; Nakhasi, H.L. Leptin Functions in Infectious Diseases. Front. Immunol. 2018, 9, 2741. [Google Scholar] [CrossRef]
- Naylor, C.; Petri, W.A., Jr. Leptin regulation of immune responses. Trends Mol. Med. 2016, 22, 88–98. [Google Scholar] [CrossRef]
- Maldonado-Ruiz, R.; Fuentes-Mera, L.; Camacho, A. Central modulation of neuroinflammation by neuropeptides and energy-sensing hormones during obesity. Biomed. Res. Int. 2017, 2017, 7949582. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, E.V.; Liu, A.; Matarese, G.; La Cava, A. Leptin promotes systemic lupus erythematosus by increasing autoantibody production and inhibiting immune regulation. Proc. Natl. Acad. Sci. USA 2016, 113, 10637–10642. [Google Scholar]
- Palatucci, A.T.; Piantedosi, D.; Rubino, V.; Giovazzino, A.; Guccione, J.; Pernice, V.; Ruggiero, G.; Cortese, L.; Terrazzano, G. Circulating regulatory T cells (Treg), leptin and induction of proinflammatory activity in obese Labrador Retriever dogs. Vet. Immunol. Immunopathol. 2018, 202, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, I.M.; Jeremias, J.T.; Takeara, P.; de Souza, D.F.; Balieiro, J.C.C.; Pfrimer, K.; Brunetto, M.A.; Pontieri, C.F.F. Effect of dietary protein intake on the body composition and metabolic parameters of neutered dogs. J. Nutr. Sci. 2017, 6, e40. [Google Scholar] [CrossRef]
- Kim, A.Y.; Kim, H.S.; Kang, J.H.; Yang, M.P. Serum adipokine concentrations in dogs with diabetes mellitus: A pilot study. J. Vet. Sci. 2015, 16, 333–340. [Google Scholar] [CrossRef][Green Version]
- Lam, Q.L.; Lu, L. Role of leptin in immunity. Cell. Mol. Immunol. 2007, 4, 1–13. [Google Scholar]
- Francisco, V.; Pino, J.; Campos-Cabaleiro, V.; Ruiz-Fernández, C.; Mera, A.; Gonzalez-Gay, M.A.; Gómez, R.; Gualillo, O. Obesity, fat mass and immune system: Role for leptin. Front. Physiol. 2018, 9, 640. [Google Scholar] [CrossRef]
- Friedman, J.M.; Halaas, J.L. Leptin and the regulation of body weight in mammals. Nature 1998, 395, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, L.A.; Dembski, M.; Weng, X.; Deng, N.; Culpepper, J.; Devos, R.; Richards, G.J.; Campfield, L.A.; Clark, F.T.; Deeds, J.; et al. Identification and expression cloning of a leptin receptor, OB-R. Cell 1995, 83, 1263–1271. [Google Scholar] [CrossRef]
- Matarese, G.; La Cava, A. The intricate interface between immune system and metabolism. Trends Immunol. 2004, 25, 193–200. [Google Scholar] [CrossRef]
- Chan, J.L.; Matarese, G.; Shetty, G.K.; Raciti, P.; Kelesidis, I.; Aufiero, D.; De Rosa, V.; Perna, F.; Fontana, S.; Mantzoros, C.S. Differential regulation of metabolic, neuroendocrine, and immune function by leptin in humans. Proc. Natl. Acad. Sci. USA 2006, 103, 8481–8486. [Google Scholar] [CrossRef]
- Procaccini, C.; Jirillo, E.; Matarese, G. Leptin as an immunomodulator. Mol. Aspects Med. 2012, 33, 35–45. [Google Scholar] [CrossRef]
- La Cava, A.; Matarese, G. The weight of leptin in immunity. Nat. Rev. Immunol. 2004, 4, 371–379. [Google Scholar] [CrossRef]
- Bernotiene, E.; Palmer, G.; Gabay, C. The role of leptin in innate and adaptive immune responses. Arthritis. Res. Ther. 2006, 8, 217. [Google Scholar] [CrossRef][Green Version]
- Hsu, A.; Aronoff, D.M.; Phipps, J.; Goel, D.; Mancuso, P. Leptin improves pulmonary bacterial clearance and survival in ob/ob mice during pneumococcal pneumonia. Clin. Exp. Immunol. 2007, 150, 332–339. [Google Scholar] [CrossRef]
- Gruen, M.L.; Hao, M.; Piston, D.W.; Hasty, A.H. Leptin requires canonical migratory signaling pathways for induction of monocyte and macrophage chemotaxis. Am. J. Physiol. Cell Physiol. 2007, 293, C1481–C1488. [Google Scholar] [CrossRef]
- Dib, L.H.; Ortega, M.T.; Fleming, S.D.; Chapes, S.K.; Melgarejo, T. Bone marrow leptin signaling mediates obesity-associated adipose tissue inflammation in male mice. Endocrinology 2014, 155, 40–46. [Google Scholar] [CrossRef]
- Conde, J.; Scotece, M.; Gómez, R.; Gómez-Reino, J.J.; Lago, F.; Gualillo, O. At the crossroad between immunity and metabolism: Focus on leptin. Expert. Rev. Clin. Immunol. 2010, 6, 801–808. [Google Scholar] [CrossRef]
- Sánchez-Pozo, C.; Rodriguez-Baño, J.; Domínguez-Castellano, A.; Muniain, M.A.; Goberna, R.; Sánchez-Margalet, V. Leptin stimulates the oxidative burst in control monocytes but attenuates the oxidative burst in monocytes from HIV-infected patients. Clin. Exp. Immunol. 2003, 134, 464–469. [Google Scholar] [CrossRef]
- Kohchi, C.; Inagawa, H.; Nishizawa, T.; Soma, G. ROS and innate immunity. Anticancer Res. 2009, 29, 817–821. [Google Scholar]
- Najib, S.; Sánchez-Margalet, V. Human leptin promotes survival of human circulating blood monocytes prone to apoptosis by activation of p42/44 MAPK pathway. Cell Immunol. 2002, 220, 143–149. [Google Scholar] [CrossRef]
- Faggioni, R.; Feingold, K.R.; Grunfeld, C. Leptin regulation of the immune response and the immunodeficiency of malnutrition. FASEB J. 2001, 15, 2565–2571. [Google Scholar] [CrossRef]
- Bruno, A.; Conus, S.; Schmid, I.; Simon, H.U. Apoptotic pathways are inhibited by leptin receptor activation in neutrophils. J. Immunol. 2005, 174, 8090–8096. [Google Scholar] [CrossRef] [PubMed]
- Zarkesh-Esfahani, H.; Pockley, A.G.; Wu, Z.; Hellewell, P.G.; Weetman, A.P.; Ross, R.J. Leptin indirectly activates human neutrophils via induction of TNF-alpha. J. Immunol. 2004, 172, 1809–1814. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, B.; Straface, E.; Quaranta, M.G.; Giordani, L.; Viora, M. Leptin promotes differentiation and survival of human dendritic cells and licenses them for Th1 priming. J. Immunol. 2005, 174, 6820–6828. [Google Scholar] [CrossRef]
- Lam, Q.L.K.; Liu, S.; Cao, X.; Lu, L. Involvement of leptin signaling in the survival and maturation of bone marrow-derived dendritic cells. Eur. J. Immunol. 2006, 36, 3118–3130. [Google Scholar] [CrossRef]
- Laue, T.; Wrann, C.D.; Hoffmann-Castendiek, B.; Pietsch, D.; Hübner, L.; Kielstein, H. Altered NK cell function in obese healthy humans. BMC Obes. 2015, 2, 1. [Google Scholar] [CrossRef] [PubMed]
- Jahn, J.; Spielau, M.; Brandsch, C.; Stangl, G.I.; Delank, K.S.; Bähr, I.; Berreis, T.; Wrann, C.D.; Kielstein, H. Decreased NK cell functions in obesity can be reactivated by fat mass reduction. Obesity 2015, 23, 2233–2241. [Google Scholar] [CrossRef]
- Tian, Z.; Sun, R.; Wei, H.; Gao, B. Impaired natural killer (NK) cell activity in leptin receptor deficient mice: Leptin as a critical regulator in NK cell development and activation. Biochem. Biophys. Res. Commun. 2002, 298, 297–302. [Google Scholar] [CrossRef]
- Howard, J.K.; Lord, G.M.; Matarese, G.; Vendetti, S.; Ghatei, M.A.; Ritter, M.A.; Lechler, R.I.; Bloom, S.R. Leptin protects mice from starvation-induced lymphoid atrophy and increases thymic cellularity in ob/ob mice. J. Clin. Investig. 1999, 104, 1051–1059. [Google Scholar] [CrossRef]
- Sánchez-Margalet, V.; Martín-Romero, C.; González-Yanes, C.; Goberna, R.; Rodríguez-Baño, J.; Muniain, M.A. Leptin receptor (Ob-R) expression is induced in peripheral blood mononuclear cells by in vitro activation and in vivo in HIV-infected patients. Clin. Exp. Immunol. 2002, 129, 119–124. [Google Scholar] [CrossRef]
- Martín-Romero, C.; Santos-Alvarez, J.; Goberna, R.; Sánchez-Margalet, V. Human leptin enhances activation and proliferation of human circulating T lymphocytes. Cell Immunol. 2000, 199, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Lord, G.M.; Matarese, G.; Howard, J.K.; Baker, R.J.; Bloom, S.R.; Lechler, R.I. Leptin modulates the T-cell immune response and reverses starvation-induced immunosuppression. Nature 1998, 394, 897–901. [Google Scholar] [CrossRef]
- Batra, A.; Okur, B.; Glauben, R.; Erben, U.; Ihbe, J.; Stroh, T.; Fedke, I.; Chang, H.D.; Zeitz, M.; Siegmund, B. Leptin: A critical regulator of CD4+ T-cell polarization in vitro and in vivo. Endocrinology 2010, 151, 56–62. [Google Scholar] [CrossRef]
- Deng, J.; Liu, Y.; Yang, M.; Wang, S.; Zhang, M.; Wang, X.; Ko, K.H.; Hua, Z.; Sun, L.; Cao, X.; et al. Leptin exacerbates collagen-induced arthritis via enhancement of Th17 cell response. Arthritis Rheum. 2012, 64, 3564–3573. [Google Scholar] [CrossRef]
- Reis, B.S.; Lee, K.; Fanok, M.H.; Mascaraque, C.; Amoury, M.; Cohn, L.B.; Rogoz, A.; Dallner, O.S.; Moraes-Vieira, P.M.; Domingos, A.I.; et al. Leptin receptor signaling in T cells is required for Th17 differentiation. J. Immunol. 2015, 194, 5253–5260. [Google Scholar] [CrossRef]
- Lam, Q.L.K.; Wang, S.; Ko, O.K.H.; Kincade, P.W.; Lu, L. Leptin signaling maintains B-cell homeostasis via induction of Bcl-2 and Cyclin D1. Proc. Natl Acad. Sci. USA 2010, 107, 13812–13817. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S. The origin of FOXP3-expressing CD4+ regulatory T cells: Thymus or periphery. J. Clin. Investig. 2003, 112, 1310–1312. [Google Scholar] [CrossRef]
- De Rosa, V.; Procaccini, C.; Calì, G.; Pirozzi, G.; Fontana, S.; Zappacosta, S.; La Cava, A.; Matarese, G. A key role of leptin in the control of regulatory T cell proliferation. Immunity 2007, 26, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Silva Morales, M.; Mueller, D. Anergy into T regulatory cells: An integration of metabolic cues and epigenetic changes at the Foxp3 conserved non-coding sequence 2. F1000Research 2018, 7, 1938. [Google Scholar] [CrossRef]
- Kumar, P.; Bhattacharya, P.; Prabhakar, B.S. A comprehensive review on the role of co-signaling receptors and Treg homeostasis in autoimmunity and tumor immunity. J. Autoimmun. 2018, 95, 77–99. [Google Scholar] [CrossRef]
- Matarese, G.; Procaccini, C.; De Rosa, V.; Horvath, T.L. La Cava, A. Regulatory T cells in obesity: The leptin connection. Trends Mol. Med. 2010, 16, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.E.; Torgerson, T.R.; Schubert, L.A.; Anover, S.D.; Ocheltree, E.L.; Ochs, H.D.; Ziegler, S.F. Analysis of FOXP3 reveals multiple domains required for its function as a transcriptional repressor. J. Immunol. 2006, 177, 3133–3142. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Hanabuchi, S.; Wang, Y.H.; Park, W.R.; Arima, K.; Bover, L.; Qin, F.X.; Gilliet, M.; Liu, Y.J. Two functional subsets of FOXP3+ regulatory T cells in human thymus and periphery. Immunity 2008, 28, 870–880. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Huang, C.; Zhou, B.; Ziegler, S.F. Isoform-specific inhibition of ROR alpha-mediated transcriptional activation by human FOXP3. J. Immunol. 2008, 180, 4785–4792. [Google Scholar] [CrossRef]
- Zhou, L.; Lopes, J.E.; Chong, M.M.; Ivanov, I.I.; Min, R.; Victora, G.D.; Shen, Y.; Du, J.; Rubtsov, Y.P.; Rudensky, A.Y.; et al. TGF-beta-induced Foxp3 inhibits T(H)17 cell differentiation by antagonizing RORgammat function. Nature 2008, 453, 236–240. [Google Scholar] [CrossRef]
- Sambucci, M.; Gargano, F.; De Rosa, V.; De Bardi, M.; Picozza, M.; Placido, R.; Ruggieri, S.; Capone, A.; Gasperini, C.; Matarese, G.; et al. FoxP3 isoforms and PD-1 expression by T regulatory cells in multiple sclerosis. Sci. Rep. 2018, 8, 3674. [Google Scholar] [CrossRef]
- De Rosa, V.; Galgani, M.; Porcellini, A.; Colamatteo, A.; Santopaolo, M.; Zuchegna, C.; Romano, A.; De Simone, S.; Procaccini, C.; La Rocca, C.; et al. Glycolysis controls the induction of human regulatory T cells by modulating the expression of FOXP3 exon 2 splicing variants. Nat. Immunol. 2015, 16, 1174–1184. [Google Scholar] [CrossRef]
- Mailer, R.K.; Joly, A.L.; Liu, S.; Elias, S.; Tegner, J.; Andersson, J. IL-1β promotes Th17 differentiation by inducing alternative splicing of FOXP3. Sci. Rep. 2015, 5, 14674. [Google Scholar] [CrossRef] [PubMed]
- McLeod, L.E.; Proud, C.G. ATP depletion increases phosphorylation of elongation factor eEF2 in adult cardiomyocytes independently of inhibition of mTOR signalling. FEBS Lett. 2002, 531, 448–452. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell. Sci. 2009, 122, 3589–3594. [Google Scholar] [CrossRef]
- Galgani, M.; Procaccini, C.; De Rosa, V.; Carbone, F.; Chieffi, P.; La Cava, A.; Matarese, G. Leptin modulates the survival of autoreactive CD4+ T cells through the nutrient/energy-sensing mammalian target of rapamycin signaling pathway. J. Immunol. 2010, 185, 7474–7479. [Google Scholar] [CrossRef]
- Zeng, H.; Yang, K.; Cloer, C.; Neale, G.; Vogel, P.; Chi, H. mTORC1 couples immune signals and metabolic programming to establish T(reg)-cell function. Nature 2013, 499, 485–490. [Google Scholar] [CrossRef]
- Wang, R.; Dillon, C.P.; Shi, L.Z.; Milasta, S.; Carter, R.; Finkelstein, D.; McCormick, L.L.; Fitzgerald, P.; Chi, H.; Munger, J.; et al. The transcription factor Myc controls metabolic reprogramming upon T lymphocyte activation. Immunity 2011, 35, 871–882. [Google Scholar] [CrossRef]
- Donnelly, R.P.; Finlay, D.K. Glucose, glycolysis and lymphocyte responses. Mol. Immunol. 2015, 68, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Blagih, J.; Coulombe, F.; Vincent, E.E.; Dupuy, F.; Galicia-Vázquez, G.; Yurchenko, E.; Raissi, T.C.; van der Windt, G.J.; Viollet, B.; Pearce, E.L.; et al. The energy sensor AMPK regulates T cell metabolic adaptation and effector responses in vivo. Immunity 2015, 42, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Loftus, R.M.; Finlay, D.K. Immunometabolism: Cellular metabolism turns immune regulator. J. Biol. Chem. 2016, 291, 1–10. [Google Scholar] [CrossRef]
- Gerriets, V.A.; Rathmell, J.C. Metabolic pathways in T cell fate and function. Trends Immunol. 2012, 33, 168–173. [Google Scholar] [CrossRef]
- Procaccini, C.; Carbone, F.; Galgani, M.; La Rocca, C.; De Rosa, V.; Cassano, S.; Matarese, G. Obesity and susceptibility to autoimmune diseases. Expert. Rev. Clin. Immunol. 2011, 7, 287–294. [Google Scholar] [CrossRef]
- Lee, B.C.; Lee, J. Cellular and molecular players in adipose tissue inflammation in the development of obesity-induced insulin resistance. Biochim. Biophys Acta 2014, 1842, 446–462. [Google Scholar] [CrossRef]
- Wang, M.; Chen, F.; Wang, J.; Zeng, Z.; Yang, Q.; Shao, S. Th17 and Treg lymphocytes in obesity and Type 2 diabetic patients. Clin. Immunol. 2018, 197, 77–78. [Google Scholar] [CrossRef]
- Yun, J.M.; Jialal, I.; Devaraj, S. Effects of epigallocatechin gallate on regulatory T cell number and function in obese v. lean volunteers. Br. J. Nutr. 2010, 103, 1771–1777. [Google Scholar] [CrossRef]
- Wagner, N.M.; Brandhorst, G.; Czepluch, F.; Lankeit, M.; Eberle, C.; Herzberg, S.; Faustin, V.; Riggert, J.; Oellerich, M.; Hasenfuss, G.; et al. Circulating regulatory T cells are reduced in obesity and may identify iubjects at increased metabolic and cardiovascular risk. Obesity 2013, 2, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Deiuliis, J.; Shah, Z.; Shah, N.; Needleman, B.; Mikami, D.; Narula, V.; Perry, K.; Hazey, J.; Kampfrath, T.; Kollengode, M.; et al. Visceral adipose inflammation in obesity is associated with critical alterations in T regulatory cell numbers. PLoS ONE 2011, 6, e16376. [Google Scholar] [CrossRef]
- Taleb, S.; Herbin, O.; Ait-Oufella, H.; Verreth, W.; Gourdy, P.; Barateau, V.; Merval, R.; Esposito, B.; Clément, K.; Holvoet, P.; et al. Defective leptin/leptin receptor signaling improves regulatory T cell immune response and protects mice from atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2691–2698. [Google Scholar] [CrossRef]
- Antuna-Puente, B.; Feve, B.; Fellahi, S.; Bastard, J.P. Adipokines: The missing link between insulin resistance and obesity. Diabetes Metab. 2008, 34, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Tvarijonaviciute, A.; Tecles, F.; Martinez-Subiela, S.; Ceron, J.J. Effect of weight loss on inflammatory biomarkers in obese dogs. Vet. J. 2012, 193, 570–572. [Google Scholar] [CrossRef] [PubMed]
- Wakshlag, J.J.; Struble, A.M.; Levine, C.B.; Bushey, J.J.; Laflamme, D.P.; Long, G.M. The effects of weight loss on adipokines and markers of inflammation in dogs. Br. J. Nutr. 2011, 106, S11–S14. [Google Scholar] [CrossRef]
- Bastien, B.C.; Patil, A.; Satyaraj, E. The impact of weight loss on circulating cytokines in Beagle dogs. Vet. Immunol. Immunopathol. 2015, 163, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Sagawa, M.M.; Nakadomo, F.; Honjoh, T.; Ishioka, K.; Saito, M. Correlation between plasma leptin concentration and body fat content in dogs. Am. J. Vet. Res. 2002, 63, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Ishioka, K.; Soliman, M.M.; Sagawa, M.; Nakadomo, F.; Shibata, H.; Honjoh, T.; Hashimoto, A.; Kitamura, H.; Kimura, K.; Saito, M. Experimental and clinical studies on plasma leptin in obese dogs. J. Vet. Med. Sci. 2002, 64, 349–353. [Google Scholar] [CrossRef]
- Ishioka, K.; Hosoya, K.; Kitagawa, H.; Shibata, H.; Honjoh, T.; Kimura, K.; Saito, M. Plasma leptin concentration in dogs: Effects of body condition score, age, gender and breeds. Res. Vet. Sci. 2007, 82, 11–15. [Google Scholar] [CrossRef]
- Jeusette, I.C.; Detilleux, J.; Shibata, H.; Saito, M.; Honjoh, T.; Delobel, A.; Istasse, L.; Diez, M. Effects of chronic obesity and weight loss on plasma ghrelin and leptin concentrations in dogs. Res. Vet. Sci. 2005, 79, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Segal, K.R.; Landt, M.; Klein, S. Relationship between insulin sensitivity and plasma leptin concentration in lean and obese men. Diabetes 1996, 45, 988–991. [Google Scholar] [CrossRef]
- Gayet, C.; Bailhache, E.; Dumon, H.; Martin, L.; Siliart, B.; Nguyen, P. Insulin resistance and changes in plasma concentration of TNFalpha, IGF1, and NEFA in dogs during weight gain and obesity. J. Anim. Physiol. Anim. Nutr. 2004, 88, 157–165. [Google Scholar] [CrossRef]
- Veiga, A.P.M.; Price, C.A.; de Oliveira, S.T.; dos Santos, A.P.; Campos, R.; Barbosa, P.R.; Gonzalez, F.H.D. Association of canine obesity with reduced serum levels of C-reactive protein. J. Vet. Diagn. Investig. 2008, 20, 224–228. [Google Scholar] [CrossRef]
- Van de Velde, H.; Janssens, G.P.; Stuyven, E.; Cox, E.; Buyse, J.; Hesta, M. Short-term increase of body weight triggers immunological variables in dogs. Vet. Immunol. Immunopathol. 2012, 145, 431–437. [Google Scholar] [CrossRef]
- Van de Velde, H.; Janssens, G.P.; Rochus, K.; Duchateau, L.; Scharek-Tedin, L.; Zentek, J.; Nguyen, P.; Cox, E.; Buyse, J.; Biourge, V.; et al. Proliferation capacity of T-lymphocytes is affected transiently after a long-term weight gain in Beagle dogs. Vet. Immunol. Immunopathol. 2013, 152, 237–244. [Google Scholar] [CrossRef]
- Frank, L.; Mann, S.; Levine, C.B.; Cummings, B.P.; Wakshlag, J.J. Increasing body condition score is positively associated interleukin-6 and monocyte chemoattractant protein-1 in Labrador retrievers. Vet. Immunol. Immunopathol. 2015, 167, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Piantedosi, D.; Di Loria, A.; Guccione, J.; De Rosa, A.; Fabbri, S.; Cortese, L.; Carta, S.; Ciaramella, P. Serum biochemistry profile, inflammatory cytokines, adipokines and cardiovascular findings in obese dogs. Vet. J. 2016, 216, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Wieland, C.W.; Florquin, S.; Chan, E.D.; Leemans, J.C.; Weijer, S.; Verbon, A.; Fantuzzi, G.; van der Poll, T. Pulmonary Mycobacterium tuberculosis infection in leptin deficient ob/ob mice. Int. Immunol. 2005, 17, 1399–1408. [Google Scholar] [CrossRef]
- Vedantama, G.; Viswanathana, V.K. Leptin signaling protects the gut from Entamoeba histolytica infection. Gut Microbes. 2012, 3, 1. [Google Scholar]
- Madan, R.; Guo, X.; Naylor, C.; Buonomo, E.L.; Mackay, D.; Noor, Z.; Concannon, P.; Scully, K.W.; Pramoonjago, P.; Kolling, G.L.; et al. Role of leptin-mediated colonic inflammation in defense against Clostridium difficile Colitis. Infect. Immun. 2014, 82, 341–349. [Google Scholar] [CrossRef]
- Shivahare, R.; Ali, W.; Vishwakarma, P.; Natu, S.M.; Puri, S.K.; Gupta, S. Leptin augments protective immune responses in murine macrophages and enhances potential of miltefosine against experimental visceral leishmaniasis. Acta. Tropica 2015, 150, 35–41. [Google Scholar] [CrossRef]
- Dayakar, A.; Chandrasekaran, S.; Veronica, J. Maurya, R.S. Leptin induces the phagocytosis and protective immune response in Leishmania donovani infected THP-1 cell line and human PBMCs. Exp. Parasitol. 2016, 160, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.; Graniel, J.; Ortiz, R. Effect of leptin on activation and cytokine synthesis in peripheral blood lymphocytes of malnourished infected children. Clin. Exp. Immunol. 2007, 148, 478–485. [Google Scholar] [CrossRef]
- Ehrlich, A.; Moreno Castilho, T.; Goldsmith-Pestana, K.; Chae, W.J.; Bothwell, A.L.; Sparwasser, T.; McMahon-Pratt, D. The immunotherapeutic role of regulatory T cells in Leishmania (Viannia) panamensis infection. J. Immunol. 2014, 193, 2961–2970. [Google Scholar] [CrossRef] [PubMed]
- Cortese, L.; Annunziatella, M.; Palatucci, A.T.; Rubino, V.; Piantedosi, D.; Di Loria, A.; Ruggiero, G.; Ciaramella, P.; Terrazzano, G. Regulatory T cells, Cytotoxic T lymphocytes and a T(H)1 cytokine profile in dogs naturally infected by Leishmania infantum. Res. Vet. Sci. 2013, 95, 942–949. [Google Scholar] [CrossRef]
- Adalid-Peralta, L.; Fragoso, G.; Fleury, A.; Sciutto, E. Mechanisms underlying the induction of regulatory T cells and its relevance in the adaptive immune response in parasitic infections. Int. J. Biol. Sci. 2011, 7, 1412–1426. [Google Scholar] [CrossRef]
- Di Loria, A.; Squillacioti, C.; De Luca, A.; Veneziano, V.; Mirabella, N.; Guccione, J.; Santoro, D. Increased leptin mRNA expression in the blood of dogs naturally infected by Leishmania infantum. Vet. J. 2014, 202, 634–636. [Google Scholar] [CrossRef]
- Chang, S.; Buzdar, A.U.; Hursting, S.D. Inflammatory breast cancer and body mass index. J. Clin. Oncol. 1998, 16, 3731–3735. [Google Scholar] [CrossRef]
- Marconato, L.; Romanelli, G.; Stefanello, D.; Giacoboni, C.; Bonfanti, U.; Bettini, G.; Finotello, R.; Verganti, S.; Valenti, P.; Ciaramella, L. Prognostic factors for dogs with mammary inflammatory carcinoma: 43 cases (2003–2008). J. Am. Vet. Med. Assoc. 2009, 235, 967–972. [Google Scholar] [CrossRef]
- Chan, D.S.; Vieira, A.R.; Aune, D.; Bandera, E.V.; Greenwood, D.C.; McTiernan, A.; Navarro Rosenblatt, D.; Thune, I.; Vieira, R.; Norat, T. Body mass index and survival in women with breast cancer-systematic literature review and meta-analysis of 82 follow-up studies. Ann. Oncol. 2014, 25, 1901–1914. [Google Scholar] [CrossRef]
- Lim, H.Y.; Im, K.S.; Kim, N.H.; Kim, H.W.; Shin, J.I.; Yhee, J.Y.; Sur, J.H. Effects of obesity and obesity-related molecules on canine mammary gland tumors. Vet. Pathol. 2015, 52, 1045–1051. [Google Scholar] [CrossRef]
- Lorincz, A.M.; Sukumar, S. Molecular links between obesity and breast cancer. Endocr. Relat. Cancer 2006, 13, 279–292. [Google Scholar] [CrossRef]
- Laud, K.; Gourdou, I.; Pessemesse, L.; Peyrat, J.; Djiane, J. Identification of leptin receptors in human breast cancer: Functional activity in the T47-D breast cancer cell line. Mol. Cell. Endocrinol. 2002, 188, 219–226. [Google Scholar] [CrossRef]
- Jardé, T.; Caldefie-Chezet, F.; Damez, M.; Mishellany, F.; Penault-Llorca, F.; Guillot, J.; Vasson, M.P. Leptin and leptin receptor involvement in cancer development: A study on human primary breast carcinoma. Oncol. Rep. 2008, 19, 905–912. [Google Scholar] [CrossRef]
- Jardé, T.; Perrier, S.; Vasson, M.P.; Caldefie-Chézet, F. Molecular mechanisms of leptin and adiponectin in breast cancer. Eur. J. Cancer. 2011, 47, 33–43. [Google Scholar] [CrossRef]
- Ray, A. Cancer and comorbidity: The role of leptin in breast cancer and associated pathologies. World J. Clin. Cases. 2018, 6, 483–492. [Google Scholar] [CrossRef]
- Rose, D.P.; Gilhooly, E.M.; Nixon, D.W. Adverse effects of obesity on breast cancer prognosis, and the biological actions of leptin. Int. J. Oncol. 2002, 21, 1285. [Google Scholar] [CrossRef]
- Ghanim, H.; Aljada, A.; Hofmeyer, D.; Syed, T.; Mohanty, P.; Dandona, P. Circulating mononuclear cells in the obese are in a proinflammatory state. Circulation 2004, 110, 1564–1571. [Google Scholar] [CrossRef]
- Nicolini, A.; Carpi, A.; Rossi, G. Cytokines in breast cancer. Cytokine Growth Factor Rev. 2006, 17, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Morris, P.G.; Hudis, C.A.; Giri, D.; Morrow, M.; Falcone, D.J.; Zhou, X.K.; Du, B.; Brogi, E.; Crawford, C.B.; Kopelovich, L. Inflammation and increased aromatase expression occur in the breast tissue of obese women with breast cancer. Cancer Prev. Res. 2011, 4, 1021–1029. [Google Scholar] [CrossRef]
- Arendt, L.M.; McCready, J.; Keller, P.J.; Baker, D.D.; Naber, S.P.; Seewaldt, V.; Kuperwasser, C. Obesity promotes breast cancer by CCL2-mediated macrophage recruitment and angiogenesis. Cancer Res. 2013, 73, 6080–6093. [Google Scholar] [CrossRef]
- Qian, B.Z.; Li, J.; Zhang, H.; Kitamura, T.; Zhang, J.; Campion, L.R.; Kaiser, E.A.; Snyder, L.A.; Pollard, J.W. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 2011, 475, 222–225. [Google Scholar] [CrossRef]
- Bhardwaj, P.; Du, B.; Zhou, X.K.; Sue, E.; Harbus, M.D.; Falcone, D.J.; Giri, D.; Hudis, C.A.; Kopelovich, L.; Subbaramaiah, K. Caloric restriction reverses obesity- induced mammary gland inflammation in mice. Cancer Prev. Res. 2013, 6, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Mehlman, E.; Bright, J.M.; Jeckel, K.; Porsche, C.; Veeramachaneni, D.N.; Frye, M. Echocardiographic evidence of left ventricular hypertrophy in obese dogs. J. Vet. Intern. Med. 2013, 27, 62–68. [Google Scholar] [CrossRef]
- Adolphe, J.L.; Silver, T.I.; Childs, H.; Drew, M.D.; Weber, L.P. Short-term obesity results in detrimental metabolic and cardiovascular changes that may not be reversed with weight loss in an obese dog model. Br. J. Nutr. 2014, 112, 647–656. [Google Scholar] [CrossRef]
- Tropf, M.; Nelson, O.L.; Lee, P.M.; Weng, H.Y. Cardiac and metabolic variables in obese dogs. J. Vet. Intern. Med. 2017, 31, 1000–1007. [Google Scholar] [CrossRef]
- Fonfara, S.; Hetzel, U.; Tew, S.R.; Dukes-McEwan, J.; Cripps, P.; Clegg, P.D. Leptin expression in dogs with cardiac disease and congestive heart failure. J. Vet. Intern. Med. 2011, 25, 1017–1024. [Google Scholar] [CrossRef]
- Kim, H.S.; Kang, J.H.; Jeung, E.B.; Yang, M.P. Serum concentrations of leptin and adiponectin in dogs with myxomatous mitral valve disease. J. Vet. Intern. Med. 2016, 30, 1589–1600. [Google Scholar] [CrossRef]
- Cuspidi, C.; Rescaldani, M.; Sala, C.; Grassi, G. Left-ventricular hypertrophy and obesity: A systematic review and meta-analysis of echocardiographic studies. J. Hypertens. 2014, 32, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Murdolo, G.; Angeli, F.; Reboldi, G.; Di Giacomo, L.; Aita, A.; Bartolini, C.; Vedecchia, P. Left ventricular hypertrophy and obesity: Only a matter of fat? High Blood Press. Cardiovasc. Prev. 2015, 22, 29–41. [Google Scholar] [CrossRef]
- Pérez-Sánchez, A.P.; Del-Angel-Caraza, J.; Quijano-Hernández, I.A.; Barbosa-Mireles, M.A. Obesity-hypertension and its relation to other diseases in dogs. Vet. Res. Commun. 2015, 39, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Verwaerde, P.; Sénard, J.M.; Galinier, M.; Rougé, P.; Massabuau, P.; Galitzky, J.; Berlan, M.; Lafontan, M.; Montastruc, J.L. Changes in short-term variability of blood pressure and heart rate during the development of obesity-associated hypertension in high-fat fed dogs. J. Hypertens. 1999, 17, 1135–1143. [Google Scholar] [CrossRef]
- Montoya, J.A.; Morris, P.J.; Bautista, I.; Juste, M.C.; Suarez, L.; Peña, C.; Hackett, R.M.; Rawlings, J. Hypertension: A risk factor associated with weight status in dogs. J. Nutr. 2006, 136, 2011S–2013S. [Google Scholar] [CrossRef] [PubMed]
- Schulze, P.C.; Kratzsch, J. Leptin as a new diagnostic tool in chronic heart failure. Clin. Chim. Acta 2005, 362, 1–11. [Google Scholar] [CrossRef]
- Purdham, D.M.; Zou, M.X.; Rajapurohitam, V. Rat heart is a site of leptin production and action. Am. J. Physiol. Heart. Circ. Physiol. 2004, 287, H2877–H2884. [Google Scholar] [CrossRef] [PubMed]
- Karmazyn, M.; Purdham, D.M.; Rajapurohitam, V.; Zeidan, A. Leptin as a cardiac hypertrophic factor: A potential target for therapeutics. Trends Cardiovasc. Med. 2007, 17, 206–211. [Google Scholar] [CrossRef]
- Sierra-Honigmann, M.R.; Nath, A.K.; Murakami, C.; García-Cardeña, G.; Papapetropoulos, A.; Sessa, W.C.; Madge, L.A.; Schechner, J.S.; Schwabb, M.B.; Polverini, P.J.; et al. Biological action of leptin as an angiogenic factor. Science 1998, 281, 1683–1686. [Google Scholar] [CrossRef]
- Sweeney, G. Cardiovascular effects of leptin. Nat. Rev. Cardiol. 2010, 7, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; McNeill, J.H. The emerging roles of leptin and ghrelin in cardiovascular physiology and pathophysiology. Curr. Vasc. Pharmacol. 2005, 3, 169–180. [Google Scholar] [CrossRef]
- Cheng, K.H.; Chu, C.S.; Lee, K.T.; Lin, T.H.; Hsieh, C.C.; Chiu, C.C.; Voon, W.C.; Sheu, S.H.; Lai, W.T. Adipocytokines and proinflammatory mediators from abdominal and epicardial adipose tissue in patients with coronary artery disease. Int. J. Obes. 2008, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Knudson, J.D.; Payne, G.A.; Borbouse, L.; Tune, J.D. Leptin and mechanisms of endothelial dysfunction and cardiovascular disease. Curr. Hypertens. Rep. 2008, 10, 434–439. [Google Scholar] [CrossRef]
- Anker, S.D.; Chua, T.P.; Ponikowski, P.; Hrrington, D.; Swan, J.W.; Kox, W.J.; Poole-Wilson, P.A.; Coats, A.J. Hormonal changes and catabolic/anabolic imbalance in chronic heart failure and their importance for cardiac cachexia. Circulation 1997, 96, 526–534. [Google Scholar] [CrossRef]
- Doehner, W.; Pflaum, C.D.; Rauchhaus, M.; Godsland, I.F.; Egerer, K.; Cicoira, M.; Florea, V.G.; Sharma, R.; Bolger, A.P.; Coats, A.J.; et al. Leptin, insulin sensitivity and growth hormone binding protein in chronic heart failure with and without cardiac cachexia. Eur. J. Endocrinol. 2001, 145, 727–735. [Google Scholar] [CrossRef][Green Version]
- Haynes, W.G.; Morgan, D.A.; Walsh, S.A.; Mark, A.L.; Sivitz, W.I. Receptor- mediated regional sympathetic nerve activation by leptin. J. Clin. Investig. 1997, 100, 270–278. [Google Scholar] [CrossRef]
- Atkinson, L.L.; Fischer, M.A.; Lopaschuk, G.D. Leptin activates cardiac fatty acid oxidation independent of changes in the AMP-activated protein kinase-acetyl-CoA carboxylase-malonyl- CoA axis. J. Biol. Chem. 2002, 277, 29424–29430. [Google Scholar] [CrossRef]
- Schram, K.; De Girolamo, S.; Madani, S.; Munoz, D.; Thong, F.; Sweeney, G. Leptin regulates MMP-2, TIMP-1 and collagen synthesis via p38 MAPK in HL-1 murine cardiomyocytes. Cell Mol. Biol. Lett. 2010, 15, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Barouch, L.A.; Gao, D.; Chen, L.; Miller, K.L.; Xu, W.; Phan, A.C.; Kittleson, M.M.; Minhas, K.M.; Berkowitz, D.E.; Wei, C.; et al. Cardiac myocyte apoptosis is associated with increased DNA damage and decreased survival in murine models of obesity. Circ. Res. 2006, 98, 119–124. [Google Scholar] [CrossRef] [PubMed]
- McGaffin, K.R.; Witham, W.G.; Yester, K.A.; Romano, L.C.; O’Doherty, R.M.; McTiernan, C.F.; O’Donnell, C.P. Cardiac-specific leptin receptor deletion exacerbates ischaemic heart failure in mice. Cardiovasc. Res. 2011, 89, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Mazaki-Tovi, M.; Feuermann, Y.; Segev, G.; Klement, E.; Yas-Natan, E.; Farkas, A.; Kol, A.; Shamay, A. Increased serum leptin and insulin concentrations in canine hypothyroidism. Vet. J. 2010, 183, 109–114. [Google Scholar] [CrossRef]
- Greene, V.R.; Wilson, H.; Pfent, C.; Roethele, J.; Carwile, J.; Qin, Y.; Grimm, E.; Ellerhorst, J.A. Expression of leptin and iNOS in oral melanomas in dogs. J. Vet. Intern. Med. 2013, 27, 1278–1282. [Google Scholar] [CrossRef]
- Schaffler, A.; Landfried, K.; Volk, M.; Fürst, A.; Büchler, C.; Schölmerich, J.; Herfarth, H. Potential of adipocytokines in predicting peripancreatic necrosis and severity in acute pancreatitis: Pilot study. J. Gastroenterol. Hepatol. 2007, 22, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Muddana, V.; Lamb, J.; Greer, J.; Papachristou, G.I.; Whitcomb, D.C. Low serum adiponectin levels are associated with systemic organ failure in acute pancreatitis. Pancreas 2009, 38, 907–912. [Google Scholar] [CrossRef]
- Schaffler, A.; Hamer, O.; Dickopf, J.; Goetz, A.; Landfried, K.; Voelk, M.; Herfarth, H.; Kopp, A.; Büchler, C.; Schölmerich, J.; et al. Admission resistin levels predict peripancreatic necrosis and clinical severity in acute pancreatitis. Am. J. Gastroenterol. 2010, 105, 2474–2484. [Google Scholar] [CrossRef]
- Schaffler, A.; Hamer, O.W.; Dickopf, J.; Goetz, A.; Landfried, K.; Voelk, M.; Herfarth, H.; Kopp, A.; Buechler, C.; Schölmerich, J.; et al. Admission visfatin levels predict pancreatic and peripancreatic necrosis in acute pancreatitis and correlate with clinical severity. Am. J. Gastroenterol. 2011, 106, 957–967. [Google Scholar] [CrossRef]
- Kerem, M.; Bedirli, A.; Pasaoglu, H.; Unsal, C.; Yilmaz, T.U.; Ofluoglu, E.; Sahin, T.T. Role of ghrelin and leptin in predicting the severity of acute pancreatitis. Dig. Dis. Sci. 2007, 52, 950–955. [Google Scholar] [CrossRef]
- Zyromski, N.J.; Mathur, A.; Pitt, H.A.; Lu, D.; Gripe, J.T.; Walker, J.J.; Yancey, K.; Wade, T.E.; Swartz-Basile, D.A. A murine model of obesity implicates the adipokine milieu in the pathogenesis of severe acute pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G552–G558. [Google Scholar] [CrossRef]
- Franco-Pons, N.; Gea-Sorlì, S.; Closa, D. Release of inflammatory mediators by adipose tissue during acute pancreatitis. J. Pathol. 2010, 221, 175–182. [Google Scholar] [CrossRef]
- Paek, J.; Kang, J.H.; Kim, H.S.; Lee, I.; Seo, K.W.; Yang, M.P. Serum adipokine concentrations in dogs with acute pancreatitis. J. Vet. Intern. Med. 2014, 28, 1760–1769. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, A.; Kweon, O.K.; Kim, W.H. Presence and distribution of leptin and leptin receptor in the canine gallbladder. Acta Histochem. 2016, 118, 674–678. [Google Scholar] [CrossRef][Green Version]
- Lee, S.; Kweon, O.K.; Kim, W.H. Increased leptin and leptin receptor expression in dogs with gallbladder mucocele. J. Vet. Intern. Med. 2017, 31, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Haluzík, M.; Fiedler, J.; Nedvidkova, J.; Ceska, R. Serum leptin concentrations in patients with combined hyperlipidemia: Relationship to serum lipids and lipoproteins. Physiol. Res. 1998, 48, 363–368. [Google Scholar]
- Haluzík, M.; Fiedler, J.; Nedvídkova, J.; Ceska, R. Serum leptin levels in patients with hyperlipidemias. Nutrition 2000, 16, 429–433. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, S.E.; Oh, J.H.; Seo, K.W.; Song, K.H. Leptin, adiponectin and serotonin levels in lean and obese dogs. BMC Vet. Res. 2014, 10, 113. [Google Scholar] [CrossRef] [PubMed]
- Gonzaga, N.C.; Medeiros, C.C.; de Carvalho, D.F.; Alves, J.G. Leptin and cardiometabolic risk factors in obese children and adolescents. J. Paediatr. Child Health 2014, 50, 707–712. [Google Scholar] [CrossRef]
- Cuevas, A.; Miquel, J.F.; Reyes, M.S.; Zanlungo, S.; Nervi, F. Diet as a risk factor for cholesterol gallstone disease. J. Am. Coll. Nutr. 2004, 23, 187–196. [Google Scholar] [CrossRef]
- Kim, J.M.; Lee, H.L.; Moon, W.; Koh, D.H.; Lee, O.Y.; Yoon, B.C.; Choi, H.S.; Hahm, J.S.; Lee, M.H.; Lee, D.H.; et al. Association between insulin, insulin resistance, and gallstone disease in Korean general population. Korean J. Gastroenterol. 2007, 50, 183–187. [Google Scholar]
- Lee, S.; Kweon, O.K.; Kim, W.H. Associations between serum leptin levels, hyperlipidemia, and cholelithiasis in dogs. PLoS ONE 2017, 12, e0187315. [Google Scholar] [CrossRef]
- Hardie, L.; Trayhurn, P.; Abramovich, D.; Fowler, P. Circulating leptin in women: A longitudinal study in the menstrual cycle and during pregnancy. Clin. Endocrinol. 1997, 47, 101–106. [Google Scholar] [CrossRef]
- Amico, J.A.; Thomas, A.; Crowley, R.S.; Burmeister, L.A. Concentrations of leptin in the serum of pregnant, lactating, and cycling rats and of leptin messenger ribonucleic acid in rat placental tissue. Life Sci. 1998, 63, 1387–1395. [Google Scholar] [CrossRef]
- Balogh, O.; Staub, L.P.; Gram, A.; Boos, A.; Kowalewski, M.P.; Reichler, I.M. Leptin in the canine uterus and placenta: Possible implications in pregnancy. Reprod. Biol. Endocrinol. 2015, 13, 13. [Google Scholar] [CrossRef]
- Cojocaru, M.; Cojocaru, I.M.; Silosi, I.; Rogoz, S. Role of Leptin in Autoimmune Diseases. Maedica J. Clin. Med. 2013, 8, 75–79. [Google Scholar]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortese, L.; Terrazzano, G.; Pelagalli, A. Leptin and Immunological Profile in Obesity and Its Associated Diseases in Dogs. Int. J. Mol. Sci. 2019, 20, 2392. https://doi.org/10.3390/ijms20102392
Cortese L, Terrazzano G, Pelagalli A. Leptin and Immunological Profile in Obesity and Its Associated Diseases in Dogs. International Journal of Molecular Sciences. 2019; 20(10):2392. https://doi.org/10.3390/ijms20102392
Chicago/Turabian StyleCortese, Laura, Giuseppe Terrazzano, and Alessandra Pelagalli. 2019. "Leptin and Immunological Profile in Obesity and Its Associated Diseases in Dogs" International Journal of Molecular Sciences 20, no. 10: 2392. https://doi.org/10.3390/ijms20102392
APA StyleCortese, L., Terrazzano, G., & Pelagalli, A. (2019). Leptin and Immunological Profile in Obesity and Its Associated Diseases in Dogs. International Journal of Molecular Sciences, 20(10), 2392. https://doi.org/10.3390/ijms20102392