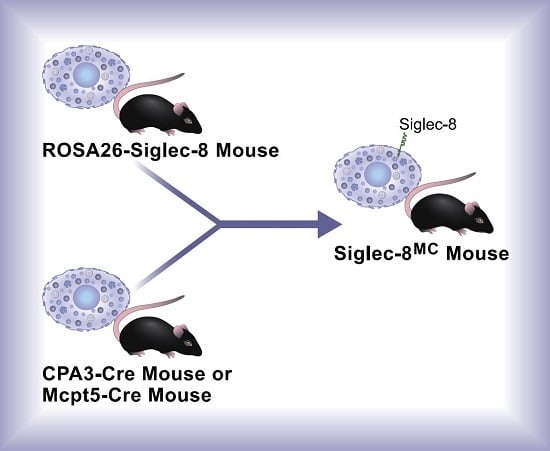
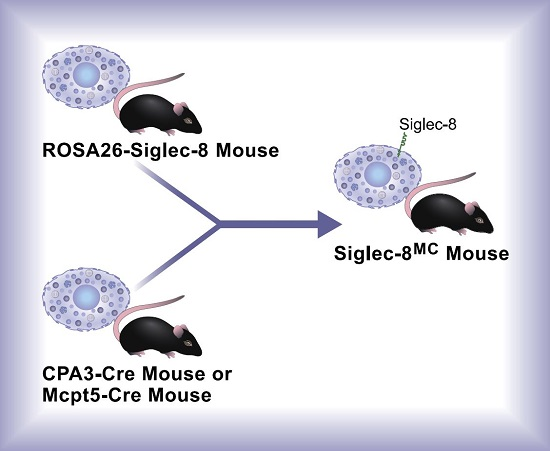
Mast Cell-Specific Expression of Human Siglec-8 in Conditional Knock-in Mice

 , , and
, , and
Abstract
1. Introduction
2. Results
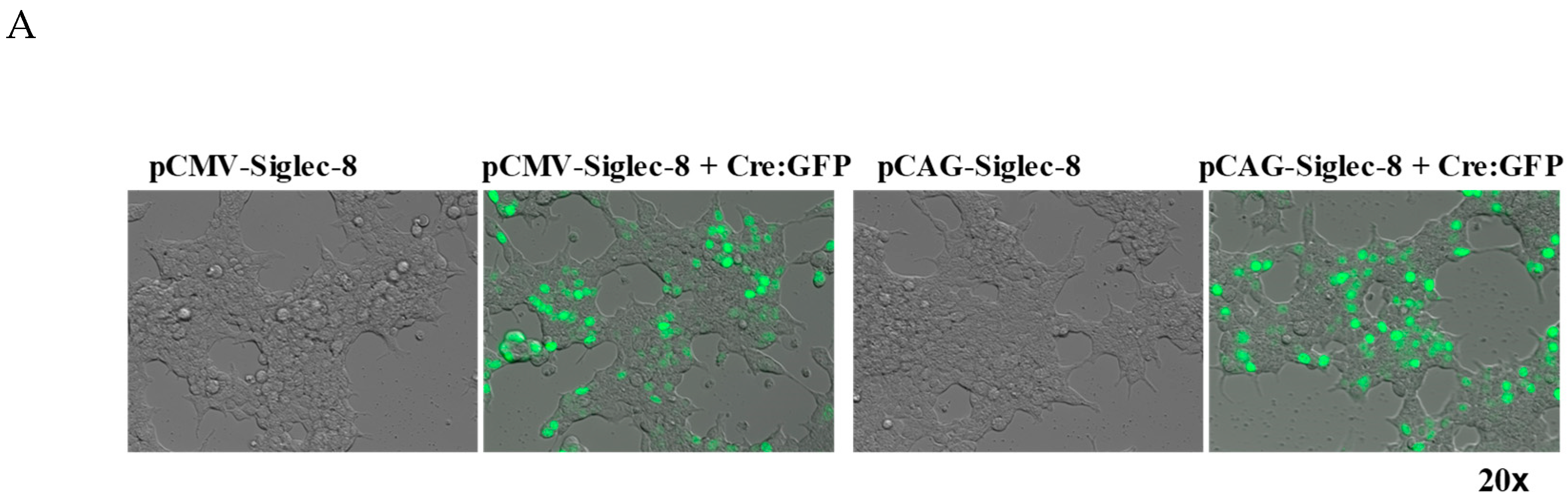
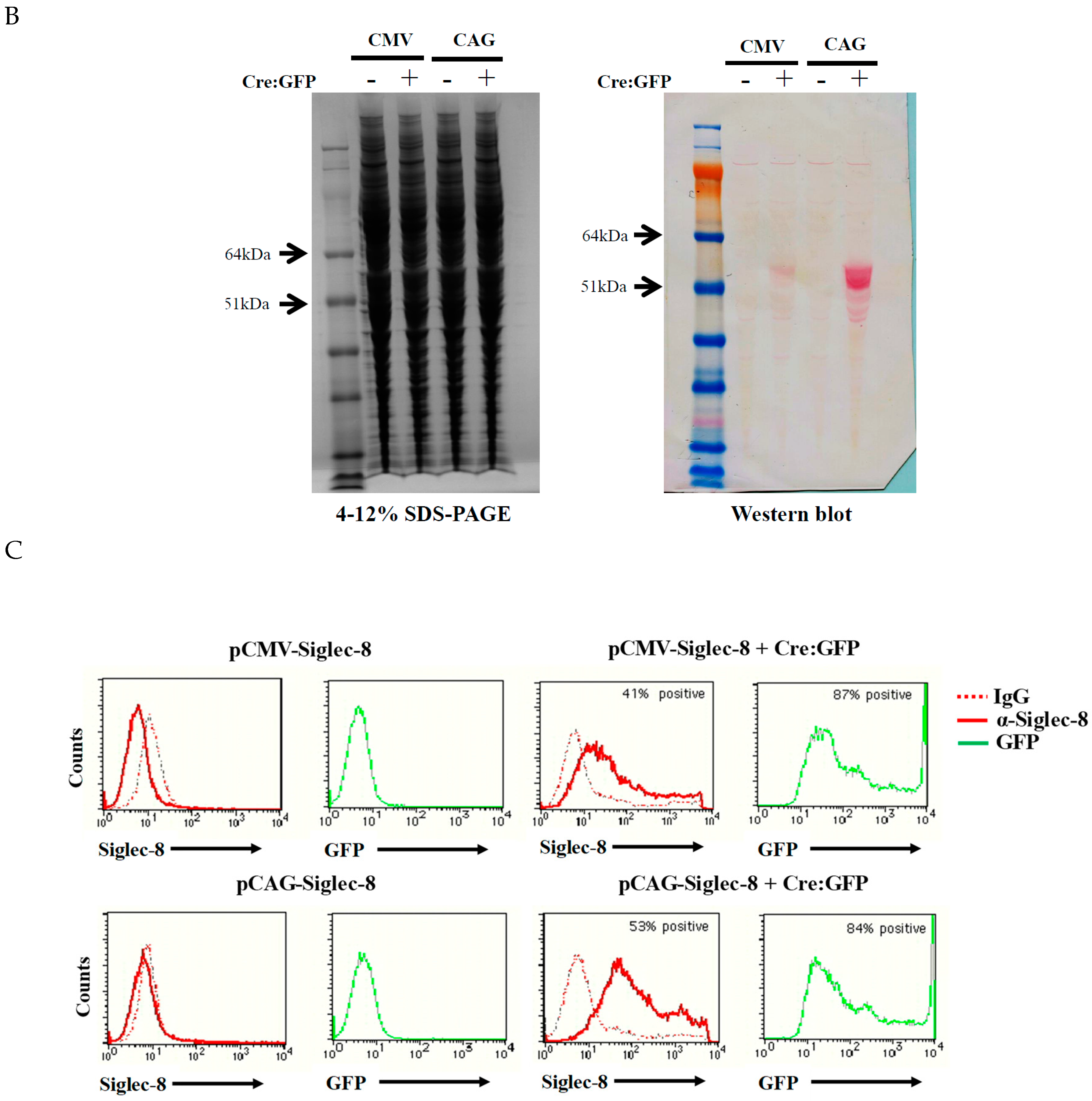
2.1. In Vitro Testing of ROSA26-Siglec-8 Constructs in HEK 293T Cells
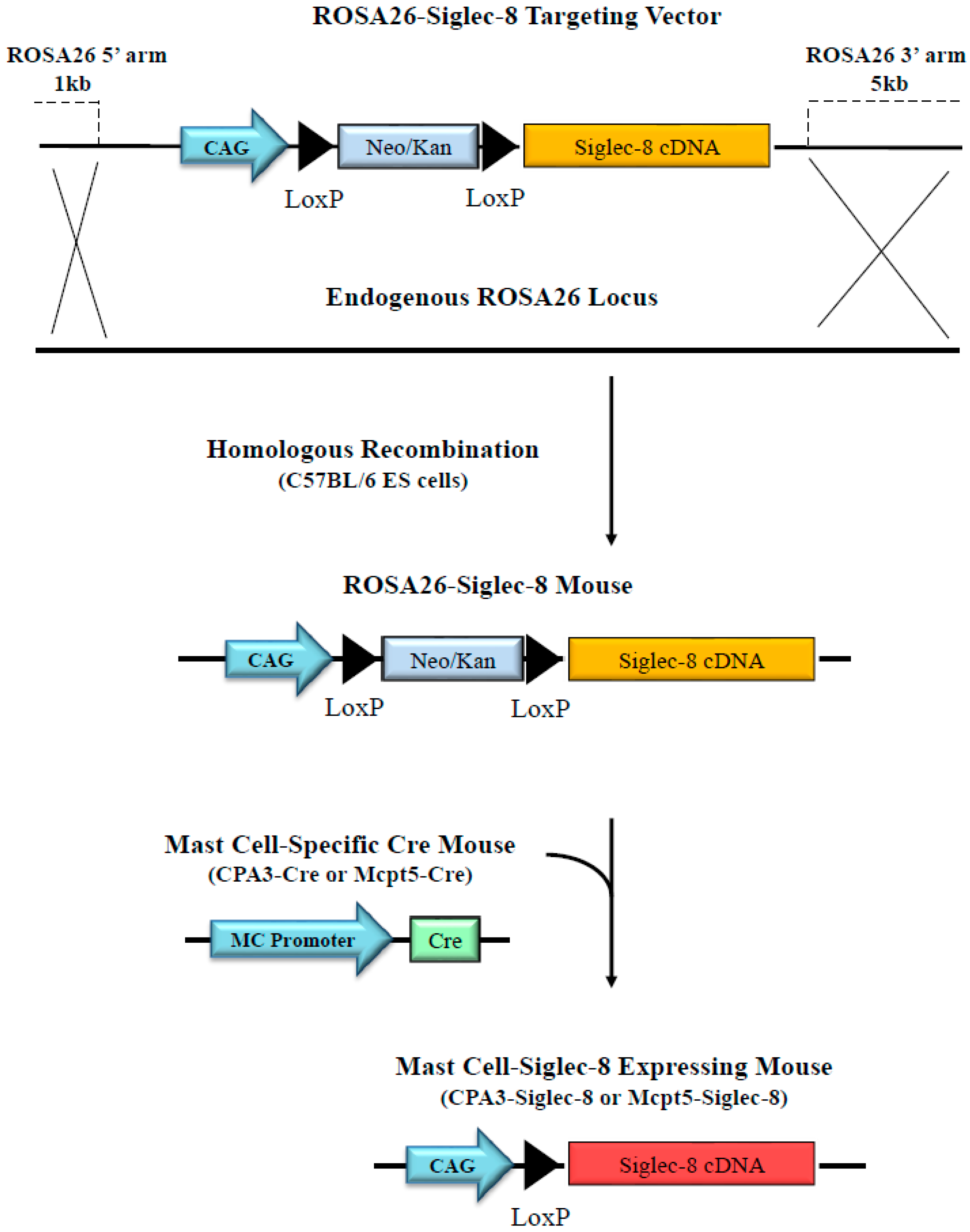
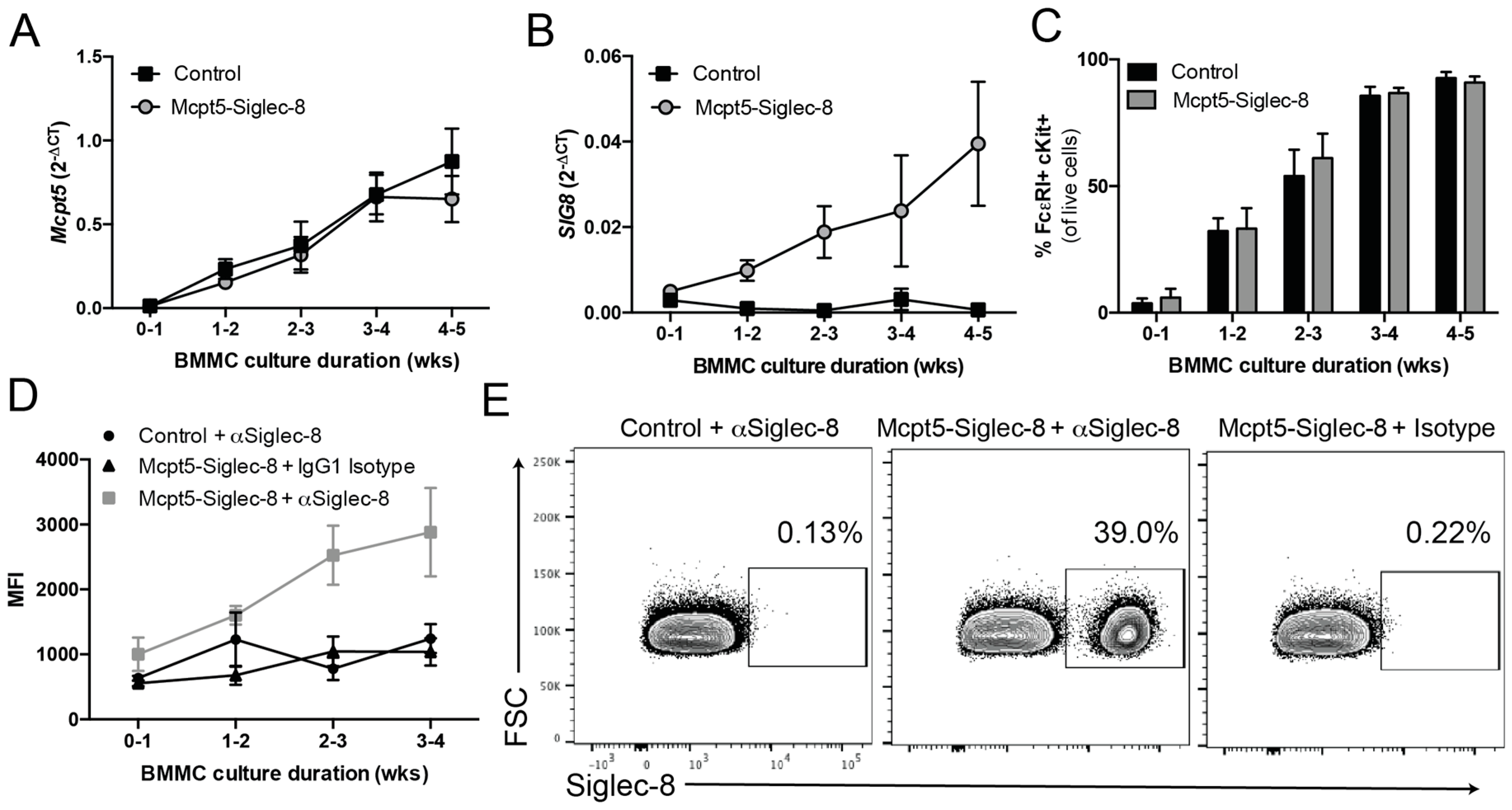
2.2. Generation of Conditional Knock-in ROSA26-Siglec-8 Mice and Targeting of Siglec-8 Expression to Mast Cells
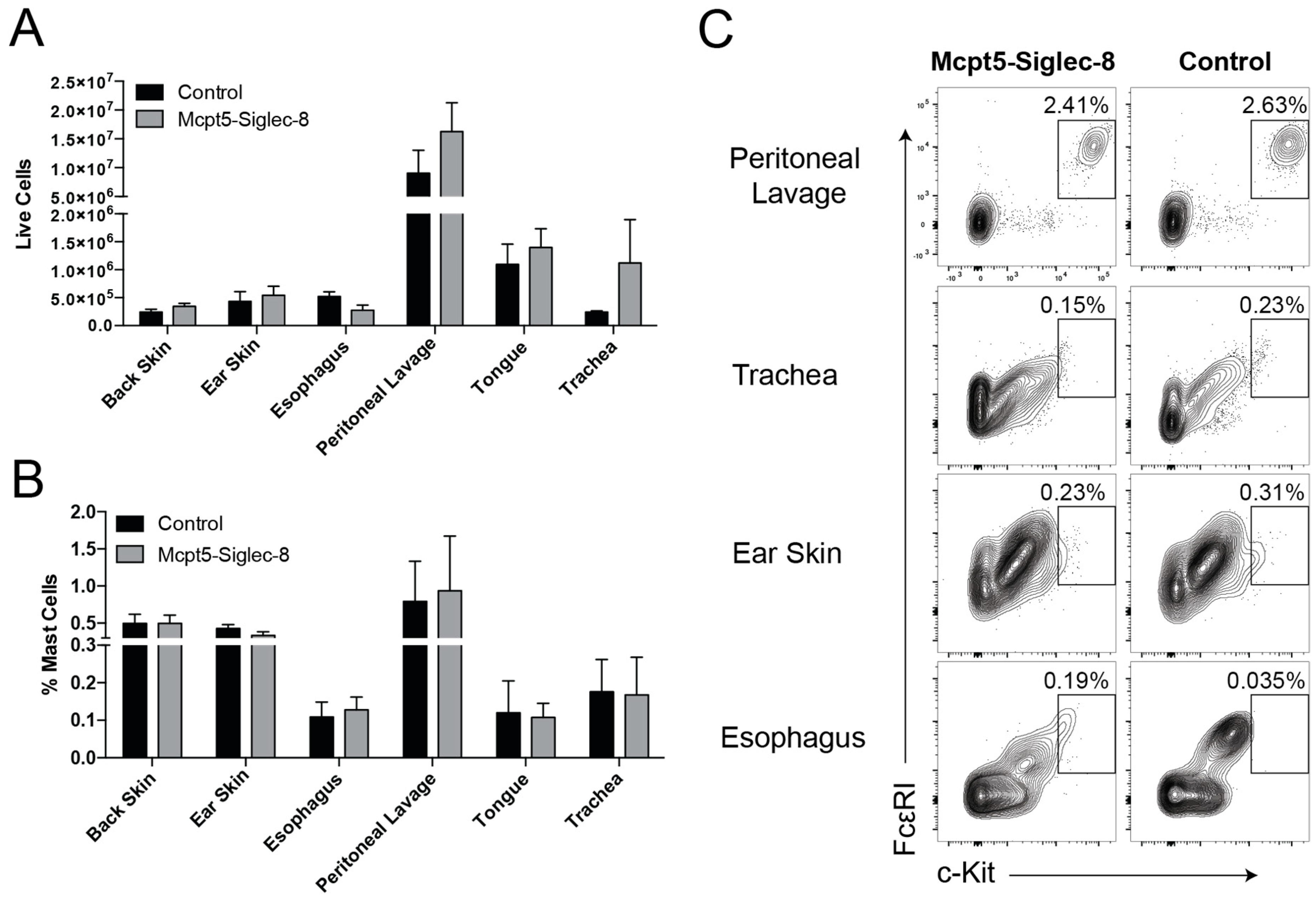
2.3. Equal Numbers of Mast Cells in Tissues of Mcpt5-Siglec-8 and Control Mice
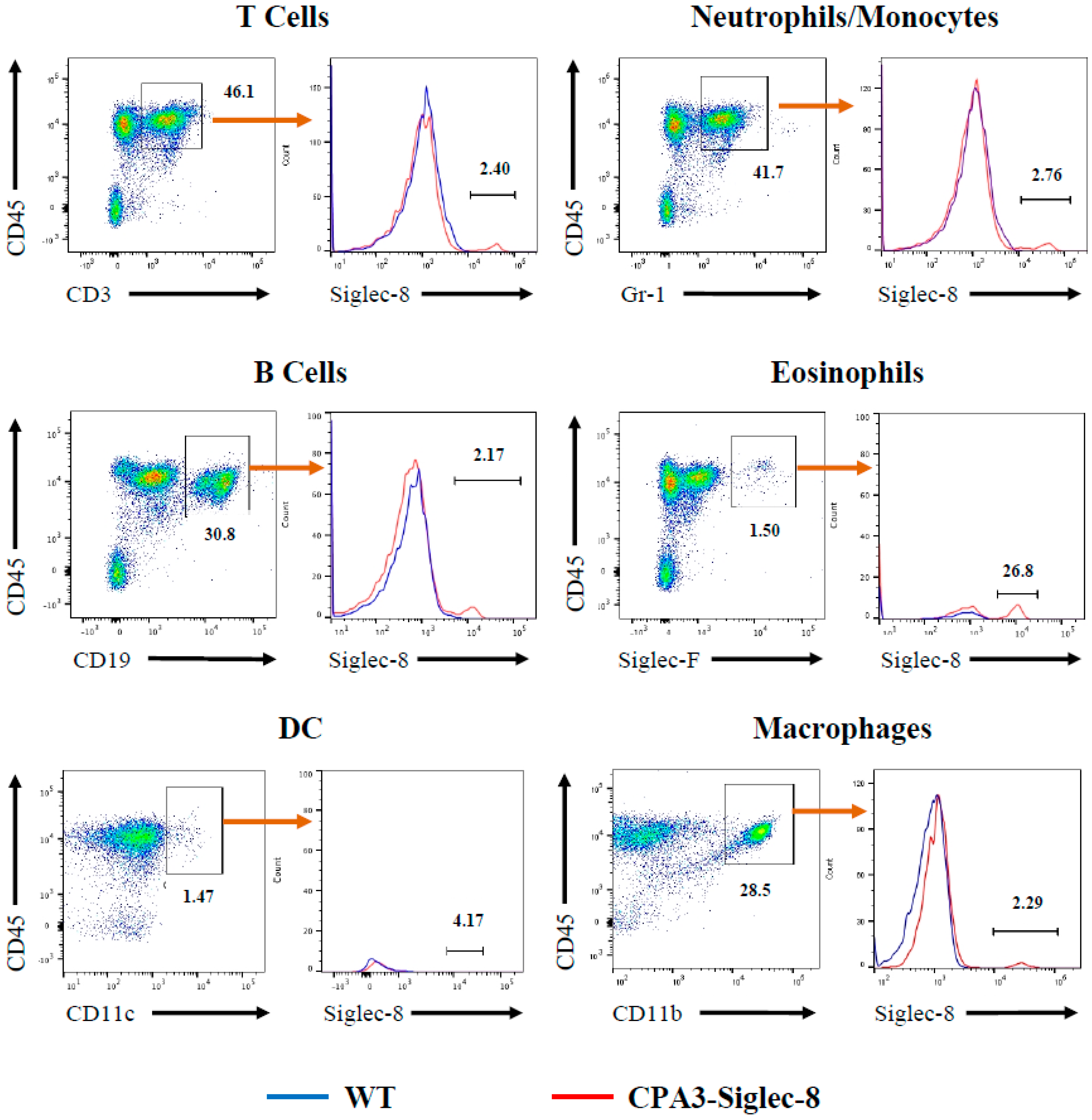
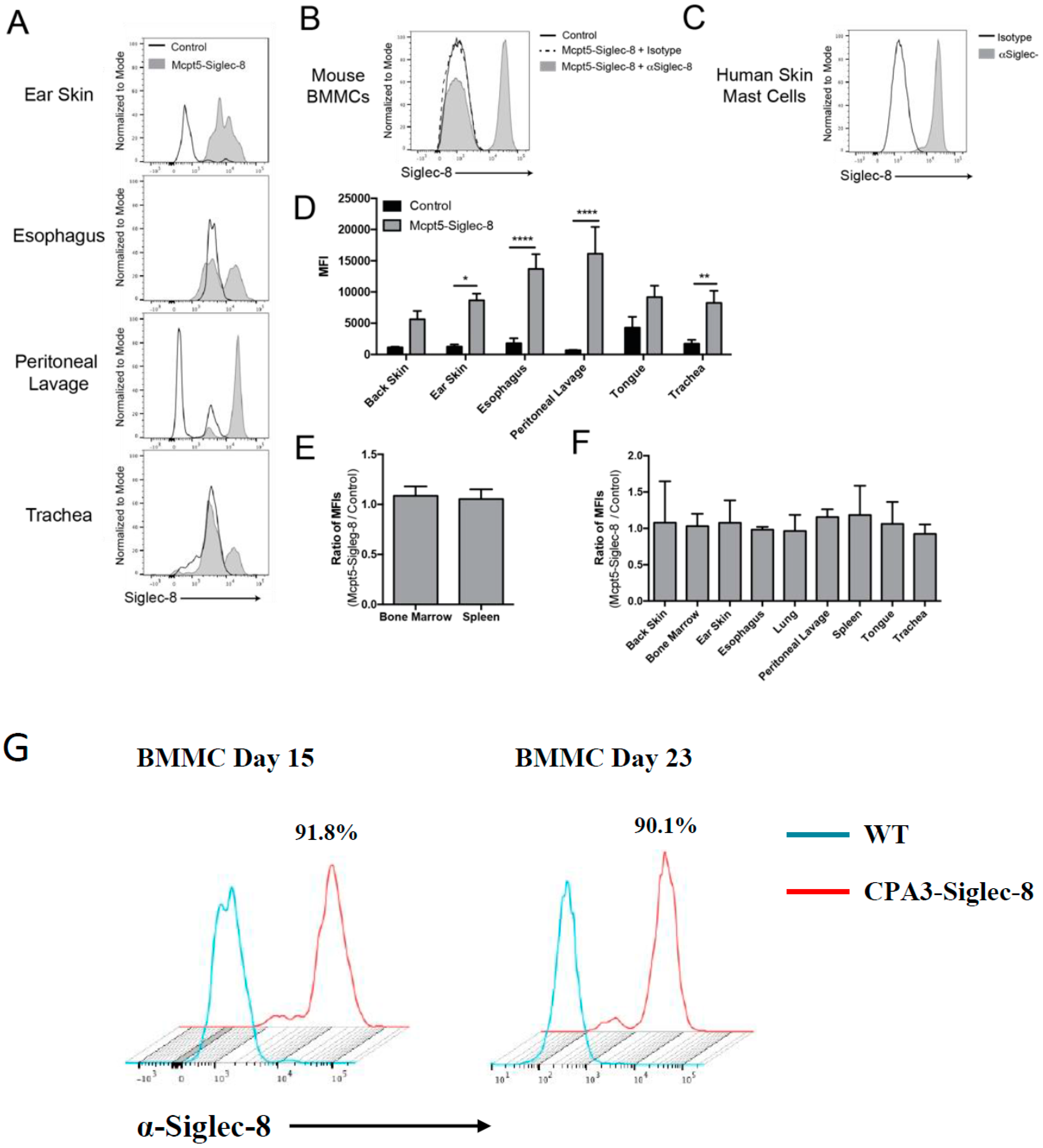
2.4. Expression of Siglec-8 on Mast Cells and Basophils in CPA3-Siglec-8 Mice on Mast Cells in Mcpt5-Siglec-8 Mice
2.5. Expression of Siglec-8 on Mast cells and Its Tissue Distribution
3. Discussion
4. Materials and Methods
4.1. Generation of a ROSA26-Siglec-8 Construct and Transfection of the ROSA26-Siglec-8 Construct into HEK 293T Cells
4.2. Generation of Human Siglec-8 Conditional Knock-in Mice
4.3. Targeting Human Siglec-8 Expression to Mouse Mast Cells
4.4. Genotyping of ROSA26-Siglec-8 KI Mice and Cre Transgenic Mice
4.5. RT-PCR Analysis of Mcpt5 and Siglec-8 mRNA Expression in BMMCs
4.6. Antibodies and Flow Cytometric Analysis of Siglec-8 Expression in Cells and Tissues
4.7. Bone Marrow-Derived Mast Cell (BMMC) Culture
4.8. Tissue Digestion for Mouse Mast Cell Enumeration
4.9. Human Skin Mast Cell Isolation and Culture
4.10. Western Blot
4.11. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BMMC | Bone marrow derived mast cell |
| CAG promoter | A hybrid promoter consisting of CMV enhancer, chicken β-actin promoter and rabbit β-globin splice acceptor |
| CPA3 | Carboxypeptidase A3 |
| Cre | Cre recombinase |
| ES cells | Embryonic stem cells |
| GFP | Green fluorescence protein |
| KI | Knock-in |
| Mcpt5 | Mast cell protease 5 |
| pCAG-Neo-ROSA26-Siglec-8 | ROSA26-Siglec-8 construct with the CAG promoter |
| pCMV-Neo-ROSA26-Siglec-8 | ROSA26-Siglec-8 construct with the CMV promoter |
References
- Floyd, H.; Ni, J.; Cornish, A.L.; Zeng, Z.; Liu, D.; Carter, K.C.; Steel, J.; Crocker, P.R. Siglec-8. A novel eosinophil-specific member of the immunoglobulin superfamily. J. Biol. Chem. 2000, 275, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Kikly, K.K.; Bochner, B.S.; Freeman, S.D.; Tan, K.B.; Gallagher, K.T.; D’alessio, K.J.; Holmes, S.D.; Abrahamson, J.A.; Erickson-Miller, C.L.; Murdock, P.R. Identification of SAF-2, a novel siglec expressed on eosinophils, mast cells, and basophils. J. Allergy Clin. Immunol. 2000, 105, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Bochner, B.S. “Siglec”ting the allergic response for therapeutic targeting. Glycobiology 2016, 26, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.W.; Kelly, E.A.; Nguyen, C.L.; Jarjour, N.N.; Bochner, B.S. Characterization of Siglec-8 Expression on Lavage Cells after Segmental Lung Allergen Challenge. Int. Arch. Allergy Immunol. 2018, 177, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.S.; Shimizu, K.; Grant, A.V.; Rafaels, N.; Zhou, L.F.; Hudson, S.A.; Konno, S.; Zimmermann, N.; Araujo, M.I.; Ponte, E.V.; et al. Polymorphisms in the sialic acid-binding immunoglobulin-like lectin-8 (Siglec-8) gene are associated with susceptibility to asthma. Eur. J. Hum. Genet. 2010, 18, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Nutku, E.; Aizawa, H.; Hudson, S.A.; Bochner, B.S. Ligation of Siglec-8: A selective mechanism for induction of human eosinophil apoptosis. Blood 2003, 101, 5014–5020. [Google Scholar] [CrossRef] [PubMed]
- Hudson, S.A.; Bovin, N.V.; Schnaar, R.L.; Crocker, P.R.; Bochner, B.S. Eosinophil-selective binding and proapoptotic effect in vitro of a synthetic Siglec-8 ligand, polymeric 6′-sulfated sialyl Lewis x. J. Pharmacol. Exp. Ther. 2009, 330, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, H.; Choi, O.H.; Hubbard, W.W.; Lee, H.S.; Canning, B.J.; Lee, H.H.; Ryu, S.D.; von Gunten, S.S.; Bickel, C.A.; Hudson, S.A.; et al. Inhibition of FcepsilonRI-dependent mediator release and calcium flux from human mast cells by sialic acid-binding immunoglobulin-like lectin 8 engagement. J. Allergy Clin. Immunol. 2008, 121, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Carroll, D.J.; O’Sullivan, J.A.; Nix, D.B.; Cao, Y.Y.; Tiemeyer, M.M.; Bochner, B.S. Sialic acid-binding immunoglobulin-like lectin 8 (Siglec-8) is an activating receptor mediating beta2-integrin-dependent function in human eosinophils. J. Allergy Clin. Immunol. 2018, 141, 2196–2207. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.A.; Carroll, D.J.; Cao, Y.; Salicru, A.N.; Bochner, B.S. Leveraging Siglec-8 endocytic mechanisms to kill human eosinophils and malignant mast cells. J. Allergy Clin. Immunol. 2018, 141, 1774–1785. [Google Scholar] [CrossRef]
- Aizawa, H.; Zimmermann, N.N.; Carrigan, P.E.; Lee, J.J.; Rothenberg, M.E.; Bochner, B.S. Molecular analysis of human Siglec-8 orthologs relevant to mouse eosinophils: Identification of mouse orthologs of Siglec-5 (mSiglec-F) and Siglec-10 (mSiglec-G). Genomics 2003, 82, 521–530. [Google Scholar] [CrossRef]
- Tateno, H.; Crocker, P.R.; Paulson, J.C. Mouse Siglec-F and human Siglec-8 are functionally convergent paralogs that are selectively expressed on eosinophils and recognize 6′-sulfo-sialyl Lewis X as a preferred glycan ligand. Glycobiology 2005, 15, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Kano, G.G.; Hudson, S.A.; Brummet, M.M.; Zimmermann, N.N.; Zhu, Z.Z.; Bochner, B.S. Mechanisms of Siglec-F-induced eosinophil apoptosis: A role for caspases but not for SHP-1, Src kinases, NADPH oxidase or reactive oxygen. PLoS ONE 2013, 8, e68143. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, D.D. Mast cells and mastocytosis. Blood 2008, 112, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Akin, C. Mast cell activation syndromes. J. Allergy Clin. Immunol. 2017, 140, 349–355. [Google Scholar] [CrossRef]
- Church, M.K.; Kolkhir, P.; Metz, M.; Maurer, M. The role and relevance of mast cells in urticaria. Immunol. Rev. 2018, 282, 232–247. [Google Scholar] [CrossRef]
- Yokoi, H.; Myers, A.A.; Matsumoto, K.K.; Crocker, P.R.; Saito, H.H.; Bochner, B.S. Alteration and acquisition of Siglecs during in vitro maturation of CD34+ progenitors into human mast cells. Allergy 2006, 61, 769–776. [Google Scholar] [CrossRef]
- Hudson, S.A.; Herrmann, H.H.; Du, J.J.; Cox, P.P.; EL Haddad, B.; Butler, B.B.; Crocker, P.R.; Ackerman, S.J.; Valent, P.P.; Bochner, B.S. Developmental, malignancy-related, and cross-species analysis of eosinophil, mast cell, and basophil siglec-8 expression. J. Clin. Immunol. 2011, 31, 1045–1053. [Google Scholar] [CrossRef]
- O’Sullivan, J.A.; Wei, Y.Y.; Carroll, D.J.; Moreno-Vinasco, L.L.; Cao, Y.Y.; Zhang, F.F.; Lee, J.J.; Zhu, Z.Z.; Bochner, B.S. Characterization of a novel mouse strain expressing human Siglec-8 only on eosinophils. J. Leukoc. Biol. 2018, 104, 11–19. [Google Scholar] [CrossRef]
- Foussias, G.; Yousef, G.M.; Diamandis, E.P. Molecular characterization of a Siglec8 variant containing cytoplasmic tyrosine-based motifs, and mapping of the Siglec8 gene. Biochem. Biophys. Res. Commun. 2000, 278, 775–781. [Google Scholar] [CrossRef]
- Lilla, J.N.; Chen, C.C.; Mukai, K.K.; BenBarak, M.J.; Franco, C.B.; Kalesnikoff, J.J.; Yu, M.; Tsai, M.; Piliponsky, A.M.; Galli, S.J. Reduced mast cell and basophil numbers and function in Cpa3-Cre; Mcl-1fl/fl mice. Blood 2011, 118, 6930–6938. [Google Scholar] [CrossRef] [PubMed]
- Peschke, K.; Dudeck, A.; Rabenhorst, A.; Hartmann, K.; Roers, A. Cre/loxP-based mouse models of mast cell deficiency and mast cell-specific gene inactivation. Methods Mol. Biol. 2015, 1220, 403–421. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Brandal, S.; Kapur, R.; Zhu, Z.; Takemoto, C.M. Global microRNA expression is essential for murine mast cell development in vivo. Exp. Hematol. 2014, 42, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Scholten, J.; Hartmann, K.; Gerbaulet, A.; Krieg, T.; Müller, W.; Testa, G.; Roers, A. Mast cell-specific Cre/loxP-mediated recombination in vivo. Transgenic Res. 2008, 17, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Hohenstein, P.; Slight, J.; Ozdemir, D.D.; Burn, S.F.; Berry, R.; Hastie, N.D. High-efficiency Rosa26 knock-in vector construction for Cre-regulated overexpression and RNAi. Pathogenetics 2008, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, T.; Cepko, C.L. Controlled expression of transgenes introduced by in vivo electroporation. Proc. Natl. Acad. Sci. USA 2007, 104, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, D.F.; Barrett, N.A.; Austen, K.F.; Immunological Genome Project Consortium. Expression profiling of constitutive mast cells reveals a unique identity within the immune system. Nat. Immunol. 2016, 17, 878–887. [Google Scholar] [CrossRef]
- Kambe, N.; Kambe, M.; Kochan, J.P.; Schwartz, L.B. Human skin-derived mast cells can proliferate while retaining their characteristic functional and protease phenotypes. Blood 2001, 97, 2045–2052. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Clone | Fluorophore Conjugation | Source |
|---|---|---|---|
| Anti-Siglec-8 (mouse IgG1) | 2E2 | Alexa Fluor 488 | Produced in house |
| Mouse IgG1 Isotype | MOPC-21 | Unconjugated, Alexa Fluor 647 or Alexa Fluor 488 | Tonbo Biosciences, San Diego, CA, USA Conjugated to Alexa Fluor 488 in house |
| Anti-CD3 | 500A2 | Pacific Blue | Invitrogen |
| Anti-CD11b | M1/70 | APC-Cy7 | BD Pharmingen |
| Anti-CD11c | N418 | APC | Biolegend |
| Anti-CD19 | 1D3 | PE | BD Biosciences |
| Anti-CD45 | 1O4 | PE-Cy7 | BD Pharmingen |
| Anti-CD45 | 30-F11 | APC-Cy7 | BD Pharmingen |
| Anti-CD49b | DX5 | PE | BD Pharmingen |
| Anti-CD117 (c-Kit) | 2B8 | BV421 | BD Pharmingen |
| Anti-CD117 (c-Kit) | 2B8 | APC | eBioscience |
| Anti-CD117 (c-Kit) | 2B8 | FITC | eBioscience |
| Anti-FcεRI | MAR-1 | PE-Cy7 | Biolegend |
| Anti-FcεRI | MAR-1 | PE-Cy7 | eBioscience |
| Anti-FcεRI | MAR-1 | APC | Biolegend |
| Anti-Ly-6G and Ly-6C (Gr-1) | RB6-8C5 | Pacific Blue | Invitrogen |
| Anti-Ly-6G and Ly-6C (Gr-1) | RB6-8C5 | eFluor450 | eBioscience |
| Anti-Siglec-F | E50-2440 | PE | BD Pharmingen |
| Anti-Siglec-8 (mouse IgG1) | 2C4 | Alexa Fluor 647 | Produced in house |
| Anti-Siglec-8 (mouse IgG1) | 7C9 | PE | Biolegend |
| Anti-Siglec-8 (mouse IgG1) | 7C9 | APC | Biolegend |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Y.; Chhiba, K.D.; Zhang, F.; Ye, X.; Wang, L.; Zhang, L.; Robida, P.A.; Moreno-Vinasco, L.; Schnaar, R.L.; Roers, A.; et al. Mast Cell-Specific Expression of Human Siglec-8 in Conditional Knock-in Mice. Int. J. Mol. Sci. 2019, 20, 19. https://doi.org/10.3390/ijms20010019
Wei Y, Chhiba KD, Zhang F, Ye X, Wang L, Zhang L, Robida PA, Moreno-Vinasco L, Schnaar RL, Roers A, et al. Mast Cell-Specific Expression of Human Siglec-8 in Conditional Knock-in Mice. International Journal of Molecular Sciences. 2019; 20(1):19. https://doi.org/10.3390/ijms20010019
Chicago/Turabian StyleWei, Yadong, Krishan D. Chhiba, Fengrui Zhang, Xujun Ye, Lihui Wang, Li Zhang, Piper A. Robida, Liliana Moreno-Vinasco, Ronald L. Schnaar, Axel Roers, and et al. 2019. "Mast Cell-Specific Expression of Human Siglec-8 in Conditional Knock-in Mice" International Journal of Molecular Sciences 20, no. 1: 19. https://doi.org/10.3390/ijms20010019
APA StyleWei, Y., Chhiba, K. D., Zhang, F., Ye, X., Wang, L., Zhang, L., Robida, P. A., Moreno-Vinasco, L., Schnaar, R. L., Roers, A., Hartmann, K., Lee, C.-M., Demers, D., Zheng, T., Bochner, B. S., & Zhu, Z. (2019). Mast Cell-Specific Expression of Human Siglec-8 in Conditional Knock-in Mice. International Journal of Molecular Sciences, 20(1), 19. https://doi.org/10.3390/ijms20010019