The Influence of Capsaicin on the Integrity of Microvascular Endothelial Cell Monolayers

 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Quantified Impact of Vanilloids on the Viability of cEND Cells Using MTT Assays
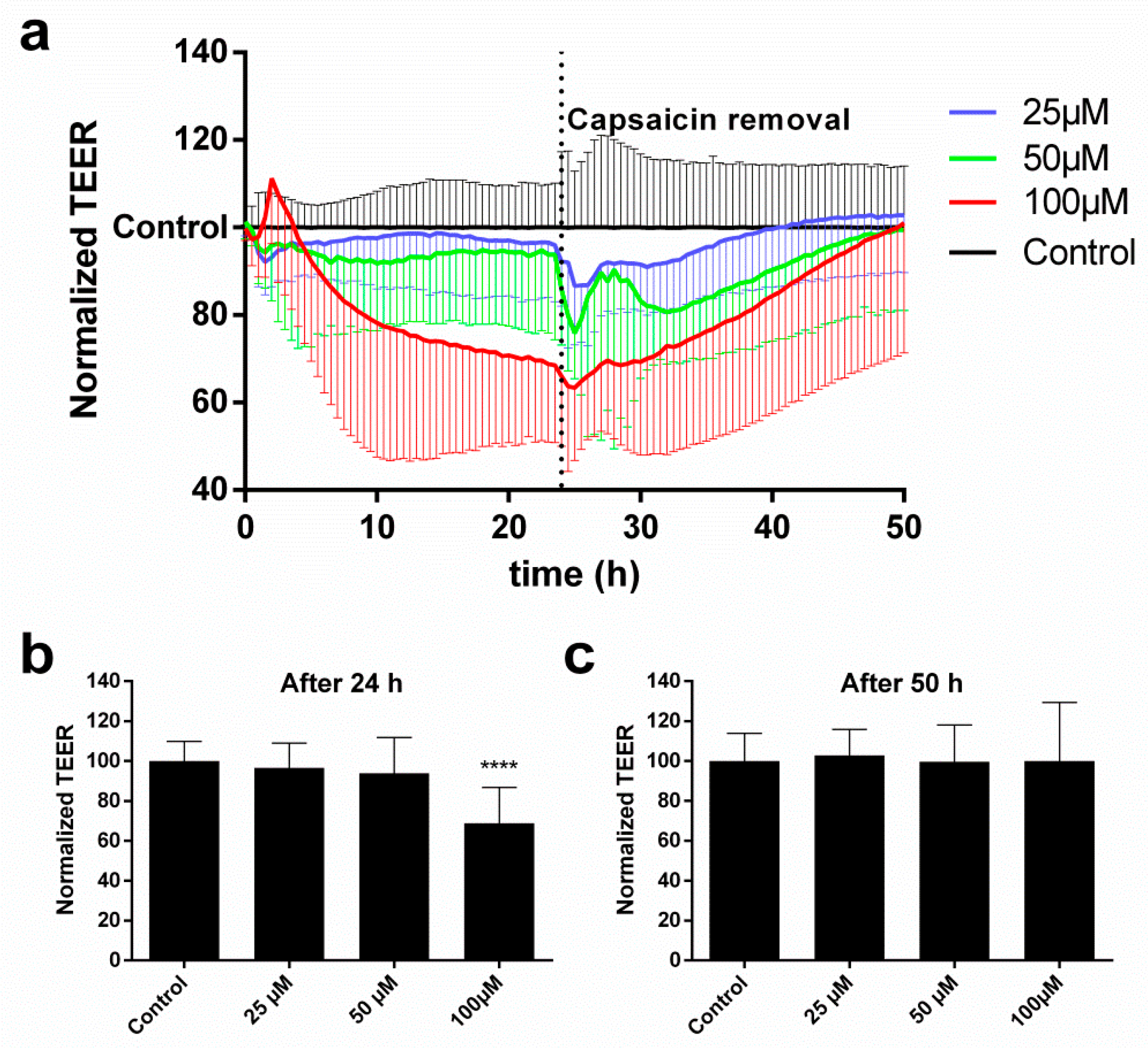
2.2. cEND Cells Showed Reversible Tight Junction Opening after Temporal Vanilloid Treatment
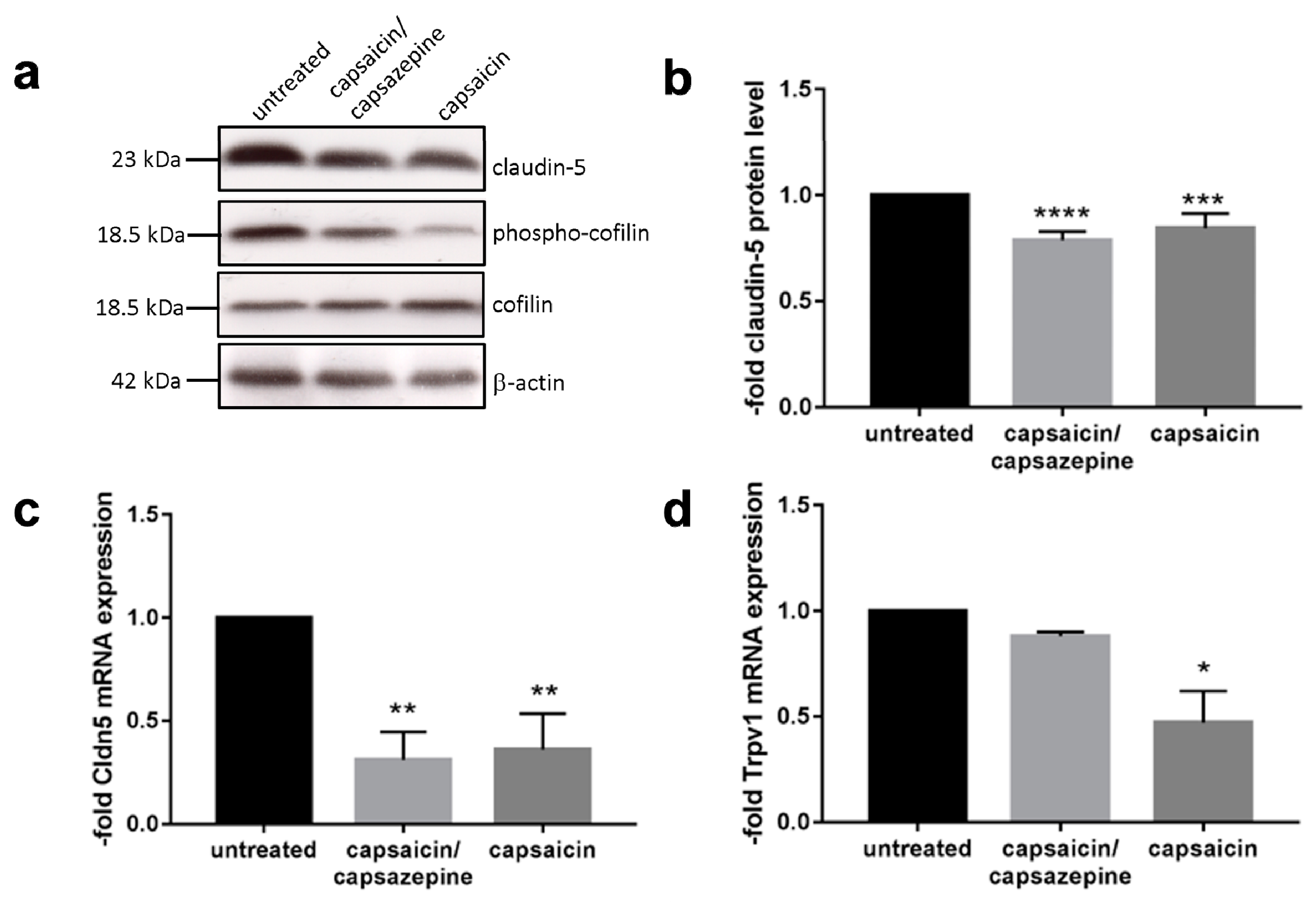
2.3. Capsaicin Decreased Expression Levels of Claudin 5 and Induced Dephosphorylation of Cofilin
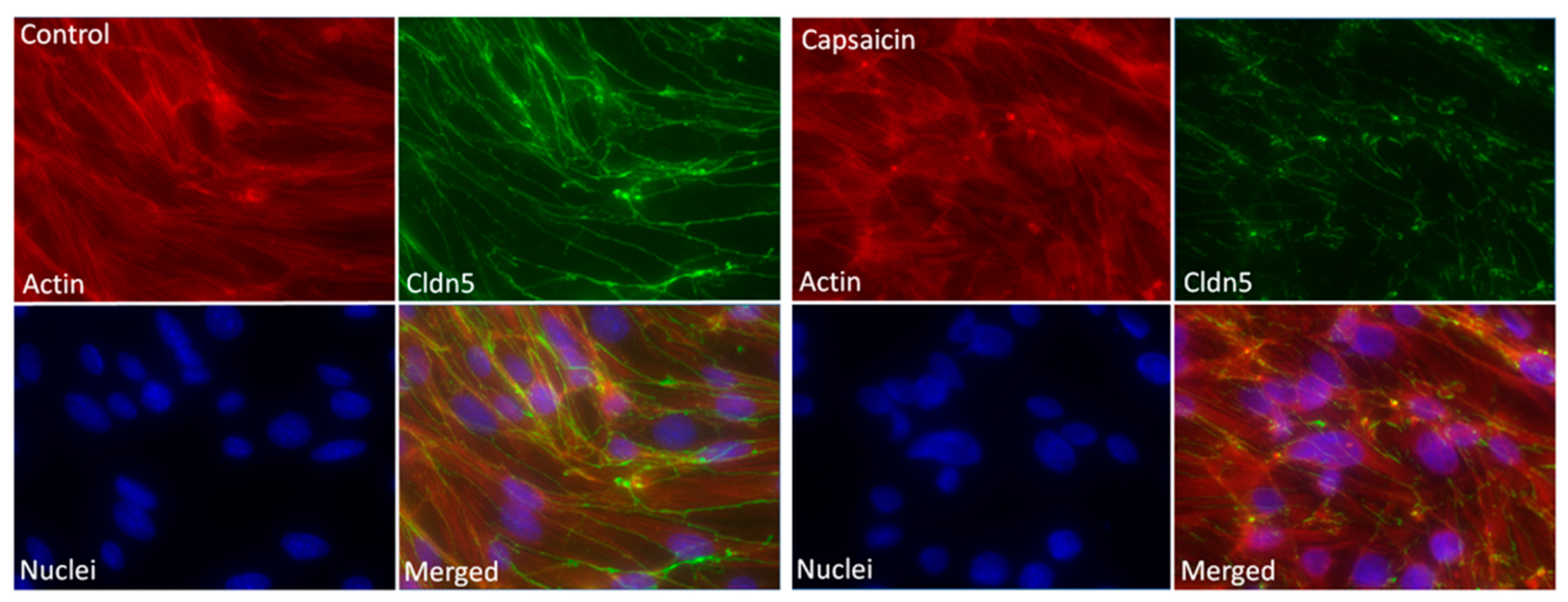
2.4. Fluorescence and Digital Holographic Microscopy Revealed Morphological Changes of Endothelial Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) Assay
4.3. Electrical Impedance Measurements
4.4. Structured Illumination Fluorescence Microscopy (SIFM)
4.5. Western Blot
4.6. Real-Time Quantitative PCR
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BBB | Blood-brain barrier |
| CCL | capacitance |
| cEND | brain microvascular endothelial cells |
| Cldn5 | claudin 5 |
| DHM | digital holographic microscopy |
| ECM | endothelial cell medium |
| IK | incubation buffer |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| TEER | transendothelial electric resistance |
| TRITC | tetramethylrhodamine |
| SIFM | structured illumination fluorescence microscopy |
| ZO-1 | zonula occludens 1 |
References
- Korel, F.; Bagdatlioglu, N.; Balaban, M.Ö.; Hisil, Y. Ground Red Peppers: Capsaicinoids Content, Scoville Scores, and Discrimination by an Electronic Nose. J. Agric. Food Chem. 2002, 50, 3257–3261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Liu, D.Y.; Ma, L.Q.; Luo, Z.D.; Cao, T.B.; Zhong, J.; Yan, Z.C.; Wang, L.J.; Zhao, Z.G.; Zhu, S.J. Activation of Transient Receptor Potential Vanilloid Type-1 Channel Prevents Adipogenesis and Obesity. Circ. Res. 2007, 100, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.; Brooks, J.R. Capsaicin-Based Therapies for Pain Control. In Capsaicin as a Therapeutic Molecule; Abdel-Salam, O.M.E., Ed.; Springer: Basel, Switzerland, 2014; pp. 129–146. [Google Scholar]
- Shiobara, T.; Usui, T.; Han, J.; Isoda, H.; Nagumo, Y. The Reversible Increase in Tight Junction Permeability Induced by Capsaicin is Mediated Via Cofilin-Actin Cytoskeletal Dynamics and Decreased Level of Occludin. PLoS ONE 2013, 8, e79954. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, M.; Goycoolea, F.M. Vanilloids and Their Effect on Mammalian Biological Barriers. In Capsaicin: Food Sources, Medical Uses and Health Implications; Gilliam, B., Ed.; Nova Science Publishers: Hapac, NY, USA, 2014; pp. 53–74. [Google Scholar]
- Kaiser, M.; Pereira, S.; Pohl, L.; Ketelhut, S.; Kemper, B.; Gorzelanny, C.; Galla, H.; Moerschbacher, B.M.; Goycoolea, F.M. Chitosan Encapsulation Modulates the Effect of Capsaicin on the Tight Junctions of MDCK Cells. Sci. Rep. 2015, 5, 10048. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, M.; Kirsch, B.; Hauser, H.; Schneider, D.; Seuß-Baum, I.; Goycoolea, F.M. In Vitro and Sensory Evaluation of Capsaicin-Loaded Nanoformulations. PLoS ONE 2015, 10, e0141017. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, M.; Chalapala, S.; Gorzelanny, C.; Perali, R.S.; Goycoolea, F.M. The Effect of Capsaicin Derivatives on Tight-Junction Integrity and Permeability of MDCK Cells. J. Pharm. Sci. 2016, 105, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, L. Modern Methods for Delivery of Drugs Across the Blood–brain Barrier. Adv. Drug Deliv. Rev. 2012, 64, 640–665. [Google Scholar] [CrossRef] [PubMed]
- Beggs, S.; Liu, X.J.; Kwan, C.; Salter, M.W. Peripheral Nerve Injury and TRPV1-Expressing Primary Afferent C-Fibers Cause Opening of the Blood-Brain Barrier. Mol. Pain 2010, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Easton, A.; Fraser, P. TRPV1 Activation Results in Disruption of the Blood-Brain Barrier in the Rat. Br. J. Pharmacol. 2005, 146, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Burek, M.; Salvador, E.; Förster, C.Y. Generation of an Immortalized Murine Brain Microvascular Endothelial Cell Line as an in Vitro Blood Brain Barrier Model. J. Vis. Exp. 2012, 66, 4022. [Google Scholar] [CrossRef] [PubMed]
- Forster, C.; Silwedel, C.; Golenhofen, N.; Burek, M.; Kietz, S.; Mankertz, J.; Drenckhahn, D. Occludin as Direct Target for Glucocorticoid-Induced Improvement of Blood-Brain Barrier Properties in a Murine in Vitro System. J. Physiol. Lond. 2005, 565, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Kim, J.; Kim, S.Y.; Jang, S.M.; Lee, S.M.; Choi, I.H.; Park, S.W.; Shin, J.S.; Yoo, K.H. Capacitance-Based Assay for Real-Time Monitoring of Endocytosis and Cell Viability. Miniat. Chem. Biol. 2012, 12, 2377–2384. [Google Scholar] [CrossRef] [PubMed]
- Wegener, J.; Keese, C.R.; Giaever, I. Electric Cell-Substrate Impedance Sensing (ECIS) as a Noninvasive Means to Monitor the Kinetics of Cell Spreading to Artificial Surfaces. Exp. Cell Res. 2000, 259, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Nagumo, Y.; Han, J.; Bellila, A.; Isoda, H.; Tanaka, T. Cofilin Mediates Tight-Junction Opening by Redistributing Actin and Tight-Junction Proteins. Biochem. Biophys. Res. Commun. 2008, 377, 921–925. [Google Scholar]
- Kaiser, M.; Lankamp, F.; Goycoolea, F.M. Nanoencapsulation of Capsaicin Attenuates the Cytotoxic Effect on Caco-2 Cells. In Gums and Stabilisers for the Food Industry 18: Hydrocolloid Functionality for Affordable and Sustainable Global Food Solutions; Williams, P., Phillips, G., Eds.; The Royal Society of Chemistry: London, UK, 2016; pp. 176–181. [Google Scholar]
- Kaiser, M.; Pohl, L.; Ketelhut, S.; Kastl, L.; Gorzelanny, C.; Götte, M.; Schnekenburger, J.; Goycoolea, F.M.; Kemper, B. Nanoencapsulated Capsaicin Changes Migration Behavior and Morphology of Madin Darby Canine Kidney Cell Monolayers. PLoS ONE 2017, 12, e0187497. [Google Scholar] [CrossRef] [PubMed]
- Dilling, C.; Roewer, N.; Forster, C.Y.; Burek, M. Multiple Protocadherins are Expressed in Brain Microvascular Endothelial Cells and might Play a Role in Tight Junction Protein Regulation. J. Cerebr. Blood Flow Metabol. 2017, 37, 3391–3400. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaiser, M.; Burek, M.; Britz, S.; Lankamp, F.; Ketelhut, S.; Kemper, B.; Förster, C.; Gorzelanny, C.; Goycoolea, F.M. The Influence of Capsaicin on the Integrity of Microvascular Endothelial Cell Monolayers. Int. J. Mol. Sci. 2019, 20, 122. https://doi.org/10.3390/ijms20010122
Kaiser M, Burek M, Britz S, Lankamp F, Ketelhut S, Kemper B, Förster C, Gorzelanny C, Goycoolea FM. The Influence of Capsaicin on the Integrity of Microvascular Endothelial Cell Monolayers. International Journal of Molecular Sciences. 2019; 20(1):122. https://doi.org/10.3390/ijms20010122
Chicago/Turabian StyleKaiser, Mathias, Malgorzata Burek, Stefan Britz, Frauke Lankamp, Steffi Ketelhut, Björn Kemper, Carola Förster, Christian Gorzelanny, and Francisco M. Goycoolea. 2019. "The Influence of Capsaicin on the Integrity of Microvascular Endothelial Cell Monolayers" International Journal of Molecular Sciences 20, no. 1: 122. https://doi.org/10.3390/ijms20010122
APA StyleKaiser, M., Burek, M., Britz, S., Lankamp, F., Ketelhut, S., Kemper, B., Förster, C., Gorzelanny, C., & Goycoolea, F. M. (2019). The Influence of Capsaicin on the Integrity of Microvascular Endothelial Cell Monolayers. International Journal of Molecular Sciences, 20(1), 122. https://doi.org/10.3390/ijms20010122