The First Dose of Fingolimod Affects Circulating Extracellular Vesicles in Multiple Sclerosis Patients

 , , , , ,
, , , , ,
Abstract

1. Introduction
2. Results
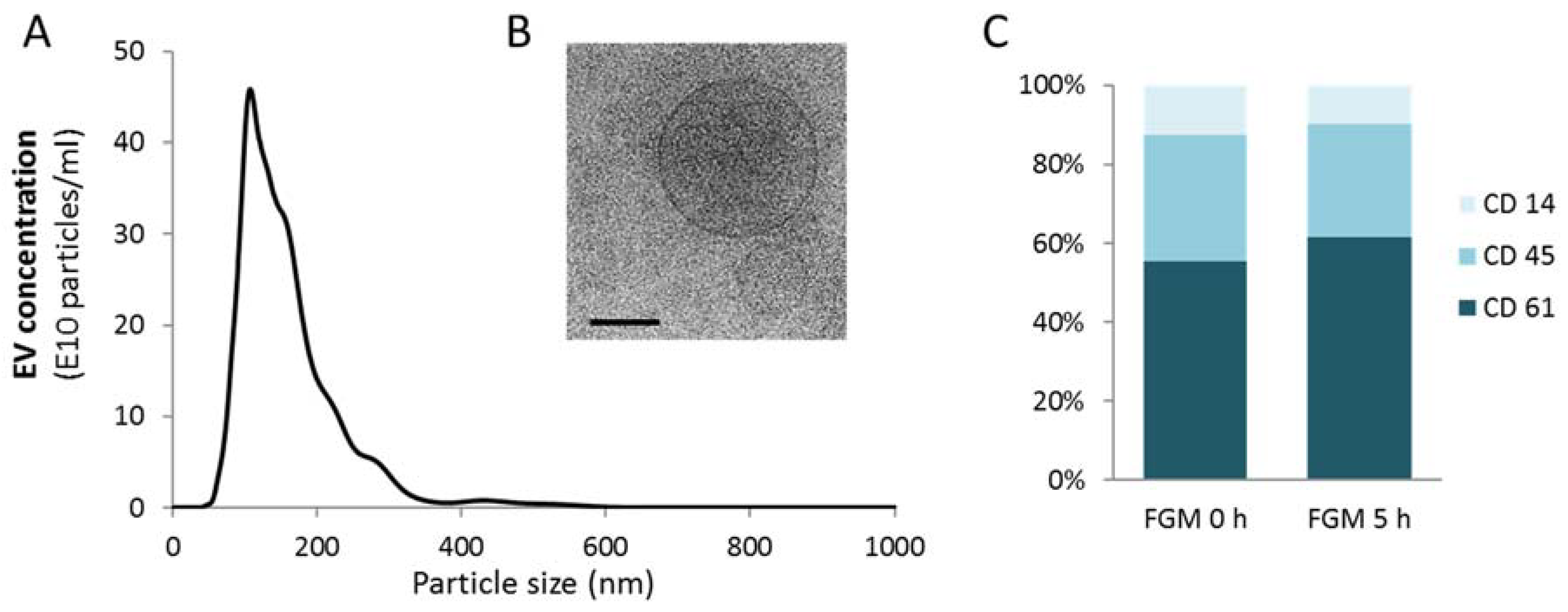
2.1. EV Characterization
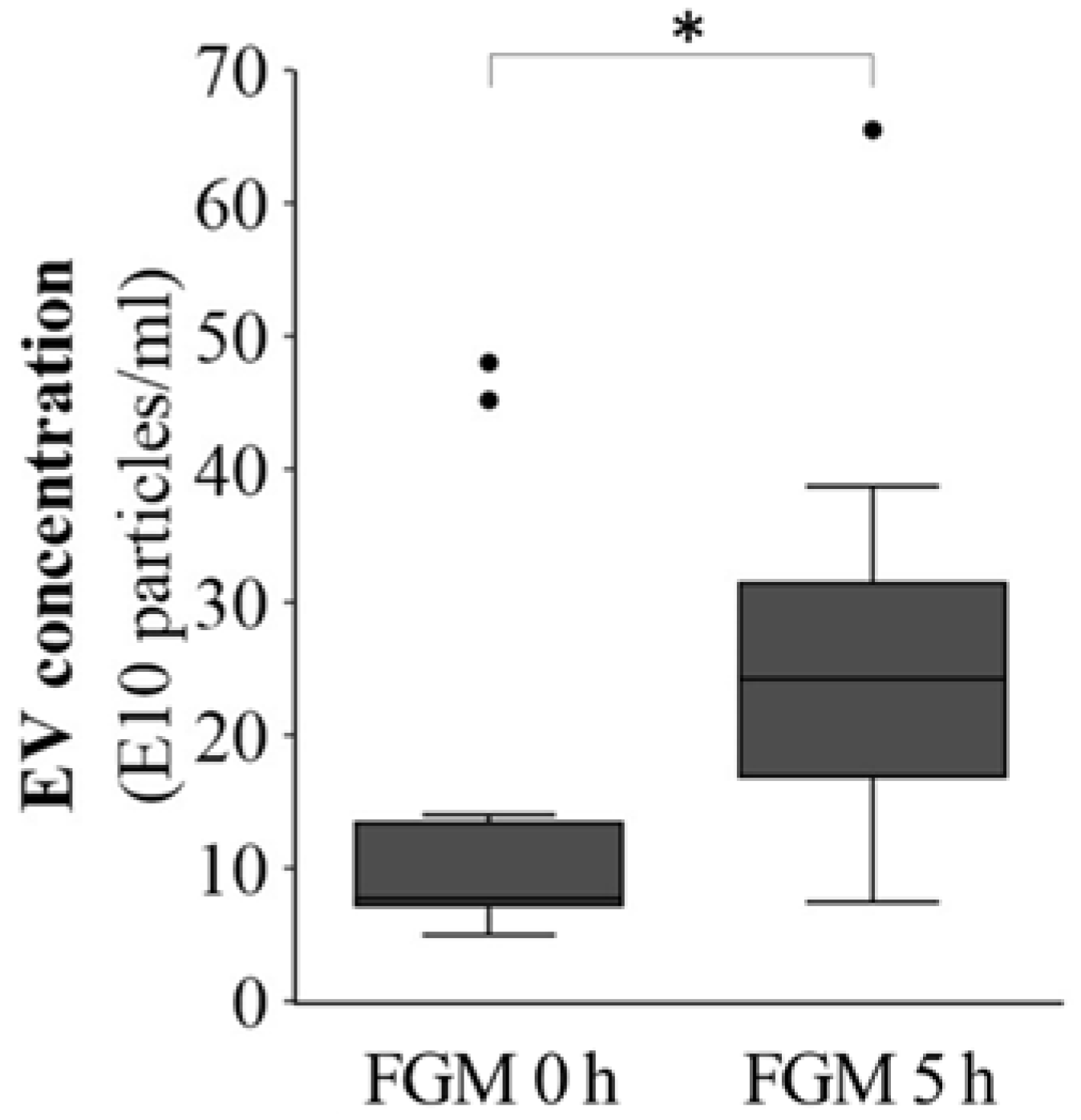
2.2. FMG Affects the EV Concentration in the Initial Hours
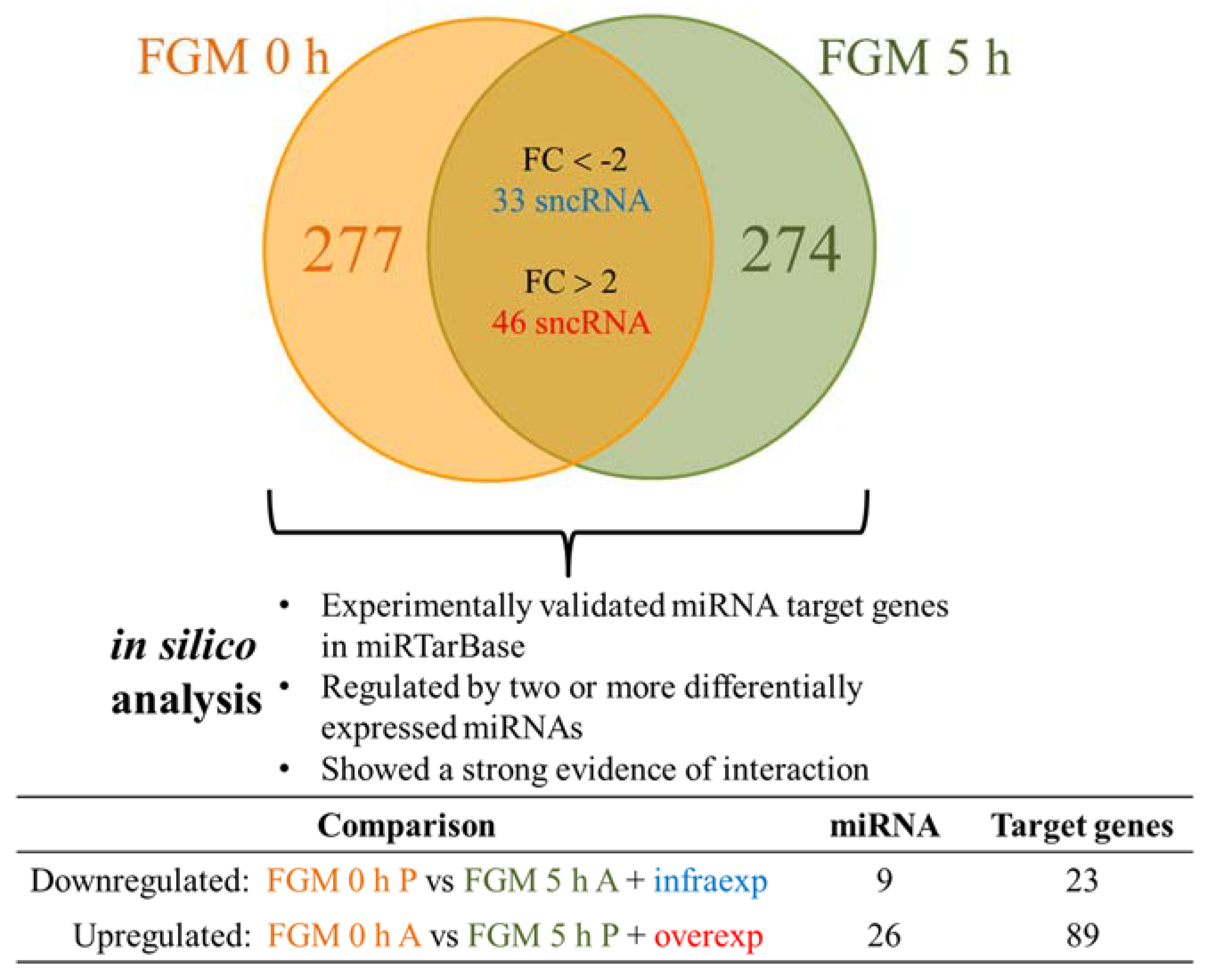
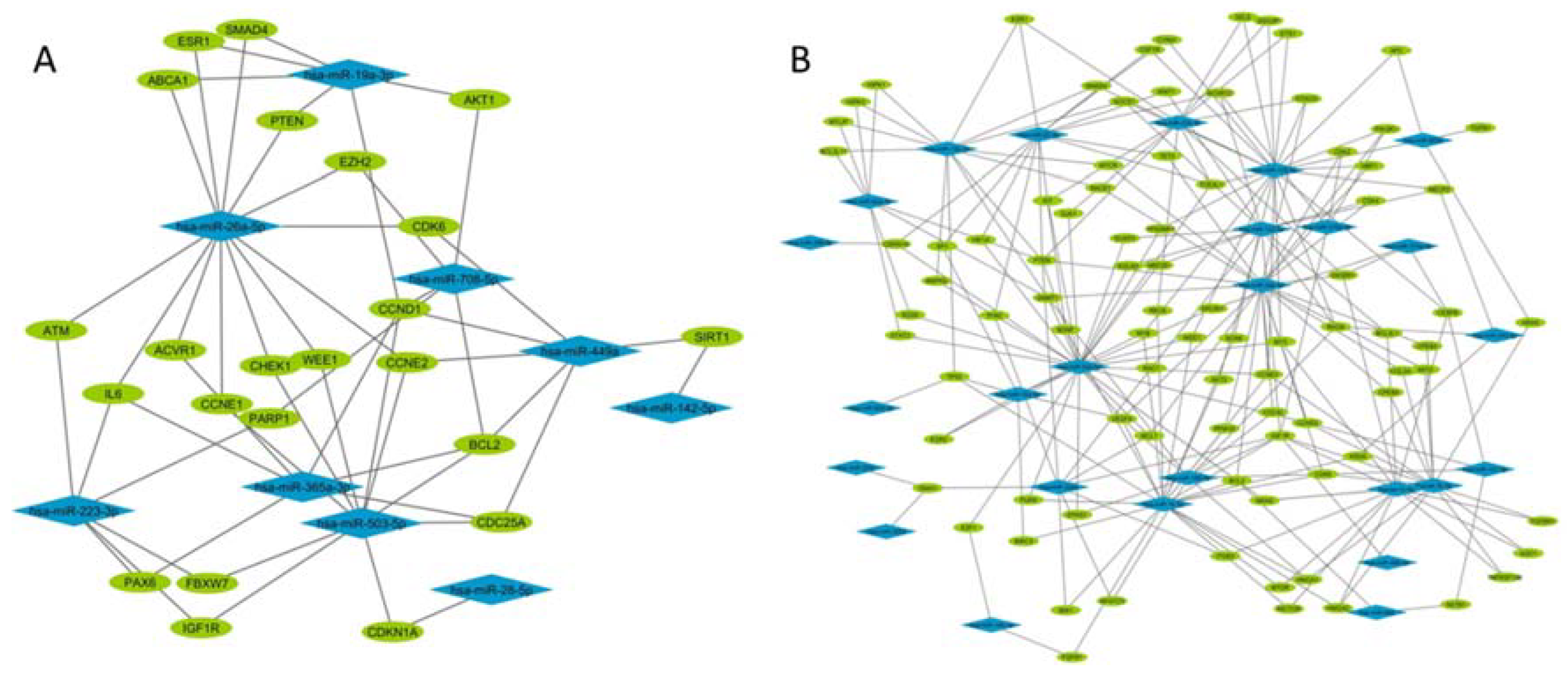
2.3. FGM Modulates the sncRNA Cargo in the First 5 h
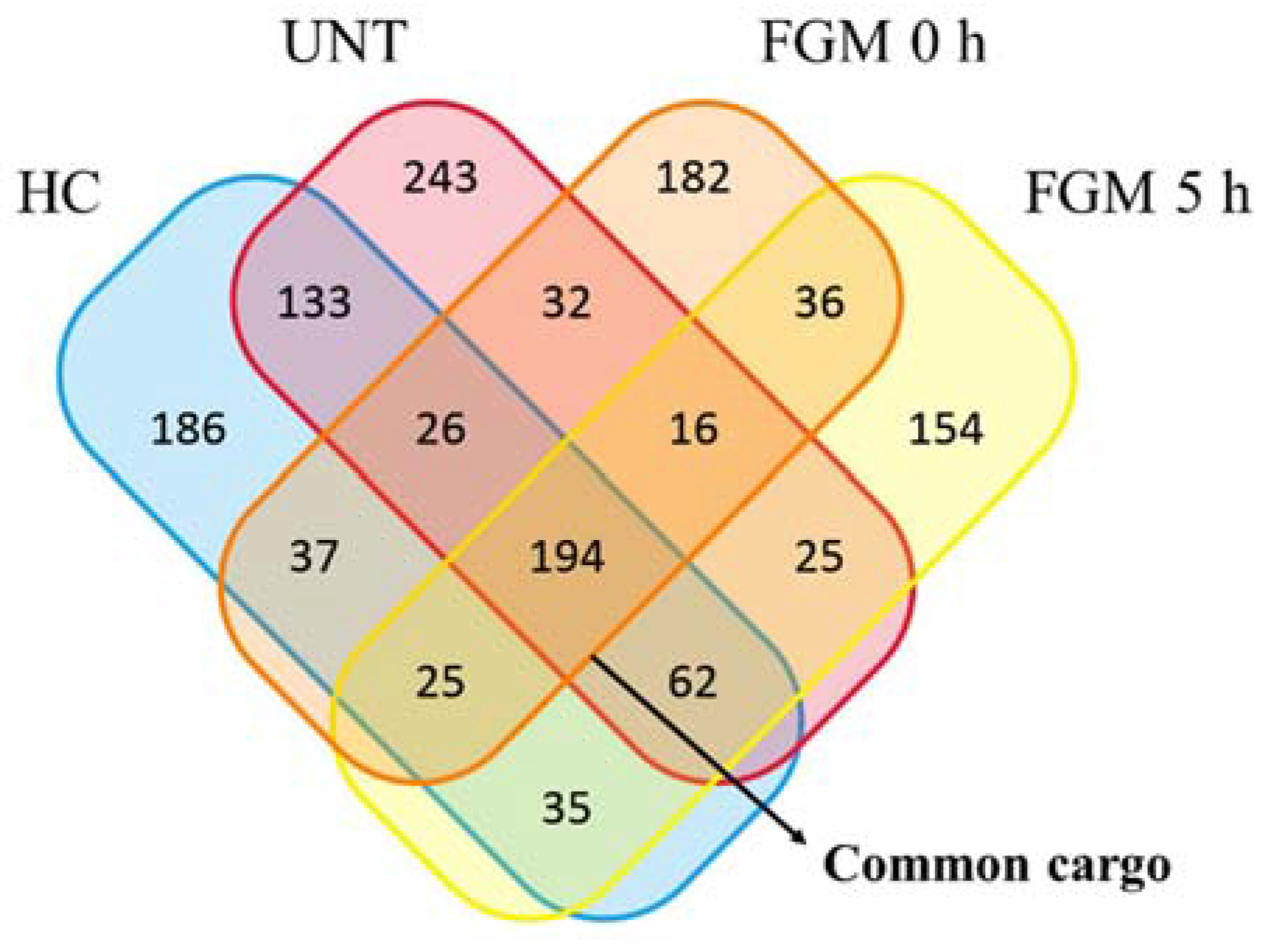
2.4. 30% of sncRNA Cargo of EVs Is Commonly Expressed in All Conditions
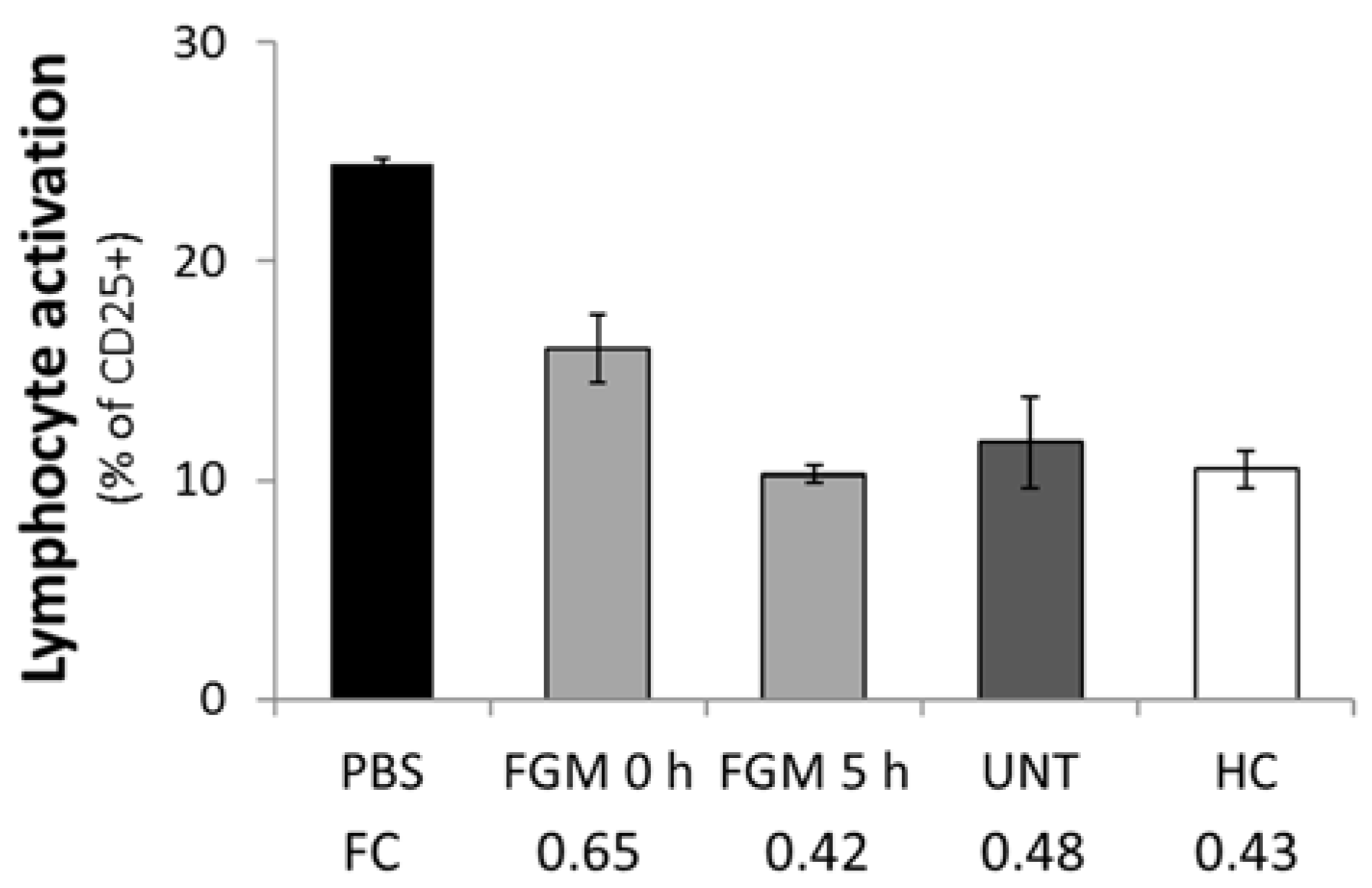
2.5. FGM Affects the Functional Capacity of Circulating EVs
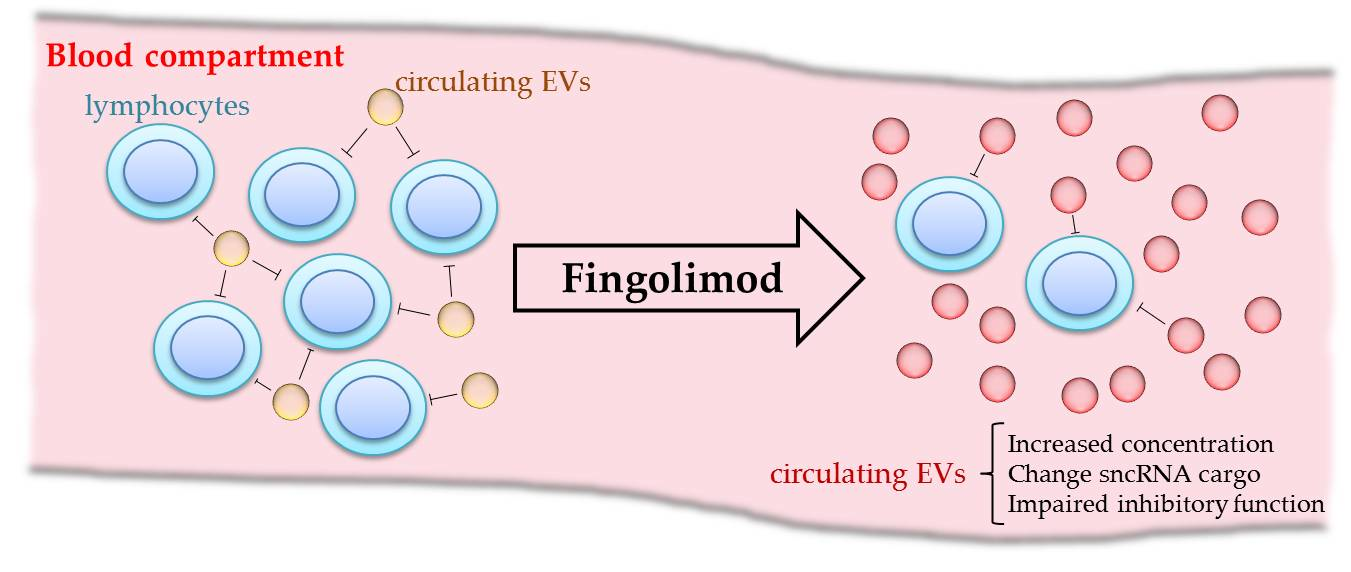
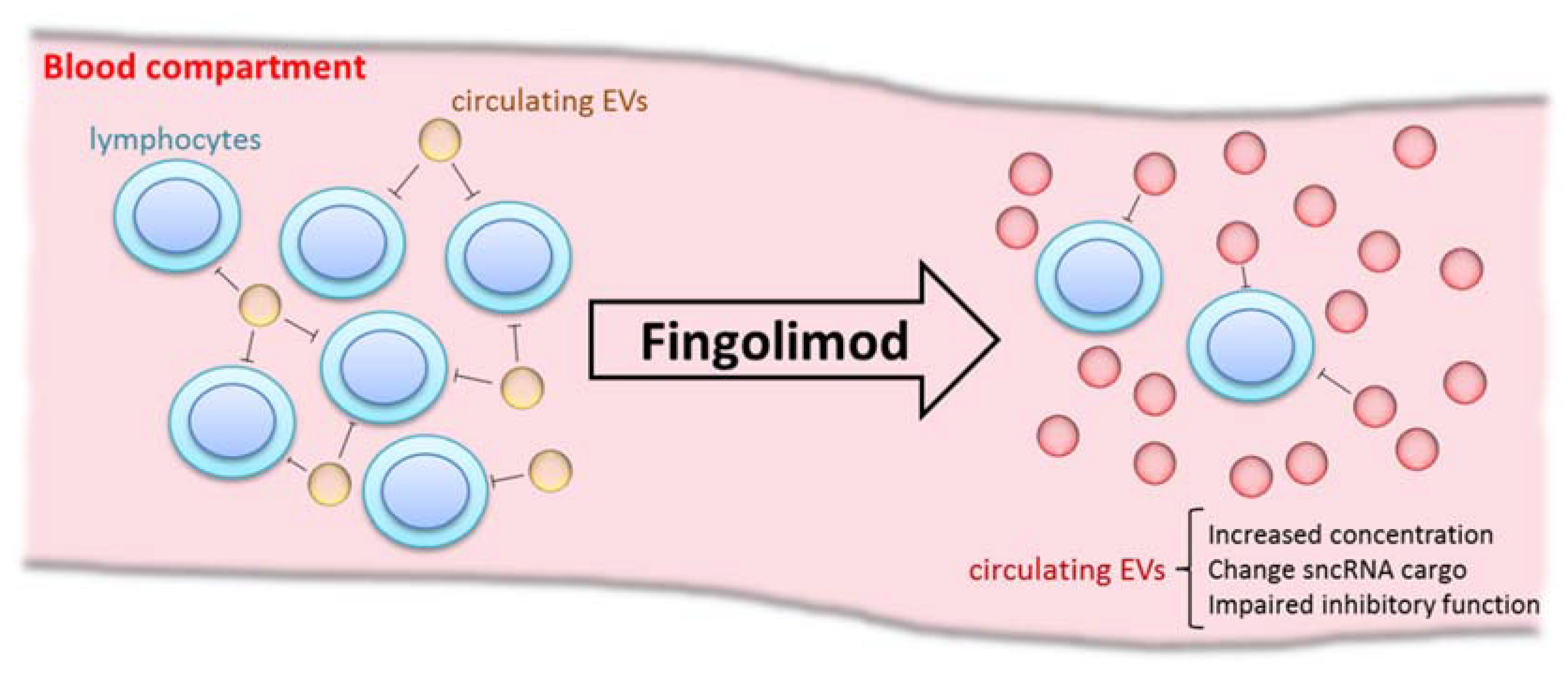
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Isolation of Extracellular Vesicles
4.3. Nanoparticle Tracking Analysis (NTA)
4.4. Flow Cytometry of Extracellular Vesicles
4.5. Small Noncoding RNA (sncRNAs) Arrays
4.6. Coculture of EVs and PBMCs
4.7. Flow Cytometry of PBMCs
4.8. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EGRF | Epidermal growth factor receptor |
| EV | Extracellular vesicle |
| FC | Fold change |
| FGM | Fingolimod |
| HC | Healthy control |
| MS | Multiple sclerosis |
| NTA | Nanoparticle Tracking Analysis |
| PBMC | Peripheral blood mononuclear cell |
| PHA | Phytohaemagglutinin |
| sncRNA | Small noncoding RNA |
| S1PR | Sphingosine 1 phosphate receptor |
| TRAIL | Tumor necrosis factor related apoptosis-inducing ligand |
| UNT | Untreated MS patient |
References
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Arraud, N.; Linares, R.; Tan, S.; Gounou, C.; Pasquet, J.M.; Mornet, S.; Brisson, A.R. Extracellular vesicles from blood plasma: Determination of their morphology, size, phenotype and concentration. J. Thromb. Haemost. 2014, 12, 614–627. [Google Scholar] [CrossRef] [PubMed]
- Goetzl, E.J.; Boxer, A.; Schwartz, J.B.; Abner, E.L.; Petersen, R.C.; Miller, B.L.; Kapogiannis, D. Altered lysosomal proteins in neural-derived plasma exosomes in preclinical Alzheimer disease. Neurology 2015, 85, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654. [Google Scholar] [CrossRef] [PubMed]
- Mittelbrunn, M.; Gutierrez-Vazquez, C.; Villarroya-Beltri, C.; Gonzalez, S.; Sanchez-Cabo, F.; Gonzalez, M.A.; Bernad, A.; Sanchez-Madrid, F. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat. Commun. 2011, 2, 282. [Google Scholar] [CrossRef] [PubMed]
- Chaput, N.; Théry, C. Exosomes: Immune properties and potential clinical implementations. Semin. Immunopathol. 2011, 33, 419–440. [Google Scholar] [CrossRef] [PubMed]
- Sáenz-Cuesta, M.; Osorio-Querejeta, I.; Otaegui, D. Extracellular vesicles in multiple sclerosis: What are they telling us? Front. Cell. Neurosci. 2014, 8, 100. [Google Scholar] [CrossRef] [PubMed]
- Minagar, A.; Jy, W.; Jimenez, J.J.; Sheremata, W.A.; Mauro, L.M.; Mao, W.W.; Horstman, L.L.; Ahn, Y.S. Elevated plasma endothelial microparticles in multiple sclerosis. Neurology 2001, 56, 1319–1324. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.S.; Chervenak, R.; Weinstock-Guttman, B.; Tsunoda, I.; Ramanathan, M.; Martinez, N.; Omura, S.; Sato, F.; Chaitanya, G.V.; Minagar, A.; et al. Blood circulating microparticle species in relapsing–remitting and secondary progressive multiple sclerosis. A case–control, cross sectional study with conventional MRI and advanced iron content imaging outcomes. J. Neurol. Sci. 2015, 355, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Sáenz-Cuesta, M.; Irizar, H.; Castillo-Triviño, T.; Muñoz-Culla, M.; Osorio-Querejeta, I.; Prada, A.; Sepulveda, L.; Lopez-Mato, M.P.; Lopez de Munain, A.; Comabella, M.; et al. Circulating microparticles reflect treatment effects and clinical status in multiple sclerosis. Biomark. Med. 2014, 8, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Verderio, C.; Muzio, L.; Turola, E.; Bergami, A.; Novellino, L.; Ruffini, F.; Riganti, L.; Corradini, I.; Francolini, M.; Garzetti, L.; et al. Myeloid microvesicles are a marker and therapeutic target for neuroinflammation. Ann. Neurol. 2012, 72, 610–624. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimkhani, S.; Vafaee, F.; Young, P.E.; Hur, S.S.J.; Hawke, S.; Devenney, E.; Beadnall, H.; Barnett, M.H.; Suter, C.M.; Buckland, M.E. Exosomal microRNA signatures in multiple sclerosis reflect disease status. Sci. Rep. 2017, 7, 14293. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Hartung, H.P. Mechanism of Action of Oral Fingolimod (FTY720) in Multiple Sclerosis. Clin. Neuropharmacol. 2010, 33, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Francis, G.; Kappos, L.; O’Connor, P.; Collins, W.; Tang, D.; Mercier, F.; Cohen, J.A. Temporal profile of lymphocyte counts and relationship with infections with fingolimod therapy. Mult. Scler. J. 2014, 20, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Trkov, S.; Stenovec, M.; Kreft, M.; Potokar, M.; Parpura, V.; Davletov, B.; Zorec, R. Fingolimod-A sphingosine-like molecule inhibits vesicle mobility and secretion in astrocytes. Glia 2012, 60, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Zinger, A.; Latham, S.L.; Combes, V.; Byrne, S.; Barnett, M.H.; Hawke, S.; Grau, G.E. Plasma levels of endothelial and B-cell-derived microparticles are restored by fingolimod treatment in multiple sclerosis patients. Mult. Scler. J. 2016, 22, 1883–1887. [Google Scholar] [CrossRef] [PubMed]
- Dalla Costa, G.; Finardi, A.; Garzetti, L.; Carandini, T.; Comi, G.; Martinelli, V.; Furlan, R. Disease-modifying treatments modulate myeloid cells in multiple sclerosis patients. Neurol. Sci. 2018, 39, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.P.; Kim, E.Y.; Badr, C.E.; Weissleder, R.; Mempel, T.R.; Tannous, B.A.; Breakefield, X.O. Visualization and tracking of tumour extracellular vesicle delivery and RNA translation using multiplexed reporters. Nat. Commun. 2015, 6, 7029. [Google Scholar] [CrossRef] [PubMed]
- Oliver-Martos, B.; Arnaiz, C.; Pinto-Medel, M.J.; Suardiaz, M.; Hurtado, I.; Garcia-Leon, J.; de Baldeon, F.D.; Fernandez, O. Gene expression changes immediately after interferon beta administration in multiple sclerosis patients according to therapeutic response. Mult. Scler. J. 2013, 19, 501. [Google Scholar]
- Admyre, C.; Johansson, S.M.; Qazi, K.R.; Filen, J.J.; Lahesmaa, R.; Norman, M.; Neve, E.P.A.; Scheynius, A.; Gabrielsson, S. Exosomes with immune modulatory features are present in human breast milk. J. Immunol. 2007, 179, 1969–1978. [Google Scholar] [CrossRef] [PubMed]
- Clayton, A.; Al-Taei, S.; Webber, J.; Mason, M.D.; Tabi, Z. Cancer exosomes express CD39 and CD73, which suppress T cells through adenosine production. J. Immunol. 2011, 187, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Hohjoh, H.; Fukuoka, M.; Sato, W.; Oki, S.; Tomi, C.; Yamaguchi, H.; Kondo, T.; Takahashi, R.; Yamamura, T. Circulating exosomes suppress the induction of regulatory T cells via let-7i in multiple sclerosis. Nat. Commun. 2018, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, B.Z.; Cohen, J.A.; Conway, D.S. Sphingosine 1-Phosphate Receptor Modulators for the Treatment of Multiple Sclerosis. Neurotherapeutics 2017, 14, 859–873. [Google Scholar] [CrossRef] [PubMed]
- Sáenz-Cuesta, M.; Arbelaiz, A.; Oregi, A.; Irizar, H.; Osorio-Querejeta, I.; Muñoz-Culla, M.; Banales, J.M.; Falcon-Perez, J.M.; Olascoaga, J.; Otaegui, D. Methods for extracellular vesicles isolation in a hospital setting. Front. Immunol. 2015, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Perez, A.; Loizaga, A.; Arceo, R.; Lacasa, I.; Rabade, A.; Zorroza, K.; Mosen-Ansorena, D.; Gonzalez, E.; Aransay, A.M.; Falcon-Perez, J.M.; et al. A Pilot Study on the Potential of RNA-Associated to Urinary Vesicles as a Suitable Non-Invasive Source for Diagnostic Purposes in Bladder Cancer. Cancers 2014, 6, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.D.; Tseng, Y.T.; Shrestha, S.; Lin, Y.L.; Khaleel, A.; Chou, C.H.; Chu, C.F.; Huang, H.Y.; Lin, C.M.; Ho, S.Y.; et al. MiRTarBase update 2014: An information resource for experimentally validated miRNA-target interactions. Nucleic Acids Res. 2014, 42, D78–D85. [Google Scholar] [CrossRef] [PubMed]
- Pathan, M.; Keerthikumar, S.; Ang, C.S.; Gangoda, L.; Quek, C.Y.J.; Williamson, N.A.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A.; et al. FunRich: An open access standalone functional enrichment and interaction network analysis tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject | Age (Years) | EDSS | Evolution Time of MS (Years) | Previous Treatment |
|---|---|---|---|---|
| UNT1 | 32 | 2 | 10.5 | |
| UNT2 | 31 | 0 | 0.3 | |
| UNT3 | 37 | 0 | 0.0 | |
| UNT4 | 52 | 2 | 13.8 | |
| UNT5 | 22 | 0 | 0.0 | |
| FGM1 | 36 | 4.5 | 6.6 | INFβ 1A |
| FGM2 | 34 | 4.5 | 10.8 | INFβ 1B |
| FGM3 | 50 | 6 | 18.5 | INFβ 1A |
| FGM4 | 49 | 4 | 20.2 | NTZ |
| FGM5 | 36 | 3.5 | 16.8 | NTZ |
| FGM6 | 40 | 1 | 9.2 | NTZ |
| FGM7 | 50 | 3.5 | 22.1 | NTZ |
| FGM8 | 40 | 2 | 12.4 | INFβ 1B |
| FGM9 | 33 | 0 | 9.3 | GA |
| FGM10 | 30 | 3.5 | 4.2 | NTZ |
| FGM11 | 41 | 3 | 20.8 | GA |
| HC1 | 41 | |||
| HC2 | 28 | |||
| HC3 | 22 | |||
| HC4 | 36 | |||
| HC5 | 26 | |||
| HC6 | 33 | |||
| HC7 | 33 | |||
| HC8 | 42 |
| sncRNA Type | HC | UNT | FGM 0 h | FGM 5 h | Common Cargo | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mature miRNA | 382 | 55% | 374 | 53% | 262 | 48% | 243 | 44% | 124 | 64% |
| Immature miRNA | 137 | 20% | 148 | 21% | 115 | 21% | 127 | 23% | 28 | 14% |
| snoRNA | 140 | 20% | 139 | 20% | 134 | 24% | 134 | 24% | 32 | 16% |
| CDBox | 29 | 4% | 33 | 5% | 25 | 5% | 30 | 5% | 9 | 5% |
| H/ACAbox | 6 | 1% | 7 | 1% | 9 | 2% | 7 | 1% | 1 | 1% |
| scaRNA | 4 | 1% | 4 | 1% | 5 | 1% | 6 | 1% | 0 | 0% |
| TOTAL | 698 | 100% | 705 | 100% | 550 | 100% | 547 | 100% | 194 | 100% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sáenz-Cuesta, M.; Alberro, A.; Muñoz-Culla, M.; Osorio-Querejeta, I.; Fernandez-Mercado, M.; Lopetegui, I.; Tainta, M.; Prada, Á.; Castillo-Triviño, T.; Falcón-Pérez, J.M.; et al. The First Dose of Fingolimod Affects Circulating Extracellular Vesicles in Multiple Sclerosis Patients. Int. J. Mol. Sci. 2018, 19, 2448. https://doi.org/10.3390/ijms19082448
Sáenz-Cuesta M, Alberro A, Muñoz-Culla M, Osorio-Querejeta I, Fernandez-Mercado M, Lopetegui I, Tainta M, Prada Á, Castillo-Triviño T, Falcón-Pérez JM, et al. The First Dose of Fingolimod Affects Circulating Extracellular Vesicles in Multiple Sclerosis Patients. International Journal of Molecular Sciences. 2018; 19(8):2448. https://doi.org/10.3390/ijms19082448
Chicago/Turabian StyleSáenz-Cuesta, Matías, Ainhoa Alberro, Maider Muñoz-Culla, Iñaki Osorio-Querejeta, Marta Fernandez-Mercado, Itziar Lopetegui, Mikel Tainta, Álvaro Prada, Tamara Castillo-Triviño, Juan Manuel Falcón-Pérez, and et al. 2018. "The First Dose of Fingolimod Affects Circulating Extracellular Vesicles in Multiple Sclerosis Patients" International Journal of Molecular Sciences 19, no. 8: 2448. https://doi.org/10.3390/ijms19082448
APA StyleSáenz-Cuesta, M., Alberro, A., Muñoz-Culla, M., Osorio-Querejeta, I., Fernandez-Mercado, M., Lopetegui, I., Tainta, M., Prada, Á., Castillo-Triviño, T., Falcón-Pérez, J. M., Olascoaga, J., & Otaegui, D. (2018). The First Dose of Fingolimod Affects Circulating Extracellular Vesicles in Multiple Sclerosis Patients. International Journal of Molecular Sciences, 19(8), 2448. https://doi.org/10.3390/ijms19082448