Edema Induced by a Crotalus durissus terrificus Venom Serine Protease (Cdtsp 2) Involves the PAR Pathway and PKC and PLC Activation

Abstract
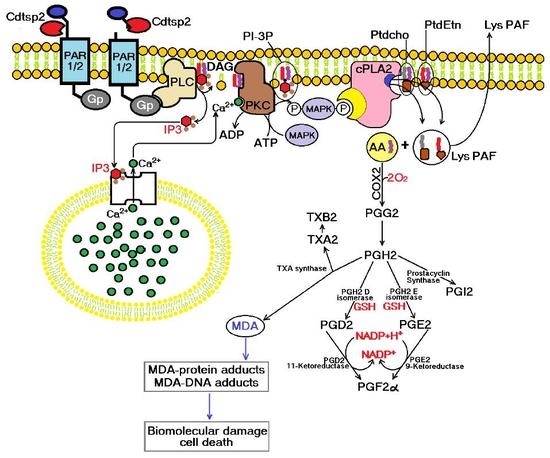
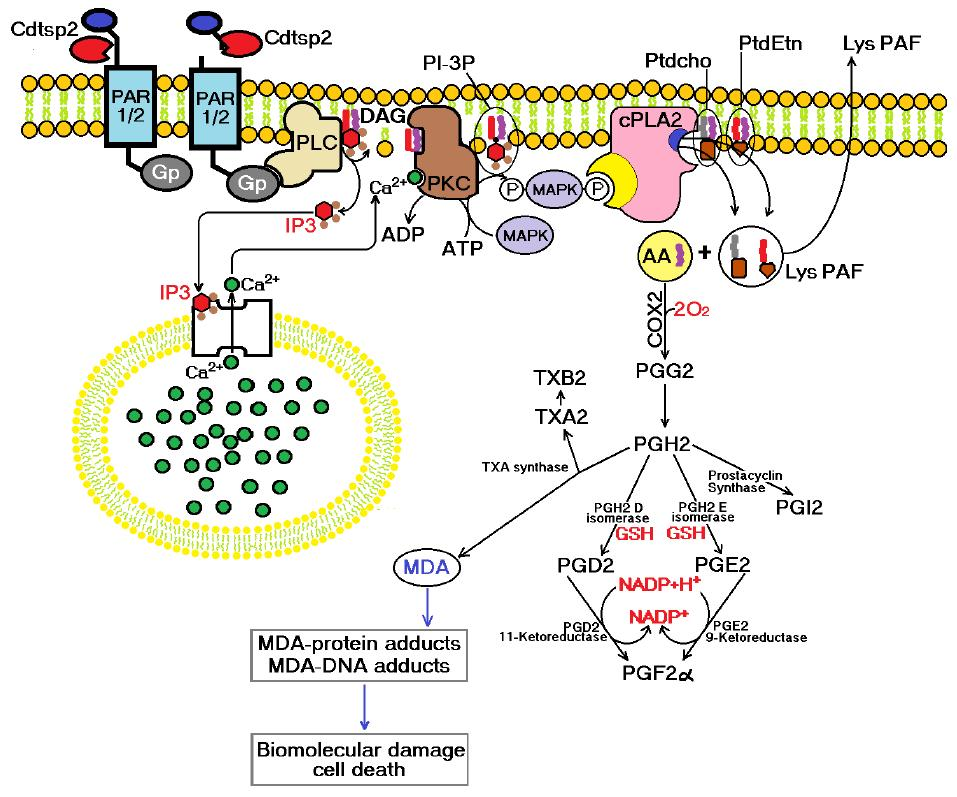{kind=link}
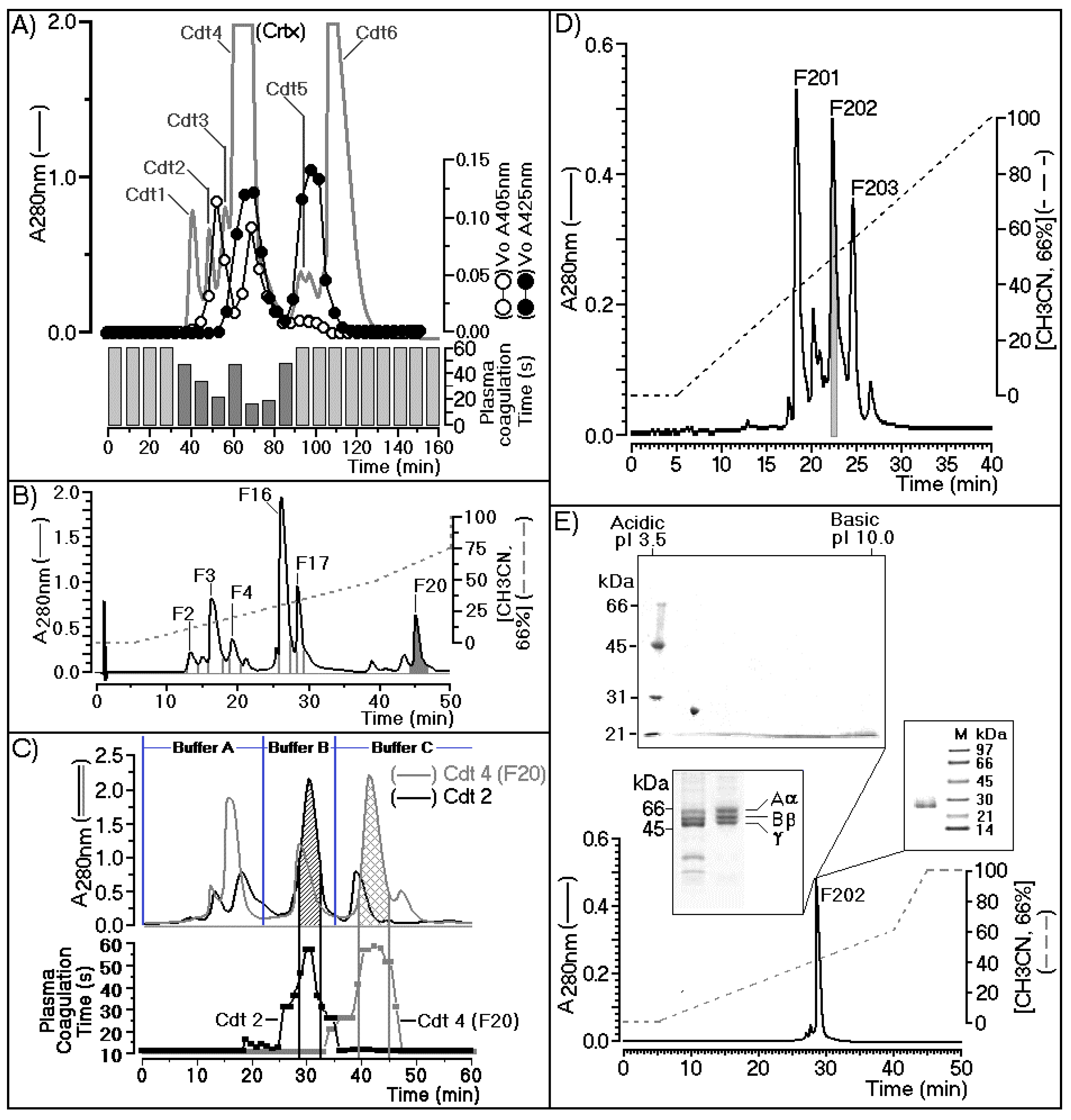{kind=link}
{kind=link}
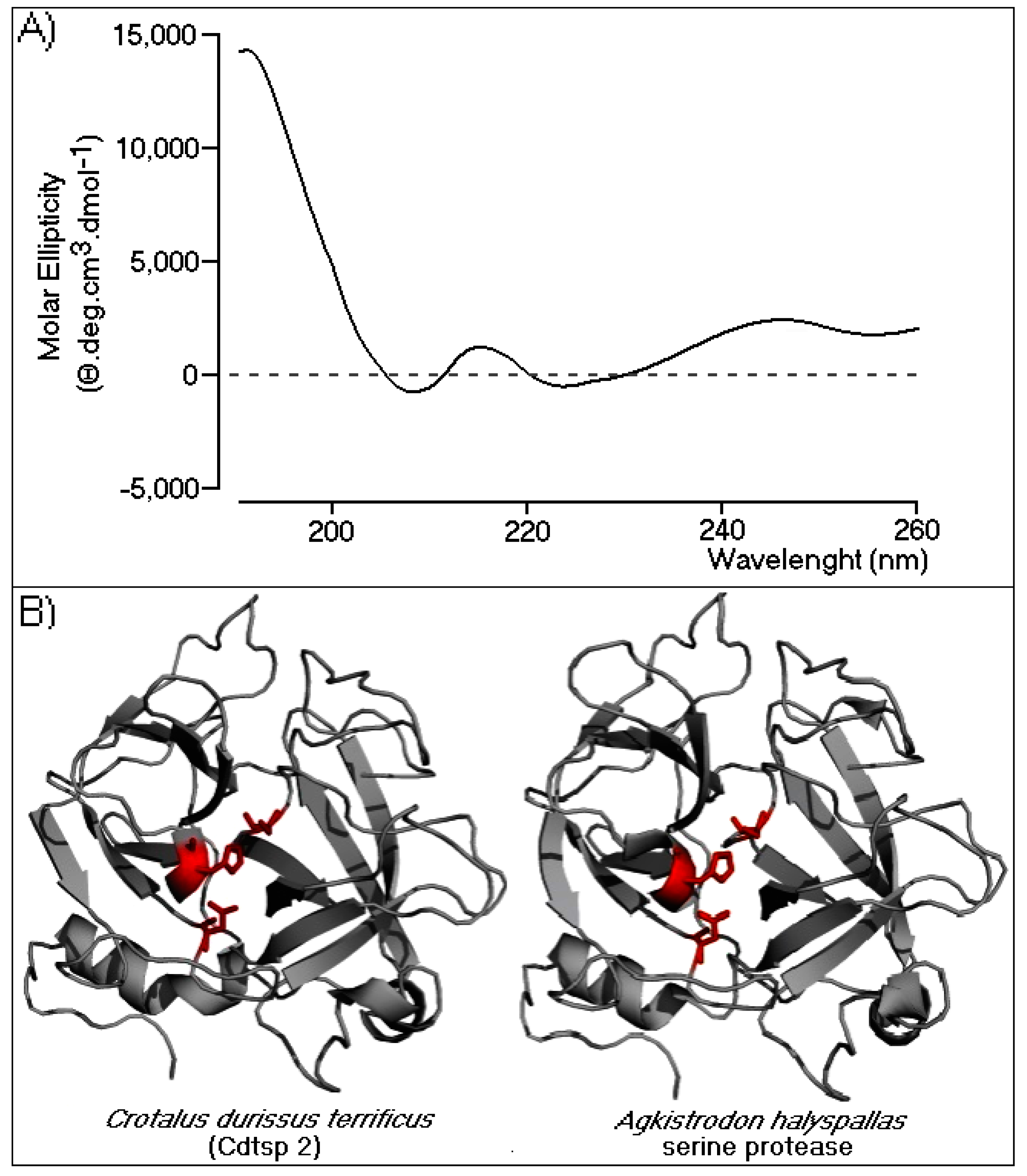{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Purification of Cdtsp 2 and Coagulation Activity
2.2. Structural Analysis
2.2.1. Amino Acid Sequence
2.2.2. Circular Dichroism and Three-Dimensional Model of Cdtsp 2
2.3. Pharmacological Characterization of Cdtsp 2
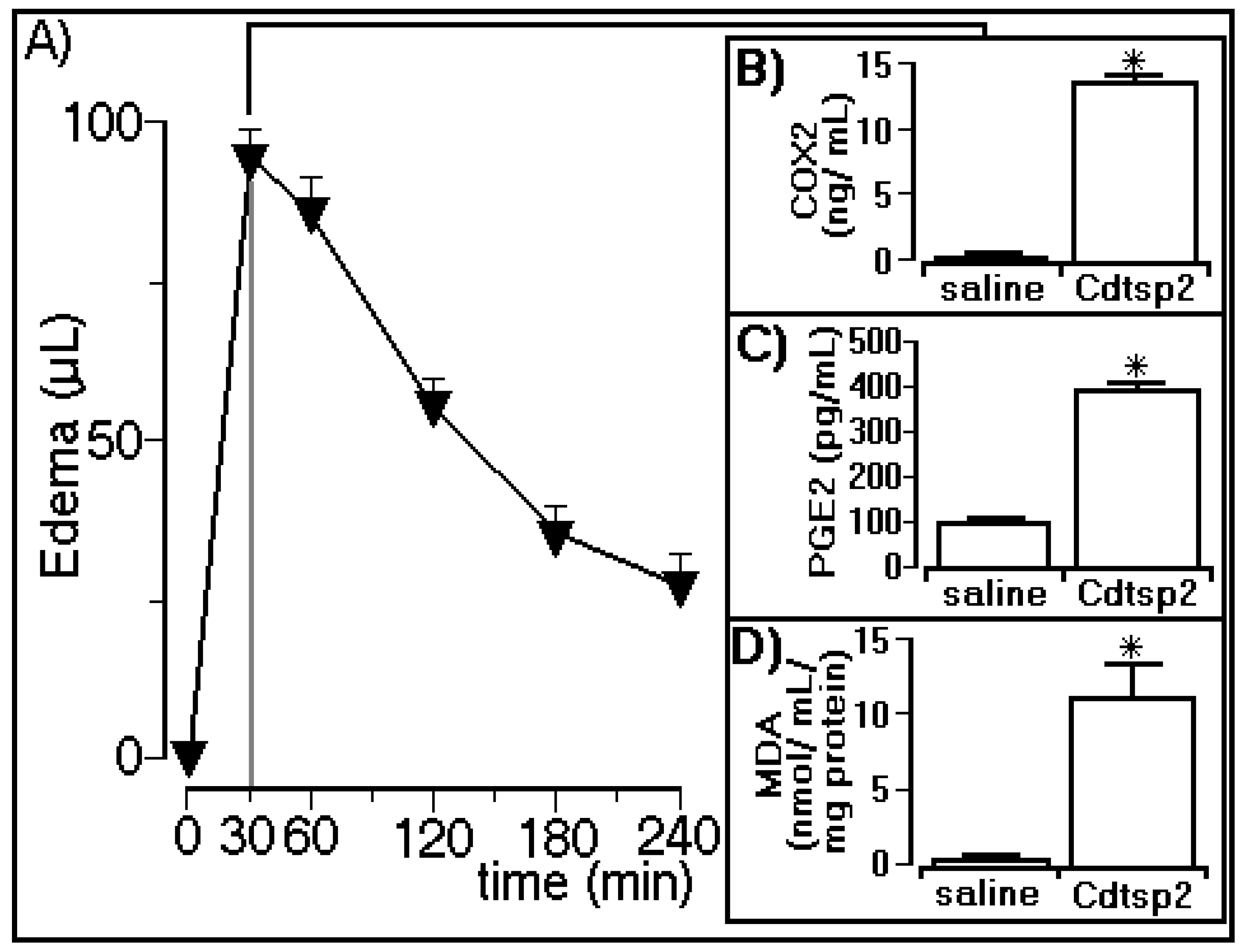
2.3.1. Edema Assay and Biochemical Determinations
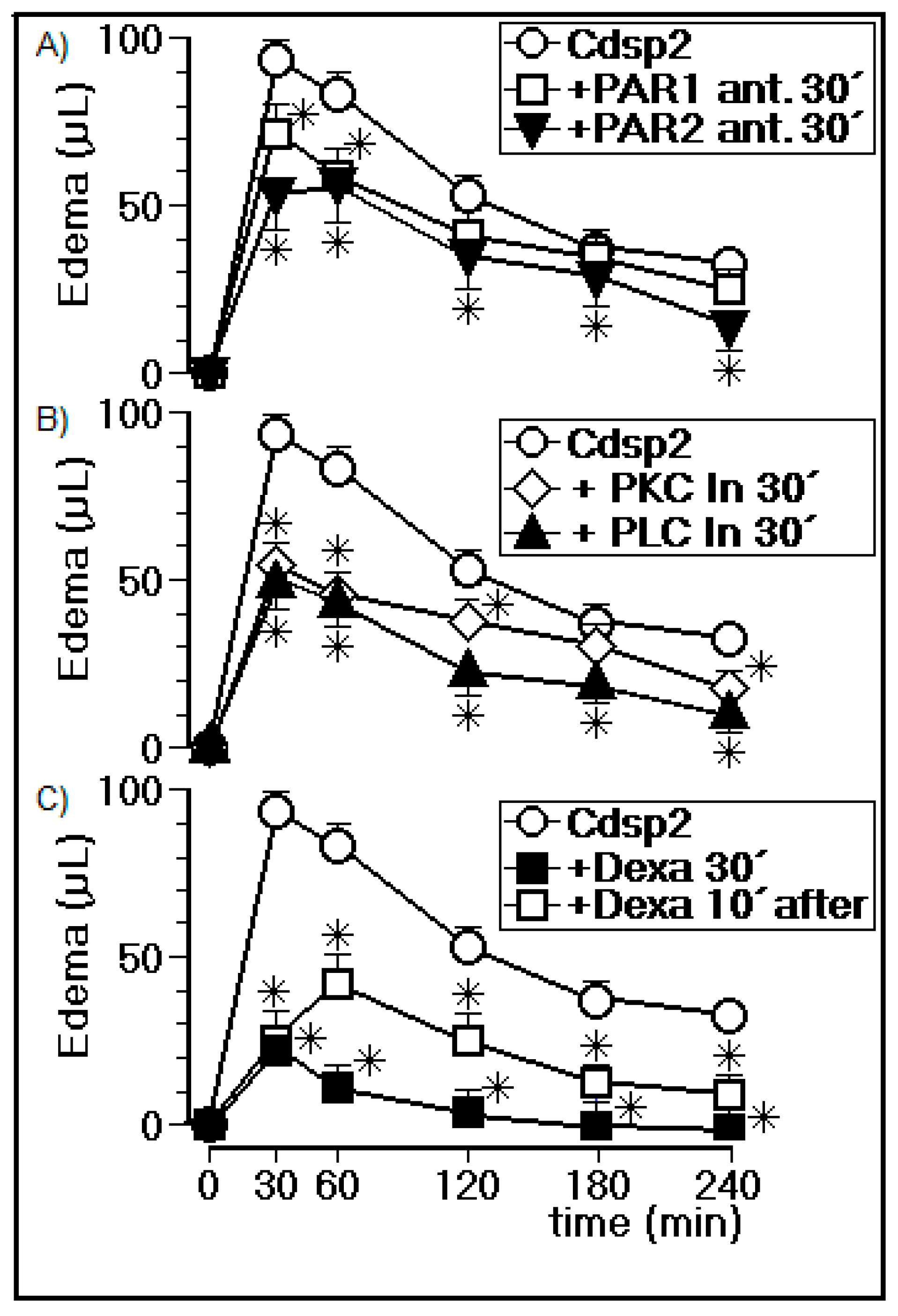
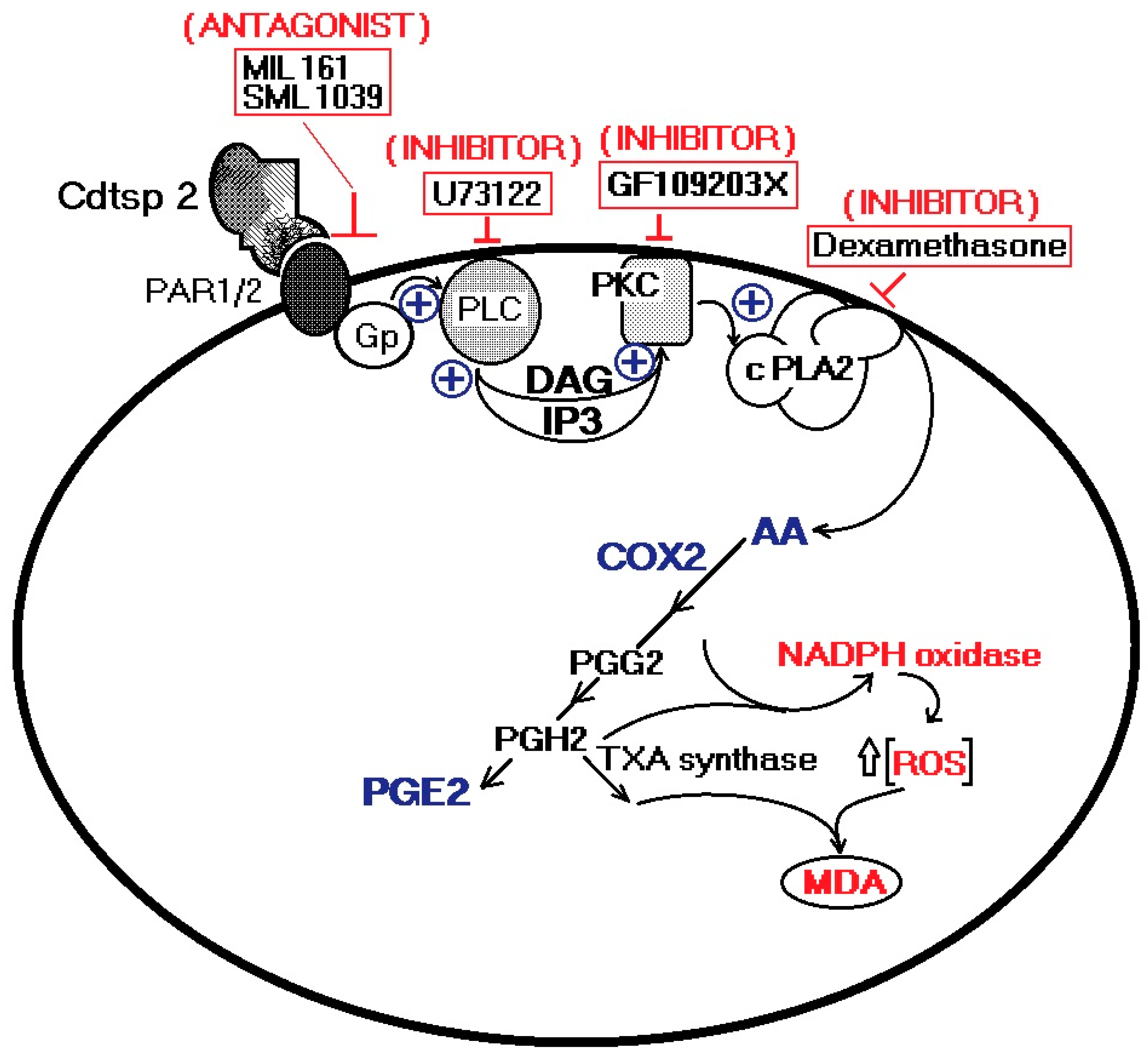
2.3.2. Edema Assay in the Presence of PLC, PKC, a cPLA2 Inhibitor, and a PAR1/2 Antagonist
3. Discussion
4. Materials and Methods
4.1. Cdtsp 2 Purification
4.2. Coagulation Activity
4.3. Primary Structure Determination
4.4. Circular Dichroism Spectroscopy
4.5. Structural Homology Modeling
4.6. Enzymatic Activity
4.7. Quantification of Enzymes
4.7.1. Evaluation of COX-2 Levels
4.7.2. Prostaglandin E2 Quantification
4.7.3. Lipid Peroxidation Determination
4.8. Evaluation of Paw Edema
4.8.1. PAR1 Antagonist (ML161) and PAR2 Antagonist (SML 1039)
4.8.2. PLC Inhibition by U73122
4.8.3. PKC Inhibition by GF109203X
4.8.4. Dexamethasone Treatment
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- De Oliveira, D.G.L.; Murakami, M.T.; Cintra, A.C.O.; Franco, J.J.; Sampaio, S.V.; Arni, R.K. Functional and structural analysis of two fibrinogen-activating enzymes isolated from the venoms of Crotalus durissus terrificus and Crotalus durissus collilineatus. Acta Biochim. Biophys. Sin. (Shanghai) 2009, 41, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Boldrini-França, J.; Rodrigues, R.S.; Santos-Silva, L.K.; de Souza, D.L.N.; Gomes, M.S.R.; Cologna, C.T.; de Pauw, E.; Quinton, L.; Henrique-Silva, F.; de Melo Rodrigues, V.; et al. Expression of a new serine protease from Crotalus durissus collilineatus venom in Pichia pastoris and functional comparison with the native enzyme. Appl. Microbiol. Biotechnol. 2015, 99, 9971–9986. [Google Scholar]
- Patiño, A.C.; Pereañez, J.A.; Gutiérrez, J.M.; Rucavado, A. Biochemical and biological characterization of two serine proteinases from Colombian Crotalus durissus cumanensis snake venom. Toxicon 2013, 63, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Toyama, M.H.; Toyama, D.D.O.; Passero, L.F.D.; Laurenti, M.D.; Corbett, C.E.; Tomokane, T.Y.; Fonseca, F.V.; Antunes, E.; Joazeiro, P.P.; Beriam, L.O.; et al. Isolation of a new l-amino acid oxidase from Crotalus durissus cascavella venom. Toxicon 2006, 47, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Yonamine, C.M.; Prieto-da-Silva, A.R.; Magalhães, G.S.; Rádis-Baptista, G.; Morganti, L.; Ambiel, F.C.; Chura-Chambi, R.M.; Yamane, T.; Camillo, M.A. Cloning of serine protease cDNAs from Crotalus durissus terrificus venom gland and expression of a functional Gyroxin homologue in COS-7 cells. Toxicon 2009, 54, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Menaldo, D.L.; Bernardes, C.P.; Pereira, J.C.; Silveira, D.S.C.; Mamede, C.C.; Stanziola, L.; Oliveira, F.D.; Pereira-Crott, L.S.; Faccioli, L.H.; Sampaio, S.V. Effects of two serine proteases from Bothrops pirajai snake venom on the complement system and the inflammatory response. Int. Immunopharmacol. 2013, 15, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Da-Silva-Freitas, D.; Boldrini-França, J.; Arantes, E.C. PEGylation: A successful approach to improve the biopharmaceutical potential of snake venom thrombin-like serine protease. Protein Pept. Lett. 2015, 22, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Menaldo, D.L.; Bernardes, C.P.; Zoccal, K.F.; Jacob-Ferreira, A.L.; Costa, T.R.; Del Lama, M.P.; Naal, R.M.; Frantz, F.G.; Faccioli, L.H.; Sampaio, S.V. Immune cells and mediators involved in the inflammatory responses induced by a P-I metalloprotease and a phospholipase A2 from Bothrops atrox venom. Mol. Immunol. 2017, 85, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Mamede, C.C.N.; de Sousa, B.B.; da Cunha Pereira, D.F.; Matias, M.S.; de Queiroz, M.R.; de Morais, N.C.G.; Vieira, S.A.P.B.; Stanziola, L.; Oliveira, F. Comparative analysis of local effects caused by Bothrops alternatus and Bothrops moojeni snake venoms: Enzymatic contributions and inflammatory modulations. Toxicon 2016, 117, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.W.; Serrano, S.M. Insights into and speculations about snake venom metalloproteinase (SVMP) synthesis, folding and disulfide bond formation and their contribution to venom complexity. FEBS J. 2008, 275, 3016–3030. [Google Scholar] [CrossRef] [PubMed]
- Zychar, B.C.; Dale, C.S.; Demarchi, D.S.; Gonçalves, L.R.C. Contribution of metalloproteases, serine proteases and phospholipases A2 to the inflammatory reaction induced by Bothrops jararaca crude venom in mice. Toxicon 2010, 55, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Leslie, C.C. Cytosolic phospholipase A(2): Physiological function and role in disease. J. Lipid. Res. 2015, 56, 1386–1402. [Google Scholar] [CrossRef] [PubMed]
- Burke, J.E.; Dennis, E.A. Phospholipase A(2) structure/function, mechanism, and signaling. J. Lipid Res. 2009, 50, S237–S242. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, C.A.; Newman, Z.L.; Morford, J.J.; Ryan, S.B.; Battani, K.A.; Su, Z. Cyclooxygenase-2, prostaglandin E2 glycerol ester and nitric oxide are involved in muscarine-induced presynaptic enhancement at the vertebrate neuromuscular junction. J. Physiol. 2013, 591, 4749–4764. [Google Scholar] [CrossRef] [PubMed]
- Norris, P.C.; Gosselin, D.; Reichart, D.; Glass, C.K.; Dennis, E.A. Phospholipase A(2) regulates eicosanoid class switching during inflammasome activation. Proc. Natl. Acad. Sci. USA 2014, 111, 12746–12751. [Google Scholar] [CrossRef] [PubMed]
- Mouchlis, V.D.; Dennis, E.A. Membrane and Inhibitor Interactions of Intracellular Phospholipases A(2). Adv. Biol. Regul. 2016, 61, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; Chuang, D.Y.; Zong, Y.; Jiang, J.; Lee, J.C.M.; Gu, Z.; Simonyi, A. Role of cytosolic phospholipase A(2) in oxidative and inflammatory signaling pathways in different cell types in the central nervous system. Mol. Neurobiol. 2014, 50, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, A.A.; Horrocks, L.A. Signaling and interplay mediated by phospholipases A2, C, and D in LA-N-1 cell nuclei. Reprod. Nutr. Dev. 2005, 45, 613–631. [Google Scholar] [CrossRef] [PubMed]
- Cosentino-Gomes, D.; Rocco-Machado, N.; Meyer-Fernandes, J.R. Cell Signaling through Protein Kinase C Oxidation and Activation. Int. J. Mol. Sci. 2012, 13, 10697–10721. [Google Scholar] [CrossRef] [PubMed]
- Cavada, B.S.; Castellón, R.E.R.; Vasconcelos, G.G.; Rocha, B.A.M.; Bezerra, G.A.; Debray, H.; Delatorre, P.; Nagano, C.S.; Toyama, M.; Pinto, V.P.; et al. Crystallization and preliminary X-ray diffraction analysis of a new chitin-binding protein from Parkia platycephala seeds. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2005, 61, 841–843. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C. Protein kinase C: Poised to signal. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E395–E402. [Google Scholar] [CrossRef] [PubMed]
- Grimsey, N.; Soto, A.G.; Trejo, J. Regulation of Protease-activated Receptor Signaling by Posttranslational Modifications. IUBMB Life 2011, 63, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Covic, L.; Gresser, A.L.; Talavera, J.; Swift, S.; Kuliopulos, A. Activation and inhibition of G protein-coupled receptors by cell-penetrating membrane-tethered peptides. Proc. Natl. Acad. Sci. USA 2002, 99, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, A.A.; Horrocks, L.A. Phospholipase A2-Generated Lipid Mediators in the Brain: The Good, the Bad, and the Ugly. Neuroscientist 2006, 12, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, B.; Liu, L.; Zhang, E.-L.; Sun, B.; Xu, G.; Chen, J.; Gao, Y.Q. Arachidonic Acid Metabolism Pathway Is Not Only Dominant in Metabolic Modulation but Associated With Phenotypic Variation After Acute Hypoxia Exposure. Front. Physiol. 2018, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; He, Y.; Chuang, D.Y.; Lee, J.C.; Gu, Z.; Simonyi, A.; Sun, A.Y. Integrating cytosolic phospholipase A2 with oxidative/nitrosative signaling pathways in neurons: A novel therapeutic strategy for AD. Mol. Neurobiol. 2012, 46, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, F.V.; Antunes, E.; Morganti, R.P.; Monteiro, H.S.; Martins, A.M.; Toyama, D.O.; Marangoni, S.; Toyama, M.H. Characterization of a new platelet aggregating factor from crotoxin Crotalus durissus cascavella venom. Protein J. 2006, 25, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.R.; Trejo, J. Challenges and Opportunities in Protease-Activated Receptor Drug Development. Ann. Rev. Pharmacol. Toxicol. 2017, 57, 349–373. [Google Scholar] [CrossRef] [PubMed]
- Costa Jde, O.; Fonseca, K.C.; Garrote-Filho, M.S.; Cunha, C.C.; de Freitas, M.V.; Silva, H.S.; Araújo, R.B.; Penha-Silva, N.; de Oliveira, F. Structural and functional comparison of proteolytic enzymes from plant latex and snake venoms. Biochimie 2010, 92, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Castro, H.C.; Silva, D.M.; Craik, C.; Zingali, R.B. Structural features of a snake venom thrombin-like enzyme: Thrombin and trypsin on a single catalytic platform? Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 2001, 1547, 183–195. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and Inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Maroun, R.C.; Serrano, S.M.T. Identification of the substrate-binding exosites of two snake venom serine proteinases: Molecular basis for the partition of two essential functions of thrombin. J. Mol. Recognit. 2003, 17, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, A.K.; Singh, A.; Mukherjee, K.J.; Panda, A.K. Refolding and purification of recombinant L-asparaginase from inclusion bodies of E. coli into active tetrameric protein. Front. Microbiol. 2014, 5, 486. [Google Scholar] [CrossRef] [PubMed]
- Goettig, P. Effects of Glycosylation on the Enzymatic Activity and Mechanisms of Proteases. Int. J. Mol. Sci. 2016, 17, 1969. [Google Scholar] [CrossRef] [PubMed]
- Coorssen, J.R. Phospholipase activation and secretion: Evidence that PLA2, PLC, and PLD are not essential to exocytosis. Am. J. Physiol. Physiol. 1996, 270, C1153–C1163. [Google Scholar] [CrossRef] [PubMed]
- Gijón, M.A.; Leslie, C.C. Regulation of arachidonic acid release and cytosolic Phospholipase A2 activation. J. Leukoc. Biol. 1999, 65, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Balsinde, J.; Balboa, M.A.; Dennis, E.A. Identification of a Third Pathway for Arachidonic Acid Mobilization and Prostaglandin Production in Activated P388D1 Macrophage-like Cells. J. Biol. Chem. 2000, 275, 22544–22549. [Google Scholar] [CrossRef] [PubMed]
- Touqui, L.; Alaoui-El-Azher, M. Mammalian Secreted Phospholipases A2 and Their Pathophysiolo-gical Significance in Inflammatory Diseases. Curr. Mol. Med. 2001, 1, 739–754. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Weng, Y.I.; Simonyi, A.; Krugh, B.W.; Liao, Z.; Weisman, G.A.; Sun, G.Y. Role of PKC and MAPK in cytosolic PLA2 phosphorylation and arachadonic acid release in primary murine astrocytes. J. Neurochem. 2002, 83, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Balboa, M.A.; Balsinde, J. Oxidative stress and arachidonic acid mobilization. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2006, 1761, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Soubhye, J.; van Antwerpen, P.; Dufrasne, F. Targeting Cytosolic Phospholipase A2α for Novel Anti-Inflammatory Agents. Curr. Med. Chem. 2018, 25, 2418–2447. [Google Scholar] [CrossRef] [PubMed]
- Yonamine, C.M.; Kondo, M.Y.; Juliano, M.A.; Icimoto, M.Y.; Baptista, G.R.; Yamane, T.; Oliveira, V.; Juliano, L.; Lapa, A.J.; Lima-Landman, M.T.; et al. Kinetic characterization of gyroxin, a serine protease from Crotalus durissus terrificus venom. Biochimie 2012, 94, 2791–2793. [Google Scholar] [CrossRef] [PubMed]
- Dale, C.; Vergnolle, N. Protease Signaling to G Protein-Coupled Receptors: Implications for Inflammation and Pain. J. Recept. Signal Transduct. Res. 2008, 28, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Houle, S.; Papez, M.D.; Ferazzini, M.; Hollenberg, M.D.; Vergnolle, N. Neutrophils and the kallikrein-kinin system in proteinase-activated receptor 4-mediated inflammation in rodents. Br. J. Pharmacol. 2005, 146, 760–678. [Google Scholar] [CrossRef] [PubMed]
- Park, K.W.; Jin, B.K. Thrombin-induced oxidative stress contributes to the death of hippocampal neurons: Role of neuronal NADPH oxidase. J. Neurosci. Res. 2008, 86, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, E.; Sunagar, K.; Almeida, D.; Vasconcelos, V.; Antunes, A. IMPACT_S: Integrated Multiprogram Platform to Analyze and Combine Tests of Selection. PLoS ONE 2014, 9, e96243. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2016, 45, D158–D169.
- Lua, R.C.; Lichtarge, O. PyETV: A PyMOL evolutionary trace viewer to analyze functional site predictions in protein complexes. Bioinformatics 2010, 26, 2981–2982. [Google Scholar] [CrossRef] [PubMed]
- Prasa, D.; Svendsen, L.; Stürzebecher, J. Inhibition of thrombin generation in plasma by inhibitors of factor Xa. Thromb. Haemost. 1997, 78, 1215–1220. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, C.R.C.; Belchor, M.N.; Rodrigues, C.F.B.; Toyama, D.D.O.; De Oliveira, M.A.; Novaes, D.P.; Toyama, M.H. Edema Induced by a Crotalus durissus terrificus Venom Serine Protease (Cdtsp 2) Involves the PAR Pathway and PKC and PLC Activation. Int. J. Mol. Sci. 2018, 19, 2405. https://doi.org/10.3390/ijms19082405
Costa CRC, Belchor MN, Rodrigues CFB, Toyama DDO, De Oliveira MA, Novaes DP, Toyama MH. Edema Induced by a Crotalus durissus terrificus Venom Serine Protease (Cdtsp 2) Involves the PAR Pathway and PKC and PLC Activation. International Journal of Molecular Sciences. 2018; 19(8):2405. https://doi.org/10.3390/ijms19082405
Chicago/Turabian StyleCosta, Caroline R. C., Mariana Novo Belchor, Caroline F. B. Rodrigues, Daniela De Oliveira Toyama, Marcos A. De Oliveira, Danielle P. Novaes, and Marcos Hikari Toyama. 2018. "Edema Induced by a Crotalus durissus terrificus Venom Serine Protease (Cdtsp 2) Involves the PAR Pathway and PKC and PLC Activation" International Journal of Molecular Sciences 19, no. 8: 2405. https://doi.org/10.3390/ijms19082405
APA StyleCosta, C. R. C., Belchor, M. N., Rodrigues, C. F. B., Toyama, D. D. O., De Oliveira, M. A., Novaes, D. P., & Toyama, M. H. (2018). Edema Induced by a Crotalus durissus terrificus Venom Serine Protease (Cdtsp 2) Involves the PAR Pathway and PKC and PLC Activation. International Journal of Molecular Sciences, 19(8), 2405. https://doi.org/10.3390/ijms19082405