Non-Ionizing Radiation for Cardiac Human Amniotic Mesenchymal Stromal Cell Commitment: A Physical Strategy in Regenerative Medicine

 , and
, and
Abstract

1. Introduction
2. Results
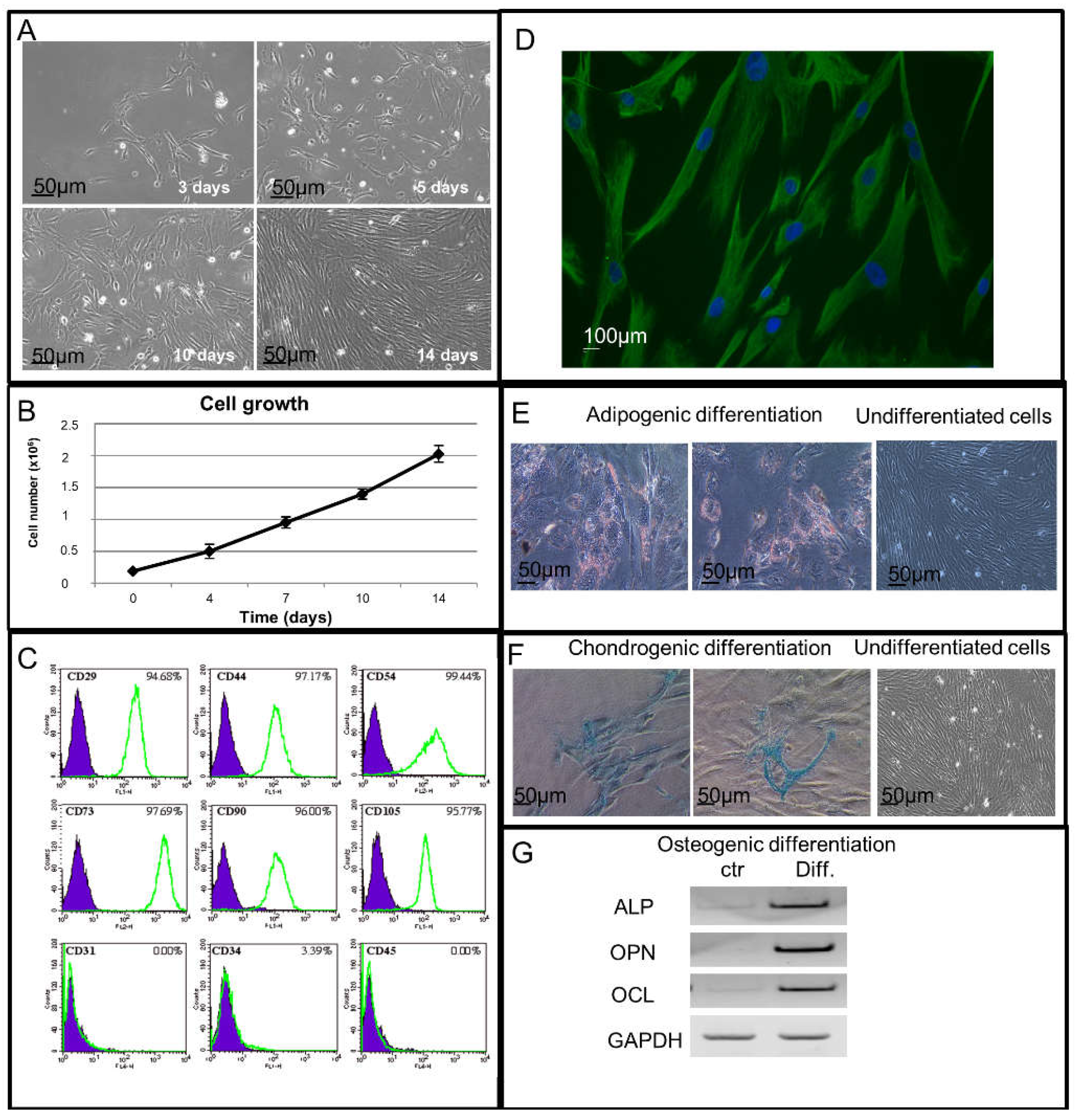
2.1. Cell Growth Study of Isolated hAMSCs
2.2. Immunophenotypical and Immunofluorescence Characterization of Isolated hAMSCs
2.3. Adipogenic, Chondrogenic and Osteogenic Potential Differentiation Study of Isolated hAMSCs
2.4. Metabolic Activity and Cell Proliferation Study of hAMSCs
2.5. Cellular Vitality and Apoptosis Study in hAMSCs
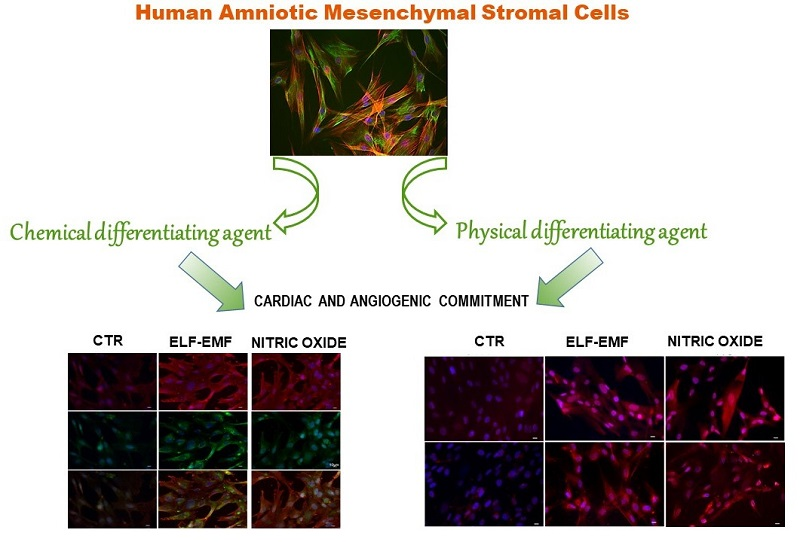
2.6. Study of Cardio-Angiogenic mRNA Expression in Chemically and Physically Treated hAMSCs
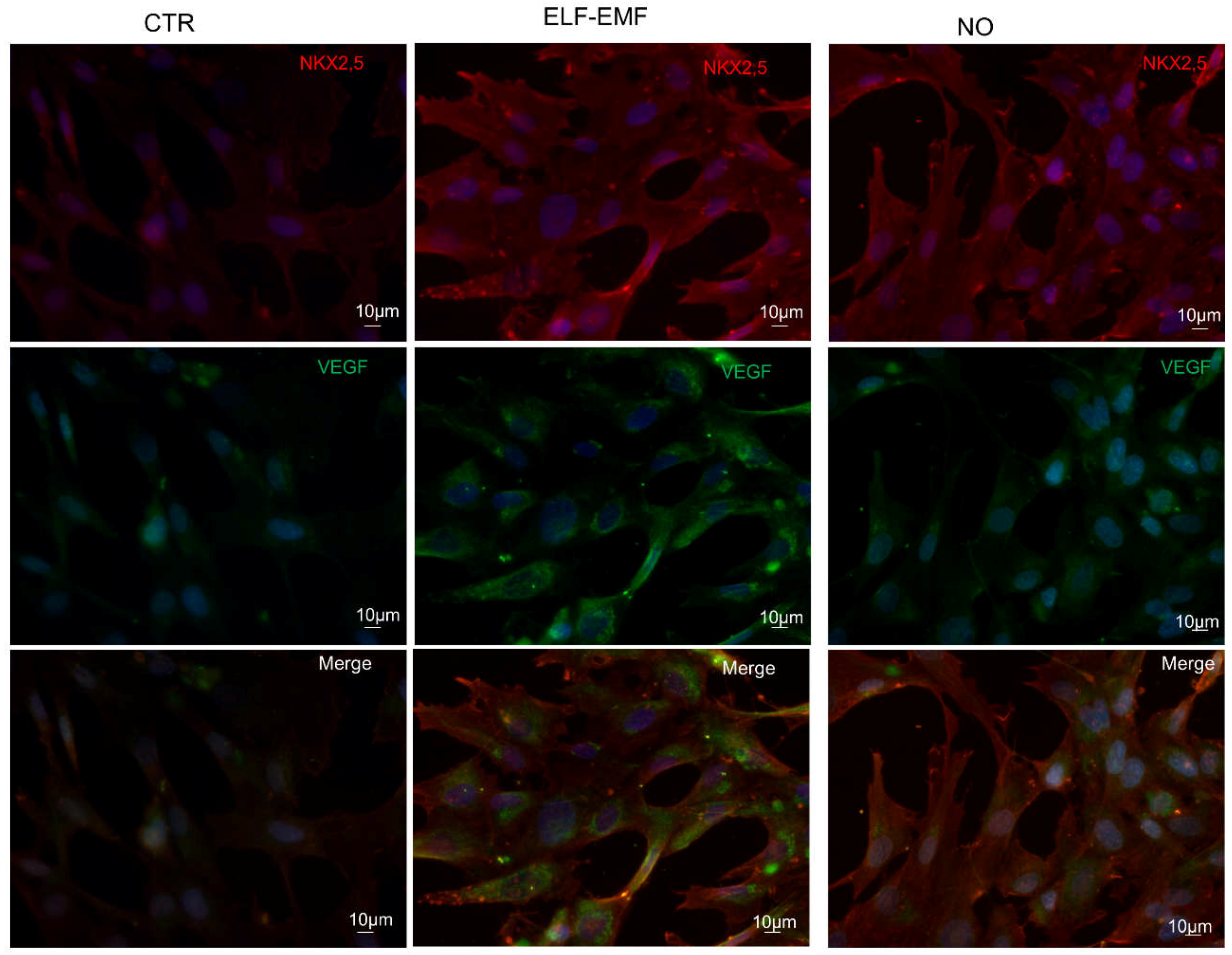
2.7. Study of Cardio-Angiogenic Protein Expression in Chemically and Physically Treated hAMSCs
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of hAMSCs
4.2. Exposure System Description and Characterization
4.4. Phase-Contrast and Immunofluorescence Analysis
4.5. hAMSCs Growth
4.6. Fluorescence-Activated Cell Sorting (FACS) Analysis
4.7. hAMSC Adipogenic, Chondrogenic and Osteogenic Differentiation Potential
4.7.1. Adipogenic Differentiation
4.7.2. Chondrogenic Differentiation
4.7.3. Osteogenic Differentiation
4.8. Cell Metabolic Activity Analysis
4.9. Cell Proliferation Analysis
4.10. Annexin-V Apoptosis Assay
4.11. RT-PCR Analysis
4.12. Real-Time Quantitative RT-PCR Analysis
4.13. Western Blotting Analysis
4.14. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bianco, P.; Robey, P.G. Stem cells in tissue engineering. Nature 2001, 414, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Kim, I.-K.; Cho, S.-W.; Cho, M.-C.; Hwang, K.-K.; Piao, H.; Piao, S.; Lim, S.H.; Hong, Y.S.; Choi, C.Y. Implantation of bone marrow mononuclear cells using injectable fibrin matrix enhances neovascularization in infarcted myocardium. Biomaterials 2005, 26, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.S.; Hwang, S.M.; Chen, K.D.; Lee, Y.S.; Hsu, L.W.; Chang, Y.J.; Wang, C.N.; Peng, H.H.; Chang, Y.L.; Chao, A.S. Functional network analysis of the transcriptomes of mesenchymal stem cells derived from amniotic fluid, amniotic membrane, cord blood, and bone marrow. Stem Cells 2007, 25, 2511–2523. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Yang, X.; Han, Z.-P.; Qu, F.-F.; Shao, L.; Shi, Y.-F. Mesenchymal stem cells: A new trend for cell therapy. Acta Pharmacol. Sin. 2013, 34, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Bianco, P.; Riminucci, M.; Gronthos, S.; Robey, P.G. Bone marrow stromal stem cells: Nature, biology, and potential applications. Stem Cells 2001, 19, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Orlic, D.; Kajstura, J.; Chimenti, S.; Jakoniuk, I.; Anderson, S.M.; Li, B.; Pickel, J.; McKay, R.; Nadal-Ginard, B.; Bodine, D.M. Bone marrow cells regenerate infarcted myocardium. Nature 2001, 410, 701–705. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.-Y.; Cho, H.-J.; Kang, H.-J.; Kim, T.-S.; Kim, M.-H.; Chung, J.-H.; Bae, J.-W.; Oh, B.-H.; Park, Y.-B.; Kim, H.-S. Pre-treatment of mesenchymal stem cells with a combination of growth factors enhances gap junction formation, cytoprotective effect on cardiomyocytes, and therapeutic efficacy for myocardial infarction. J. Am. Coll. Cardiol. 2008, 51, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Parolini, O.; Alviano, F.; Bagnara, G.P.; Bilic, G.; Buhring, H.J.R.; Evangelista, M.; Hennerbichler, S.; Liu, B.; Magatti, M.; Mao, N. Concise review: Isolation and characterization of cells from human term placenta: Outcome of the first international Workshop on Placenta Derived Stem Cells. Stem Cells 2008, 26, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Toda, A.; Okabe, M.; Yoshida, T.; Nikaido, T. The potential of amniotic membrane/amnion-derived cells for regeneration of various tissues. J. Pharmacol. Sci. 2007, 105, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Sant’Anna, E.F.; Leven, R.M.; Virdi, A.S.; Sumner, D. Effect of low intensity pulsed ultrasound and BMP2 on rat bone marrow stromal cell gene expression. J. Orthop. Res. 2005, 23, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Rego, E.B.; Inubushi, T.; Kawazoe, A.; Tanimoto, K.; Miyauchi, M.; Tanaka, E.; Takata, T.; Tanne, K. Ultrasound stimulation induces PGE2 synthesis promoting cementoblastic differentiation through EP2/EP4 receptor pathway. Ultrasound Med. Biol. 2010, 36, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.-C.; Hu, J.; Zou, S.-J.; Chen, H.-Q.; Zhou, H.-X.; Han, L.-C. Mechanical strain induces osteogenic differentiation: Cbfa1 and Ets-1 expression in stretched rat mesenchymal stem cells. Int. J. Oral Maxillofac. Surg. 2008, 37, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, L.; Hofmeister, A.; Hruska, K. Differential growth factor control of bone formation through osteoprogenitor differentiation. Bone 2004, 34, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Del Gaudio, C.; Baiguera, S.; Boieri, M.; Mazzanti, B.; Ribatti, D.; Bianco, A.; Macchiarini, P. Induction of angiogenesis using VEGF releasing genipin-crosslinked electrospun gelatin mats. Biomaterials 2013, 34, 7754–7765. [Google Scholar] [CrossRef] [PubMed]
- Qian, Q.; Qian, H.; Zhang, X.; Zhu, W.; Yan, Y.; Ye, S.; Peng, X.; Li, W.; Xu, Z.; Sun, L. 5-Azacytidine induces cardiac differentiation of human umbilical cord-derived mesenchymal stem cells by activating extracellular regulated kinase. Stem Cells Dev. 2012, 21, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Song, W.; Lu, X.; Hamilton, J.A.; Lei, M.; Peng, T.; Yee, S.-P. Development of heart failure and congenital septal defects in mice lacking endothelial nitric oxide synthase. Circulation 2002, 106, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Kanno, S.; Kim, P.K.; Sallam, K.; Lei, J.; Billiar, T.R.; Shears, L.L. Nitric oxide facilitates cardiomyogenesis in mouse embryonic stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 12277–12281. [Google Scholar] [CrossRef] [PubMed]
- Rebelatto, C.K.; Aguiar, A.M.; Senegaglia, A.C.; Aita, C.M.; Hansen, P.; Barchiki, F.; Kuligovski, C.; Olandoski, M.R.; Moutinho, J.A.; Dallagiovanna, B. Expression of cardiac function genes in adult stem cells is increased by treatment with nitric oxide agents. Biochem. Biophys. Res. Commun. 2009, 378, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Lisi, A.; Briganti, E.; Ledda, M.; Losi, P.; Grimaldi, S.; Marchese, R.; Soldani, G. A combined synthetic-fibrin scaffold supports growth and cardiomyogenic commitment of human placental derived stem cells. PLoS ONE 2012, 7, e34284. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.L.; Siriwardane, M.; Almeida-Porada, G.; Porada, C.D.; Brink, P.; Christ, G.J.; Harrison, B.S. The effect of low-frequency electromagnetic field on human bone marrow stem/progenitor cell differentiation. Stem Cell Res. 2015, 15, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Gaetani, R.; Ledda, M.; Barile, L.; Chimenti, I.; De Carlo, F.; Forte, E.; Ionta, V.; Giuliani, L.; D’Emilia, E.; Frati, G. Differentiation of human adult cardiac stem cells exposed to extremely low-frequency electromagnetic fields. Cardiovasc. Res. 2009, 82, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Ledda, M.; D’Emilia, E.; Giuliani, L.; Marchese, R.; Foletti, A.; Grimaldi, S.; Lisi, A. Nonpulsed sinusoidal electromagnetic fields as a noninvasive strategy in bone repair: The effect on human mesenchymal stem cell osteogenic differentiation. Tissue Eng. Part C Methods 2015, 21, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Ledda, M.; Megiorni, F.; Pozzi, D.; Giuliani, L.; D’Emilia, E.; Piccirillo, S.; Mattei, C.; Grimaldi, S.; Lisi, A. Non ionising radiation as a non chemical strategy in regenerative medicine: Ca2+-ICR “in vitro” effect on neuronal differentiation and tumorigenicity modulation in NT2 cells. PLoS ONE 2013, 8, e61535. [Google Scholar] [CrossRef] [PubMed]
- Cuccurazzu, B.; Leone, L.; Podda, M.V.; Piacentini, R.; Riccardi, E.; Ripoli, C.; Azzena, G.B.; Grassi, C. Exposure to extremely low-frequency (50 Hz) electromagnetic fields enhances adult hippocampal neurogenesis in C57BL/6 mice. Exp. Neurol. 2010, 226, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R.; Henderson, A.S. Exposure of salivary gland cells to low-frequency electromagnetic fields alters polypeptide synthesis. Proc. Natl. Acad. Sci. USA 1988, 85, 3928–3932. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Bañuelos, T.; Navarro, L.; González, A.; Drucker-Colín, R. Differentiation of chromaffin cells elicited by ELF MF modifies gene expression pattern. Cell Biol. Int. 2004, 28, 273–279. [Google Scholar] [CrossRef] [PubMed]
- De Carlo, F.; Ledda, M.; Pozzi, D.; Pierimarchi, P.; Zonfrillo, M.; Giuliani, L.; D’Emilia, E.; Foletti, A.; Scorretti, R.; Grimaldi, S. Nonionizing radiation as a noninvasive strategy in regenerative medicine: The effect of Ca2+-ICR on mouse skeletal muscle cell growth and differentiation. Tissue Eng. Part A 2012, 18, 2248–2258. [Google Scholar] [CrossRef] [PubMed]
- Lisi, A.; Ciotti, M.; Ledda, M.; Pieri, M.; Zona, C.; Mercanti, D.; Rieti, S.; Giuliani, L.; Grimaldi, S. Exposure to 50 Hz electromagnetic radiation promote early maturation and differentiation in newborn rat cerebellar granule neurons. J. Cell. Physiol. 2005, 204, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Ventura, C.; Maioli, M.; Asara, Y.; Santoni, D.; Mesirca, P.; Remondini, D.; Bersani, F. Turning on stem cell cardiogenesis with extremely low frequency magnetic fields. FASEB J. 2005, 19, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Czerska, E.; Casamento, J.; Ning, J.; Swicord, M.; Al-Barazi, H.; Davis, C.; Elson, E. Comparison of the effect of ELF on c-myc oncogene expression in normal and transformed human cells. Ann. N. Y. Acad. Sci. 1992, 649, 340–342. [Google Scholar] [CrossRef] [PubMed]
- Liboff, A.; Williams, T.J.; Wistar, R., Jr. Time-varying magnetic fields: Effect of DNA synthesis. Science 1984, 223, 818–821. [Google Scholar] [CrossRef] [PubMed]
- Marron, M.T.; Goodman, E.M.; Sharpe, P.T.; Greenebaum, B. Low frequency electric and magnetic fields have different effects on the cell surface. FEBS Lett. 1988, 230, 13–16. [Google Scholar] [CrossRef]
- Phillips, J.L. Effects of electromagnetic field exposure on gene transcription. J. Cell. Biochem. 1993, 51, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R.; Chizmadzhev, Y.; Shirley-Henderson, A. Electromagnetic fields and cells. J. Cell. Biochem. 1993, 51, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Sisken, B.F.; Kanje, M.; Lundborg, G.; Kurtz, W. Pulsed electromagnetic fields stimulate nerve regeneration in vitro and in vivo. Restor. Neurol. Neurosci. 1990, 1, 303–309. [Google Scholar] [PubMed]
- Alves, H.; van Ginkel, J.; Groen, N.; Hulsman, M.; Mentink, A.; Reinders, M.; van Blitterswijk, C.; de Boer, J. A mesenchymal stromal cell gene signature for donor age. PLoS ONE 2012, 7, e42908. [Google Scholar] [CrossRef] [PubMed]
- Caruso, M.; Evangelista, M.; Parolini, O. Human term placental cells: Phenotype, properties and new avenues in regenerative medicine. Int. J. Mol. Cell. Med. 2012, 1, 64. [Google Scholar] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Manni, V.; Lisi, A.; Rieti, S.; Serafino, A.; Ledda, M.; Giuliani, L.; Sacco, D.; D’Emilia, E.; Grimaldi, S. Low electromagnetic field (50 Hz) induces differentiation on primary human oral keratinocytes (HOK). Bioelectromagnetics 2004, 25, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Manni, V.; Lisi, A.; Pozzi, D.; Rieti, S.; Serafino, A.; Giuliani, L.; Grimaldi, S. Effects of extremely low frequency (50 Hz) magnetic field on morphological and biochemical properties of human keratinocytes. Bioelectromagnetics 2002, 23, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Lisi, A.; Ledda, M.; Rosola, E.; Pozzi, D.; Emilia, E.D.; Giuliani, L.; Foletti, A.; Modesti, A.; Morris, S.J.; Grimaldi, S. Extremely low frequency electromagnetic field exposure promotes differentiation of pituitary corticotrope-derived AtT20 D16V cells. Bioelectromagnetics 2006, 27, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Lednev, V. Possible mechanism for the influence of weak magnetic fields on biological systems. Bioelectromagnetics 1991, 12, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Zhadin, M.N. Combined action of static and alternating magnetic fields on ion motion in a macromolecule: Theoretical aspects. Bioelectromagnetics 1998, 19, 279–292. [Google Scholar] [CrossRef]
- Liboff, A.R. Geomagnetic cyclotron resonance in living cells. J. Biol. Phys. 1985, 13, 99–102. [Google Scholar] [CrossRef]
- Del Giudice, E.; Fleischmann, M.; Preparata, G.; Talpo, G. On the “unreasonable” effects of ELF magnetic fields upon a system of ions. Bioelectromagnetics 2002, 23, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sequence | Annealing Temperature (T) |
|---|---|---|
| VEGF | 5′-CTTGGGTGCATTGGAGCCT-3′ 5′-CTGCGCTGATAGACATCCAT-3′ | 60 °C |
| KDR | 5′-CAGCATCACCAGTAGCCAG-3′ 5′-TTATACAGATCTTCAGGAGCTT-3′ | 60 °C |
| ACTA2 | 5′-ATGAAGATCCTGACTGAGCG-3′ 5′ –GCAGTGGCCATCTCATTTTC-3′ | 60 °C |
| NKX2-5 | 5′-CAGCGACCCCGACCCAG-3′ 5′-GCTTCCTCCGCCGTCGC-3′ | 60 °C |
| MYH7 | 5′-CAGAAGAAGAAGATGGATGC-3′ 5′-CGCTGGTGTCCTGCTCCT-3′ | 60 °C |
| TnI | 5′-GGACAAGGTGGATGAAGAGA-3′ 5′-AGGGTGGGCCGCTTAAACT-3′ | 60 °C |
| GAPDH | 5′-CATCATCTCTGCCCCCTCT-3′ 5′-CAAAGTTGTCATGGATGACCT-3′ | 60 °C |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ledda, M.; D’Emilia, E.; Lolli, M.G.; Marchese, R.; De Lazzari, C.; Lisi, A. Non-Ionizing Radiation for Cardiac Human Amniotic Mesenchymal Stromal Cell Commitment: A Physical Strategy in Regenerative Medicine. Int. J. Mol. Sci. 2018, 19, 2324. https://doi.org/10.3390/ijms19082324
Ledda M, D’Emilia E, Lolli MG, Marchese R, De Lazzari C, Lisi A. Non-Ionizing Radiation for Cardiac Human Amniotic Mesenchymal Stromal Cell Commitment: A Physical Strategy in Regenerative Medicine. International Journal of Molecular Sciences. 2018; 19(8):2324. https://doi.org/10.3390/ijms19082324
Chicago/Turabian StyleLedda, Mario, Enrico D’Emilia, Maria Grazia Lolli, Rodolfo Marchese, Claudio De Lazzari, and Antonella Lisi. 2018. "Non-Ionizing Radiation for Cardiac Human Amniotic Mesenchymal Stromal Cell Commitment: A Physical Strategy in Regenerative Medicine" International Journal of Molecular Sciences 19, no. 8: 2324. https://doi.org/10.3390/ijms19082324
APA StyleLedda, M., D’Emilia, E., Lolli, M. G., Marchese, R., De Lazzari, C., & Lisi, A. (2018). Non-Ionizing Radiation for Cardiac Human Amniotic Mesenchymal Stromal Cell Commitment: A Physical Strategy in Regenerative Medicine. International Journal of Molecular Sciences, 19(8), 2324. https://doi.org/10.3390/ijms19082324