The Transcriptomic Landscape of Gastric Cancer: Insights into Epstein-Barr Virus Infected and Microsatellite Unstable Tumors


 , , and
, , and
Abstract
1. Introduction
2. Results
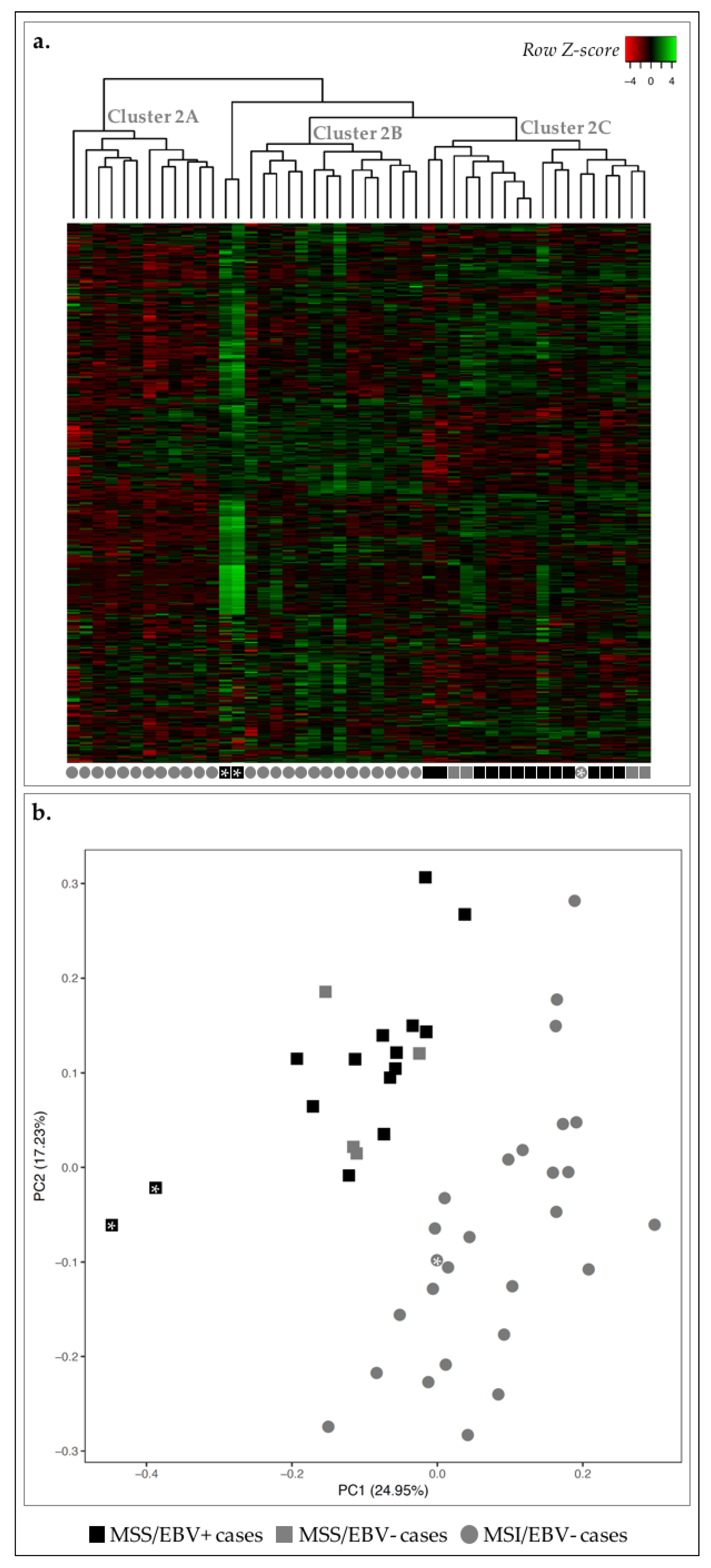
2.1. EBV+ and MSI-High GCs Displayed Distinct Transcriptomic Signatures
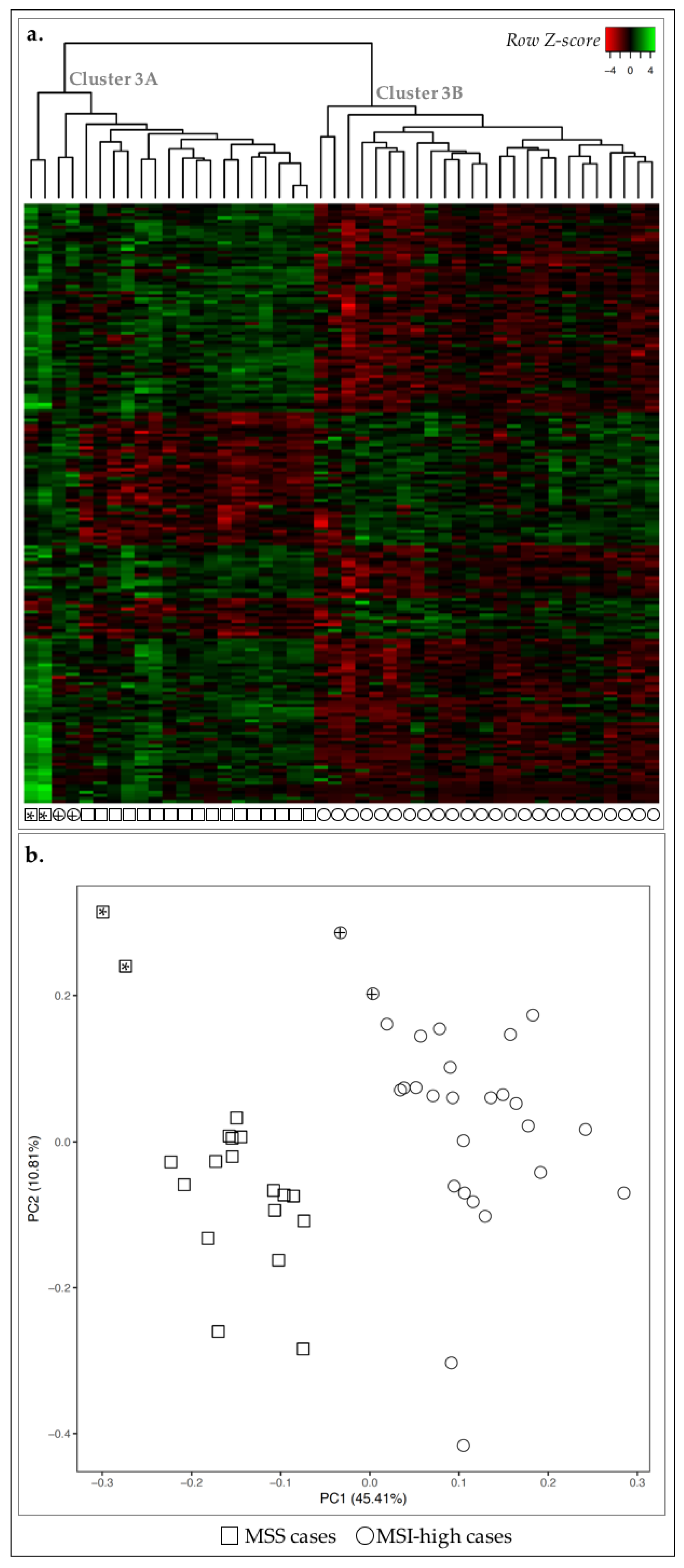
2.2. MSI-High GC Cases Displayed a Mitotic Signature, While MSS GC Cases Showed an Immune Response Signature
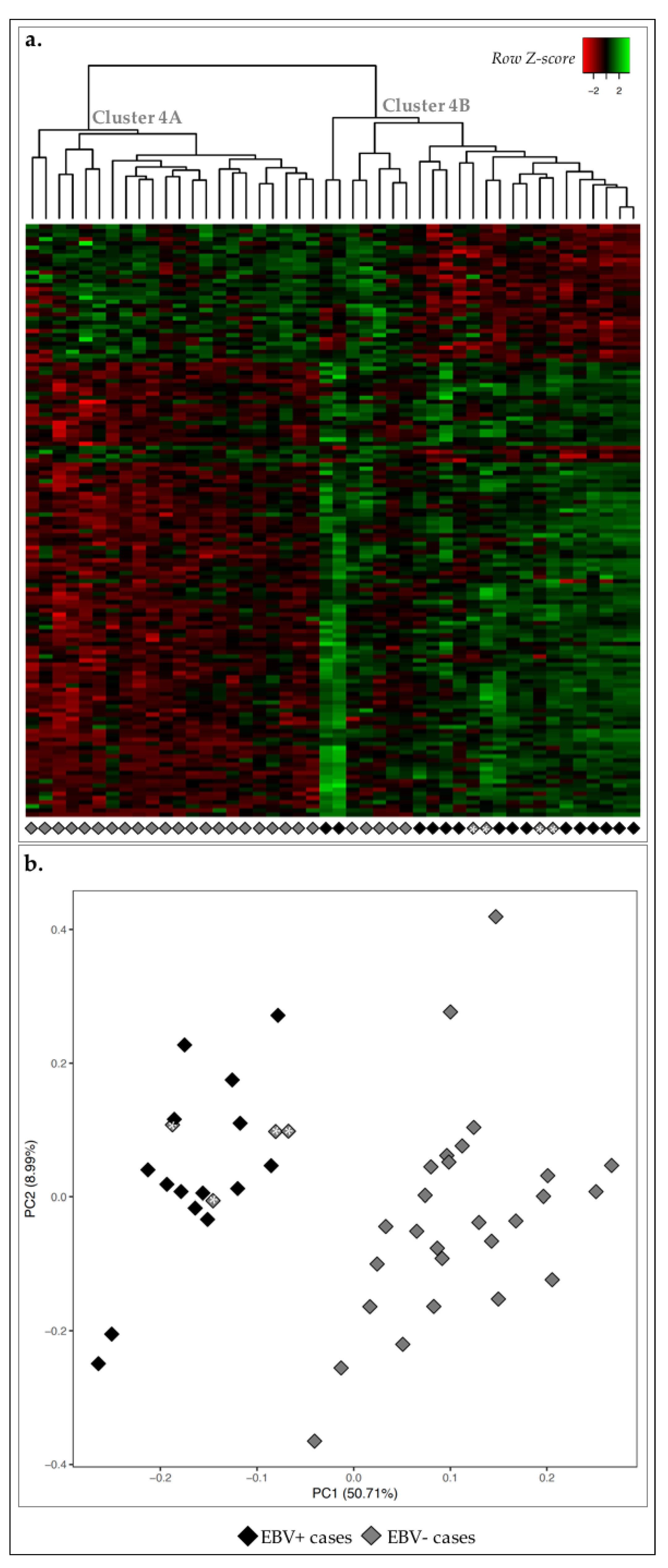
2.3. EBV+ GC Cases Were Associated with Immune Response Signature
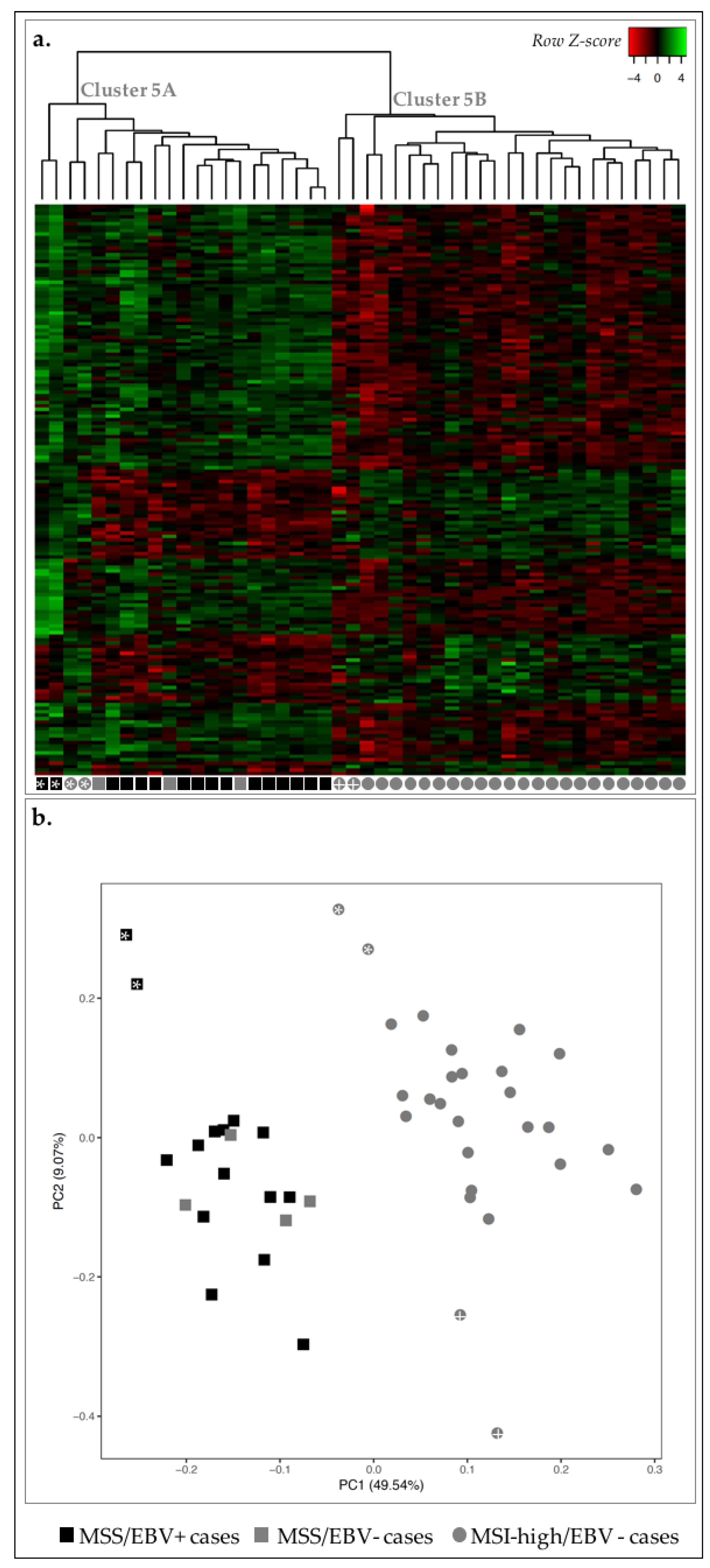
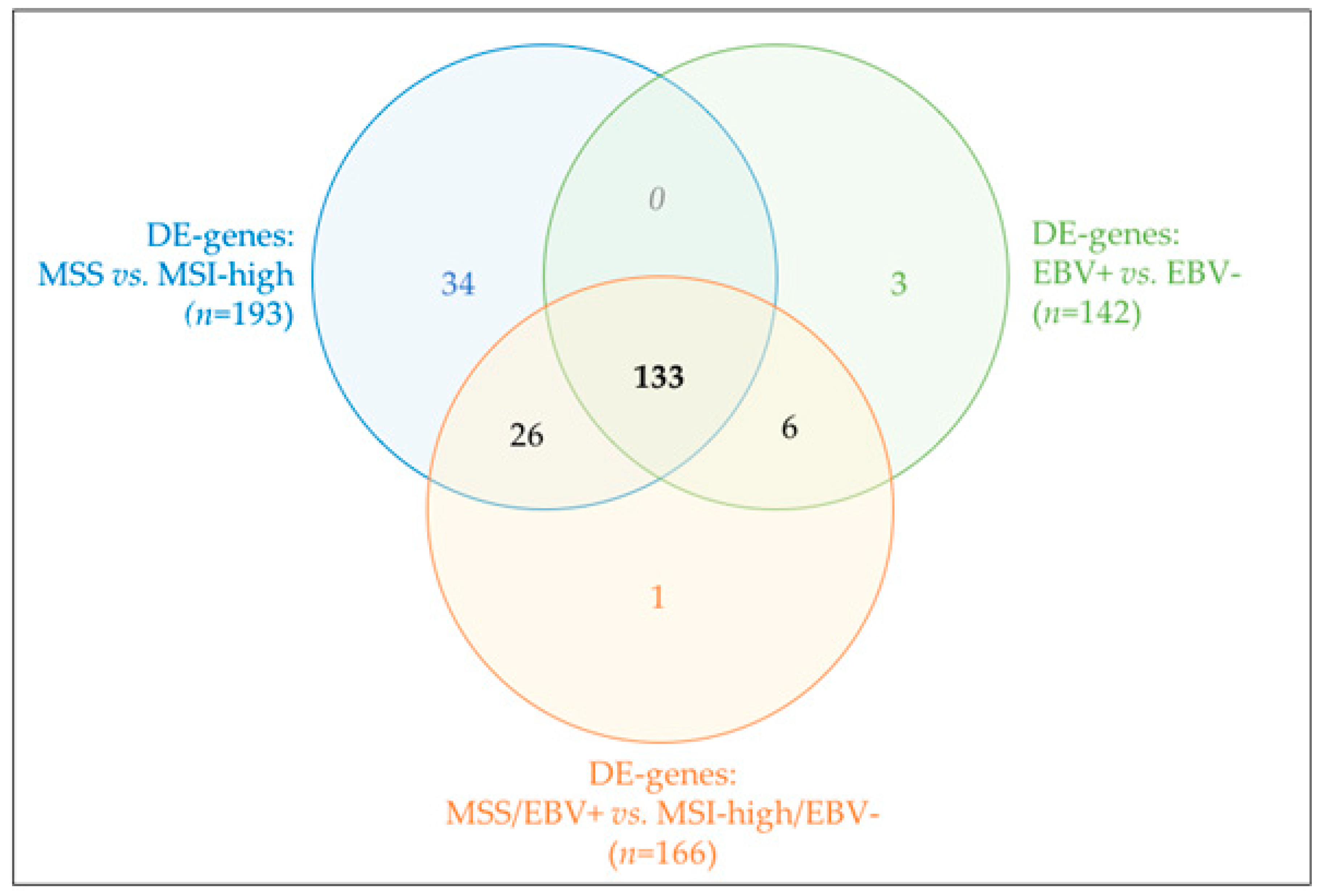
2.4. MSS/MSI Phenotype Classification Was the Major Molecular Classifier in GC
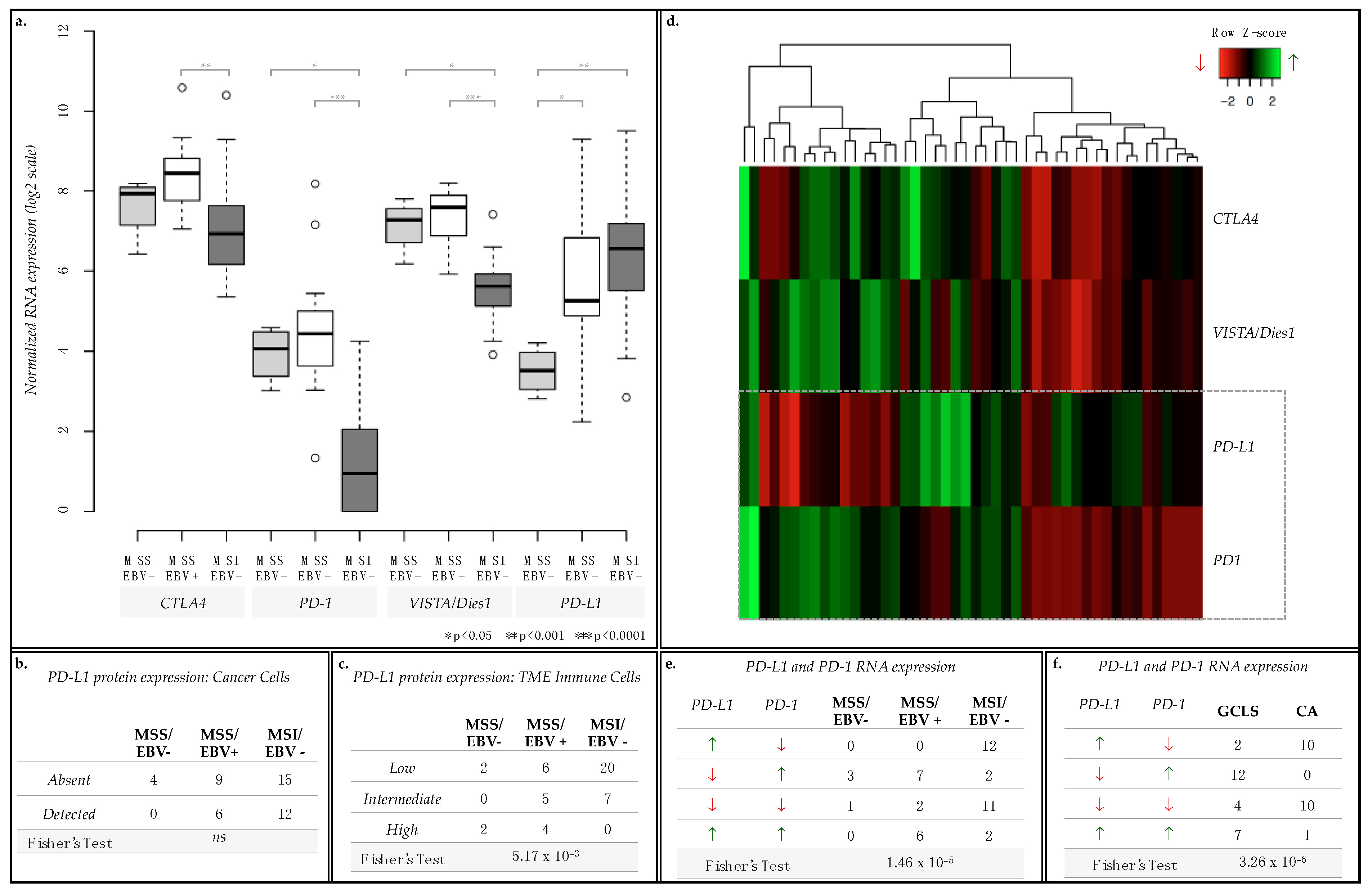
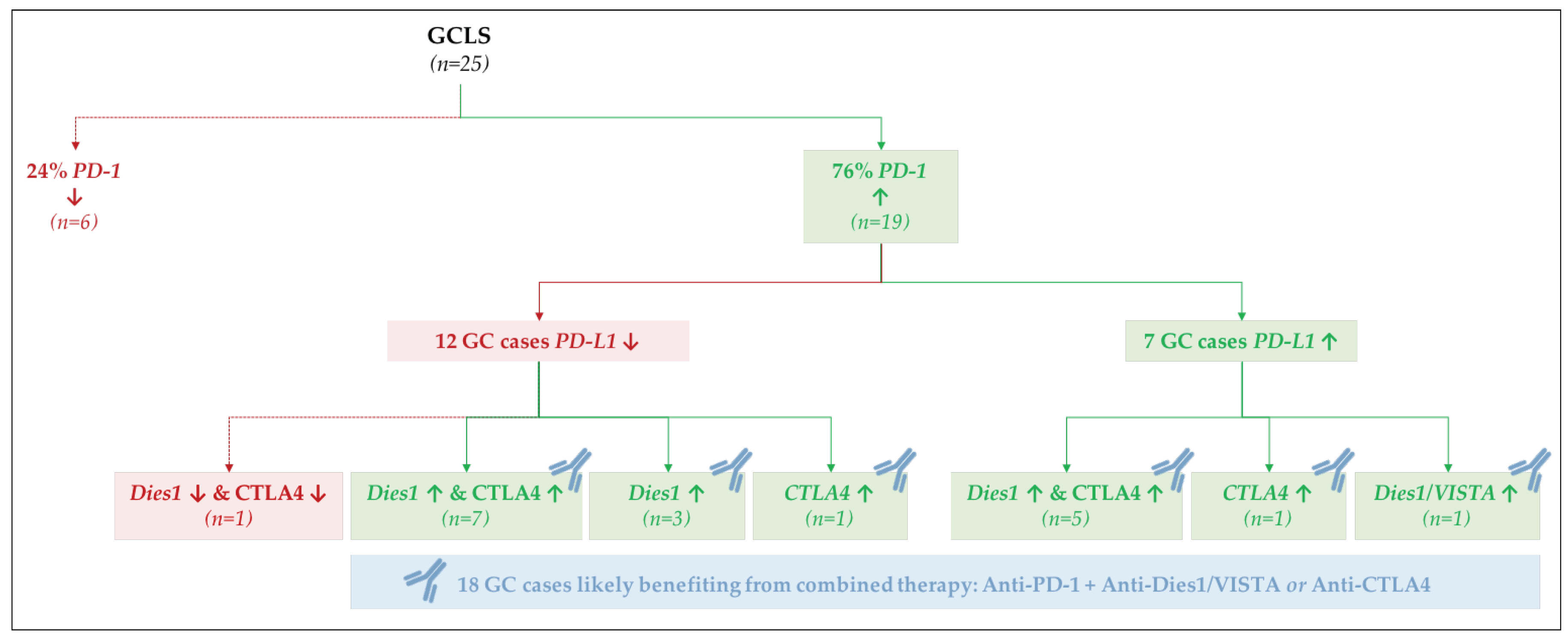
2.5. PD-L1 and PD-1 Displayed Opposite mRNA Expression Patterns and Were Differently Associated with GC Molecular Subtypes and Morphological Features
3. Discussion
4. Materials and Methods
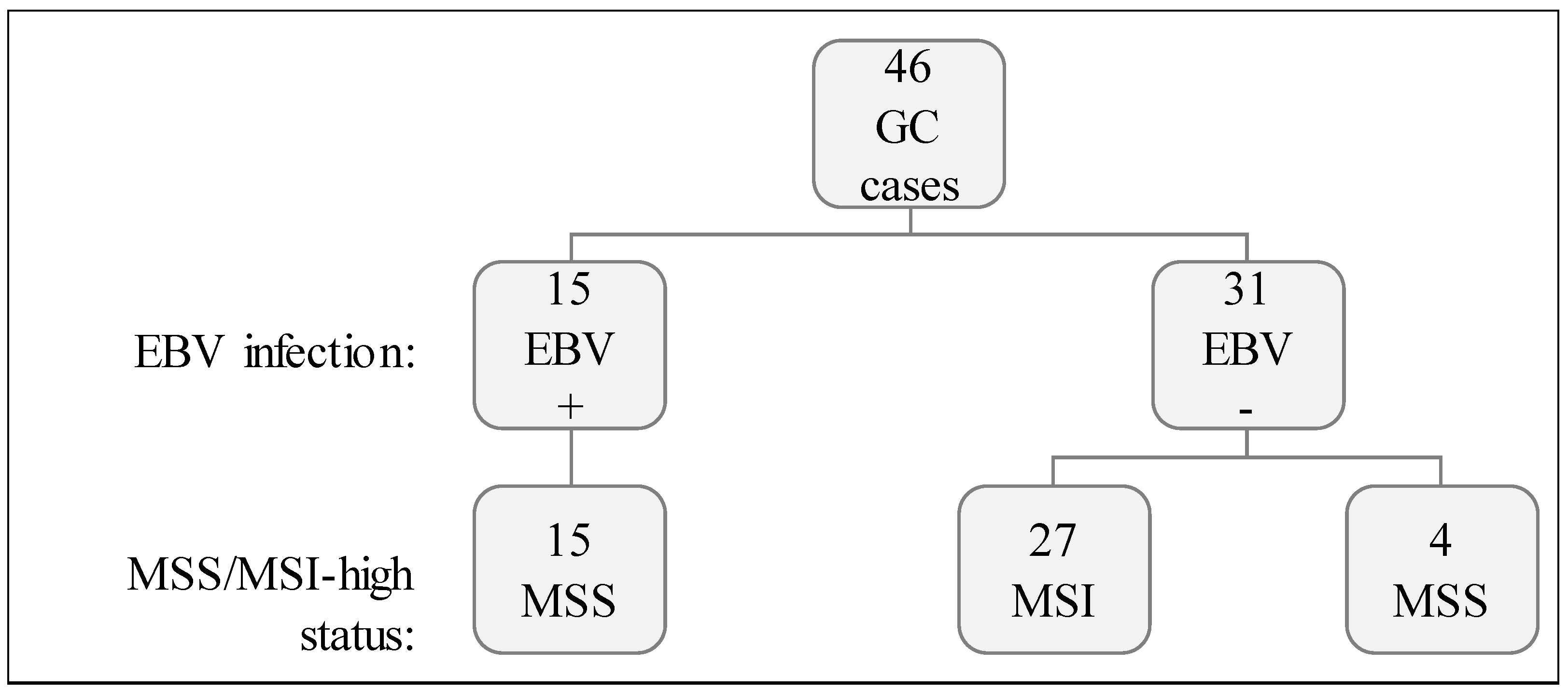
4.1. Case Series
4.2. EBV In Situ Hybridization
4.3. PCR/Fragment Analysis for MSI Status
4.4. Gene Expression Profiling by Nanostring nCounter Assay
4.5. PD-L1 Immunohistochemistry
4.6. Functional Annotation and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CA | Conventional-type Adenocarcinoma |
| DE | Differentially Expressed |
| EBV | Epstein-Barr Virus |
| ES | Enrichment Score |
| FDR | False Discovery Rate |
| GC | Gastric Cancer |
| GCLS | Gastric Cancer with Lymphoid Stroma |
| IFNγ | Interferon gamma |
| IHC | Immunohistochemistry |
| MSI-high | Microsatellite unstable |
| MSS | Microsatellite stable |
| PD-1 | Program Death 1 |
| PD-L1 | Program Death Ligand 1 |
| TCGA | The Cancer Genome Atlas |
| TME | Tumor Microenvironment |
References
- Gullo, I.; Carneiro, F.; Oliveira, C.; Almeida, G.M. Heterogeneity in Gastric Cancer: From Pure Morphology to Molecular Classifications. Pathobiology 2018, 85, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Yuen, S.T.; Leung, S.Y. Genomics Study of Gastric Cancer and Its Molecular Subtypes. Adv. Exp. Med. Biol. 2016, 908, 419–439. [Google Scholar] [CrossRef] [PubMed]
- Baraniskin, A.; Van Laethem, J.L.; Wyrwicz, L.; Guller, U.; Wasan, H.S.; Matysiak-Budnik, T.; Gruenberger, T.; Ducreux, M.; Carneiro, F.; Van Cutsem, E.; et al. Clinical relevance of molecular diagnostics in gastrointestinal (GI) cancer: European Society of Digestive Oncology (ESDO) expert discussion and recommendations from the 17th European Society for Medical Oncology (ESMO)/World Congress on Gastrointestinal Cancer, Barcelona. Eur. J. Cancer 2017, 86, 305–317. [Google Scholar] [CrossRef] [PubMed]
- TCGA. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar] [CrossRef]
- Cristescu, R.; Lee, J.; Nebozhyn, M.; Kim, K.M.; Ting, J.C.; Wong, S.S.; Liu, J.; Yue, Y.G.; Wang, J.; Yu, K.; et al. Molecular analysis of gastric cancer identifies subtypes associated with distinct clinical outcomes. Nat. Med. 2015, 21, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Sohn, B.H.; Hwang, J.E.; Jang, H.J.; Lee, H.S.; Oh, S.C.; Shim, J.J.; Lee, K.W.; Kim, E.H.; Yim, S.Y.; Lee, S.H.; et al. Clinical Significance of Four Molecular Subtypes of Gastric Cancer Identified by The Cancer Genome Atlas Project. Clinical cancer research. Off. J. Am. Assoc. Cancer Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Seruca, R.; Seixas, M.; Sobrinho-Simões, M. The Clinicopathological Features of Gastric Carcinomas with Microsatellite Instability May Be Mediated by Mutations of Different “Target Genes”: A Study of the TGFβ RII, IGFII R.; and BAX Genes. Am. J. Pathol. 1998, 153, 1211–1219. [Google Scholar] [CrossRef]
- Derks, S.; Liao, X.; Chiaravalli, A.M.; Xu, X.; Camargo, M.C.; Solcia, E.; Sessa, F.; Fleitas, T.; Freeman, G.J.; Rodig, S.J.; et al. Abundant PD-L1 expression in Epstein-Barr Virus-infected gastric cancers. Oncotarget 2016, 7, 32925–32932. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Cho, J.; Lee, J.; Kang, S.Y.; An, J.Y.; Choi, M.G.; Lee, J.H.; Sohn, T.S.; Bae, J.M.; Kim, S.; et al. Host immune response index in gastric cancer identified by comprehensive analyses of tumor immunity. Oncoimmunology 2017, 6, e1356150. [Google Scholar] [CrossRef] [PubMed]
- Solcia, E.; Klersy, C.; Mastracci, L.; Alberizzi, P.; Candusso, M.E.; Diegoli, M.; Tava, F.; Riboni, R.; Manca, R.; Luinetti, O. A combined histologic and molecular approach identifies three groups of gastric cancer with different prognosis. Virchows Arch. 2009, 455, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Grogg, K.L.; Lohse, C.M.; Pankratz, V.S.; Halling, K.C.; Smyrk, T.C. Lymphocyte-rich gastric cancer: Associations with Epstein-Barr virus, microsatellite instability, histology, and survival. Mod. Pathol. 2003, 16, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Hissong, E.; Ramrattan, G.; Zhang, P.; Zhou, X.K.; Young, G.; Klimstra, D.S.; Shia, J.; Fernandes, H.; Yantiss, R.K. Gastric Carcinomas With Lymphoid Stroma: An Evaluation of the Histopathologic and Molecular Features. Am. J. Surg. Pathol. 2018, 42, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Gullo, I.; Oliveira, P.; Athelogou, M.; Gonçalves, G.; Pinto, M.L.; Carvalho, J.; Valente, A.; Pinheiro, H.; Andrade, S.; Almeida, G.M.; et al. New insights into the inflamed tumor immune microenvironment of Gastric Cancer with Lymphoid Stroma: From morphology and digital analysis to gene expression. Gastric Cancer 2018. [Google Scholar] [CrossRef] [PubMed]
- Bosman, F.T.; Carneiro, F.; Hruban, R.H.; Theise, N.D. WHO Classification of Tumours of the Digestive System, 4th ed.; IARC: Lyon, France, 2010. [Google Scholar]
- Fuchs, C.S.; Doi, T.; Jang, R.W.-J.; Muro, K.; Satoh, T.; Machado, M.; Sun, W.; Jalal, S.I.; Shah, M.A.; Metges, J.-P.; et al. KEYNOTE-059 cohort 1: Efficacy and safety of pembrolizumab (pembro) monotherapy in patients with previously treated advanced gastric cancer. J. Clin. Oncol. 2017, 35, 4003. [Google Scholar] [CrossRef]
- FDA Approves Merck’s KEYTRUDA® (pembrolizumab) for Previously Treated Patients with Recurrent Locally Advanced or Metastatic Gastric or Gastroesophageal Junction Cancer Whose Tumors Express PD-L1 (CPS Greater Than or Equal to 1). Merck. Available online: http://investors.merck.com/home/default.aspx (accessed on 17 July 2018).
- FDA Grants Accelerated Approval to Pembrolizumab for Tissue/Site Agnostic Indication. Available online: https://www.fda.gov/drugs/informationondrugs/ approveddrugs/ucm560040.htm (accessed on 18 January 2018).
- Muro, K.; Chung, H.C.; Shankaran, V.; Geva, R.; Catenacci, D.; Gupta, S.; Eder, J.P.; Golan, T.; Le, D.T.; Burtness, B.; et al. Pembrolizumab for patients with PD-L1-positive advanced gastric cancer (KEYNOTE-012): A multicentre, open-label, phase 1b trial. Lancet Oncol. 2016, 17, 717–726. [Google Scholar] [CrossRef]
- Kang, Y.K.; Boku, N.; Satoh, T.; Ryu, M.H.; Chao, Y.; Kato, K.; Chung, H.C.; Chen, J.S.; Muro, K.; Kang, W.K.; et al. Nivolumab in patients with advanced gastric or gastro-oesophageal junction cancer refractory to, or intolerant of, at least two previous chemotherapy regimens (ONO-4538-12, ATTRACTION-2): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 390, 2461–2471. [Google Scholar] [CrossRef]
- Fuchs, C.S.; Doi, T.; Jang, R.W.; Muro, K.; Satoh, T.; Machado, M.; Sun, W.; Jalal, S.I.; Shah, M.A.; Metges, J.P.; et al. Safety and Efficacy of Pembrolizumab Monotherapy in Patients With Previously Treated Advanced Gastric and Gastroesophageal Junction Cancer: Phase 2 Clinical KEYNOTE-059 Trial. JAMA Oncol. 2018, 4, e180013. [Google Scholar] [CrossRef] [PubMed]
- Ayers, M.; Lunceford, J.; Nebozhyn, M.; Murphy, E.; Loboda, A.; Kaufman, D.R.; Albright, A.; Cheng, J.D.; Kang, S.P.; Shankaran, V.; et al. IFN-gamma-related mRNA profile predicts clinical response to PD-1 blockade. J. Clin. Investig. 2017, 127, 2930–2940. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Mellman, I. Elements of cancer immunity and the cancer-immune set point. Nature 2017, 541, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Chan, X.B.; Epstein, D.; Teh, B.T.; Kim, K.-M.; Kim, S.T.; Park, S.H.; Kang, W.K.; Rozen, S.; Lee, J.; et al. NanoString expression profiling identifies candidate biomarkers of RAD001 response in metastatic gastric cancer. ESMO Open 2016, 1, e000009. [Google Scholar] [CrossRef] [PubMed]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Boger, C.; Behrens, H.M.; Kruger, S.; Rocken, C. The novel negative checkpoint regulator VISTA is expressed in gastric carcinoma and associated with PD-L1/PD-1: A future perspective for a combined gastric cancer therapy? Oncoimmunology 2017, 6, e1293215. [Google Scholar] [CrossRef] [PubMed]
- Chiaravalli, A.M.; Cornaggia, M.; Furlan, D.; Capella, C.; Fiocca, R.; Tagliabue, G.; Klersy, C.; Solcia, E. The role of histological investigation in prognostic evaluation of advanced gastric cancer. Analysis of histological structure and molecular changes compared with invasive pattern and stage. Virchows Arch. 2001, 439, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Velho, S.; Fernandes, M.S.; Leite, M.; Figueiredo, C.; Seruca, R. Causes and consequences of microsatellite instability in gastric carcinogenesis. World J. Gastroenterol. 2014, 20, 16433–16442. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, I.; Saavedra, K.; Espinoza, J.A.; Weber, H.; Garcia, P.; Nervi, B.; Garrido, M.; Corvalan, A.H.; Roa, J.C.; Bizama, C. Molecular classification of gastric cancer: Towards a pathway-driven targeted therapy. Oncotarget 2015, 6, 24750–24779. [Google Scholar] [CrossRef] [PubMed]
- Michael-Robinson, J.M.; Reid, L.E.; Purdie, D.M.; Biemer-Huttmann, A.E.; Walsh, M.D.; Pandeya, N.; Simms, L.A.; Young, J.P.; Leggett, B.A.; Jass, J.R.; et al. Proliferation, apoptosis, and survival in high-level microsatellite instability sporadic colorectal cancer. Clinical cancer research. Off. J. Am. Assoc. Cancer Res. 2001, 7, 2347–2356. [Google Scholar]
- Shinozaki-Ushiku, A.; Kunita, A.; Fukayama, M. Update on Epstein-Barr virus and gastric cancer (review). Int. J. Oncol. 2015, 46, 1421–1434. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Park, C.; Kim, H.J.; Park, J.; Hwang, J.; Kim, J.I.; Choi, M.G.; Kim, S.; Kim, K.M.; Kang, M.S. Deregulation of immune response genes in patients with Epstein-Barr virus-associated gastric cancer and outcomes. Gastroenterology 2015, 148, 137–147.e9. [Google Scholar] [CrossRef] [PubMed]
- Strong, M.J.; Xu, G.; Coco, J.; Baribault, C.; Vinay, D.S.; Lacey, M.R.; Strong, A.L.; Lehman, T.A.; Seddon, M.B.; Lin, Z.; et al. Differences in gastric carcinoma microenvironment stratify according to EBV infection intensity: Implications for possible immune adjuvant therapy. PLoS Pathog. 2013, 9, e1003341. [Google Scholar] [CrossRef] [PubMed]
- Veldman-Jones, M.H.; Brant, R.; Rooney, C.; Geh, C.; Emery, H.; Harbron, C.G.; Wappett, M.; Sharpe, A.; Dymond, M.; Barrett, J.C.; et al. Evaluating Robustness and Sensitivity of the NanoString Technologies nCounter Platform to Enable Multiplexed Gene Expression Analysis of Clinical Samples. Cancer Res. 2015, 75, 2587–2593. [Google Scholar] [CrossRef] [PubMed]
- Takagi, S.; Kumagai, S.; Kinouchi, Y.; Hiwatashi, N.; Nagashima, F.; Takahashi, S.; Shimosegawa, T. High Ki-67 labeling index in human colorectal cancer with microsatellite instability. Anticancer Res. 2002, 22, 3241–3244. [Google Scholar] [PubMed]
- Wu, Y.; Cao, D.; Qu, L.; Cao, X.; Jia, Z.; Zhao, T.; Wang, Q.; Jiang, J. PD-1 and PD-L1 co-expression predicts favorable prognosis in gastric cancer. Oncotarget 2017, 8, 64066–64082. [Google Scholar] [CrossRef] [PubMed]
- Tie, Y.; Ma, X.; Zhu, C.; Mao, Y.; Shen, K.; Wei, X.; Chen, Y.; Zheng, H. Safety and efficacy of nivolumab in the treatment of cancers: A meta-analysis of 27 prospective clinical trials. Int. J. Cancer 2017, 140, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Goode, E.F.; Smyth, E.C. Immunotherapy for Gastroesophageal Cancer. J. Clin. Med. 2016, 5, 84. [Google Scholar] [CrossRef] [PubMed]
- Janjigian, Y.Y.; Bendell, J.C.; Calvo, E.; Kim, J.W.; Ascierto, P.A.; Sharma, P.; Ott, P.A.; Bono, P.; Jaeger, D.; Evans, T.R.J.; et al. CheckMate-032: Phase, I./II, open-label study of safety and activity of nivolumab (nivo) alone or with ipilimumab (ipi) in advanced and metastatic (A/M) gastric cancer (GC). J. Clin. Oncol. 2016, 34, 4010. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2008. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin, Germany, 2009. [Google Scholar]
- Tang, Y.; Horikoshi, M.; Li, W. Ggfortify: Unified Interface to Visualize Statistical. R J. 2016, 8, 474–485. [Google Scholar]
- Horikoshi, M.; Tang, Y. Ggfortify: Data Visualization Tools for Statistical Analysis Results. Available online: https://CRAN.R-project.org/package=ggfortify (accessed on 17 July 2018).
- Boger, C.; Behrens, H.M.; Mathiak, M.; Kruger, S.; Kalthoff, H.; Rocken, C. PD-L1 is an independent prognostic predictor in gastric cancer of Western patients. Oncotarget 2016, 7, 24269–24283. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Value of k | Cluster ID | MSI Status | EBV Infection | ||
|---|---|---|---|---|---|
| MSI-High (n = 27) | MSS (n = 19) | EBV+ (n = 15) | EBV− (n = 31) | ||
| 2 | I | 25 | 1 | 1 | 26 |
| II | 2 | 18 | 14 | 6 | |
| 3 | I | 27 | 0 | 0 | 27 |
| II | 0 | 2 | 2 | 0 | |
| III | 0 | 17 | 13 | 4 | |
| 4 | I | 13 | 0 | 0 | 13 |
| II | 14 | 0 | 0 | 14 | |
| III | 0 | 17 | 13 | 4 | |
| IV | 0 | 2 | 2 | 0 | |
| 5 | I | 0 | 12 | 9 | 3 |
| II | 12 | 0 | 0 | 12 | |
| III | 0 | 2 | 2 | 0 | |
| IV | 12 | 0 | 0 | 12 | |
| V | 3 | 5 | 4 | 4 | |
| Biological Term | Count | FDR |
|---|---|---|
| Signal Peptide | 73 | 1.24 × 10−11 |
| Disulfide bond | 64 | 1.05 × 10−09 |
| Secreted | 44 | 2.67 × 10−7 |
| GO:0042127~regulation of cell proliferation | 31 | 1.77 × 10−6 |
| GO:0006935~chemotaxis | 15 | 3.16 × 10−6 |
| GO:0042330~taxis | 15 | 3.16 × 10−6 |
| IPR001811:Small chemokine, interleukin-8-like | 9 | 8.69 × 10−6 |
| GO:0007626~locomotory behavior | 18 | 1.08 × 10−5 |
| GO:0045321~leukocyte activation | 17 | 1.26 × 10−5 |
| GO:0008009~chemokine activity | 9 | 2.10 × 10−5 |
| Cluster ID | Term | Count | FDR |
|---|---|---|---|
| Upregulated DE-genes in MSI-high vs. MSS cases (n = 55) | |||
| Cluster 1: ES = 9.5 | GO:0022402~cell cycle process | 17 | 1.46 × 10−8 |
| GO:0000279~M phase | 14 | 3.24 × 10−8 | |
| GO:0000280~nuclear division | 12 | 1.17 × 10−7 | |
| GO:0007067~mitosis | 12 | 1.17 × 10−7 | |
| GO:0000278~mitotic cell cycle | 14 | 1.40 × 10−7 | |
| GO:0000087~M phase of mitotic cell cycle | 12 | 1.43 × 10−7 | |
| GO:0048285~organelle fission | 12 | 1.81 × 10−7 | |
| GO:0022403~cell cycle phase | 14 | 5.62 × 10−7 | |
| mitosis | 10 | 1.09 × 10−6 | |
| GO:0007049~cell cycle | 17 | 1.62 × 10−6 | |
| cell division | 11 | 1.48 × 10−6 | |
| cell cycle | 12 | 2.50 × 10−5 | |
| GO:0051301~cell division | 11 | 4.02 × 10−5 | |
| Cluster 2: ES = 4.3 | GO:0005819~spindle | 9 | 2.00 × 10−5 |
| GO:0015630~microtubule cytoskeleton | 10 | 5.42 × 10−2 | |
| GO:0044430~cytoskeletal part | 10 | 3.05 | |
| GO:0005856~cytoskeleton | 12 | 3.72 | |
| Cluster 3: ES = 3.7 | GO:0007052~mitotic spindle organization | 4 | 2.13 × 10−2 |
| GO:0007051~spindle organization | 4 | 6.18 × 10−1 | |
| GO:0000226~microtubule cytoskeleton organization | 5 | 1.92 | |
| Downregulated DE-genes in MSI-high vs. MSS cases (n = 138) | |||
| Cluster 1: ES = 16.0 | disulfide bond | 57 | 1.67 × 10−13 |
| signal peptide | 61 | 1.67 × 10−13 | |
| signal | 61 | 1.44 × 10−13 | |
| disulfide bond | 57 | 5.66 × 10−13 | |
| Cluster 2: ES = 9.7 | GO:0007626~locomotory behavior | 17 | 2.98 × 10−7 |
| GO:0042330~taxis | 14 | 3.03 × 10−7 | |
| GO:0006935~chemotaxis | 14 | 3.03 × 10−7 | |
| Cluster 3: ES = 8.2 | GO:0046649~lymphocyte activation | 14 | 4.51 × 10−6 |
| GO:0045321~leukocyte activation | 15 | 5.12 × 10−6 | |
| GO:0001775~cell activation | 15 | 4.50 × 10−5 | |
| Cluster 4: ES = 5.4 | IPR001811: Small chemokine, interleukin-8-like | 8 | 1.75 × 10−5 |
| GO:0008009~chemokine activity | 8 | 5.02 × 10−5 | |
| GO:0042379~chemokine receptor binding | 8 | 7.91 × 10−5 | |
| SM00199:SCY | 8 | 1.88 × 10−4 | |
| cytokine | 9 | 2.47 × 10−2 | |
| 109.Chemokine_families | 8 | 6.13 × 10−2 | |
| GO:0005125~cytokine activity | 9 | 1.18 × 10−1 | |
| hsa04062:Chemokine signaling pathway | 10 | 4.13 × 10−1 | |
| hsa04060:Cytokine-cytokine receptor interaction | 11 | 1.19 | |
| Cluster 5: ES = 5.1 | GO:0009719~response to endogenous stimulus | 15 | 2.99 × 10−3 |
| GO:0009725~response to hormone stimulus | 14 | 5.54 × 10−3 | |
| GO:0010033~response to organic substance | 17 | 1.38 × 10−1 | |
| Cluster 6: ES = 4.9 | GO:0043067~regulation of programmed cell death | 20 | 1.27 × 10−2 |
| GO:0010941~regulation of cell death | 20 | 1.34 × 10−2 | |
| GO:0042981~regulation of apoptosis | 19 | 4.16 × 10−2 | |
| Cluster 7: ES = 4.9 | GO:0016477~cell migration | 12 | 1.10 × 10−2 |
| GO:0006928~cell motion | 15 | 1.88 × 10−2 | |
| GO:0048870~cell motility | 12 | 2.99 × 10−2 | |
| GO:0051674~localization of cell | 12 | 2.99 × 10−2 | |
| Cluster 8: ES = 4.3 | GO:0030247~polysaccharide binding | 9 | 2.18 × 10−2 |
| GO:0001871~pattern binding | 9 | 2.18 × 10−2 | |
| GO:0005539~glycosaminoglycan binding | 8 | 9.63 × 10−2 | |
| GO:0030246~carbohydrate binding | 11 | 3.40 × 10−1 | |
| Biological Term | Count | FDR |
|---|---|---|
| signal peptide | 54 | 1.38 × 10−8 |
| IPR001811:Small chemokine, interleukin-8-like | 10 | 1.40 × 10−8 |
| GO:0042330~taxis | 15 | 3.13 × 10−8 |
| GO:0006935~chemotaxis | 15 | 3.13 × 10−8 |
| disulfide bond | 49 | 5.16 × 10−8 |
| GO:0008009~chemokine activity | 10 | 6.98 × 10−8 |
| GO:0042379~chemokine receptor binding | 10 | 1.28 × 10−7 |
| SM00199:SCY | 10 | 1.99 × 10−7 |
| GO:0007626~locomotory behavior | 17 | 4.65 × 10−7 |
| GO:0006955~immune response | 25 | 5.25 × 10−7 |
| Cluster ID | Term | Count | FDR |
|---|---|---|---|
| Upregulated DE-genes in EBV+ vs. EBV− cases (n = 105) | |||
| Cluster 1: ES = 12.2 | disulfide bond | 44 | 1.36 × 10−10 |
| disulfide bond | 44 | 3.56 × 10−10 | |
| signal | 45 | 2.94 × 10−9 | |
| signal peptide | 45 | 4.07 × 10−9 | |
| Cluster 2: ES = 10.3 | GO:0042330~taxis | 14 | 5.99 × 10−9 |
| GO:0006935~chemotaxis | 14 | 5.99 × 10−9 | |
| GO:0007626~locomotory behavior | 16 | 3.41 × 10−8 | |
| GO:0007610~behavior | 16 | 5.57 × 10−5 | |
| Cluster 3: ES = 10 | GO:0045321~leukocyte activation | 15 | 8.26 × 10−8 |
| GO:0046649~lymphocyte activation | 14 | 9.54 × 10−8 | |
| GO:0042110~T-cell activation | 12 | 1.43 × 10−7 | |
| GO:0001775~cell activation | 15 | 7.88 × 10−7 | |
| Cluster 4: ES = 7.4 | IPR001811:Small chemokine, interleukin-8-like | 9 | 3.94 × 10−8 |
| GO:0008009~chemokine activity | 9 | 1.52 × 10−7 | |
| GO:0042379~chemokine receptor binding | 9 | 2.60 × 10−7 | |
| SM00199:SCY | 9 | 4.73 × 10−7 | |
| cytokine | 10 | 2.20 × 10−4 | |
| GO:0005125~cytokine activity | 10 | 1.39 × 10−3 | |
| hsa04062:Chemokine signaling pathway | 11 | 1.09 × 10−2 | |
| 109.Chemokine_families | 9 | 1.22 × 10−2 | |
| hsa04060:Cytokine-cytokine receptor interaction | 12 | 3.67 × 10−2 | |
| Cluster 5: ES = 5.2 | GO:0002520~immune system development | 11 | 4.50 × 10−3 |
| GO:0030097~hemopoiesis | 10 | 9.75 × 10−3 | |
| GO:0002521~leukocyte differentiation | 8 | 1.28 × 10−2 | |
| GO:0048534~hemopoietic or lymphoid organ development | 10 | 2.13 × 10−2 | |
| Cluster 6: ES = 5.2 | GO:0030217~T-cell differentiation | 7 | 2.48 × 10−3 |
| GO:0002521~leukocyte differentiation | 8 | 1.28 × 10−2 | |
| GO:0030098~lymphocyte differentiation | 7 | 3.64 × 10−2 | |
| Downregulated DE-genes in EBV+ vs. EBV− cases (n = 137) | |||
| Cluster 1: ES = 5.2 | GO:0000279~M phase | 8 | 5.32 × 10−3 |
| cell division | 7 | 4.85 × 10−3 | |
| GO:0007067~mitosis | 7 | 7.12 × 10−3 | |
| GO:0000280~nuclear division | 7 | 7.12 × 10−3 | |
| GO:0000087~M phase of mitotic cell cycle | 7 | 7.90 × 10−3 | |
| GO:0048285~organelle fission | 7 | 8.97 × 10−3 | |
| cell cycle | 8 | 9.93 × 10−3 | |
| mitosis | 6 | 1.38 × 10−2 | |
| GO:0051301~cell division | 7 | 3.79 × 10−2 | |
| Cluster 2: ES = 5 | GO:0022402~cell cycle process | 11 | 1.46 × 10−4 |
| GO:0005819~spindle | 7 | 5.61 × 10−4 | |
| GO:0000278~mitotic cell cycle | 9 | 8.25 × 10−4 | |
| GO:0022403~cell cycle phase | 9 | 1.93 × 10−3 | |
| GO:0007049~cell cycle | 11 | 2.74 × 10−3 | |
| GO:0015630~microtubule cytoskeleton | 8 | 1.20 × 10−1 | |
| GO:0044430~cytoskeletal part | 8 | 3.21 | |
| GO:0005856~cytoskeleton | 9 | 6.62 | |
| Biological Term | Count | FDR |
|---|---|---|
| IPR001811:Small chemokine, interleukin-8-like | 10 | 6.17 × 10−8 |
| signal peptide | 58 | 1.97 × 10−7 |
| GO:0008009~chemokine activity | 10 | 2.18 × 10−7 |
| GO:0006935~chemotaxis | 15 | 3.16 × 10−7 |
| GO:0042330~taxis | 15 | 3.16 × 10−7 |
| disulfide bond | 53 | 3.22 × 10−7 |
| GO:0042379~chemokine receptor binding | 10 | 3.97 × 10−7 |
| SM00199:SCY | 10 | 8.05 × 10−7 |
| disulfide bond | 53 | 8.56 × 10−7 |
| GO:0045321~leukocyte activation | 17 | 9.73 × 10−7 |
| Cluster ID | Term | Count | FDR |
|---|---|---|---|
| Upregulated DE-genes in MSS/EBV+ cases (n = 117) | |||
| Cluster 1: ES = 12.5 | disulfide bond | 47 | 1.12 × 10−10 |
| disulfide bond | 47 | 3.06 × 10−10 | |
| signal | 49 | 7.15 × 10−10 | |
| signal peptide | 49 | 1.03 × 10−9 | |
| Cluster 2: ES = 9.6 | GO:0042330~taxis | 14 | 2.32 × 10−8 |
| GO:0006935~chemotaxis | 14 | 2.32 × 10−8 | |
| GO:0007626~locomotory behavior | 16 | 1.58 × 10−7 | |
| GO:0007610~behavior | 16 | 2.31 × 10−4 | |
| Cluster 3: ES = 8.3 | GO:0045321~leukocyte activation | 14 | 4.01 × 10−6 |
| GO:0046649~lymphocyte activation | 13 | 4.72 × 10−6 | |
| GO:0042110~T-cell activation | 11 | 7.75 × 10−6 | |
| GO:0001775~cell activation | 14 | 3.11 × 10−5 | |
| Cluster 4: ES = 6.8 | IPR001811:Small chemokine, interleukin-8-like | 9 | 9.74 × 10−8 |
| GO:0008009~chemokine activity | 9 | 3.30 × 10−7 | |
| GO:0042379~chemokine receptor binding | 9 | 5.62 × 10−7 | |
| SM00199:SCY | 9 | 1.34 × 10−6 | |
| cytokine | 9 | 5.94 × 10−3 | |
| 109.Chemokine_families | 9 | 4.48 × 10−3 | |
| hsa04062:Chemokine signaling pathway | 11 | 1.87 × 10−2 | |
| GO:0005125~cytokine activity | 9 | 2.77 × 10−2 | |
| hsa04060:Cytokine-cytokine receptor interaction | 11 | 3.23 × 10−1 | |
| Cluster 5: ES = 4.6 | GO:0001725~stress fiber | 5 | 2.38 × 10−2 |
| GO:0032432~actin filament bundle | 5 | 3.31 × 10−2 | |
| GO:0042641~actomyosin | 5 | 3.87 × 10−2 | |
| Downregulated DE-genes in MSS/EBV+ cases (n = 49) | |||
| Cluster 1: ES = 9.9 | GO:0000278~mitotic cell cycle | 14 | 2.51 × 10−8 |
| GO:0000280~nuclear division | 12 | 2.79 × 10−8 | |
| GO:0007067~mitosis | 12 | 2.79 × 10−8 | |
| GO:0000087~M phase of mitotic cell cycle | 12 | 3.39 × 10−8 | |
| GO:0048285~organelle fission | 12 | 4.30 × 10−8 | |
| GO:0022403~cell cycle phase | 14 | 1.02 × 10−7 | |
| GO:0000279~M phase | 13 | 1.15 × 10−7 | |
| mitosis | 10 | 3.25 × 10−7 | |
| cell division | 11 | 3.84 × 10−7 | |
| cell cycle | 12 | 5.83 × 10−6 | |
| GO:0051301~cell division | 11 | 1.15 × 10−5 | |
| Cluster 2: ES = 3.5 | GO:0005819~spindle | 7 | 4.12 × 10−3 |
| GO:0015630~microtubule cytoskeleton | 9 | 1.60 × 10−1 | |
| GO:0005856~cytoskeleton | 11 | 5.27 | |
| GO:0044430~cytoskeletal part | 9 | 5.64 | |
| mRNA Expression | MSS/EBV− (n = 4) | MSS/EBV+ (n = 15) | MSI/EBV− (n = 27) | ||||
|---|---|---|---|---|---|---|---|
| PD-L1 | PD-1 | CA (n = 0) | GCLS (n = 4) | CA (n = 0) | GCLS (n = 15) | CA (n = 21) | GCLS (n = 6) |
| ↗ | ↙ | 0 | 0 | 0 | 0 | 10 | 2 |
| ↙ | ↗ | 0 | 3 | 0 | 7 | 0 | 2 |
| ↙ | ↙ | 0 | 1 | 0 | 2 | 10 | 1 |
| ↗ | ↗ | 0 | 0 | 0 | 6 | 1 | 1 |
| Fisher’s Exact Test: p = 3.71 × 10−6 | |||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gullo, I.; Carvalho, J.; Martins, D.; Lemos, D.; Monteiro, A.R.; Ferreira, M.; Das, K.; Tan, P.; Oliveira, C.; Carneiro, F.; et al. The Transcriptomic Landscape of Gastric Cancer: Insights into Epstein-Barr Virus Infected and Microsatellite Unstable Tumors. Int. J. Mol. Sci. 2018, 19, 2079. https://doi.org/10.3390/ijms19072079
Gullo I, Carvalho J, Martins D, Lemos D, Monteiro AR, Ferreira M, Das K, Tan P, Oliveira C, Carneiro F, et al. The Transcriptomic Landscape of Gastric Cancer: Insights into Epstein-Barr Virus Infected and Microsatellite Unstable Tumors. International Journal of Molecular Sciences. 2018; 19(7):2079. https://doi.org/10.3390/ijms19072079
Chicago/Turabian StyleGullo, Irene, Joana Carvalho, Diana Martins, Diana Lemos, Ana Rita Monteiro, Marta Ferreira, Kakoli Das, Patrick Tan, Carla Oliveira, Fátima Carneiro, and et al. 2018. "The Transcriptomic Landscape of Gastric Cancer: Insights into Epstein-Barr Virus Infected and Microsatellite Unstable Tumors" International Journal of Molecular Sciences 19, no. 7: 2079. https://doi.org/10.3390/ijms19072079
APA StyleGullo, I., Carvalho, J., Martins, D., Lemos, D., Monteiro, A. R., Ferreira, M., Das, K., Tan, P., Oliveira, C., Carneiro, F., & Oliveira, P. (2018). The Transcriptomic Landscape of Gastric Cancer: Insights into Epstein-Barr Virus Infected and Microsatellite Unstable Tumors. International Journal of Molecular Sciences, 19(7), 2079. https://doi.org/10.3390/ijms19072079