Ca2+/Calmodulin-Dependent AtSR1/CAMTA3 Plays Critical Roles in Balancing Plant Growth and Immunity

Abstract

1. Introduction
2. Results
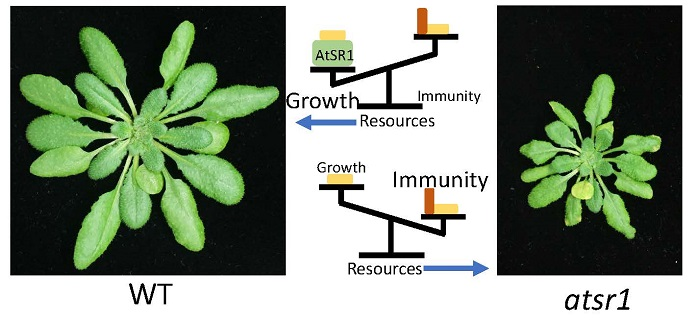
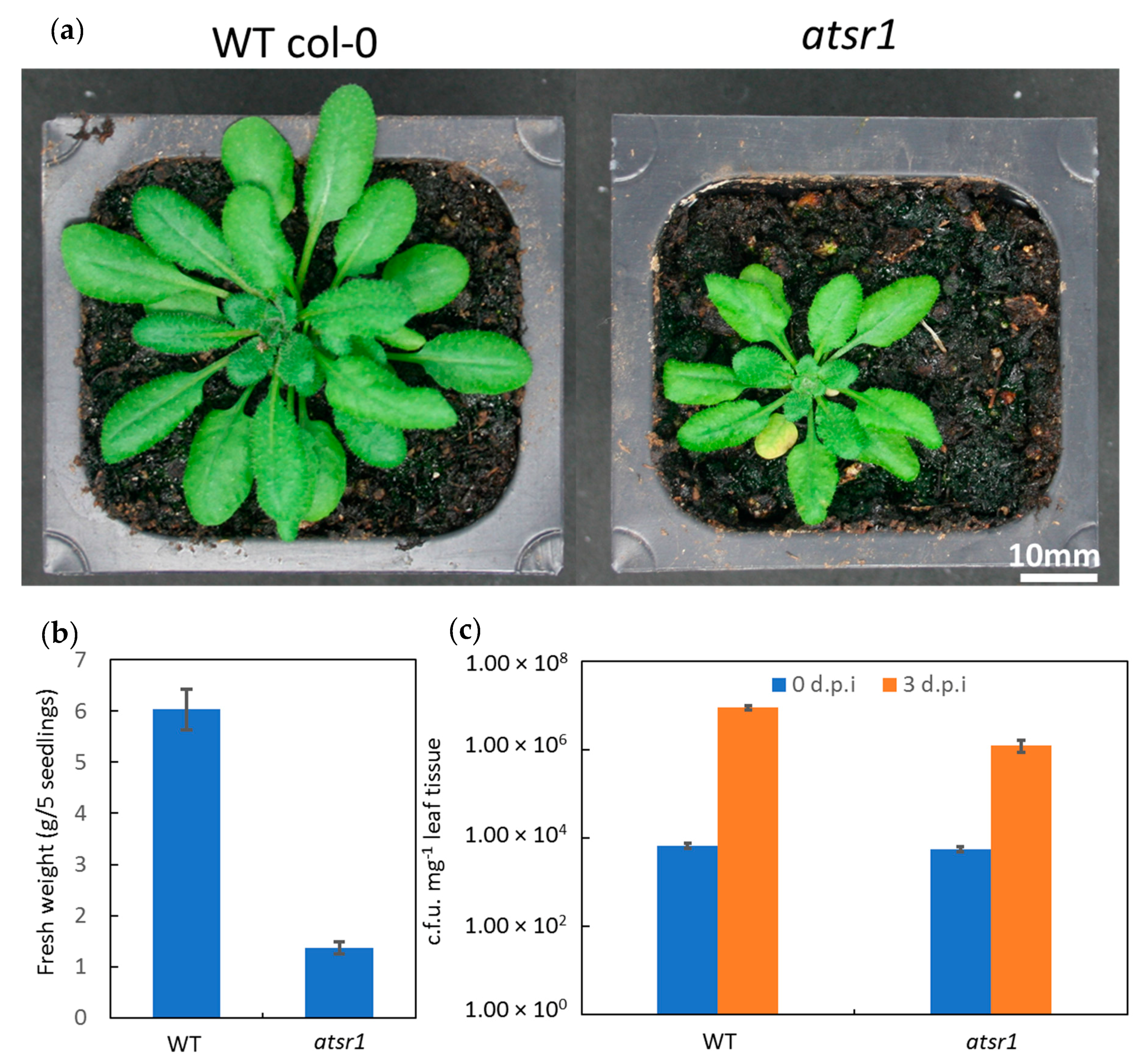
2.1. AtSR1 Is a Negative Regulator of Immune Response and a Putative Positive Regulator of Growth and Development in Plants
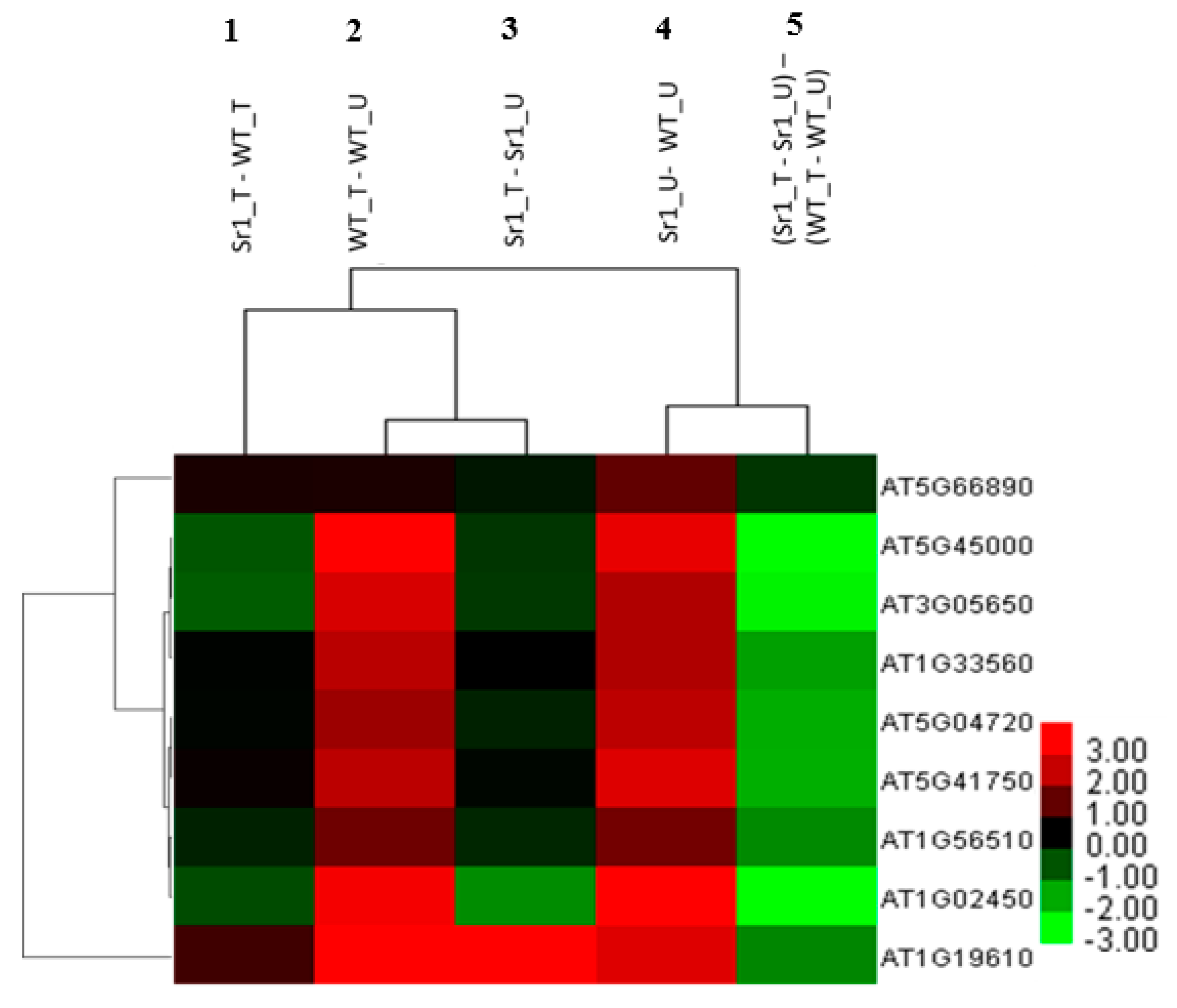
2.2. AtSR1-Mediated Transcriptome Landscape in Plant Growth and Immune Responses
2.3. AtSR1 Negatively Regulates Plant Immune Pathway in the Absence of Pathogen
2.4. AtSR1-Regulated Immune Pathway Response to Pathogen Attack
2.5. AtSR1 Positively Regulates Auxin- and BRs-Mediated Plant growth and Development in the Absence of Pathogen
2.6. AtSR1-Regulates Plant Growth-Related Pathway in Response to Pathogen Attack
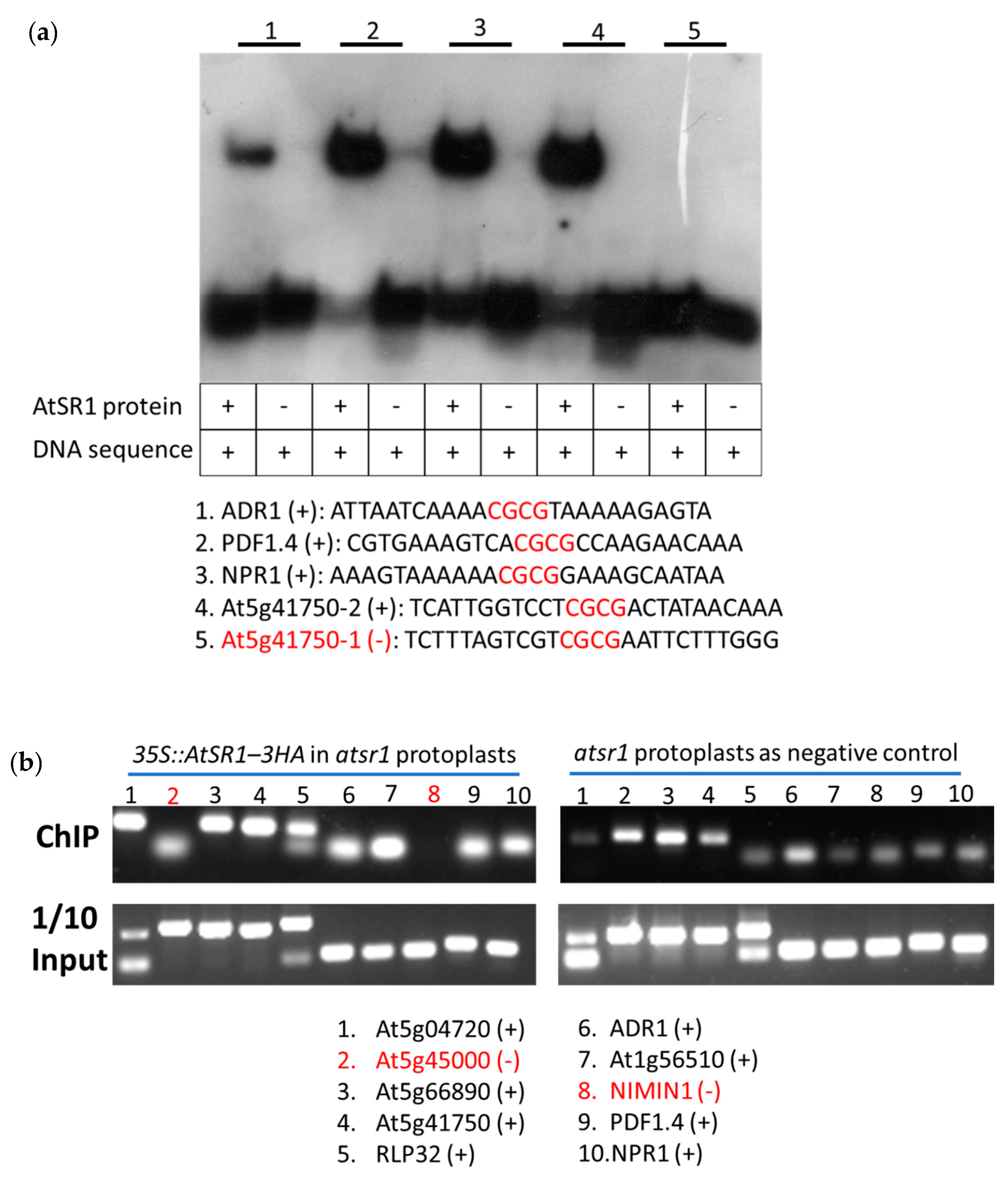
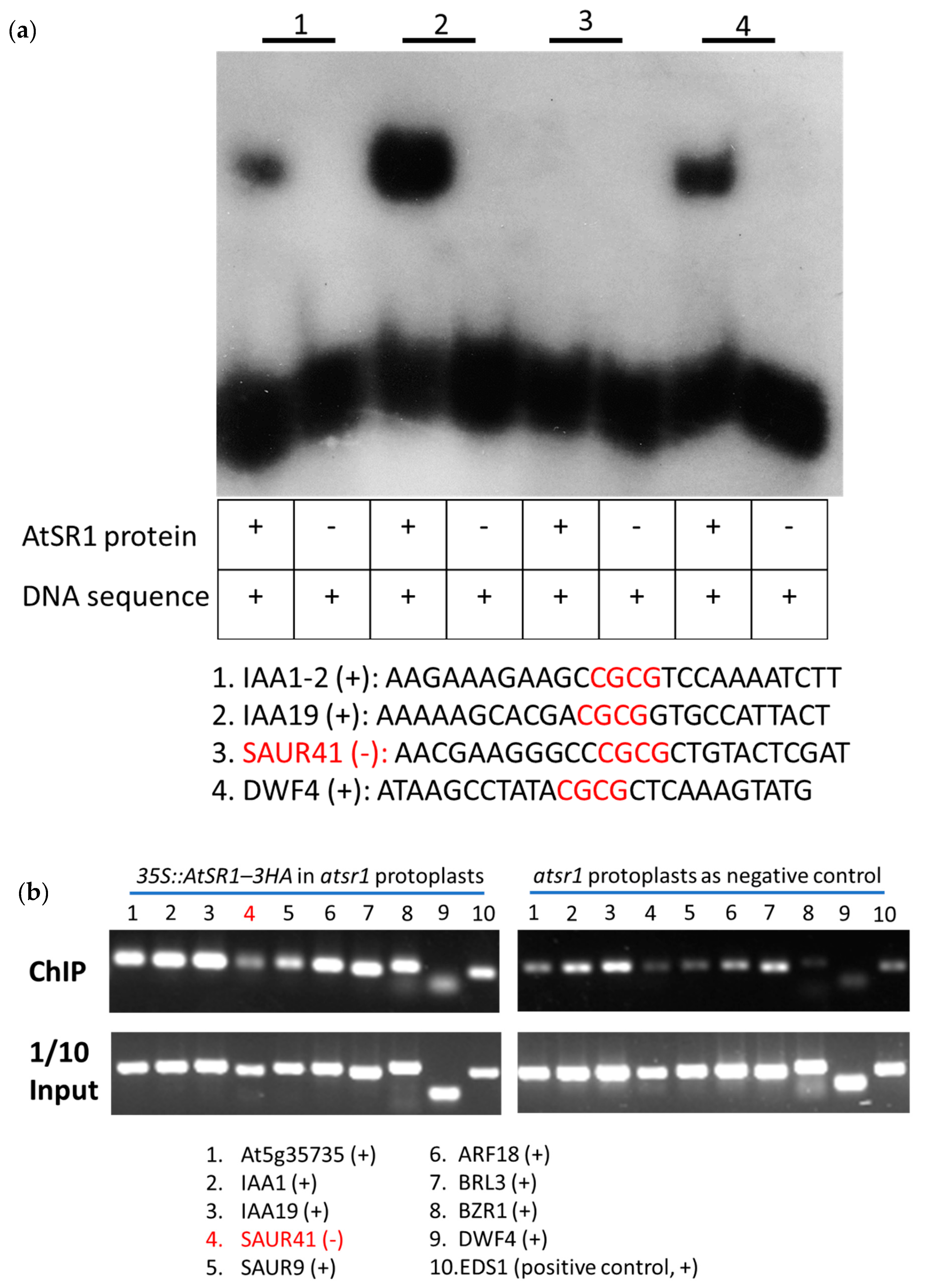
2.7. AtSR1 Could Bind to the CG-Box Motif in the Promotor of Target Genes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth
4.2. Plant Pathogen Infection and Disease Resistance Assays
4.3. Total RNA Extraction and mRNA Isolation
4.4. Differential Expression Genes Analysis of Microarray Data
4.5. Clustering of Microarray Data
4.6. EMSA for Confirmation of the Interaction between AtSR1 Protein and Short DNA for the Promotor of Target Genes
4.7. Mesophyll Protoplast Isolation and Transfection for Transient AtSR1 Expression
4.8. ChIP (Chromatin Immunoprecipitation)-PCR
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Yuan, P.; Jauregui, E.; Du, L.; Tanaka, K.; Poovaiah, B.W. Calcium signatures and signaling events orchestrate plant–microbe interactions. Curr. Opin. Plant Biol. 2017, 38, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Steinhorst, L.; Kudla, J. Signaling in cells and organisms—Calcium holds the line. Curr. Opin. Plant Biol. 2014, 22, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Kracher, B.; Mine, A.; Seyfferth, C.; Blanvillain-Baufumé, S.; Parker, J.E.; Tsuda, K.; Schulze-Lefert, P.; Maekawa, T. A dominant-interfering CAMTA3 mutation compromises primary transcriptional outputs mediated by both cell surface and intracellular immune receptors in arabidopsis thaliana. New Phytol. 2018, 217, 1667–1680. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Ali, G.S.; Simons, K.A.; Hou, J.; Yang, T.; Reddy, A.S.N.; Poovaiah, B.W. Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 2009, 457, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Doherty, C.J.; Van Buskirk, H.A.; Myers, S.J.; Thomashow, M.F. Roles for arabidopsis camta transcription factors in cold-regulated gene expression and freezing tolerance. Plant Cell 2009, 21, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Tanaka, K.; Du, L.; Poovaiah, B.W. Calcium signaling in plant autoimmunity: A guard model for ATSR1/CAMTA3-mediated immune response. Mol. Plant 2018, 11, 637–639. [Google Scholar] [CrossRef] [PubMed]
- Bjornson, M.; Benn, G.; Song, X.; Comai, L.; Franz, A.K.; Dandekar, A.; Drakakaki, G.; Dehesh, K. Distinct roles for MAPK signaling and CAMTA3 in regulating the peak time and amplitude of the plant general stress response. Plant Physiol. 2014, 166, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Zhao, C.; Wu, G.; Wu, Y.; Chen, Y.; Tang, D. SR1, a calmodulin-binding transcription factor, modulates plant defense and ethylene-induced senescence by directly regulating NDR1 and EIN3. Plant Physiol. 2012, 158, 1847–1859. [Google Scholar] [CrossRef] [PubMed]
- Jing, B.; Xu, S.; Xu, M.; Li, Y.; Li, S.; Ding, J.; Zhang, Y. Brush and spray: A high-throughput systemic acquired resistance assay suitable for large-scale genetic screening. Plant Physiol. 2011, 157, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Eichmann, R.; Schäfer, P. Growth versus immunity—A redirection of the cell cycle? Curr. Opin. Plant Biol. 2015, 26, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-Y.; Sae-Seaw, J.; Wang, Z.-Y. Brassinosteroid signalling. Development 2013, 140, 1615–1620. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O. Auxin signaling. Plant Physiol. 2018, 176, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lee, S.; So, J.h.; Dharmasiri, S.; Dharmasiri, N.; Ge, L.; Jensen, C.; Hangarter, R.; Hobbie, L.; Estelle, M. The IAA1 protein is encoded by AXR5 and is a substrate of SCFTIR1. Plant J. 2004, 40, 772–782. [Google Scholar] [CrossRef] [PubMed]
- Gray, W.M.; Kepinski, S.; Rouse, D.; Leyser, O.; Estelle, M. Auxin regulates SCFTIR1-dependent degradation of aux/iaa proteins. Nature 2001, 414, 271. [Google Scholar] [CrossRef] [PubMed]
- Okushima, Y.; Overvoorde, P.J.; Arima, K.; Alonso, J.M.; Chan, A.; Chang, C.; Ecker, J.R.; Hughes, B.; Lui, A.; Nguyen, D.; et al. Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: Unique and overlapping functions of ARF7 and ARF19. Plant Cell 2005, 17, 444–463. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; Friml, J. Auxin: A trigger for change in plant development. Cell 2009, 136, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.; Dilkes, B.P.; Fujioka, S.; Takatsuto, S.; Sakurai, A.; Feldmann, K.A. The DWF4 gene of arabidopsis encodes a cytochrome P450 that mediates multiple 22α-hydroxylation steps in brassinosteroid biosynthesis. Plant Cell 1998, 10, 231–243. [Google Scholar] [PubMed]
- Du, L.; Poovaiah, B.W. Ca2+/calmodulin is critical for brassinosteroid biosynthesis and plant growth. Nature 2005, 437, 741. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Qi, Z.; Berkowitz, G.A. Teaching an old hormone new tricks: Cytosolic ca2+ elevation involvement in plant brassinosteroid signal transduction cascades. Plant Physiol. 2013, 163, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Fàbregas, N.; Li, N.; Boeren, S.; Nash, T.E.; Goshe, M.B.; Clouse, S.D.; de Vries, S.; Caño-Delgado, A.I. The BRASSINOSTEROID INSENSITIVE1–LIKE3 signalosome complex regulates Arabidopsis root development. Plant Cell 2013, 25, 3377–3388. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.H.; Breda, A.; Hardtke, C.S. Brassinosteroid signaling directs formative cell divisions and protophloem differentiation in Arabidopsis root meristems. Development 2017, 144, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Martins, S.; Montiel-Jorda, A.; Cayrel, A.; Huguet, S.; Roux, C.P.-L.; Ljung, K.; Vert, G. Brassinosteroid signaling-dependent root responses to prolonged elevated ambient temperature. Nat. Commun. 2017, 8, 309. [Google Scholar] [CrossRef] [PubMed]
- Caño-Delgado, A.; Yin, Y.; Yu, C.; Vafeados, D.; Mora-García, S.; Cheng, J.-C.; Nam, K.H.; Li, J.; Chory, J. BRL1 and BRL3 are novel brassinosteroid receptors that function in vascular differentiation in Arabidopsis. Development 2004, 131, 5341–5351. [Google Scholar] [CrossRef] [PubMed]
- He, J.-X.; Gendron, J.M.; Sun, Y.; Gampala, S.S.L.; Gendron, N.; Sun, C.Q.; Wang, Z.-Y. Bzr1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses. Science 2005, 307, 1634–1638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Du, L.; Shen, C.; Yang, Y.; Poovaiah, B.W. Regulation of plant immunity through ubiquitin-mediated modulation of ca2+–calmodulin–ATSR1/CAMTA3 signaling. Plant J. 2014, 78, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Day, B.; Dahlbeck, D.; Staskawicz, B.J. Ndr1 interaction with rin4 mediates the differential activation of multiple disease resistance pathways in Arabidopsis. Plant Cell 2006, 18, 2782–2791. [Google Scholar] [CrossRef] [PubMed]
- Wiermer, M.; Feys, B.J.; Parker, J.E. Plant immunity: The eds1 regulatory node. Curr. Opin. Plant Biol. 2005, 8, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Falk, A.; Feys, B.J.; Frost, L.N.; Jones, J.D.G.; Daniels, M.J.; Parker, J.E. EDS1, an essential component of R gene-mediated disease resistance in Arabidopsis has homology to eukaryotic lipases. Proc Natl. Acad. Sci. USA 1999, 96, 3292–3297. [Google Scholar] [CrossRef] [PubMed]
- Wildermuth, M.C.; Dewdney, J.; Wu, G.; Ausubel, F.M. Isochorismate synthase is required to synthesize salicylic acid for plant defence. Nature 2001, 414, 562. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.H.; Mou, Z.; Tada, Y.; Spivey, N.W.; Genschik, P.; Dong, X. Proteasome-mediated turnover of the transcription co-activator NPR1 plays dual roles in regulating plant immunity. Cell 2009, 137, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Weigel, R.R.; Pfitzner, U.M.; Gatz, C. Interaction of nimin1 with NPR1 modulates Pr gene expression in arabidopsis. Plant Cell 2005, 17, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.; Boden, E.; Arias, J. Salicylic acid and NPR1 induce the recruitment of Trans-activating TGA factors to a defense gene promoter in Arabidopsis. Plant Cell 2003, 15, 1846–1858. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Xi, J.; Du, L.; Suttle, J.C.; Poovaiah, B.W. Coupling calcium/calmodulin-mediated signaling and herbivore-induced plant response through calmodulin-binding transcription factor ATSR1/CAMTA3. Plant Mol. Biol. 2012, 79, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Tateda, C.; Zhang, Z.; Shrestha, J.; Jelenska, J.; Chinchilla, D.; Greenberg, J.T. Salicylic acid regulates Arabidopsis microbial pattern receptor kinase levels and signaling. Plant Cell Online 2014, 26, 4171–4187. [Google Scholar] [CrossRef] [PubMed]
- Seyfferth, C.; Tsuda, K. Salicylic acid signal transduction: The initiation of biosynthesis, perception and transcriptional reprogramming. Front. Plant Sci. 2014, 5, 697. [Google Scholar] [CrossRef] [PubMed]
- Cesari, S.; Bernoux, M.; Moncuquet, P.; Kroj, T.; Dodds, P. A novel conserved mechanism for plant nlr protein pairs: The ‘integrated decoy’ hypothesis. Front. Plant Sci. 2014, 5, 606. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Vernaldi, S.; Maekawa, T. Evolution and conservation of plant NLR functions. Front. Immunol. 2013, 4, 297. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Chen, X.; Lin, W.; Chen, S.; Lu, D.; Niu, Y.; Li, L.; Cheng, C.; McCormack, M.; Sheen, J.; et al. Bifurcation of Arabidopsis NLR immune signaling via Ca2+-dependent protein kinases. PLoS Pathog. 2013, 9, e1003127. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.J.; Chini, A.; Basu, D.; Loake, G.J. Targeted activation tagging of the Arabidopsis NBS-LRR gene, ADR1, conveys resistance to virulent pathogens. Mol. Plant Microbe Interact. 2003, 16, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dodds, P.N.; Bernoux, M. What do we know about nod-like receptors in plant immunity? Ann. Rev. Phytopathol. 2017, 55, 205–229. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.T.N.; Chung, E.-H.; Habring-Müller, A.; Demar, M.; Schwab, R.; Dangl, J.L.; Weigel, D.; Chae, E. Activation of a plant NLR complex through heteromeric association with an autoimmune risk variant of another NLR. Curr. Biol. 2017, 27, 1148–1160. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Hake, K.; Wang, W.; Zhao, T.; Romeis, T.; Tang, D. Calcium-dependent protein kinase5 associates with the truncated NLR protein TIR-NBS2 to contribute to exo70B1-mediated immunity. Plant Cell 2017, 29, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Stael, S.; Kmiecik, P.; Willems, P.; Van Der Kelen, K.; Coll, N.S.; Teige, M.; Van Breusegem, F. Plant innate immunity–sunny side up? Trends Plant Sci. 2015, 20, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Kuai, X.; MacLeod, B.J.; Després, C. Integrating data on the arabidopsis NPR1/NPR3/NPR4 salicylic acid receptors; a differentiating argument. Front. Plant Sci. 2015, 6, 235. [Google Scholar] [CrossRef] [PubMed]
- Matschi, S.; Hake, K.; Herde, M.; Hause, B.; Romeis, T. The calcium-dependent protein kinase CPK28 regulates development by inducing growth phase-specific, spatially restricted alterations in jasmonic acid levels independent of defense responses in arabidopsis. Plant Cell 2015, 27, 591–606. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.L.; Kang, J.-H.; Howe, G.A. Jasmonate-triggered plant immunity. J. Chem. Ecol. 2014, 40, 657–675. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, B.C.; Calil, I.P.; Machado, J.P.B.; Santos, A.A.; Fontes, E.P.B. Immune receptors and co-receptors in antiviral innate immunity in plants. Front. Microbiol. 2017, 7, 2139. [Google Scholar] [CrossRef] [PubMed]
- Cao, X. Self-regulation and cross-regulation of pattern-recognition receptor signalling in health and disease. Nat. Rev. Immunol. 2016, 16, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Lolle, S.; Greeff, C.; Petersen, K.; Roux, M.; Jensen, M.K.; Bressendorff, S.; Rodriguez, E.; Sømark, K.; Mundy, J.; Petersen, M. Matching nlr immune receptors to autoimmunity in CAMTA3 mutants using Antimorphic NLR alleles. Cell Host Microbe 2017, 21, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; An, C.; Park, S.; Gilmour, S.J.; Wang, L.; Renna, L.; Brandizzi, F.; Grumet, R.; Thomashow, M. Camta-mediated regulation of salicylic acid immunity pathway genes in arabidopsis exposed to low temperature and pathogen infection. Plant Cell 2017. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Park, S.; Gilmour, S.J.; Thomashow, M.F. Roles of camta transcription factors and salicylic acid in configuring the low-temperature transcriptome and freezing tolerance of arabidopsis. Plant J. 2013, 75, 364–376. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Lin, W.; Gao, X.; Wu, S.; Cheng, C.; Avila, J.; Heese, A.; Devarenne, T.P.; He, P.; Shan, L. Direct ubiquitination of pattern recognition receptor FLS2 attenuates plant innate immunity. Science 2011, 332, 1439–1442. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.V.S.K.; Abdel-Hameed, A.A.E.; Xing, D.; Reddy, A.S.N. Global gene expression analysis using rna-seq uncovered a new role for SR1/CAMTA3 transcription factor in salt stress. Sci. Rep. 2016, 6, 27021. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.S.; Kim, M.C.; Yoo, J.H.; Moon, B.C.; Koo, S.C.; Park, B.O.; Lee, J.H.; Koo, Y.D.; Han, H.J.; Lee, S.Y.; et al. Isolation of a calmodulin-binding transcription factor from rice (Oryza sativa L.). J. Biol. Chem. 2005, 280, 40820–40831. [Google Scholar] [CrossRef] [PubMed]
- Ms medium for arabidopsis. Cold Spring Harbor Protoc. 2010. [CrossRef]
- Lohse, M.; Bolger, A.M.; Nagel, A.; Fernie, A.R.; Lunn, J.E.; Stitt, M.; Usadel, B. Robina: A user-friendly, integrated software solution for rna-seq-based transcriptomics. Nucleic Acids Res. 2012, 40, W622–W627. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.P.; Giorgi, F.M.; Lohse, M.; Kvederaviciute, K.; Klages, S.; Usadel, B.; Meskiene, I.; Reinhardt, R.; Hincha, D.K. Transcriptome sequencing and microarray design for functional genomics in the extremophile arabidopsis relative thellungiella salsuginea (eutrema salsugineum). BMC Genom. 2013, 14, 793. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, F.M.; Del Fabbro, C.; Licausi, F. Comparative study of rna-seq- and microarray-derived coexpression networks in arabidopsis thaliana. Bioinformatics 2013, 29, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Deeken, R.; Ache, P.; Kajahn, I.; Klinkenberg, J.; Bringmann, G.; Hedrich, R. Identification of Arabidopsis thaliana phloem RNAS provides a search criterion for phloem-based transcripts hidden in complex datasets of microarray experiments. Plant J. 2008, 55, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-H.; Tian, X.; Li, Y.-J.; Wu, C.-A.; Zheng, C.-C. Microarray-based analysis of stress-regulated micrornas in arabidopsis thaliana. RNA 2008, 14, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Kleber-Janke, T.; Becker, W.-M. Use of modified BL21(DE3) Escherichia coli cells for high-level expression of recombinant peanut allergens affected by poor codon usage. Protein Expr. Purif. 2000, 19, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Assadi-Porter, F.M.; Patry, S.; Markley, J.L. Efficient and rapid protein expression and purification of small high disulfide containing sweet protein brazzein in E. Coli. Protein Expr. Purif. 2008, 58, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jin, S.; Kim, S.Y.; Kim, W.; Ahn, J.H. A fast, efficient chromatin immunoprecipitation method for studying protein-DNA binding in arabidopsis mesophyll protoplasts. Plant Methods 2017, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Nanjareddy, K.; Arthikala, M.-K.; Blanco, L.; Arellano, E.S.; Lara, M. Protoplast isolation, transient transformation of leaf mesophyll protoplasts and improved agrobacterium-mediated leaf disc infiltration of phaseolus vulgaris: Tools for rapid gene expression analysis. BMC Biotechnol. 2016, 16, 53. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-Y.; Seo, P.J.; Cho, S.-H. Preparation of leaf mesophyll protoplasts for transient gene expression in brachypodium distachyon. J. Plant Biol. 2012, 55, 390–397. [Google Scholar] [CrossRef]
- Yoo, S.-D.; Cho, Y.-H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Adli, M.; Bernstein, B.E. Whole-genome chromatin profiling from limited numbers of cells using nano-chip-seq. Nat. Protoc. 2011, 6, 1656–1668. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ratio | Direction | AGI Code | Functional Description | ChIP/SR1 | EMSA/SR1 |
|---|---|---|---|---|---|
| 1.90 | ↑ | At5g04720 | Encodes a member of the ADR1 family NB-LRR immune receptors | + | + |
| 2.36 | ↑ | At5g45000 | Disease resistance protein (TIR-NBS-LRR class) family | − | − |
| 0.99 | ↑ | At5g66890 | Disease resistance protein (CC-NBS-LRR class) family | + | + |
| 2.24 | ↑ | At5g41750 | Disease resistance protein (TIR-NBS-LRR class) family | + | −,+,+ |
| 1.77 | ↑ | At3g05650 | receptor like protein 32 (RLP32); FUNCTIONS IN: kinase activity | + | + |
| 1.77 | ↑ | At1g33560 | ACTIVATED DISEASE RESISTANCE 1, ADR1. | + | + |
| 1.16 | ↑ | At1g56510 | TIR-NB-LRR protein confers resistance to Albugo candidate | + | − |
| 3.23 | ↑ | At1g02450 | NIMIN1 modulates PR gene expression with other TF(s) | − | −,+,−,− |
| 2.25 | ↑ | At1g19610 | LCR78, LOW-MOLECULAR-WEIGHT CYSTEINE-RICH 78, PDF1.4 | + | + |
| 1.00 | ↑ | AT1G64280 | NPR1 (NONEXPRESSER OF PR GENES 1); Co-transcription factor | + | + |
| Ratio | Direction | AGI Code | Functional Description | ChIP/SR1 | EMSA/SR1 |
|---|---|---|---|---|---|
| 2.04 | ↑ | At5g35735 | auxin-responsive family protein with unknown function | + | +,− |
| 1.57 | ↓ | At4g14560 | auxin-responsive protein/indoleacetic acid-induced protein 1 (IAA1) | + | −,+ |
| 0.44 | ↑ | At3g15540 | auxin-responsive protein/indoleacetic acid-induced protein 19 (IAA19) | + | + |
| 1.40 | ↑ | At1g16510 | SAUR41, small auxin up-regulated protein 41 | − | − |
| 1.27 | ↑ | At4g36110 | SAUR9, small auxin up-regulated protein 9 | + | + |
| 0.34 | ↓ | At3g61830 | ARF18, auxin-responsive factor gene family, AUX/IAA-related | + | +,+ |
| 1.34 | ↑ | At3g13380 | An LRR receptor-like kinase similar to BRI (BRL3) | + | + |
| 1.47 | ↓ | At1g75080 | brassinosteroid signaling positive regulator (BZR1) | + | - |
| 0.69 | ↓ | At3g50660 | steroid 22-alpha-hydroxylase (CYP90B1) (DWF4) | + | + |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, P.; Du, L.; Poovaiah, B.W. Ca2+/Calmodulin-Dependent AtSR1/CAMTA3 Plays Critical Roles in Balancing Plant Growth and Immunity. Int. J. Mol. Sci. 2018, 19, 1764. https://doi.org/10.3390/ijms19061764
Yuan P, Du L, Poovaiah BW. Ca2+/Calmodulin-Dependent AtSR1/CAMTA3 Plays Critical Roles in Balancing Plant Growth and Immunity. International Journal of Molecular Sciences. 2018; 19(6):1764. https://doi.org/10.3390/ijms19061764
Chicago/Turabian StyleYuan, Peiguo, Liqun Du, and B. W. Poovaiah. 2018. "Ca2+/Calmodulin-Dependent AtSR1/CAMTA3 Plays Critical Roles in Balancing Plant Growth and Immunity" International Journal of Molecular Sciences 19, no. 6: 1764. https://doi.org/10.3390/ijms19061764
APA StyleYuan, P., Du, L., & Poovaiah, B. W. (2018). Ca2+/Calmodulin-Dependent AtSR1/CAMTA3 Plays Critical Roles in Balancing Plant Growth and Immunity. International Journal of Molecular Sciences, 19(6), 1764. https://doi.org/10.3390/ijms19061764