Immunological Processes Driving IgE Sensitisation and Disease Development in Males and Females


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Atopy and IgE Sensitisation
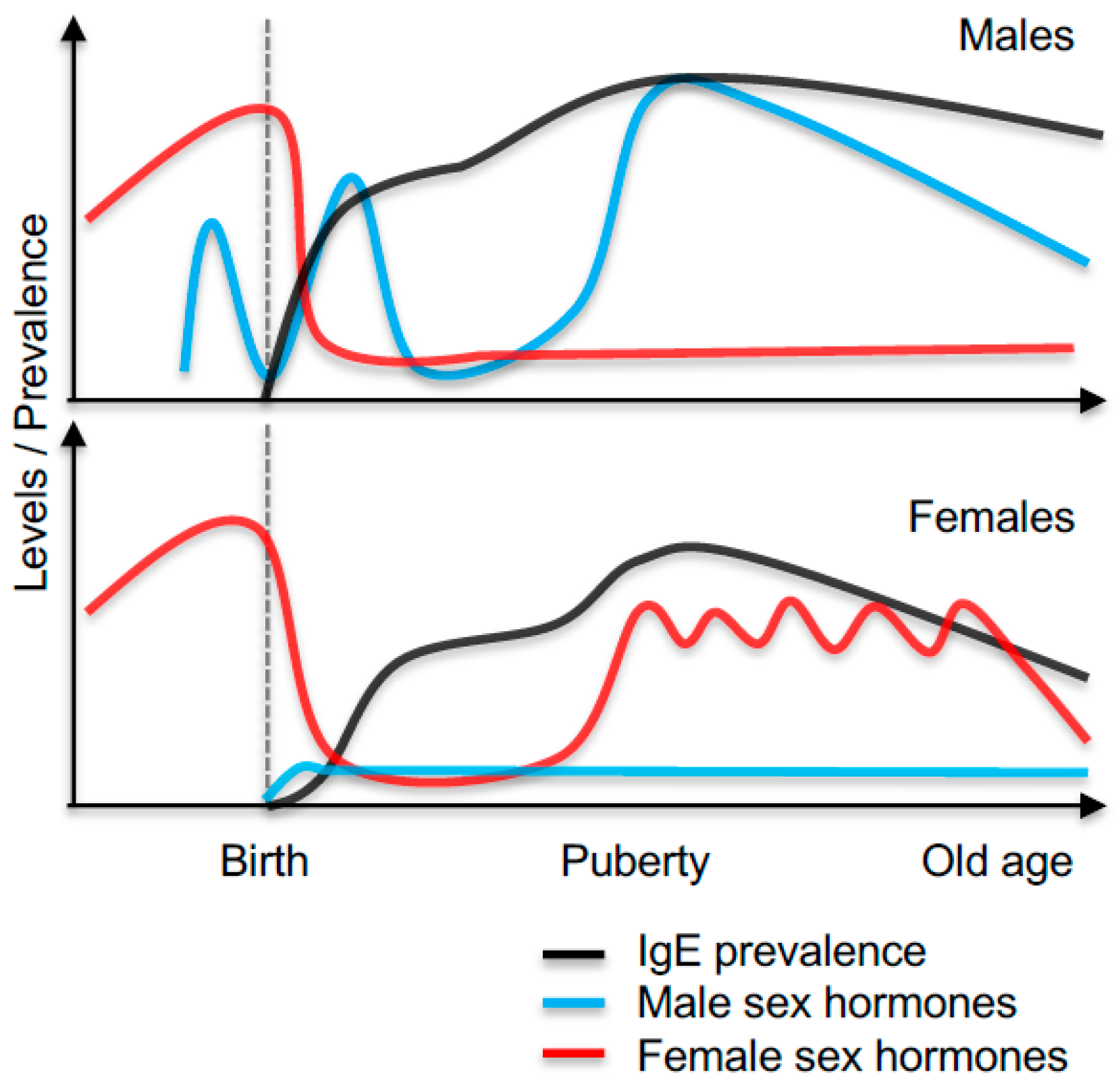
IgE Sensitisation in Male and Females
3. Respiratory Allergic Sensitisation: The Mechanisms in Rodents and Human
3.1. Aeroallergen Uptake and Presentation by Airway Dendritic Cells
3.2. The Influence of Sex Hormones on DC Function
3.3. The Influence of Sex Hormones on Innate Immune Sensing in the Airways
3.4. Sex Hormone Influences on the Induction of CD4+ T Cell-Mediated Immune Responses
3.5. T Cell Development and Activation Are Influenced by Sex Hormones
3.6. Sex Hormone Influences on B Cell Responses and IgE Sensitisation
4. After Sensitisation—Translation into Clinical Allergic Disease
Sex Differences in Asthma Pathology
5. Sex Hormones and Impact on Therapies for Allergic Sensitisation
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Roved, J.; Westerdahl, H.; Hasselquist, D. Sex differences in immune responses: Hormonal effects, antagonistic selection, and evolutionary consequences. Horm. Behav. 2017, 88, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Foo, Y.Z.; Nakagawa, S.; Rhodes, G.; Simmons, L.W. The effects of sex hormones on immune function: A meta-analysis. Biol. Rev. Camb. Philos. Soc. 2017, 92, 551–571. [Google Scholar] [CrossRef] [PubMed]
- Keselman, A.; Heller, N. Estrogen signaling modulates allergic inflammation and contributes to sex differences in asthma. Front. Immunol. 2015, 6, 568. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; O’Malley, B.W. Unfolding the action of progesterone receptors. J. Biol. Chem. 2003, 278, 39261–39264. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.H.; Li, J.; Xu, H.E.; Melcher, K.; Yong, E.L. Androgen receptor: Structure, role in prostate cancer and drug discovery. Acta Pharmacol. Sin. 2015, 36, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Gilliver, S.C. Sex steroids as inflammatory regulators. J. Steroid Biochem. Mol. Biol. 2010, 120, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Pierdominici, M.; Maselli, A.; Colasanti, T.; Giammarioli, A.M.; Delunardo, F.; Vacirca, D.; Sanchez, M.; Giovannetti, A.; Malorni, W.; Ortona, E. Estrogen receptor profiles in human peripheral blood lymphocytes. Immunol. Lett. 2010, 132, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Cordoba-Aguilar, A.; Munguia-Steyer, R. The sicker sex: Understanding male biases in parasitic infection, resource allocation and fitness. PLoS ONE 2013, 8, e76246. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Pinart, M.; Keller, T.; Reich, A.; Frohlich, M.; Cabieses, B.; Hohmann, C.; Postma, D.S.; Bousquet, J.; Anto, J.M.; Keil, T. Sex-related allergic rhinitis prevalence switch from childhood to adulthood: A systematic review and meta-analysis. Int. Arch. Allergy Immunol. 2017, 172, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Barbarot, S.; Auziere, S.; Gadkari, A.; Girolomoni, G.; Puig, L.; Simpson, E.L.; Margolis, D.J.; de Bruin-Weller, M.; Eckert, L. Epidemiology of atopic dermatitis in adults: Results from an international survey. Allergy 2018. [Google Scholar] [CrossRef] [PubMed]
- Patelis, A.; Gunnbjornsdottir, M.; Borres, M.P.; Burney, P.; Gislason, T.; Toren, K.; Forsberg, B.; Alving, K.; Malinovschi, A.; Janson, C. Natural history of perceived food hypersensitivity and IgE sensitisation to food allergens in a cohort of adults. PLoS ONE 2014, 9, e85333. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.H.; Jaramillo, R.; Sicherer, S.H.; Wood, R.A.; Bock, S.A.; Burks, A.W.; Massing, M.; Cohn, R.D.; Zeldin, D.C. National prevalence and risk factors for food allergy and relationship to asthma: Results from the national health and nutrition examination survey 2005–2006. J. Allergy Clin. Immunol. 2010, 126, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Dittmar, D.; Uter, W.; Bauer, A.; Fortina, A.B.; Bircher, A.J.; Czarnecka-Operacz, M.; Dugonik, A.; Elsner, P.; Gallo, R.; Ghaffar, S.A.; et al. European surveillance system on contact allergies (ESSCA): Polysensitization, 2009–2014. Contact Dermat. 2018. [Google Scholar] [CrossRef] [PubMed]
- Afify, S.M.; Pali-Scholl, I. Adverse reactions to food: The female dominance—A secondary publication and update. World Allergy Organ. J. 2017, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Salo, P.M.; Arbes, S.J., Jr.; Jaramillo, R.; Calatroni, A.; Weir, C.H.; Sever, M.L.; Hoppin, J.A.; Rose, K.M.; Liu, A.H.; Gergen, P.J.; et al. Prevalence of allergic sensitization in the United States: Results from the national health and nutrition examination survey (NHANES) 2005–2006. J. Allergy Clin. Immunol. 2014, 134, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Schoos, A.M.; Chawes, B.L.; Rasmussen, M.A.; Bloch, J.; Bonnelykke, K.; Bisgaard, H. Atopic endotype in childhood. J. Allergy Clin. Immunol. 2016, 137, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Salo, P.M.; Calatroni, A.; Gergen, P.J.; Hoppin, J.A.; Sever, M.L.; Jaramillo, R.; Arbes, S.J., Jr.; Zeldin, D.C. Allergy-related outcomes in relation to serum IgE: Results from the national health and nutrition examination survey 2005–2006. J. Allergy Clin. Immunol. 2011, 127, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- Tariq, S.M.; Matthews, S.M.; Hakim, E.A.; Stevens, M.; Arshad, S.H.; Hide, D.W. The prevalence of and risk factors for atopy in early childhood: A whole population birth cohort study. J. Allergy Clin. Immunol. 1998, 101, 587–593. [Google Scholar] [CrossRef]
- Marinho, S.; Simpson, A.; Soderstrom, L.; Woodcock, A.; Ahlstedt, S.; Custovic, A. Quantification of atopy and the probability of rhinitis in preschool children: A population-based birth cohort study. Allergy 2007, 62, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Simpson, A.; Soderstrom, L.; Ahlstedt, S.; Murray, C.S.; Woodcock, A.; Custovic, A. IgE antibody quantification and the probability of wheeze in preschool children. J. Allergy Clin. Immunol. 2005, 116, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Perzanowski, M.S.; Ronmark, E.; Platts-Mills, T.A.; Lundback, B. Effect of cat and dog ownership on sensitization and development of asthma among preteenage children. Am. J. Respir. Crit. Care Med. 2002, 166, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.M.; Hrusch, C.L.; Gozdz, J.; Igartua, C.; Pivniouk, V.; Murray, S.E.; Ledford, J.G.; Marques dos Santos, M.; Anderson, R.L.; Metwali, N.; et al. Innate immunity and asthma risk in Amish and Hutterite farm children. N. Engl. J. Med. 2016, 375, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Braun-Fahrlander, C.; Gassner, M.; Grize, L.; Neu, U.; Sennhauser, F.H.; Varonier, H.S.; Vuille, J.C.; Wuthrich, B. Prevalence of hay fever and allergic sensitization in farmer’s children and their peers living in the same rural community. Scarpol team. Swiss study on childhood allergy and respiratory symptoms with respect to air pollution. Clin. Exp. Allergy 1999, 29, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.; Belgrave, D.; Papastamoulis, P.; Simpson, A.; Rattray, M.; Custovic, A. Evolution of IgE responses to multiple allergen components throughout childhood. J. Allergy Clin. Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Gough, H.; Grabenhenrich, L.; Reich, A.; Eckers, N.; Nitsche, O.; Schramm, D.; Beschorner, J.; Hoffmann, U.; Schuster, A.; Bauer, C.P.; et al. Allergic multimorbidity of asthma, rhinitis and eczema over 20 years in the German birth cohort MAS. Pediatr. Allergy Immunol. 2015, 26, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Bantz, S.K.; Zhu, Z.; Zheng, T. The atopic march: Progression from atopic dermatitis to allergic rhinitis and asthma. J. Clin. Cell. Immunol. 2014, 5. [Google Scholar] [CrossRef]
- Jones, C.A.; Holloway, J.A.; Warner, J.O. Does atopic disease start in foetal life. Allergy 2000, 55, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Bonnelykke, K.; Pipper, C.B.; Bisgaard, H. Sensitization does not develop in utero. J. Allergy Clin. Immunol. 2008, 121, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, E.J.; Yoon, D.; Lee, J.K.; Chang, W.S.; Lim, Y.M.; Park, J.W.; Lee, J.S. Prevalence of self-reported allergic diseases and IgE levels: A 2010 knhanes analysis. Allergy Asthma Immunol. Res. 2017, 9, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Uekert, S.J.; Akan, G.; Evans, M.D.; Li, Z.; Roberg, K.; Tisler, C.; Dasilva, D.; Anderson, E.; Gangnon, R.; Allen, D.B.; et al. Sex-related differences in immune development and the expression of atopy in early childhood. J. Allergy Clin. Immunol. 2006, 118, 1375–1381. [Google Scholar] [CrossRef] [PubMed]
- Sherrill, D.L.; Stein, R.; Halonen, M.; Holberg, C.J.; Wright, A.; Martinez, F.D. Total serum IgE and its association with asthma symptoms and allergic sensitization among children. J. Allergy Clin. Immunol. 1999, 104, 28–36. [Google Scholar] [CrossRef]
- Sears, M.R.; Burrows, B.; Flannery, E.M.; Herbison, G.P.; Hewitt, C.J.; Holdaway, M.D. Relation between airway responsiveness and serum IgE in children with asthma and in apparently normal-children. N. Engl. J. Med. 1991, 325, 1067–1071. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, H.P.; Herskind, A.M.; Nielsen, B.W.; Husby, S. Ige in unselected like-sexed monozygotic and dizygotic twins at birth and at 6 to 9 years of age: High but dissimilar genetic influence on IgE levels. J. Allergy Clin. Immunol. 2001, 107, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Nickel, R.; Illi, S.; Lau, S.; Sommerfeld, C.; Bergmann, R.; Kamin, W.; Forster, J.; Schuster, A.; Niggemann, B.; Wahn, U.; et al. Variability of total serum immunoglobulin E levels from birth to the age of 10 years. A prospective evaluation in a large birth cohort (german multicenter allergy study). Clin. Exp. Allergy 2005, 35, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Barbee, R.A.; Halonen, M.; Kaltenborn, W.; Lebowitz, M.; Burrows, B. A longitudinal-study of serum IgE in a community cohort—Correlations with age, sex, smoking, and atopic status. J. Allergy Clin. Immunol. 1987, 79, 919–927. [Google Scholar] [CrossRef]
- Govaere, E.; Van Gysel, D.; Massa, G.; Verhamme, K.M.; Doli, E.; De Baets, F. The influence of age and gender on sensitization to aero-allergens. Pediatr. Allergy Immunol. 2007, 18, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Hollams, E.M.; Deverell, M.; Serralha, M.; Suriyaarachchi, D.; Parsons, F.; Zhang, G.; de Klerk, N.; Holt, B.J.; Ladyman, C.; Sadowska, A.; et al. Elucidation of asthma phenotypes in atopic teenagers through parallel immunophenotypic and clinical profiling. J. Allergy Clin. Immunol. 2009, 124, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Daniluk, U.; Alifier, M.; Kaczmarski, M.; Stasiak-Barmuta, A.; Lebensztejn, D. Longitudinal observation of children with enhanced total serum IgE. Ann. Allergy Asthma Immunol. 2015, 114, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Shin, Y.H.; Han, M.Y. Determinants of sensitization to allergen in infants and young children. Korean J. Pediatr. 2014, 57, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.M.; Toppari, J.; Haavisto, A.M.; Petersen, J.H.; Simell, T.; Simell, O.; Skakkebaek, N.E. Longitudinal reproductive hormone profiles in infants: Peak of inhibin B levels in infant boys exceeds levels in adult men. J. Clin. Endocrinol. Metab. 1998, 83, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, D.; Luczynska, C.; Chinn, S.; Burney, P. The association of age, gender and smoking with total IgE and specific IgE. Clin. Exp. Allergy 1995, 25, 1083–1091. [Google Scholar] [PubMed]
- Jarvis, D.; Zock, J.P.; Heinrich, J.; Svanes, C.; Verlato, G.; Olivieri, M.; Villani, S.; Ponzio, M.; Leynaert, B.; Sunyer, J.; et al. Cat and dust mite allergen levels, specific IgG and IgG4, and respiratory symptoms in adults. J. Allergy Clin. Immunol. 2007, 119, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Omenaas, E.; Bakke, P.; Elsayed, S.; Hanoa, R.; Gulsvik, A. Total and specific serum IgE levels in adults: Relationship to sex, age and environmental factors. Clin. Exp. Allergy 1994, 24, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.F.S.; Newson, R.B.; Abramson, M.J.; Anto, J.M.; Bono, R.; Corsico, A.G.; de Marco, R.; Demoly, P.; Forsberg, B.; Gislason, T.; et al. Changes in IgE sensitization and total IgE levels over 20 years of follow-up. J. Allergy Clin. Immunol. 2016, 137, 1788–1795. [Google Scholar] [CrossRef] [PubMed]
- Vellutini, M.; Viegi, G.; Parrini, D.; Pedreschi, M.; Baldacci, S.; Modena, P.; Biavati, P.; Simoni, M.; Carrozzi, L.; Giuntini, C. Serum immunoglobulins E are related to menstrual cycle. Eur. J. Epidemiol. 1997, 13, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, G.; Mansueto, P.; Melluso, M.; Morici, G.; Norrito, F.; Esposito Pellitteri, M.; Di Salvo, A.; Colombo, A.; Candore, G.; Caruso, C. Non-specific airway hyperresponsiveness in mono-sensitive sicilian patients with allergic rhinitis. Its relationship to total serum IgE levels and blood eosinophils during and out of the pollen season. Clin. Exp. Allergy 1997, 27, 1052–1059. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Salomon, B.; Klatzmann, D.; Pauwels, R.A. Dendritic cells are required for the development of chronic eosinophilic airway inflammation in response to inhaled antigen in sensitized mice. J. Immunol. 1998, 160, 4090–4097. [Google Scholar] [PubMed]
- Paul, W.E.; Zhu, J. How are t(h)2-type immune responses initiated and amplified? Nat. Rev. Immunol. 2010, 10, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Holt, P.G.; Stumbles, P.A. Regulation of immunologic homeostasis in peripheral tissues by dendritic cells: The respiratory tract as a paradigm. J. Allergy Clin. Immunol. 2000, 105, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Holt, P.G.; Schonhegrad, M.A.; Phillips, M.J.; Mcmenamin, P.G. Ia-positive dendritic cells form a tightly meshed network within the human airway epithelium. Clin. Exp. Allergy 1989, 19, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Dutertre, C.A.; Scott, C.L.; McGovern, N.; Sichien, D.; Chakarov, S.; Van Gassen, S.; Chen, J.; Poidinger, M.; De Prijck, S.; et al. Unsupervised high-dimensional analysis aligns dendritic cells across tissues and species. Immunity 2016, 45, 669–684. [Google Scholar] [CrossRef] [PubMed]
- Serbina, N.V.; Pamer, E.G. Monocyte emigration from bone marrow during bacterial infection requires signals mediated by chemokine receptor CCR2. Nat. Immunol. 2006, 7, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Plantinga, M.; Guilliams, M.; Vanheerswynghels, M.; Deswarte, K.; Branco-Madeira, F.; Toussaint, W.; Vanhoutte, L.; Neyt, K.; Killeen, N.; Malissen, B.; et al. Conventional and monocyte-derived CD11b+ dendritic cells initiate and maintain T helper 2 cell-mediated immunity to house dust mite allergen. Immunity 2013, 38, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Medoff, B.D.; Seung, E.; Hong, S.; Thomas, S.Y.; Sandall, B.P.; Duffield, J.S.; Kuperman, D.A.; Erle, D.J.; Luster, A.D. CD11b+ myeloid cells are the key mediators of Th2 cell homing into the airway in allergic inflammation. J. Immunol. 2008, 182, 623–635. [Google Scholar] [CrossRef]
- Yoshida, T.; Hachimura, S.; Ishimori, M.; Ise, W.; Totsuka, M.; Ametani, A.; Kaminogawa, S. Interleukin 12 and CD86 regulate Th1 and Th2 development induced by a range of antigen doses presented by peyer’s patch and spleen cells. Cytotechnology 2003, 43, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Leffler, J.; Mincham, K.T.; Mok, D.; Blank, F.; Holt, P.G.; Stumbles, P.A.; Strickland, D.H. Functional differences in airway dendritic cells determine susceptibility to IgE-sensitization. Immunol. Cell. Biol. 2018, 96, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Akbari, O.; DeKruyff, R.H.; Umetsu, D.T. Pulmonary dendritic cells producing IL-10 mediate tolerance induced by respiratory exposure to antigen. Nat. Immunol. 2001, 2, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Meiler, F.; Zumkehr, J.; Klunker, S.; Ruckert, B.; Akdis, C.A.; Akdis, M. In vivo switch to IL-10-secreting T regulatory cells in high dose allergen exposure. J. Exp. Med. 2008, 205, 2887–2898. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.C.; Yen, H.R.; Jiang, R.S.; Wang, R.C.; Huang, W.C.; Chen, S.C.; Lin, B.S.; Liang, K.L. Dose-response relationship of specific allergen exposure-induced immunological tolerance: A mouse model. Int. Forum Allergy Rhinol. 2015, 5, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Cook, P.C.; MacDonald, A.S. Dendritic cells in lung immunopathology. Semin. Immunopathol. 2016, 38, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Woodfolk, J.A. High-dose allergen exposure leads to tolerance. Clin. Rev. Allergy Immunol. 2005, 28, 43–58. [Google Scholar] [CrossRef]
- Flutter, B.; Nestle, F.O. What on “IRF” is this gene 4? IRF4 transcription-factor-dependent dendritic cells are required for T helper 2 cell responses in murine skin. Immunity 2013, 39, 625–627. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Nish, S.A.; Jiang, R.; Hou, L.; Licona-Limon, P.; Weinstein, J.S.; Zhao, H.; Medzhitov, R. Control of T helper 2 responses by transcription factor IRF4-dependent dendritic cells. Immunity 2013, 39, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Khare, A.; Krishnamoorthy, N.; Oriss, T.B.; Fei, M.; Ray, P.; Ray, A. Cutting edge: Inhaled antigen upregulates retinaldehyde dehydrogenase in lung CD103+ but not plasmacytoid dendritic cells to induce Foxp3 de novo in CD4+ T cells and promote airway tolerance. J. Immunol. 2013, 191, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Everts, B.; Tussiwand, R.; Dreesen, L.; Fairfax, K.C.; Huang, S.C.; Smith, A.M.; O’Neill, C.M.; Lam, W.Y.; Edelson, B.T.; Urban, J.F., Jr.; et al. Migratory CD103+ dendritic cells suppress helminth-driven type 2 immunity through constitutive expression of IL-12. J. Exp. Med. 2016, 213, 35–51. [Google Scholar] [CrossRef] [PubMed]
- De Heer, H.J.; Hammad, H.; Soullie, T.; Hijdra, D.; Vos, N.; Willart, M.A.; Hoogsteden, H.C.; Lambrecht, B.N. Essential role of lung plasmacytoid dendritic cells in preventing asthmatic reactions to harmless inhaled antigen. J. Exp. Med. 2004, 200, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Oriss, T.B.; Ostroukhova, M.; Seguin-Devaux, C.; Dixon-McCarthy, B.; Stolz, D.B.; Watkins, S.C.; Pillemer, B.; Ray, P.; Ray, A. Dynamics of dendritic cell phenotype and interactions with CD4+ T cells in airway inflammation and tolerance. J. Immunol. 2005, 174, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Desch, A.N.; Randolph, G.J.; Murphy, K.; Gautier, E.L.; Kedl, R.M.; Lahoud, M.H.; Caminschi, I.; Shortman, K.; Henson, P.M.; Jakubzick, C.V. CD103+ pulmonary dendritic cells preferentially acquire and present apoptotic cell-associated antigen. J. Exp. Med. 2011, 208, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P.; Mazzone, S.B.; Rogers, M.J.; Arikkatt, J.J.; Loh, Z.; Pritchard, A.L.; Upham, J.W.; Phipps, S. The plasmacytoid dendritic cell: At the cross-roads in asthma. Eur. Respir. J. 2014, 43, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Laffont, S.; Seillet, C.; Guery, J.C. Estrogen receptor-dependent regulation of dendritic cell development and function. Front. Immunol. 2017, 8, 108. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, Y.; Ran, S.; Segal, S. Sex-associated differences in the regulation of immune responses controlled by the mhc of the mouse. J. Immunol. 1984, 132, 656–661. [Google Scholar] [PubMed]
- Seillet, C.; Rouquie, N.; Foulon, E.; Douin-Echinard, V.; Krust, A.; Chambon, P.; Arnal, J.F.; Guery, J.C.; Laffont, S. Estradiol promotes functional responses in inflammatory and steady-state dendritic cells through differential requirement for activation function-1 of estrogen receptor alpha. J. Immunol. 2013, 190, 5459–5470. [Google Scholar] [CrossRef] [PubMed]
- Anipindi, V.C.; Bagri, P.; Roth, K.; Dizzell, S.E.; Nguyen, P.V.; Shaler, C.R.; Chu, D.K.; Jimenez-Saiz, R.; Liang, H.; Swift, S.; et al. Estradiol enhances CD4+ T-cell anti-viral immunity by priming vaginal DCs to induce Th17 responses via an IL-1-dependent pathway. PLoS Pathog. 2016, 12, e1005589. [Google Scholar]
- Paharkova-Vatchkova, V.; Maldonado, R.; Kovats, S. Estrogen preferentially promotes the differentiation of CD11c+ CD11bintermediate dendritic cells from bone marrow precursors. J. Immunol. 2004, 172, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Douin-Echinard, V.; Laffont, S.; Seillet, C.; Delpy, L.; Krust, A.; Chambon, P.; Gourdy, P.; Arnal, J.F.; Guery, J.C. Estrogen receptor α, but not β, is required for optimal dendritic cell differentiation and CD40-induced cytokine production. J. Immunol. 2008, 180, 3661–3669. [Google Scholar] [CrossRef] [PubMed]
- Melgert, B.N.; Oriss, T.B.; Qi, Z.; Dixon-McCarthy, B.; Geerlings, M.; Hylkema, M.N.; Ray, A. Macrophages: Regulators of sex differences in asthma? Am. J. Respir. Cell Mol. Biol. 2010, 42, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, A.K.; Ryan, E.J.; Giordano, D.; Magaletti, D.M.; Clark, E.A. 17beta-estradiol (E2) modulates cytokine and chemokine expression in human monocyte-derived dendritic cells. Blood 2004, 104, 1404–1410. [Google Scholar] [CrossRef] [PubMed]
- Helft, J.; Bottcher, J.; Chakravarty, P.; Zelenay, S.; Huotari, J.; Schraml, B.U.; Goubau, D.; Reis e Sousa, C. GM-CSF mouse bone marrow cultures comprise a heterogeneous population of CD11c+MHCII+ macrophages and dendritic cells. Immunity 2015, 42, 1197–1211. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; He, H.; Li, C.; Shi, Y.; Wang, Q.; Li, W.; Song, W. Immunosuppressive effect of progesterone on dendritic cells in mice. J. Reprod. Immunol. 2011, 91, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Xiu, F.; Anipindi, V.C.; Nguyen, P.V.; Boudreau, J.; Liang, H.; Wan, Y.; Snider, D.P.; Kaushic, C. High physiological concentrations of progesterone reverse estradiol-mediated changes in differentiation and functions of bone marrow derived dendritic cells. PLoS ONE 2016, 11, e0153304. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.; Kyurkchiev, D.; Altankova, I.; Dimitrov, J.; Binakova, E.; Kyurkchiev, S. CD83+ monocyte-derived dendritic cells are present in human decidua and progesterone induces their differentiation in vitro. Am. J. Reprod. Immunol. 2005, 53, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Corrales, J.J.; Almeida, M.; Miralles, J.M.; Orfao, A. Persistence of androgenic effects on the production of proinflammatory cytokines by circulating antigen-presenting cells after withdrawal of testosterone treatment in aging type 2 diabetic men with partial androgen deficiency. Fertil. Steril. 2009, 92, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Koh, Y.T.; Gray, A.; Higgins, S.A.; Hubby, B.; Kast, W.M. Androgen ablation augments prostate cancer vaccine immunogenicity only when applied after immunization. Prostate 2009, 69, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Seillet, C.; Laffont, S.; Tremollieres, F.; Rouquie, N.; Ribot, C.; Arnal, J.F.; Douin-Echinard, V.; Gourdy, P.; Guery, J.C. The TLR-mediated response of plasmacytoid dendritic cells is positively regulated by estradiol in vivo through cell-intrinsic estrogen receptor alpha signaling. Blood 2012, 119, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Griesbeck, M.; Ziegler, S.; Laffont, S.; Smith, N.; Chauveau, L.; Tomezsko, P.; Sharei, A.; Kourjian, G.; Porichis, F.; Hart, M.; et al. Sex differences in plasmacytoid dendritic cell levels of IRF5 drive higher IFN-alpha production in women. J. Immunol. 2015, 195, 5327–5336. [Google Scholar] [CrossRef] [PubMed]
- Pelka, K.; Latz, E. IRF5, IRF8, and IRF7 in human pDCs—The good, the bad, and the insignificant? Eur. J. Immunol. 2013, 43, 1693–1697. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Zhang, L.; Madera, R.F.; Woda, M.; Libraty, D.H. Plasmacytoid dendritic cell interferon-alpha production to R-848 stimulation is decreased in male infants. BMC Immunol. 2012, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Papazian, D.; Hansen, S.; Wurtzen, P.A. Airway responses towards allergens—From the airway epithelium to T cells. Clin. Exp. Allergy 2015, 45, 1268–1287. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H. Allergens and the airway epithelium response: Gateway to allergic sensitization. J. Allergy Clin. Immunol. 2014, 134, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Kohl, J.; Baelder, R.; Lewkowich, I.P.; Pandey, M.K.; Hawlisch, H.; Wang, L.; Best, J.; Herman, N.S.; Sproles, A.A.; Zwirner, J.; et al. A regulatory role for the C5a anaphylatoxin in type 2 immunity in asthma. J. Clin. Investig. 2006, 116, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Drouin, S.M.; Corry, D.B.; Kildsgaard, J.; Wetsel, R.A. Cutting edge: The absence of C3 demonstrates a role for complement in Th2 effector functions in a murine model of pulmonary allergy. J. Immunol. 2001, 167, 4141–4145. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Chieppa, M.; Perros, F.; Willart, M.A.; Germain, R.N.; Lambrecht, B.N. House dust mite allergen induces asthma via toll-like receptor 4 triggering of airway structural cells. Nat. Med. 2009, 15, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Sheih, A.; Parks, W.C.; Ziegler, S.F. GM-CSF produced by the airway epithelium is required for sensitization to cockroach allergen. Mucosal Immunol. 2017, 10, 705–715. [Google Scholar] [CrossRef] [PubMed]
- McAlees, J.W.; Whitehead, G.S.; Harley, I.T.; Cappelletti, M.; Rewerts, C.L.; Holdcroft, A.M.; Divanovic, S.; Wills-Karp, M.; Finkelman, F.D.; Karp, C.L.; et al. Distinct TLR4-expressing cell compartments control neutrophilic and eosinophilic airway inflammation. Mucosal Immunol. 2015, 8, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Iijima, K.; Kobayashi, T.; Hara, K.; Kephart, G.M.; Ziegler, S.F.; McKenzie, A.N.; Kita, H. IL-33 and thymic stromal lymphopoietin mediate immune pathology in response to chronic airborne allergen exposure. J. Immunol. 2014, 193, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; O’Connor, B.; Ratoff, J.; Meng, Q.; Mallett, K.; Cousins, D.; Robinson, D.; Zhang, G.; Zhao, J.; Lee, T.H.; et al. Thymic stromal lymphopoietin expression is increased in asthmatic airways and correlates with expression of Th2-attracting chemokines and disease severity. J. Immunol. 2005, 174, 8183–8190. [Google Scholar] [CrossRef] [PubMed]
- Mjosberg, J.M.; Trifari, S.; Crellin, N.K.; Peters, C.P.; van Drunen, C.M.; Piet, B.; Fokkens, W.J.; Cupedo, T.; Spits, H. Human IL-25- and IL-33-responsive type 2 innate lymphoid cells are defined by expression of CRTH2 and CD161. Nat. Immunol. 2011, 12, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Licona-Limon, P.; Kim, L.K.; Palm, N.W.; Flavell, R.A. Th2, allergy and group 2 innate lymphoid cells. Nat. Immunol. 2013, 14, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Halim, T.Y.; Steer, C.A.; Matha, L.; Gold, M.J.; Martinez-Gonzalez, I.; McNagny, K.M.; McKenzie, A.N.; Takei, F. Group 2 innate lymphoid cells are critical for the initiation of adaptive T helper 2 cell-mediated allergic lung inflammation. Immunity 2014, 40, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Haapakoski, R.; Karisola, P.; Fyhrquist, N.; Savinko, T.; Lehtimaki, S.; Wolff, H.; Lauerma, A.; Alenius, H. Toll-like receptor activation during cutaneous allergen sensitization blocks development of asthma through IFN-gamma-dependent mechanisms. J. Investig. Dermatol. 2013, 133, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Ege, M.J.; Mayer, M.; Normand, A.C.; Genuneit, J.; Cookson, W.O.; Braun-Fahrlander, C.; Heederik, D.; Piarroux, R.; von Mutius, E.; Group, G.T.S. Exposure to environmental microorganisms and childhood asthma. N. Engl. J. Med. 2011, 364, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Schroder, P.C.; Illi, S.; Casaca, V.I.; Lluis, A.; Bock, A.; Roduit, C.; Depner, M.; Frei, R.; Genuneit, J.; Pfefferle, P.I.; et al. A switch in regulatory T cells through farm exposure during immune maturation in childhood. Allergy 2017, 72, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Baumann, C.; Bonilla, W.V.; Frohlich, A.; Helmstetter, C.; Peine, M.; Hegazy, A.N.; Pinschewer, D.D.; Lohning, M. T-bet- and stat4-dependent IL-33 receptor expression directly promotes antiviral Th1 cell responses. Proc. Natl. Acad. Sci. USA 2015, 112, 4056–4061. [Google Scholar] [CrossRef] [PubMed]
- Komai-Koma, M.; Wang, E.; Kurowska-Stolarska, M.; Li, D.; McSharry, C.; Xu, D. Interleukin-33 promoting th1 lymphocyte differentiation dependents on IL-12. Immunobiology 2016, 221, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Souyris, M.; Cenac, C.; Azar, P.; Daviaud, D.; Canivet, A.; Grunenwald, S.; Pienkowski, C.; Chaumeil, J.; Mejia, J.E.; Guery, J.C. TLR7 escapes × chromosome inactivation in immune cells. Sci. Immunol. 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Berghofer, B.; Frommer, T.; Haley, G.; Fink, L.; Bein, G.; Hackstein, H. TLR7 ligands induce higher IFN-production in females. J. Immunol. 2006, 177, 2088–2096. [Google Scholar] [CrossRef] [PubMed]
- Rettew, J.A.; Huet-Hudson, Y.M.; Marriott, I. Testosterone reduces macrophage expression in the mouse of toll-like receptor 4, a trigger for inflammation and innate immunity. Biol. Reprod. 2008, 78, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Marriott, I.; Bost, K.L.; Huet-Hudson, Y.M. Sexual dimorphism in expression of receptors for bacterial lipopolysaccharides in murine macrophages: A possible mechanism for gender-based differences in endotoxic shock susceptibility. J. Reprod. Immunol. 2006, 71, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Rettew, J.A.; Huet, Y.M.; Marriott, I. Estrogens augment cell surface TLR4 expression on murine macrophages and regulate sepsis susceptibility in vivo. Endocrinology 2009, 150, 3877–3884. [Google Scholar] [CrossRef] [PubMed]
- Alexanderson, C.; Stener-Victorin, E.; Kullberg, J.; Nilsson, S.; Levin, M.; Cajander, S.; Lonn, L.; Lonn, M.; Holmang, A. A single early postnatal estradiol injection affects morphology and gene expression of the ovary and parametrial adipose tissue in adult female rats. J. Steroid Biochem. Mol. Biol. 2010, 122, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Farkas, I.; Varju, P.; Szabo, E.; Hrabovszky, E.; Okada, N.; Okada, H.; Liposits, Z. Estrogen enhances expression of the complement C5a receptor and the C5a-agonist evoked calcium influx in hormone secreting neurons of the hypothalamus. Neurochem. Int. 2008, 52, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, R.; Smith, J.; Lapointe, M.; Marceau, P.; Tchernof, A.; Cianflone, K. Complement receptors C5ar and C5l2 are associated with metabolic profile, sex hormones, and liver enzymes in obese women pre- and postbariatric surgery. J. Obes. 2014, 2014, 1–12. [Google Scholar] [CrossRef]
- Ouyang, W.; Kolls, J.K.; Zheng, Y. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity 2008, 28, 454–467. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.H. Th9 cells: Differentiation and disease. Immunol. Rev. 2013, 252, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Strickland, D.H.; Stumbles, P.A.; Zosky, G.R.; Subrata, L.S.; Thomas, J.A.; Turner, D.J.; Sly, P.D.; Holt, P.G. Reversal of airway hyperresponsiveness by induction of airway mucosal CD4+CD25+ regulatory T cells. J. Exp. Med. 2006, 203, 2649–2660. [Google Scholar] [CrossRef] [PubMed]
- Martin-Orozco, E.; Norte-Munoz, M.; Martinez-Garcia, J. Regulatory T cells in allergy and asthma. Front. Pediatr. 2017, 5, 117. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Tato, A.; Randall, T.D.; Lund, F.E.; Spolski, R.; Leonard, W.J.; Leon, B. T follicular helper cell plasticity shapes pathogenic T helper 2 cell-mediated immunity to inhaled house dust mite. Immunity 2016, 44, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Iijima, K.; Dent, A.L.; Kita, H. Follicular helper T cells mediate IgE antibody response to airborne allergens. J. Allergy Clin. Immunol. 2017, 139, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S. T follicular helper cell differentiation, function, and roles in disease. Immunity 2014, 41, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Coquet, J.M.; Schuijs, M.J.; Smyth, M.J.; Deswarte, K.; Beyaert, R.; Braun, H.; Boon, L.; Karlsson Hedestam, G.B.; Nutt, S.L.; Hammad, H.; et al. Interleukin-21-producing CD4+ T cells promote type 2 immunity to house dust mites. Immunity 2015, 43, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.W.; Yap, H.K.; Chew, F.T.; Quah, T.C.; Prabhakaran, K.; Chan, G.S.; Wong, S.C.; Seah, C.C. Age- and sex-related changes in lymphocyte subpopulations of healthy asian subjects: From birth to adulthood. Cytometry 1996, 26, 8–15. [Google Scholar] [CrossRef]
- Amadori, A.; Zamarchi, R.; De Silvestro, G.; Forza, G.; Cavatton, G.; Danieli, G.A.; Clementi, M.; Chieco-Bianchi, L. Genetic control of the CD4/CD8 T-cell ratio in humans. Nat. Med. 1995, 1, 1279–1283. [Google Scholar] [CrossRef] [PubMed]
- Bouman, A.; Heineman, M.J.; Faas, M.M. Sex hormones and the immune response in humans. Hum. Reprod. Update 2005, 11, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Olsen, N.J.; Kovacs, W.J. Evidence that androgens modulate human thymic T cell output. J. Investig. Med. 2011, 59, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Phiel, K.L.; Henderson, R.A.; Adelman, S.J.; Elloso, M.M. Differential estrogen receptor gene expression in human peripheral blood mononuclear cell populations. Immunol. Lett. 2005, 97, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, R.; Rider, V.; Evans, M.J.; Abdou, N.I. In vitro-activated human lupus T cells express normal estrogen receptor proteins which bind to the estrogen response element. Lupus 2001, 10, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Moulton, V.R.; Holcomb, D.R.; Zajdel, M.C.; Tsokos, G.C. Estrogen upregulates cyclic AMP response element modulator alpha expression and downregulates interleukin-2 production by human T lymphocytes. Mol. Med. 2012, 18, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Chinen, T.; Kannan, A.K.; Levine, A.G.; Fan, X.; Klein, U.; Zheng, Y.; Gasteiger, G.; Feng, Y.; Fontenot, J.D.; Rudensky, A.Y. An essential role for the IL-2 receptor in treg cell function. Nat. Immunol. 2016, 17, 1322–1333. [Google Scholar] [CrossRef] [PubMed]
- Trigunaite, A.; Dimo, J.; Jorgensen, T.N. Suppressive effects of androgens on the immune system. Cell. Immunol. 2015, 294, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Karpuzoglu-Sahin, E.; Zhi-Jun, Y.; Lengi, A.; Sriranganathan, N.; Ansar Ahmed, S. Effects of long-term estrogen treatment on IFN-gamma, IL-2 and IL-4 gene expression and protein synthesis in spleen and thymus of normal C57BL/6 mice. Cytokine 2001, 14, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, H.P.; Krishnan, H.C.; Singh, R.V.; Hima, L.; Thyagarajan, S. Estrogen modulates in vitro T cell responses in a concentration- and receptor-dependent manner: Effects on intracellular molecular targets and antioxidant enzymes. Mol. Immunol. 2013, 56, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Giron-Gonzalez, J.A.; Moral, F.J.; Elvira, J.; Garcia-Gil, D.; Guerrero, F.; Gavilan, I.; Escobar, L. Consistent production of a higher Th1:Th2 cytokine ratio by stimulated T cells in men compared with women. Eur. J. Endocrinol. 2000, 143, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Pernis, A.B. Estrogen and CD4+ T cells. Curr. Opin. Rheumatol. 2007, 19, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Kissick, H.T.; Sanda, M.G.; Dunn, L.K.; Pellegrini, K.L.; On, S.T.; Noel, J.K.; Arredouani, M.S. Androgens alter T-cell immunity by inhibiting T-helper 1 differentiation. Proc. Natl. Acad. Sci. USA 2014, 111, 9887–9892. [Google Scholar] [CrossRef] [PubMed]
- Hepworth, M.R.; Hardman, M.J.; Grencis, R.K. The role of sex hormones in the development of Th2 immunity in a gender-biased model of trichuris muris infection. Eur. J. Immunol. 2010, 40, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, C.; Kasahara, T.M.; Castro, J.R.; Sacramento, P.M.; Hygino, J.; Centuriao, N.; Cassano, T.; Lopes, L.M.F.; Leite, S.; Silva, V.G.; et al. Pregnancy favors the expansion of circulating functional follicular helper T cells. J. Reprod. Immunol. 2017, 121, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Shin, E.K.; Lee, S.Y.; Her, Y.M.; Park, M.K.; Kwok, S.K.; Ju, J.H.; Park, K.S.; Kim, H.Y.; Cho, M.L.; et al. Oestrogen up-regulates interleukin-21 production by CD4+ T lymphocytes in patients with systemic lupus erythematosus. Immunology 2014, 142, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Park, H.S.; Lee, J.U.; Bothwell, A.L.; Choi, J.M. Gender-specific differences in ppargamma regulation of follicular helper T cell responses with estrogen. Sci. Rep. 2016, 6, 28495. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, D.H.; Choi, J.Y.; Kim, W.J.; Kim, J.Y.; Senejani, A.G.; Hwang, S.S.; Kim, L.K.; Tobiasova, Z.; Lee, G.R.; et al. Ppargamma negatively regulates T cell activation to prevent follicular helper T cells and germinal center formation. PLoS ONE 2014, 9, e99127. [Google Scholar]
- Aluvihare, V.R.; Kallikourdis, M.; Betz, A.G. Regulatory T cells mediate maternal tolerance to the fetus. Nat. Immunol. 2004, 5, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Walecki, M.; Eisel, F.; Klug, J.; Baal, N.; Paradowska-Dogan, A.; Wahle, E.; Hackstein, H.; Meinhardt, A.; Fijak, M. Androgen receptor modulates Foxp3 expression in CD4+CD25+Foxp3+ regulatory T-cells. Mol. Biol. Cell 2015, 26, 2845–2857. [Google Scholar] [CrossRef] [PubMed]
- Arruvito, L.; Sanz, M.; Banham, A.H.; Fainboim, L. Expansion of CD4+CD25+and Foxp3+ regulatory T cells during the follicular phase of the menstrual cycle: Implications for human reproduction. J. Immunol. 2007, 178, 2572–2578. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.P.; Peron, J.P.; Santos Franco, A.L.; Golega, B.A.; Vieira, R.P.; Ibanez, O.C.; Ribeiro, O.G.; Cabrera, W.H.; De Franco, M.; Oliveira-Filho, R.M.; et al. Ovariectomized OVA-sensitized mice display increased frequency of CD4+Foxp3+ T regulatory cells in the periphery. PLoS ONE 2013, 8, e65674. [Google Scholar] [CrossRef] [PubMed]
- Polanczyk, M.J.; Carson, B.D.; Subramanian, S.; Afentoulis, M.; Vandenbark, A.A.; Ziegler, S.F.; Offner, H. Cutting edge: Estrogen drives expansion of the CD4+CD25+ regulatory T cell compartment. J. Immunol. 2004, 173, 2227–2230. [Google Scholar] [CrossRef] [PubMed]
- Brunkow, M.E.; Jeffery, E.W.; Hjerrild, K.A.; Paeper, B.; Clark, L.B.; Yasayko, S.A.; Wilkinson, J.E.; Galas, D.; Ziegler, S.F.; Ramsdell, F. Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse. Nat. Genet. 2001, 27, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Erazo, A.; Kutchukhidze, N.; Leung, M.; Christ, A.P.; Urban, J.F., Jr.; Curotto de Lafaille, M.A.; Lafaille, J.J. Unique maturation program of the IgE response in vivo. Immunity 2007, 26, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Sullivan, B.M.; Allen, C.D. Fluorescent in vivo detection reveals that IgE+ B cells are restrained by an intrinsic cell fate predisposition. Immunity 2012, 36, 857–872. [Google Scholar] [CrossRef] [PubMed]
- Luger, E.O.; Fokuhl, V.; Wegmann, M.; Abram, M.; Tillack, K.; Achatz, G.; Manz, R.A.; Worm, M.; Radbruch, A.; Renz, H. Induction of long-lived allergen-specific plasma cells by mucosal allergen challenge. J. Allergy Clin. Immunol. 2009, 124, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Hallstrand, T.S.; Sprenger, J.D.; Agosti, J.M.; Longton, G.M.; Witherspoon, R.P.; Henderson, W.R., Jr. Long-term acquisition of allergen-specific IgE and asthma following allogeneic bone marrow transplantation from allergic donors. Blood 2004, 104, 3086–3090. [Google Scholar] [CrossRef] [PubMed]
- Vu Van, D.; Beier, K.C.; Pietzke, L.J.; Al Baz, M.S.; Feist, R.K.; Gurka, S.; Hamelmann, E.; Kroczek, R.A.; Hutloff, A. Local T/B cooperation in inflamed tissues is supported by T follicular helper-like cells. Nat. Commun. 2016, 7, 10875. [Google Scholar] [CrossRef] [PubMed]
- Laffleur, B.; Debeaupuis, O.; Dalloul, Z.; Cogne, M. B cell intrinsic mechanisms constraining IgE memory. Front. Immunol. 2017, 8, 1277. [Google Scholar] [CrossRef] [PubMed]
- Harada, M.; Magara-Koyanagi, K.; Watarai, H.; Nagata, Y.; Ishii, Y.; Kojo, S.; Horiguchi, S.; Okamoto, Y.; Nakayama, T.; Suzuki, N.; et al. IL-21-induced bepsilon cell apoptosis mediated by natural killer T cells suppresses IgE responses. J. Exp. Med. 2006, 203, 2929–2937. [Google Scholar] [CrossRef] [PubMed]
- Linhart, B.; Bigenzahn, S.; Hartl, A.; Lupinek, C.; Thalhamer, J.; Valenta, R.; Wekerle, T. Costimulation blockade inhibits allergic sensitization but does not affect established allergy in a murine model of grass pollen allergy. J. Immunol. 2007, 178, 3924–3931. [Google Scholar] [CrossRef] [PubMed]
- Marth, K.; Wollmann, E.; Gallerano, D.; Ndlovu, P.; Makupe, I.; Valenta, R.; Sibanda, E. Persistence of IgE-associated allergy and allergen-specific IgE despite CD4+ T cell loss in aids. PLoS ONE 2014, 9, e97893. [Google Scholar] [CrossRef] [PubMed]
- Butterworth, M.; McClellan, B.; Allansmith, M. Influence of sex in immunoglobulin levels. Nature 1967, 214, 1224–1225. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Denison, M.S.; Tachibana, H.; Yamada, K. Effects of estrogenic compounds on immunoglobulin production by mouse splenocytes. Biol. Pharm. Bull. 2002, 25, 1263–1267. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.; Chai, P.S.; Chong, M.Y.; Tohit, E.R.; Ramasamy, R.; Pei, C.P.; Vidyadaran, S. Gender effect on in vitro lymphocyte subset levels of healthy individuals. Cell. Immunol. 2012, 272, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Gamboa, R.; Joosten, I.; Urbano, P.C.M.; van der Molen, R.G.; van Rijssen, E.; van Cranenbroek, B.; Oosting, M.; Smeekens, S.; Jaeger, M.; Zorro, M.; et al. Differential effects of environmental and genetic factors on T and B cell immune traits. Cell Rep. 2016, 17, 2474–2487. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Dong, G.; Zhao, G.; Liu, F.; Yao, G.; Zhu, Y.; Hou, Y. Gender differences of B cell signature in healthy subjects underlie disparities in incidence and course of SLE related to estrogen. J. Immunol. Res. 2014, 2014, 814598. [Google Scholar] [CrossRef] [PubMed]
- Bynoe, M.S.; Grimaldi, C.M.; Diamond, B. Estrogen up-regulates Bcl-2 and blocks tolerance induction of naive B cells. Proc. Natl. Acad. Sci. USA 2000, 97, 2703–2708. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, C.M.; Michael, D.J.; Diamond, B. Cutting edge: Expansion and activation of a population of autoreactive marginal zone B cells in a model of estrogen-induced lupus. J. Immunol. 2001, 167, 1886–1890. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, C.M.; Cleary, J.; Dagtas, A.S.; Moussai, D.; Diamond, B. Estrogen alters thresholds for B cell apoptosis and activation. J. Clin. Investig. 2002, 109, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Peeva, E.; Venkatesh, J.; Diamond, B. Tamoxifen blocks estrogen-induced B cell maturation but not survival. J. Immunol. 2005, 175, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Yates, M.; Vandenbark, A.A.; Offner, H. Oestrogen-mediated protection of experimental autoimmune encephalomyelitis in the absence of Foxp3+ regulatory T cells implicates compensatory pathways including regulatory B cells. Immunology 2011, 132, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Canellada, A.; Blois, S.; Gentile, T.; Margni Idehu, R.A. In vitro modulation of protective antibody responses by estrogen, progesterone and interleukin-6. Am. J. Reprod. Immunol. 2002, 48, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Yates, M.A.; Li, Y.; Chlebeck, P.; Proctor, T.; Vandenbark, A.A.; Offner, H. Progesterone treatment reduces disease severity and increases IL-10 in experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2010, 220, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Kim, A.R.; Kim, D.K.; Kim, H.W.; Park, Y.H.; Jang, G.H.; Kim, B.; Park, Y.M.; You, J.S.; Kim, H.S.; et al. Interleukin-10-producing CD5+ B cells inhibit mast cells during immunoglobulin E-mediated allergic responses. Sci. Signal. 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Kanda, N.; Tsuchida, T.; Tamaki, K. Testosterone inhibits immunoglobulin production by human peripheral blood mononuclear cells. Clin. Exp. Immunol. 1996, 106, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Olsen, N.J.; Gu, X.; Kovacs, W.J. Bone marrow stromal cells mediate androgenic suppression of B lymphocyte development. J. Clin. Investig. 2001, 108, 1697–1704. [Google Scholar] [CrossRef] [PubMed]
- Viselli, S.M.; Reese, K.R.; Fan, J.; Kovacs, W.J.; Olsen, N.J. Androgens alter B cell development in normal male mice. Cell. Immunol. 1997, 182, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Lundell, A.C.; Nordstrom, I.; Andersson, K.; Strombeck, A.; Ohlsson, C.; Tivesten, A.; Rudin, A. Dihydrotestosterone levels at birth associate positively with higher proportions of circulating immature/naive CD5+ B cells in boys. Sci. Rep. 2017, 7, 15503. [Google Scholar] [CrossRef] [PubMed]
- Strombeck, A.; Nordstrom, I.; Andersson, K.; Andersson, H.; Johansen, S.; Maglio, C.; Rabe, H.; Adlerberth, I.; Wold, A.E.; Hesselmar, B.; et al. Allergic disease in 8-year-old children is preceded by delayed B cell maturation. Clin. Exp. Allergy 2017, 47, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Von Linstow, M.L.; Porsbjerg, C.; Ulrik, C.S.; Nepper-Christensen, S.; Backer, V. Prevalence and predictors of atopy among young danish adults. Clin. Exp. Allergy 2002, 32, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Strachan, D.P.; Harkins, L.S.; Johnston, I.D.; Anderson, H.R. Childhood antecedents of allergic sensitization in young british adults. J. Allergy Clin. Immunol. 1997, 99, 6–12. [Google Scholar] [CrossRef]
- Mohammad, H.R.; Belgrave, D.; Kopec Harding, K.; Murray, C.S.; Simpson, A.; Custovic, A. Age, sex and the association between skin test responses and IgE titres with asthma. Pediatr. Allergy Immunol. 2016, 27, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Couto, T.A.; Falsarella, N.; Brandão de Mattos, C.C.; Mattos, L.C. Total IgE plasma levels vary according to gender and age in brazilian patients with allergic rhinitis. Clinics 2014, 69, 740–744. [Google Scholar] [CrossRef]
- Frohlich, M.; Pinart, M.; Keller, T.; Reich, A.; Cabieses, B.; Hohmann, C.; Postma, D.S.; Bousquet, J.; Anto, J.M.; Keil, T.; et al. Is there a sex-shift in prevalence of allergic rhinitis and comorbid asthma from childhood to adulthood? A meta-analysis. Clin. Transl. Allergy 2017, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Osman, M.; Hansell, A.L.; Simpson, C.R.; Hollowell, J.; Helms, P.J. Gender-specific presentations for asthma, allergic rhinitis and eczema in primary care. Prim. Care Respir. J. 2007, 16, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Nathan, R.A.; Meltzer, E.O.; Selner, J.C.; Storms, W. Prevalence of allergic rhinitis in the United States. J. Allergy Clin. Immunol. 1997, 99, S808–S814. [Google Scholar] [CrossRef]
- Keller, T.; Hohmann, C.; Standl, M.; Wijga, A.H.; Gehring, U.; Melen, E.; Almqvist, C.; Lau, S.; Eller, E.; Wahn, U.; et al. The sex-shift in single disease and multimorbid asthma and rhinitis during puberty—A study by MeDALL. Allergy 2018, 73, 602–614. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martinez, C.E.; Sossa-Briceno, M.P.; Castro-Rodriguez, J.A. Factors predicting persistence of early wheezing through childhood and adolescence: A systematic review of the literature. J. Asthma Allergy 2017, 10, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.; Marks, G.B.; Toelle, B.G.; Belousova, E.; Peat, J.K.; Berry, G.; Woolcock, A.J. Risk factors for onset and remission of atopy, wheeze, and airway hyperresponsiveness. Thorax 2002, 57, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Fuseini, H.; Newcomb, D.C. Mechanisms driving gender differences in asthma. Curr. Allergy Asthma Rep. 2017, 17, 19. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.L.; Belanger, K.; Holford, T.R.; Bracken, M.B. Effect of fetal sex on airway lability in pregnant women with asthma. Am. J. Epidemiol. 2006, 163, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Massaro, D.; Massaro, G.D. Estrogen receptor regulation of pulmonary alveolar dimensions: Alveolar sexual dimorphism in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L866–L870. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.D.; Billimek, J.; Bar-Yoseph, R.; Radom-Aizik, S.; Cooper, D.M.; Anton-Culver, H. Sex differences in the relationship between fitness and obesity on risk for asthma in adolescents. J. Pediatr. 2016, 176, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Tam, A.; Wadsworth, S.; Dorscheid, D.; Man, S.F.; Sin, D.D. Estradiol increases mucus synthesis in bronchial epithelial cells. PLoS ONE 2014, 9, e100633. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Ray, J.M.; Pan, J.H.; Brody, S.L. Sex hormone-dependent regulation of cilia beat frequency in airway epithelium. Am. J. Respir. Cell Mol. Biol. 2012, 46, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Riffo-Vasquez, Y.; Ligeiro de Oliveira, A.P.; Page, C.P.; Spina, D.; Tavares-de-Lima, W. Role of sex hormones in allergic inflammation in mice. Clin. Exp. Allergy 2007, 37, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Adachi, Y.; Hasegawa, K.; Morimoto, M. Less sensitivity for late airway inflammation in males than females in BALB/c mice. Scand. J. Immunol. 2003, 57, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Tanabe, M.; Ito, W.; Ueki, S.; Konnno, Y.; Chihara, M.; Itoga, M.; Kobayashi, Y.; Moritoki, Y.; Kayaba, H.; et al. Gender difference in allergic airway remodelling and immunoglobulin production in mouse model of asthma. Respirology 2013, 18, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Blacquiere, M.J.; Hylkema, M.N.; Postma, D.S.; Geerlings, M.; Timens, W.; Melgert, B.N. Airway inflammation and remodeling in two mouse models of asthma: Comparison of males and females. Int. Arch. Allergy Immunol. 2010, 153, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Ligeiro de Oliveira, A.P.; Oliveira-Filho, R.M.; da Silva, Z.L.; Borelli, P.; Tavares de Lima, W. Regulation of allergic lung inflammation in rats: Interaction between estradiol and corticosterone. Neuroimmunomodulation 2004, 11, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Zhou, J.; Webb, D.C. Estrogen stimulates Th2 cytokine production and regulates the compartmentalisation of eosinophils during allergen challenge in a mouse model of asthma. Int. Arch. Allergy Immunol. 2012, 158, 252–260. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.P.; Domingos, H.V.; Cavriani, G.; Damazo, A.S.; Dos Santos Franco, A.L.; Oliani, S.M.; Oliveira-Filho, R.M.; Vargaftig, B.B.; de Lima, W.T. Cellular recruitment and cytokine generation in a rat model of allergic lung inflammation are differentially modulated by progesterone and estradiol. Am. J. Physiol. Cell Physiol. 2007, 293, C1120–C1128. [Google Scholar] [CrossRef] [PubMed]
- Hellings, P.W.; Vandekerckhove, P.; Claeys, R.; Billen, J.; Kasran, A.; Ceuppens, J.L. Progesterone increases airway eosinophilia and hyper-responsiveness in a murine model of allergic asthma. Clin. Exp. Allergy 2003, 33, 1457–1463. [Google Scholar] [CrossRef] [PubMed]
- Itoga, M.; Konno, Y.; Moritoki, Y.; Saito, Y.; Ito, W.; Tamaki, M.; Kobayashi, Y.; Kayaba, H.; Kikuchi, Y.; Chihara, J.; et al. G-protein-coupled estrogen receptor agonist suppresses airway inflammation in a mouse model of asthma through IL-10. PLoS ONE 2015, 10, e0123210. [Google Scholar]
- Dimitropoulou, C.; White, R.E.; Ownby, D.R.; Catravas, J.D. Estrogen reduces carbachol-induced constriction of asthmatic airways by stimulating large-conductance voltage and calcium-dependent potassium channels. Am. J. Respir. Cell Mol. Biol. 2005, 32, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, S.; Swasey, C.H.; Loader, J.E.; Dakhama, A.; Joetham, A.; Ohnishi, H.; Balhorn, A.; Miyahara, N.; Takeda, K.; Gelfand, E.W. Estrogen determines sex differences in airway responsiveness after allergen exposure. Am. J. Respir. Cell Mol. Biol. 2008, 38, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.K.; Yang, B.C.; Lei, H.Y.; Chen, Y.C.; Liu, Y.H.; Chen, C.C.; Liu, C.W. Attenuation of house dust mite dermatophagoides farinae-induced airway allergic responses in mice by dehydroepiandrosterone is correlated with down-regulation of Th2 response. Clin. Exp. Allergy 1999, 29, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Wulfsohn, N.L.; Politzer, W.M.; Henrico, J.S. Testosterone therapy in bronchial asthma. S. Afr. Med. J. 1964, 38, 170–172. [Google Scholar] [PubMed]
- Laffont, S.; Blanquart, E.; Savignac, M.; Cénac, C.; Laverny, G.; Metzger, D.; Girard, J.-P.; Belz, G.T.; Pelletier, L.; Seillet, C.; et al. Androgen signaling negatively controls group 2 innate lymphoid cells. J. Exp. Med. 2017, 214, 1581–1592. [Google Scholar] [CrossRef] [PubMed]
- Siroux, V.; Curt, F.; Oryszczyn, M.P.; Maccario, J.; Kauffmann, F. Role of gender and hormone-related events on IgE, atopy, and eosinophils in the epidemiological study on the genetics and environment of asthma, bronchial hyperresponsiveness and atopy. J. Allergy Clin. Immunol. 2004, 114, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Salam, M.T.; Wenten, M.; Gilliland, F.D. Endogenous and exogenous sex steroid hormones and asthma and wheeze in young women. J. Allergy Clin. Immunol. 2006, 117, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Lange, P.; Parner, J.; Prescott, E.; Ulrik, C.S.; Vestbo, J. Exogenous female sex steroid hormones and risk of asthma and asthma-like symptoms: A cross sectional study of the general population. Thorax 2001, 56, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Perez, B.; Henriquez, C.; Sarmiento, J.; Morales, N.; Folch, H.; Galesio, J.S.; Uberti, B.; Moran, G. Tamoxifen as a new therapeutic tool for neutrophilic lung inflammation. Respirology 2016, 21, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Abdou, N.I.; Rider, V.; Greenwell, C.; Li, X.; Kimler, B.F. Fulvestrant (faslodex), an estrogen selective receptor downregulator, in therapy of women with systemic lupus erythematosus. Clinical, serologic, bone density, and T cell activation marker studies: A double-blind placebo-controlled trial. J. Rheumatol. 2008, 35, 797. [Google Scholar] [PubMed]
- Madak-Erdogan, Z.; Kim, S.H.; Gong, P.; Zhao, Y.C.; Zhang, H.; Chambliss, K.L.; Carlson, K.E.; Mayne, C.G.; Shaul, P.W.; Korach, K.S.; et al. Design of pathway preferential estrogens that provide beneficial metabolic and vascular effects without stimulating reproductive tissues. Sci. Signal. 2016, 9, ra53. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leffler, J.; Stumbles, P.A.; Strickland, D.H. Immunological Processes Driving IgE Sensitisation and Disease Development in Males and Females. Int. J. Mol. Sci. 2018, 19, 1554. https://doi.org/10.3390/ijms19061554
Leffler J, Stumbles PA, Strickland DH. Immunological Processes Driving IgE Sensitisation and Disease Development in Males and Females. International Journal of Molecular Sciences. 2018; 19(6):1554. https://doi.org/10.3390/ijms19061554
Chicago/Turabian StyleLeffler, Jonatan, Philip A. Stumbles, and Deborah H. Strickland. 2018. "Immunological Processes Driving IgE Sensitisation and Disease Development in Males and Females" International Journal of Molecular Sciences 19, no. 6: 1554. https://doi.org/10.3390/ijms19061554
APA StyleLeffler, J., Stumbles, P. A., & Strickland, D. H. (2018). Immunological Processes Driving IgE Sensitisation and Disease Development in Males and Females. International Journal of Molecular Sciences, 19(6), 1554. https://doi.org/10.3390/ijms19061554