mTOR Pathway in Papillary Thyroid Carcinoma: Different Contributions of mTORC1 and mTORC2 Complexes for Tumor Behavior and SLC5A5 mRNA Expression

 , ,
, , 
Abstract
1. Introduction
2. Results
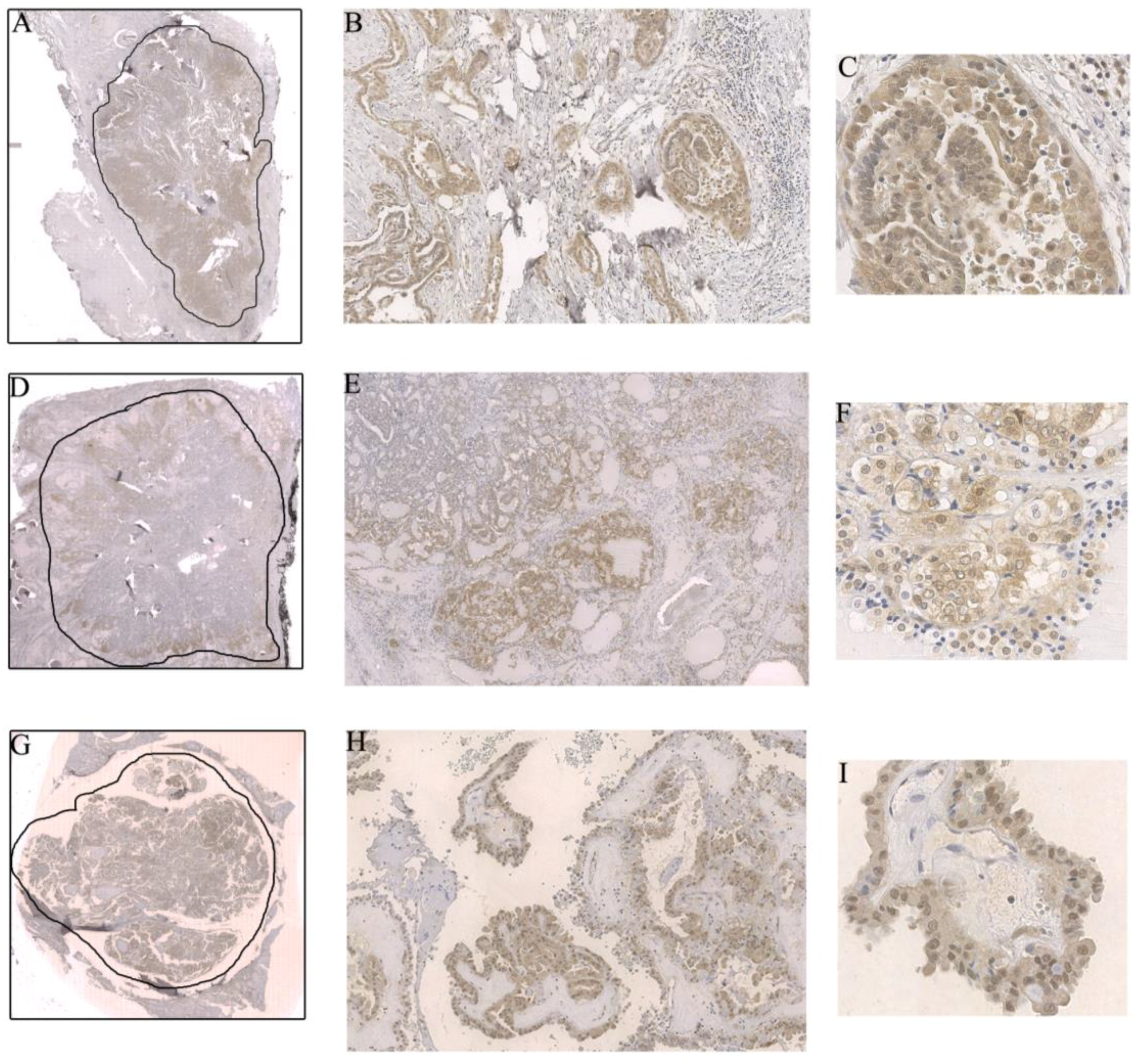
2.1. Immunoexpression of Phospho-AKT Ser473 in PTC
2.2. Relationship between the Phospho-AKT Ser473 Expression and Clinicopathological and Molecular Features
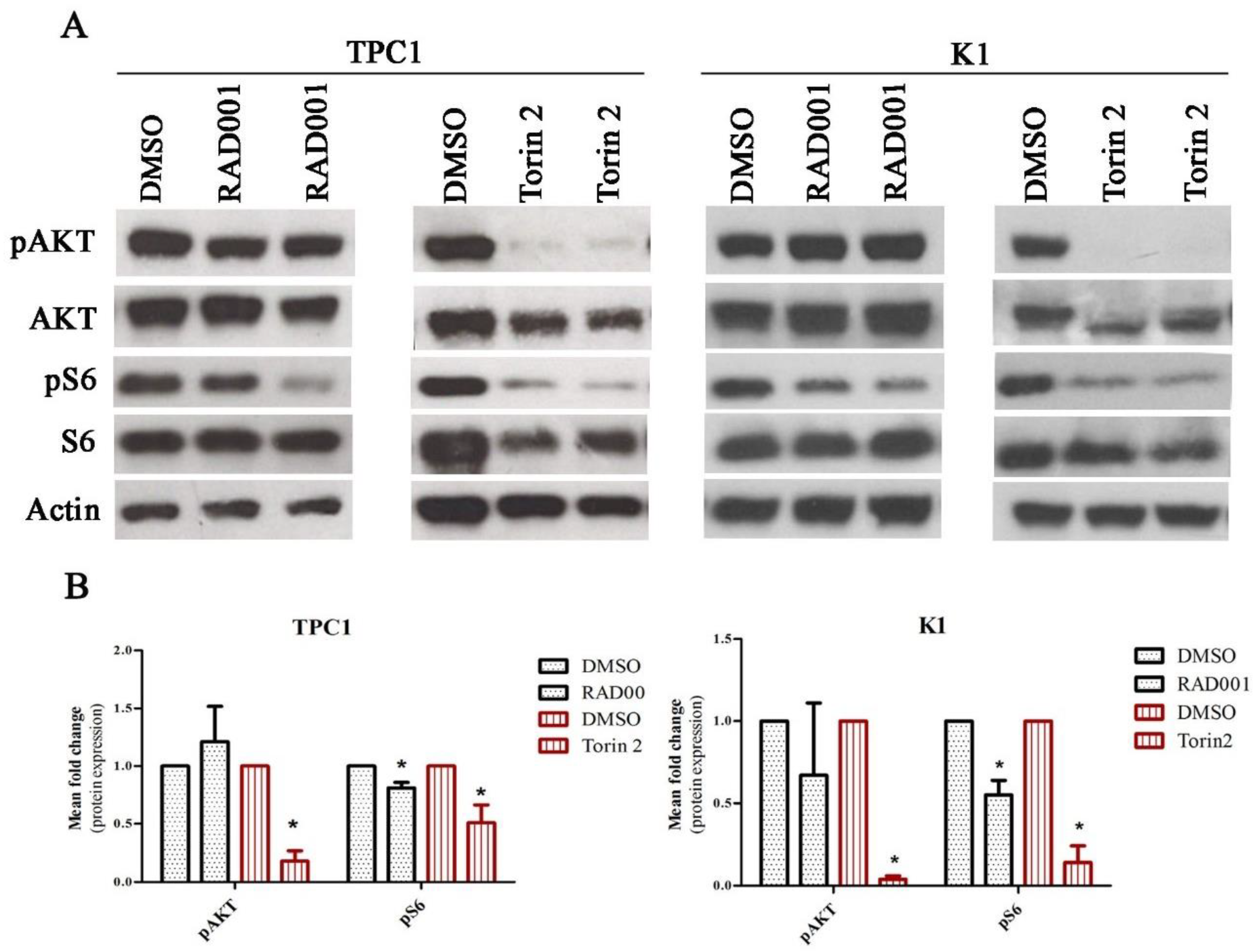
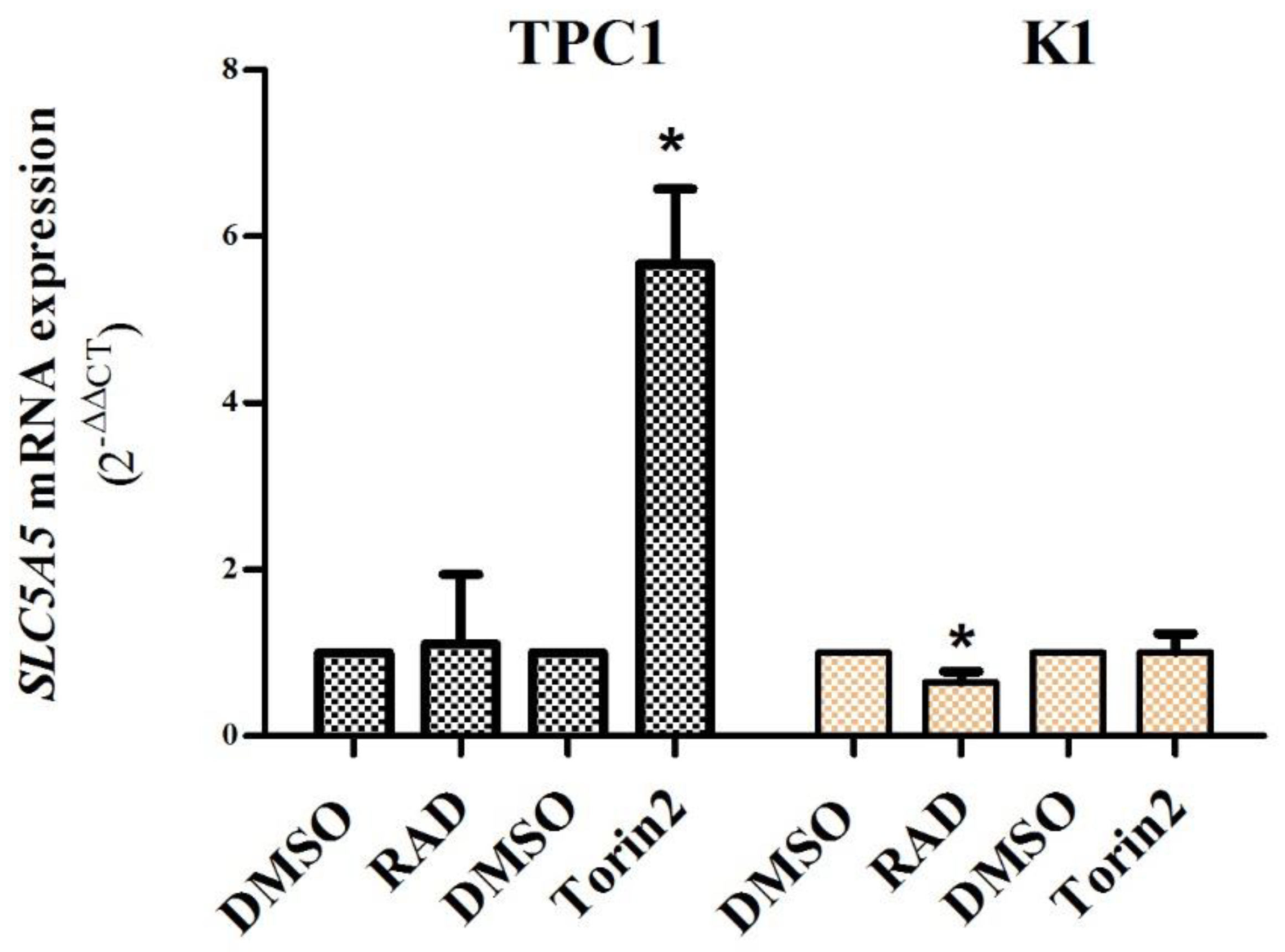
2.3. Contribution of mTORC1 and mTORC2 Complexes in the Regulation of SLC5A5 mRNA Expression
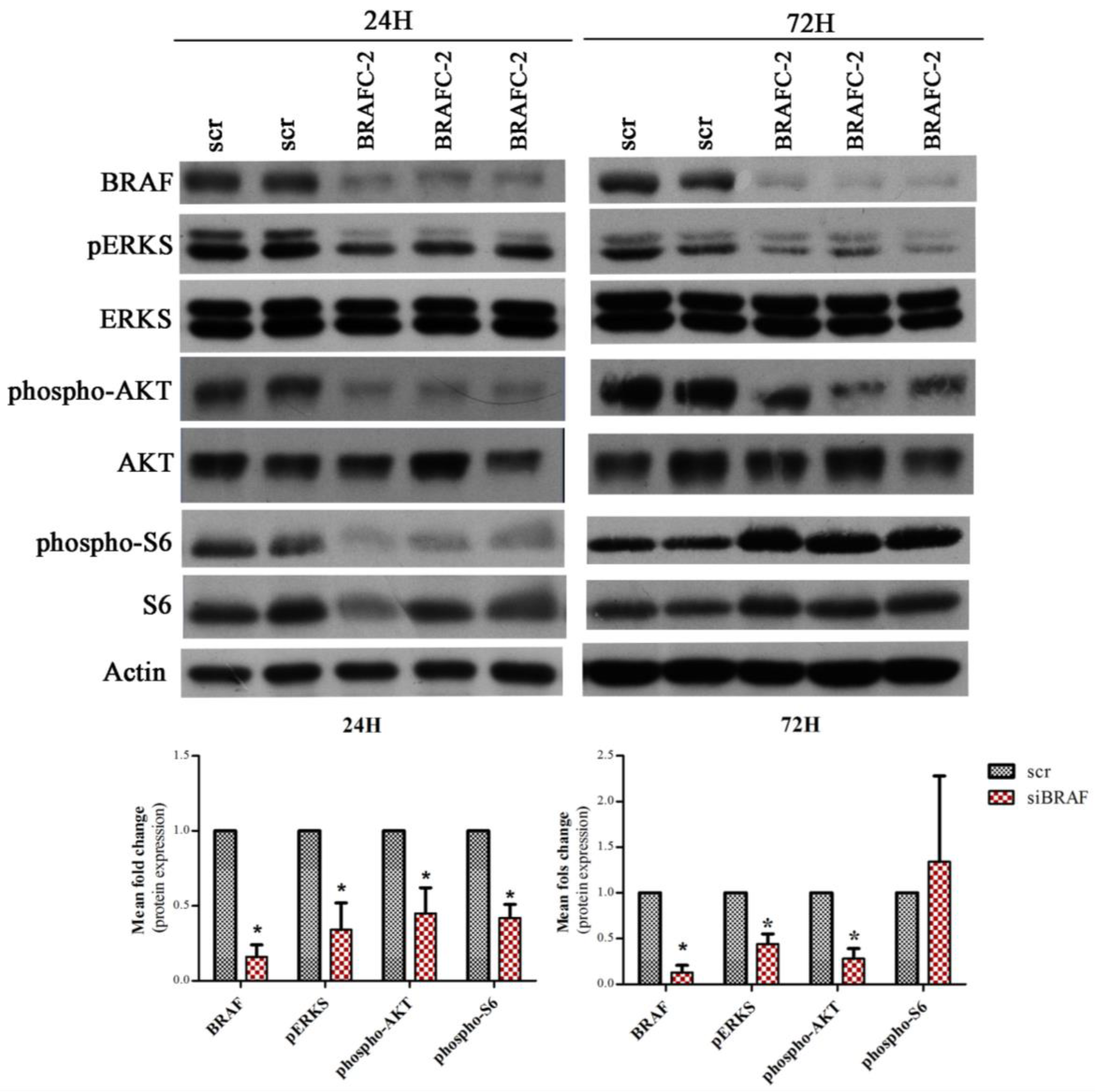
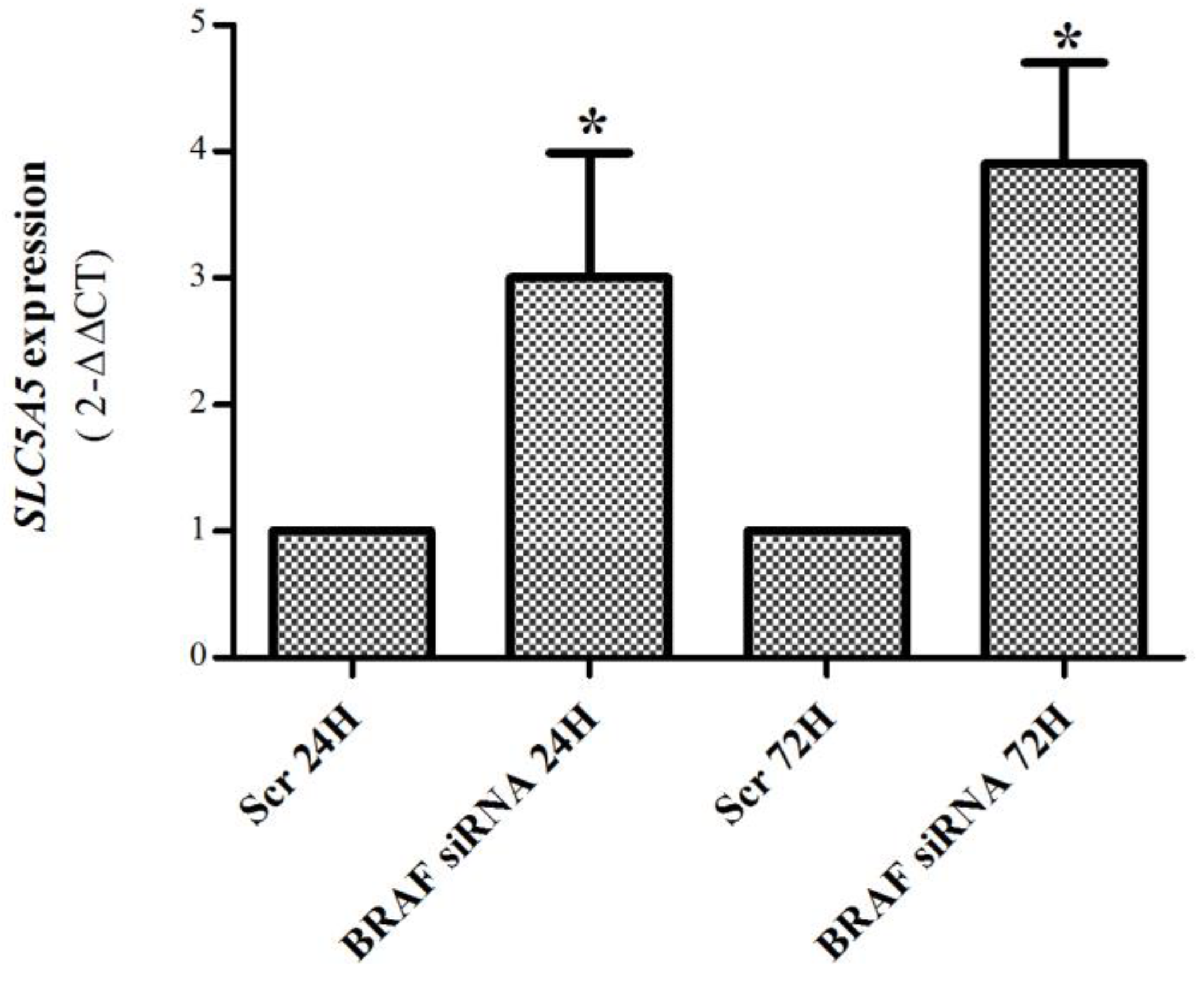
2.4. BRAF Regulation of SLC5A5 mRNA Expression and of mTORC1 and mTORC2 Status in K1 Cell Line
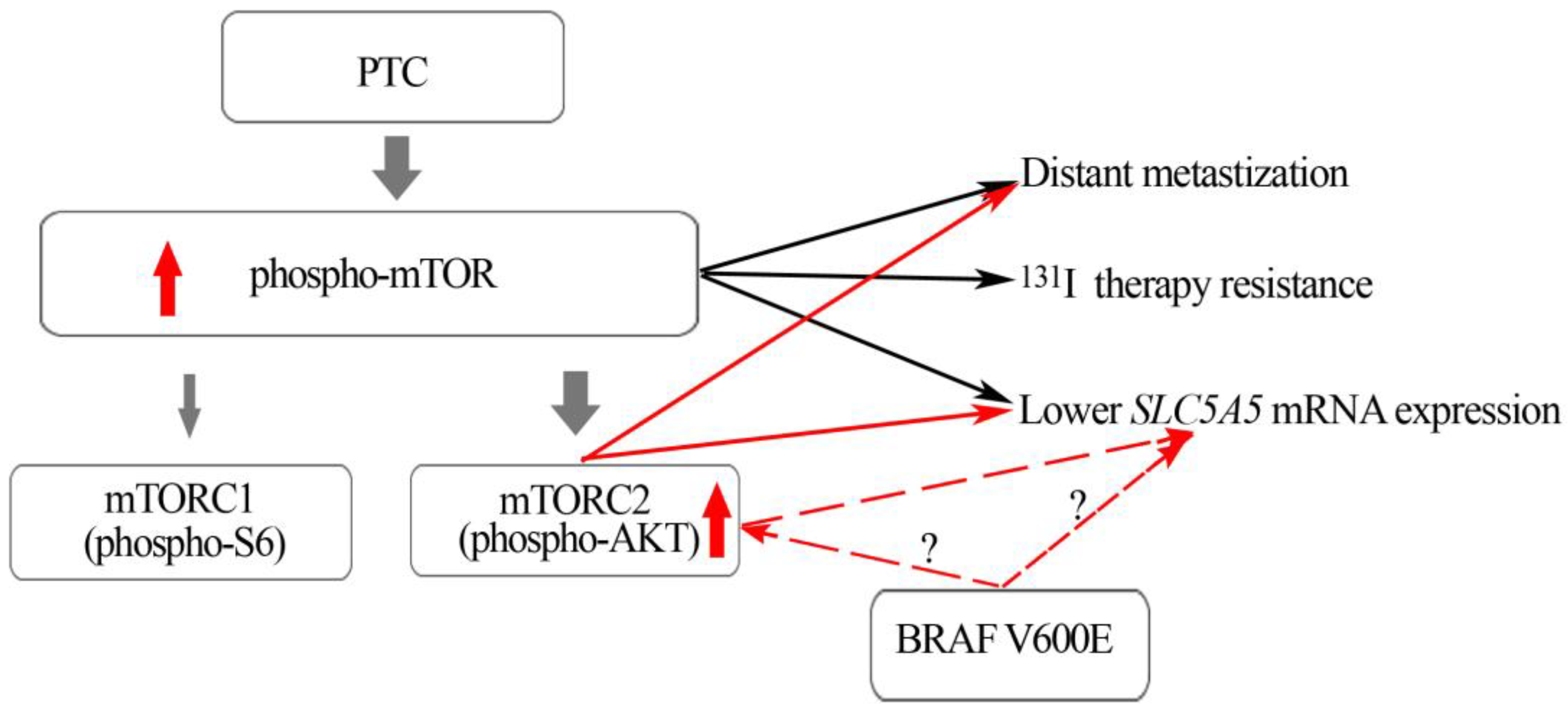
3. Discussion
4. Materials and Methods
4.1. Patient Tissue Samples
4.2. Patient’s Follow Up
4.3. Immunohistochemistry
4.4. DNA Extraction, PCR and Sanger Sequencing
4.5. Cell Lines, Treatments with RAD001 and Torin2 and Transfection Assays
4.6. RNA Extraction, Reverse Transcription and Real Time PCR
4.7. Western Blot Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| TC | thyroid cancer |
| PTC | papillary thyroid carcinoma |
| SLC5A5 | Solute Carrier Family 5 Member 5 |
| cPTC | classic PTC |
| fvPTC | follicular variant of PTC |
| NIS | sodium iodide symporter |
| RAI | radioactive iodine |
References
- Sipos, J.A.; Mazzaferri, E.L. Thyroid cancer epidemiology and prognostic variables. Clin. Oncol. 2010, 22, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Petrulea, M.S.; Plantinga, T.S.; Smit, J.W.; Georgescu, C.E.; Netea-Maier, R.T. PI3K/Akt/mTOR: A promising therapeutic target for non-medullary thyroid carcinoma. Cancer Treat. Rev. 2015, 41, 707–713. [Google Scholar] [CrossRef] [PubMed]
- LiVolsi, V.A. Papillary thyroid carcinoma: An update. Mod. Pathol. 2011, 24 (Suppl. 2), S1–S9. [Google Scholar] [CrossRef] [PubMed]
- Soares, P.; Celestino, R.; Melo, M.; Fonseca, E.; Sobrinho-Simoes, M. Prognostic biomarkers in thyroid cancer. Virchows Archiv Int. J. Pathol. 2014, 464, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Durante, C.; Haddy, N.; Baudin, E.; Leboulleux, S.; Hartl, D.; Travagli, J.P.; Caillou, B.; Ricard, M.; Lumbroso, J.D.; De Vathaire, F.; et al. Long-term outcome of 444 patients with distant metastases from papillary and follicular thyroid carcinoma: Benefits and limits of radioiodine therapy. J. Clin. Endocrinol. Metab. 2006, 91, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Fallahi, P.; Mazzi, V.; Vita, R.; Ferrari, S.M.; Materazzi, G.; Galleri, D.; Benvenga, S.; Miccoli, P.; Antonelli, A. New therapies for dedifferentiated papillary thyroid cancer. Int. J. Mol. Sci. 2015, 16, 6153–6182. [Google Scholar] [CrossRef] [PubMed]
- Vaisman, F.; Carvalho, D.P.; Vaisman, M. A new appraisal of iodine refractory thyroid cancer. Endocr.-Relat. Cancer 2015, 22, R301–R310. [Google Scholar] [CrossRef] [PubMed]
- De Souza, E.C.L.; Freitas Ferreira, A.C.; de Carvalho, D.P. The mTOR protein as a target in thyroid cancer. Expert Opin. Ther. Targets 2011, 15, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Faustino, A.; Couto, J.P.; Populo, H.; Rocha, A.S.; Pardal, F.; Cameselle-Teijeiro, J.M.; Lopes, J.M.; Sobrinho-Simoes, M.; Soares, P. mTOR pathway overactivation in BRAF mutated papillary thyroid carcinoma. J. Clin. Endocrinol. Metab. 2012, 97, E1139–E1149. [Google Scholar] [CrossRef] [PubMed]
- Populo, H.; Lopes, J.M.; Soares, P. The mTOR signalling pathway in human cancer. Int. J. Mol. Sci. 2012, 13, 1886–1918. [Google Scholar] [CrossRef] [PubMed]
- Miyakawa, M.; Tsushimma, T.; Murakami, H.; Wakai, K.; Isozaki, O.; Takano, K. Increased expression of phosphorylated p70S6 kinase and Akt in papillary thyroid cancer tissues. Endocr. J. 2003, 50, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Vasko, V.; Saji, M.; Hardy, E.; Kruhlak, M.; Larin, A.; Savchenko, V.; Miyakawa, M.; Isozaki, O.; Murakami, H.; Tsushima, T.; et al. Akt activation and localisation correlate with tumour invasion and oncogene expression in thyroid cancer. J. Med. Genet. 2004, 41, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Tavares, C.; Coelho, M.J.; Melo, M.; da Rocha, A.G.; Pestana, A.; Batista, R.; Salgado, C.; Eloy, C.; Ferreira, L.; Rios, E.; et al. pmTOR is a marker of aggressiveness in papillary thyroid carcinomas. Surgery 2016, 160, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Hau, A.M.; Beach, J.R.; Harwalker, J.; Mantuano, E.; Gonias, S.L.; Egelhoff, T.T.; Hansel, D.E. Mammalian target of rapamycin complex 2 (mTORC2) is a critical determinant of bladder cancer invasion. PLoS ONE 2013, 8, e81081. [Google Scholar] [CrossRef] [PubMed]
- Masri, J.; Bernath, A.; Martin, J.; Jo, O.D.; Vartanian, R.; Funk, A.; Gera, J. mTORC2 activity is elevated in gliomas and promotes growth and cell motility via overexpression of rictor. Cancer Res. 2007, 67, 11712–11720. [Google Scholar] [CrossRef] [PubMed]
- Bian, Y.; Wang, Z.; Xu, J.; Zhao, W.; Cao, H.; Zhang, Z. Elevated Rictor expression is associated with tumor progression and poor prognosis in patients with gastric cancer. Biochem. Biophys. Res. Commun. 2015, 21, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Maru, S.; Ishigaki, Y.; Shinohara, N.; Takata, T.; Tomosugi, N.; Nonomura, K. Inhibition of mTORC2 but not mTORC1 up-regulates E-cadherin expression and inhibits cell motility by blocking HIF-2α expression in human renal cell carcinoma. J. Urol. 2013, 189, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Matson, D.R.; Hardin, H.; Buehler, D.; Lloyd, R.V. AKT activity is elevated in aggressive thyroid neoplasms where it promotes proliferation and invasion. Exp. Mol. Pathol. 2017, 103, 288–293. [Google Scholar] [CrossRef] [PubMed]
- De Souza, E.C.; Padron, A.S.; Braga, W.M.; de Andrade, B.M.; Vaisman, M.; Nasciutti, L.E.; Ferreira, A.C.; de Carvalho, D.P. MTOR downregulates iodide uptake in thyrocytes. J. Endocrinol. 2010, 206, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Plantinga, T.S.; Heinhuis, B.; Gerrits, D.; Netea, M.G.; Joosten, L.A.; Hermus, A.R.; Oyen, W.J.; Schweppe, R.E.; Haugen, B.R.; Boerman, O.C.; et al. mTOR Inhibition promotes TTF1-dependent redifferentiation and restores iodine uptake in thyroid carcinoma cell lines. J. Clin. Endocrinol. Metab. 2014, 99, E1368–E1375. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Hussain, A.R.; Bavi, P.; Ahmed, S.O.; Al Sobhi, S.S.; Al-Dayel, F.; Uddin, S.; Al-Kuraya, K.S. High prevalence of mTOR complex activity can be targeted using Torin2 in papillary thyroid carcinoma. Carcinogenesis 2014, 35, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Sadowski, S.M.; Boufragech, M.; Zhang, L.; Mehta, A.; Kapur, P.; Zhang, Y.; Li, Z.; Shen, M.; Kebebew, E. Torin2 targets dysregulated pathways in anaplastic thyroid cancer and inhibits tumor growth and metastasis. Oncotarget 2015, 6, 18038–18049. [Google Scholar] [CrossRef] [PubMed]
- Riesco-Eizaguirre, G.; Gutierrez-Martinez, P.; Garcia-Cabezas, M.A.; Nistal, M.; Santisteban, P. The oncogene BRAF V600E is associated with a high risk of recurrence and less differentiated papillary thyroid carcinoma due to the impairment of Na+/I− targeting to the membrane. Endocr.-Relat. Cancer 2006, 13, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Ringel, M.D.; Hayre, N.; Saito, J.; Saunier, B.; Schuppert, F.; Burch, H.; Bernet, V.; Burman, K.D.; Kohn, L.D.; Saji, M. Overexpression and overactivation of Akt in thyroid carcinoma. Cancer Res. 2001, 61, 6105–6111. [Google Scholar] [PubMed]
- Saji, M.; Narahara, K.; McCarty, S.K.; Vasko, V.V.; la Perle, K.M.; Porter, K.; Jarjoura, D.; Cheng, S.-Y.; Lu, C.; Ringel, M.D. Akt deficiency delays tumor progression, vascular invasion, and distant metastases in a murine model of thyroid cancer. Oncogene 2011, 30, 4307–4315. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Vasko, V.V.; Kato, Y.; Kruhlak, M.; Saji, M.; Cheng, S.Y.; Ringel, M.D. AKT activation promotes metastasis in a mouse model of follicular thyroid carcinoma. Endocrinology 2005, 146, 4456–4463. [Google Scholar] [CrossRef] [PubMed]
- Pende, M.; Um, S.H.; Mieulet, V.; Sticker, M.; Goss, V.L.; Mestan, J.; Mueller, M.; Fumagalli, S.; Kozma, S.C.; Thomas, G. S6K1−/−/S6K2−/− Mice Exhibit Perinatal Lethality and Rapamycin-Sensitive 5′-Terminal Oligopyrimidine mRNA Translation and Reveal a Mitogen-Activated Protein Kinase-Dependent S6 Kinase Pathway. Mol. Cell Biol. 2004, 24, 3112–3124. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.P.; Shahbazian, D.; Vu, H.; Holz, M.K.; Cohen, M.S.; Taunton, J.; Sonenberg, N.; Blenis, J. RAS/ERK signaling promotes site-specific ribosomal protein S6 phosphorylation via RSK and stimulates cap-dependent translation. J. Biol. Chem. 2007, 282, 14056–14064. [Google Scholar] [CrossRef] [PubMed]
- Breuleux, M.; Klopfenstein, M.; Stephan, C.; Doughty, C.A.; Barys, L.; Maira, S.-M.; Kwiatkowski, D.; Lane, H.A. Increased AKT S473 phosphorylation after mTORC1 inhibition is rictor dependent and does not predict tumor cell response to PI3K/mTOR inhibition. Mol. Cancer Ther. 2009, 8, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Tavares, C.; Coelho, M.J.; Eloy, C.; Melo, M.; da Rocha, A.G.; Pestana, A.; Batista, R.; Ferreira, L.B.; Rios, E.; Selmi-Ruby, S.; et al. NIS expression in thyroid tumors, relation with prognosis clinicopathological and molecular features. Endocr. Connect. 2018, 7, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Meireles, A.M.; Preto, A.; Rocha, A.S.; Rebocho, A.P.; Maximo, V.; Pereira-Castro, I.; Moreira, S.; Feijao, T.; Botelho, T.; Marques, R.; et al. Molecular and genotypic characterization of human thyroid follicular cell carcinoma-derived cell lines. Thyroid Off. J. Am. Thyroid Assoc. 2007, 17, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Durante, C.; Puxeddu, E.; Ferretti, E.; Morisi, R.; Moretti, S.; Bruno, R.; Barbi, F.; Avenia, N.; Scipioni, A.; Verrienti, A.; et al. BRAF mutations in papillary thyroid carcinomas inhibit genes involved in iodine metabolism. J. Clin. Endocrinol. Metab. 2007, 92, 2840–2843. [Google Scholar] [CrossRef] [PubMed]
- DeLellis, R.A.; Lloyd, R.V.; Heitz, P.U.; Eng, C.; WHO Classification of Tumours. Pathology and Genetics of Tumours of Endocrine Organs; IARC Press: Lyon, France, 2004. [Google Scholar]
- Soares, P.; Trovisco, V.; Rocha, A.S.; Lima, J.; Castro, P.; Preto, A.; Maximo, V.; Botelho, T.; Seruca, R.; Sobrinho-Simoes, M. BRAF mutations and RET/PTC rearrangements are alternative events in the etiopathogenesis of PTC. Oncogene 2003, 22, 4578–4580. [Google Scholar] [CrossRef] [PubMed]
- Trovisco, V.; Soares, P.; Preto, A.; de Castro, I.V.; Lima, J.; Castro, P.; Maximo, V.; Botelho, T.; Moreira, S.; Meireles, A.M.; et al. Type and prevalence of BRAF mutations are closely associated with papillary thyroid carcinoma histotype and patients’ age but not with tumour aggressiveness. Virchows Archiv Int. J. Pathol. 2005, 446, 589–595. [Google Scholar] [CrossRef] [PubMed]
- De Vries, M.M.; Celestino, R.; Castro, P.; Eloy, C.; Maximo, V.; van der Wal, J.E.; Plukker, J.T.; Links, T.P.; Hofstra, R.M.; Sobrinho-Simoes, M.; et al. RET/PTC rearrangement is prevalent in follicular Hurthle cell carcinomas. Histopathology 2012, 61, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.; da Rocha, A.G.; Vinagre, J.; Batista, R.; Peixoto, J.; Tavares, C.; Celestino, R.; Almeida, A.; Salgado, C.; Eloy, C.; et al. TERT promoter mutations are a major indicator of poor outcome in differentiated thyroid carcinomas. J. Clin. Endocrinol. Metab. 2014, 99, E754–E765. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, J.; Almeida, A.; Populo, H.; Batista, R.; Lyra, J.; Pinto, V.; Coelho, R.; Celestino, R.; Prazeres, H.; Lima, L.; et al. Frequency of TERT promoter mutations in human cancers. Nat. Commun. 2013, 4, 2185. [Google Scholar] [CrossRef] [PubMed]
- Preto, A.; Goncalves, J.; Rebocho, A.P.; Figueiredo, J.; Meireles, A.M.; Rocha, A.S.; Vasconcelos, H.M.; Seca, H.; Seruca, R.; Soares, P.; et al. Proliferation and survival molecules implicated in the inhibition of BRAF pathway in thyroid cancer cells harbouring different genetic mutations. BMC Cancer 2009, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phospho-AKT Score | Frequency | % |
|---|---|---|
| 0 | 90 | 49.5 |
| 1 | 18 | 9.9 |
| 2 | 15 | 8.2 |
| 3 | 6 | 3.3 |
| 4 | 8 | 4.4 |
| 6 | 14 | 7.7 |
| 8 | 11 | 6.0 |
| 9 | 6 | 3.3 |
| 12 | 14 | 7.7 |
| Total | 182 | 100 |
| BRAF | Phospho-AKT Score | p Value |
|---|---|---|
| WT (n = 106) | 2.2 ± 3.3 | 0.04 |
| V600E (n = 74) | 3.4 ± 4.4 |
| Nuclear Expression | Distant Metastases | p Value | |
|---|---|---|---|
| Yes | No | ||
| Yes | 9 (81.82%) | 19 (47.5%) | |
| No | 2 (18.18%) | 21 (52.5%) | 0.04 |
| Total | 11 | 40 | 51 |
| Patients | Total and % | |||
|---|---|---|---|---|
| cPTC | fvPTC | other PTC variants | ||
| Gender | F n = 150 | 94 (82.5) | 41 (87.2) | 15 (71.4) |
| M n = 32 | 20 (17.5) | 6 (12.8) | 6 (28.6) | |
| Age | <45 years n = 94 | 62 (54.9) | 21 (45.7) | 11 (55.0) |
| ≥45 years n = 85 | 51 (45.1) | 25 (54.3) | 9 (45.0) | |
| Tumor size | <2cm n = 64 | 39 (36.8) | 17 (37.0) | 8 (40.0) |
| ≥2cm n = 108 | 67 (63.2) | 29 (63.0) | 12 (60.0) | |
| Tumor capsule | Present n = 83 | 42 (39.6) | 32 (71.1) | 9 (42.9) |
| Absent n = 89 | 64 (60.4) | 13 (28.9) | 12 (57.1) | |
| Tumor capsule Invasion | Yes n = 64 | 35 (89.7) | 22 (68.8) | 7 (100) |
| No n = 14 | 4 (10.3) | 10 (31.3) | 0 (0) | |
| Extra thyroid invasion | Yes n = 73 | 50 (48.1) | 12 (27.3) | 11 (55.0) |
| No n = 95 | 54 (51.9) | 32 (72.7) | 9 (45.0) | |
| Multifocality | Single n = 104 | 58 (54.7) | 32 (68.1) | 14 (70.0) |
| Multiple n = 69 | 48 (45.3) | 15 (31.9) | 6 (30.0) | |
| Lymphocytic infiltrate | Present n = 108 | 77 (70.6) | 19 (41.3) | 12 (60.0) |
| Absent n = 67 | 32 (29.4) | 27 (58.7) | 8 (40.0) | |
| Vascular invasion | Present n = 59 | 42 (40.4) | 10 (22.2) | 7 (35.0) |
| Absent n = 110 | 62 (59.6) | 35 (77.8) | 13 (65.0) | |
| Lymph node metastases | Present n = 57 | 40 (43.0) | 12 (34.3) | 5 (29.4) |
| Absent n = 88 | 53 (57.0) | 23 (65.7) | 12 (70.6) | |
| Tumor margins | Infiltrative n = 78 | 57 (79.2) | 13 (46.4) | 8 (72.7) |
| Well defined n = 33 | 15 (20.8) | 15 (53.6) | 3 (27.3) | |
| Distant metastases | Yes n = 17 | 9 (11.8) | 5 (17.9) | 3 (30.0) |
| No n = 97 | 67 (88.2) | 23 (82.1) | 7 (70.0) | |
| One year disease free survival | Yes n = 64 | 41 (56.2) | 19 (67.9) | 4 (40.0) |
| No n = 47 | 32 (43.8) | 9 (32.1) | 6 (60.0) | |
| Disease free (at the end of follow up) | Yes n = 70 | 44 (59.5) | 19 (67.9) | 7 (70.0) |
| No n = 42 | 30 (40.5) | 9 (32.1) | 3 (30.0) | |
| Deaths | Yes n = 5 | 2 (2.6) | 2 (7.1) | 1 (9.1) |
| No n = 110 | 74 (97.4) | 26 (92.9) | 10 (90.9) | |
| BRAF | WT n = 106 | 56 (49.1) | 37 (82.2) | 13 (61.9) |
| V600E n = 74 | 58 (50.9) | 8 (17.8) | 8 (38.1) | |
| NRAS | WT n = 162 | 108 (99.1) | 38 (90.5) | 16 (80.0) |
| Mut n = 9 | 1 (0.9) | 4 (9.5) | 4 (20.0) | |
| TERTp | WT n = 152 | 95 (96.0) | 40 (95.2) | 17 (100.0) |
| Mut n = 6 | 4 (4.0) | 2 (4.8) | 0 (0.0) | |
| RET/PTC | WT n = 56 | 29 (78.4) | 18 (94.7) | 9 (90.0) |
| Rearrangment n = 10 | 8 (21.6) | 1 (5.3) | 1 (10.0) | |
| Staging | I n = 64 | 45 (64.3) | 15 (60.0) | 4 (50.0) |
| II n = 6 | 3 (4.3) | 3 (12.0) | 0 (0.0) | |
| III n = 24 | 19 (27.1) | 3 (12.0) | 2 (25.0) | |
| IV n = 9 | 3 (4.3) | 4 (16.0) | 2 (25.0) | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tavares, C.; Eloy, C.; Melo, M.; Gaspar da Rocha, A.; Pestana, A.; Batista, R.; Bueno Ferreira, L.; Rios, E.; Sobrinho Simões, M.; Soares, P. mTOR Pathway in Papillary Thyroid Carcinoma: Different Contributions of mTORC1 and mTORC2 Complexes for Tumor Behavior and SLC5A5 mRNA Expression. Int. J. Mol. Sci. 2018, 19, 1448. https://doi.org/10.3390/ijms19051448
Tavares C, Eloy C, Melo M, Gaspar da Rocha A, Pestana A, Batista R, Bueno Ferreira L, Rios E, Sobrinho Simões M, Soares P. mTOR Pathway in Papillary Thyroid Carcinoma: Different Contributions of mTORC1 and mTORC2 Complexes for Tumor Behavior and SLC5A5 mRNA Expression. International Journal of Molecular Sciences. 2018; 19(5):1448. https://doi.org/10.3390/ijms19051448
Chicago/Turabian StyleTavares, Catarina, Catarina Eloy, Miguel Melo, Adriana Gaspar da Rocha, Ana Pestana, Rui Batista, Luciana Bueno Ferreira, Elisabete Rios, Manuel Sobrinho Simões, and Paula Soares. 2018. "mTOR Pathway in Papillary Thyroid Carcinoma: Different Contributions of mTORC1 and mTORC2 Complexes for Tumor Behavior and SLC5A5 mRNA Expression" International Journal of Molecular Sciences 19, no. 5: 1448. https://doi.org/10.3390/ijms19051448
APA StyleTavares, C., Eloy, C., Melo, M., Gaspar da Rocha, A., Pestana, A., Batista, R., Bueno Ferreira, L., Rios, E., Sobrinho Simões, M., & Soares, P. (2018). mTOR Pathway in Papillary Thyroid Carcinoma: Different Contributions of mTORC1 and mTORC2 Complexes for Tumor Behavior and SLC5A5 mRNA Expression. International Journal of Molecular Sciences, 19(5), 1448. https://doi.org/10.3390/ijms19051448