Hyperinsulinemia Promotes Esophageal Cancer Development in a Surgically-Induced Duodeno-Esophageal Reflux Murine Model

 ,
,  ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Macroscopic Findings in Operated Wild-Type and MKR Mice
2.2. Histological Findings in Operated Wild-Type and MKR Mice
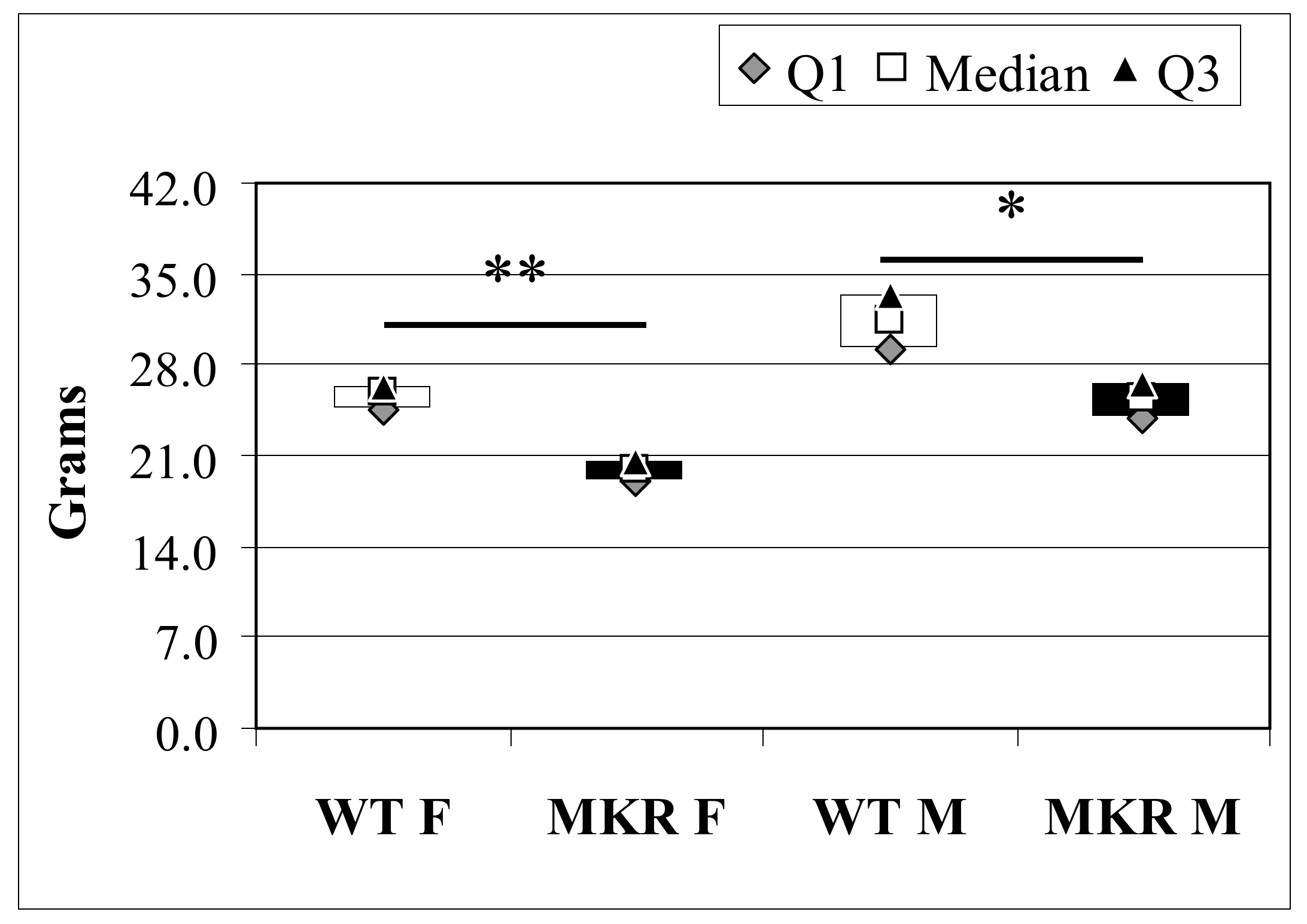
2.3. Lesion Histology and Gender Difference
2.4. Basal Serum Metabolic Parameters
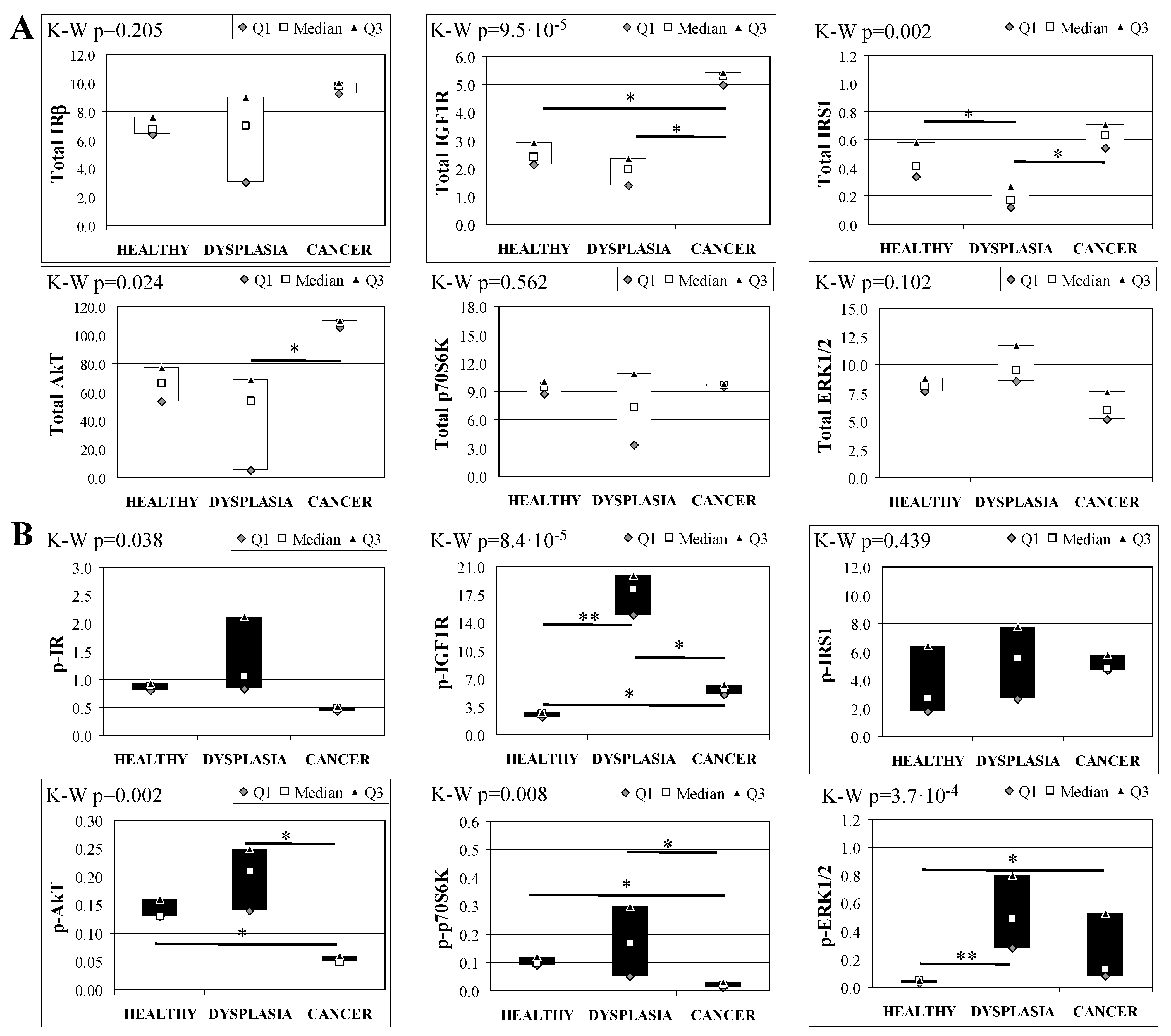
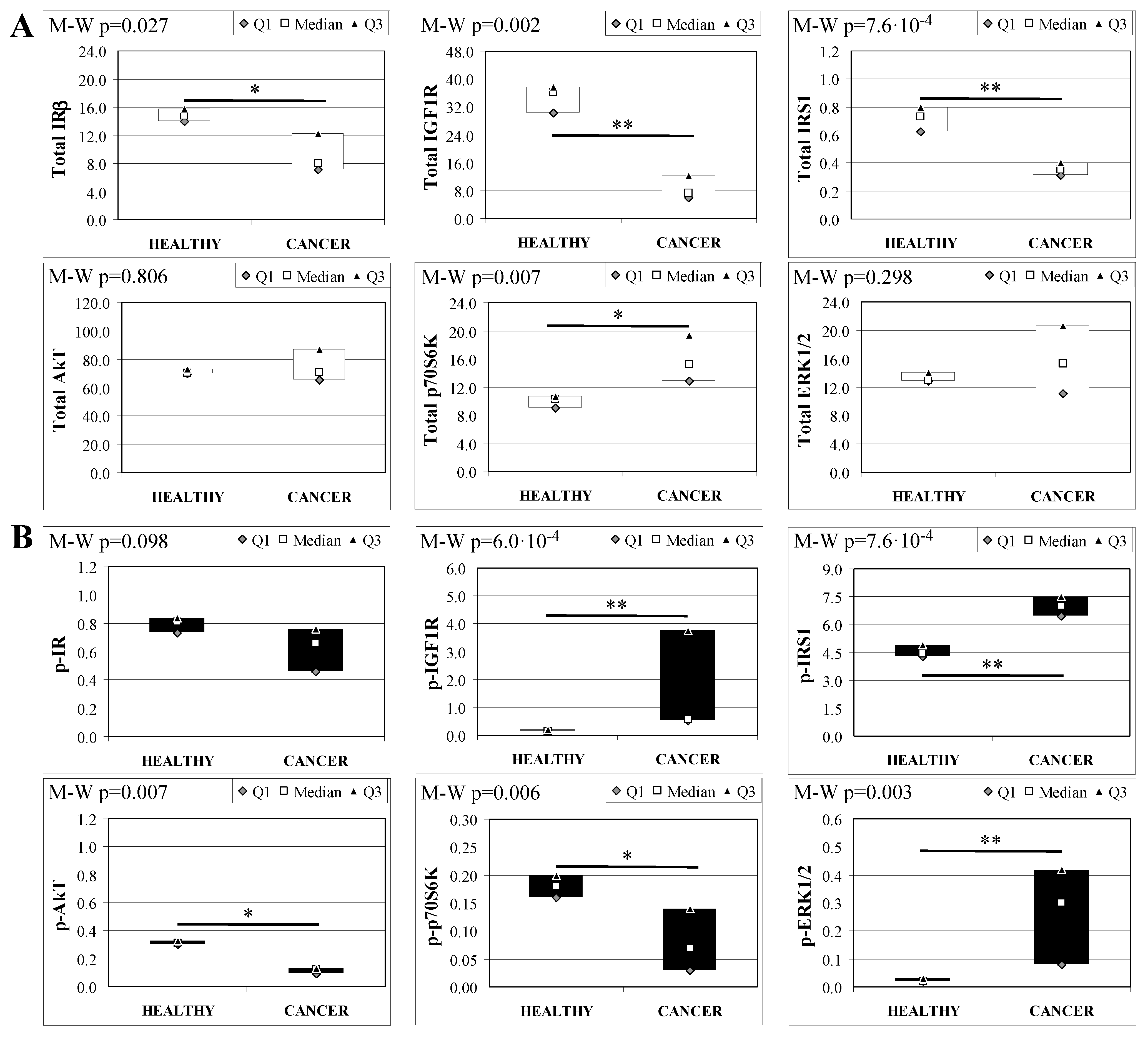
2.5. Basal Insulin Signaling in Mice Esophageal Tissue
2.6. Serum Metabolic Parameters in Operated Mice
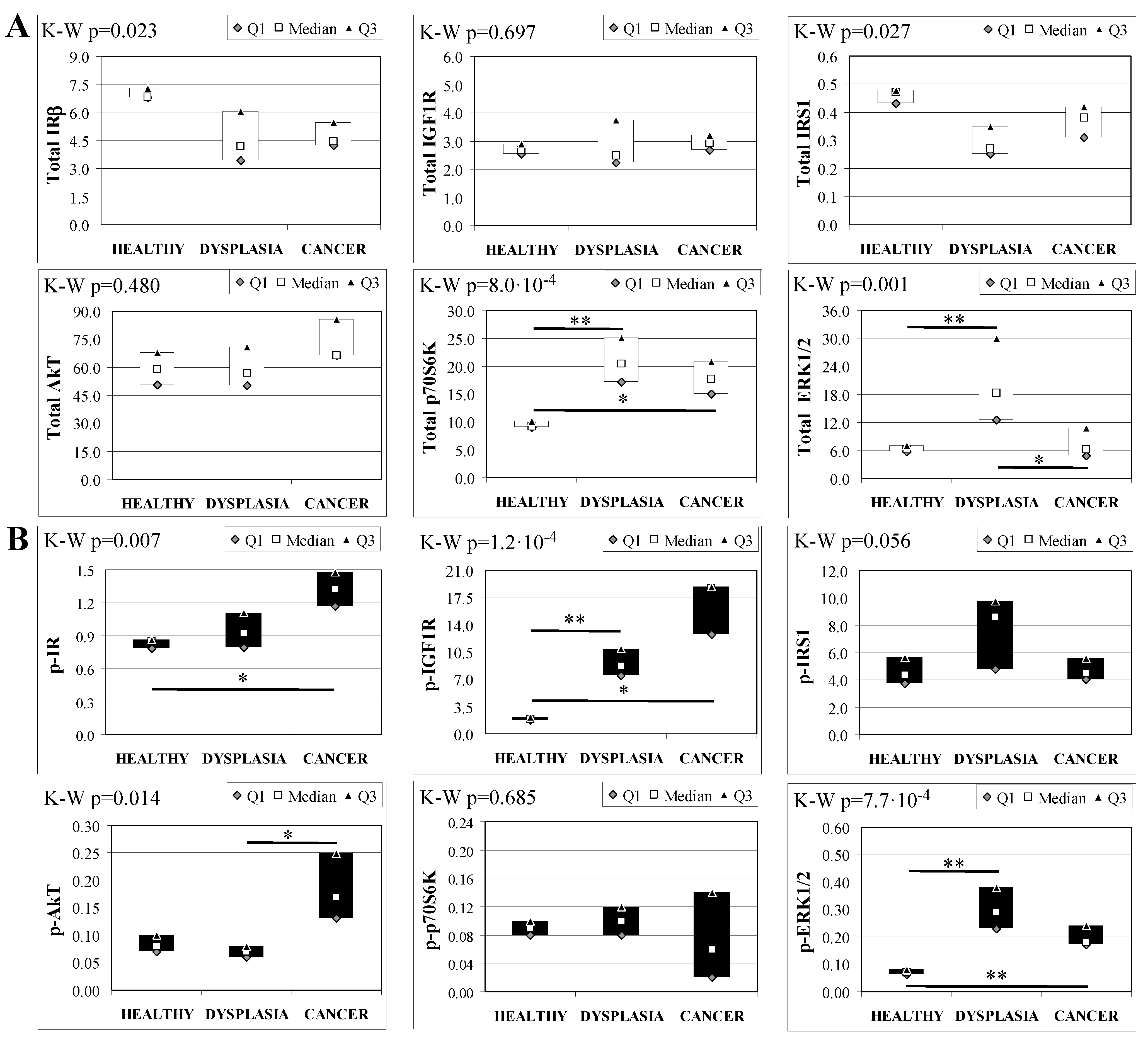
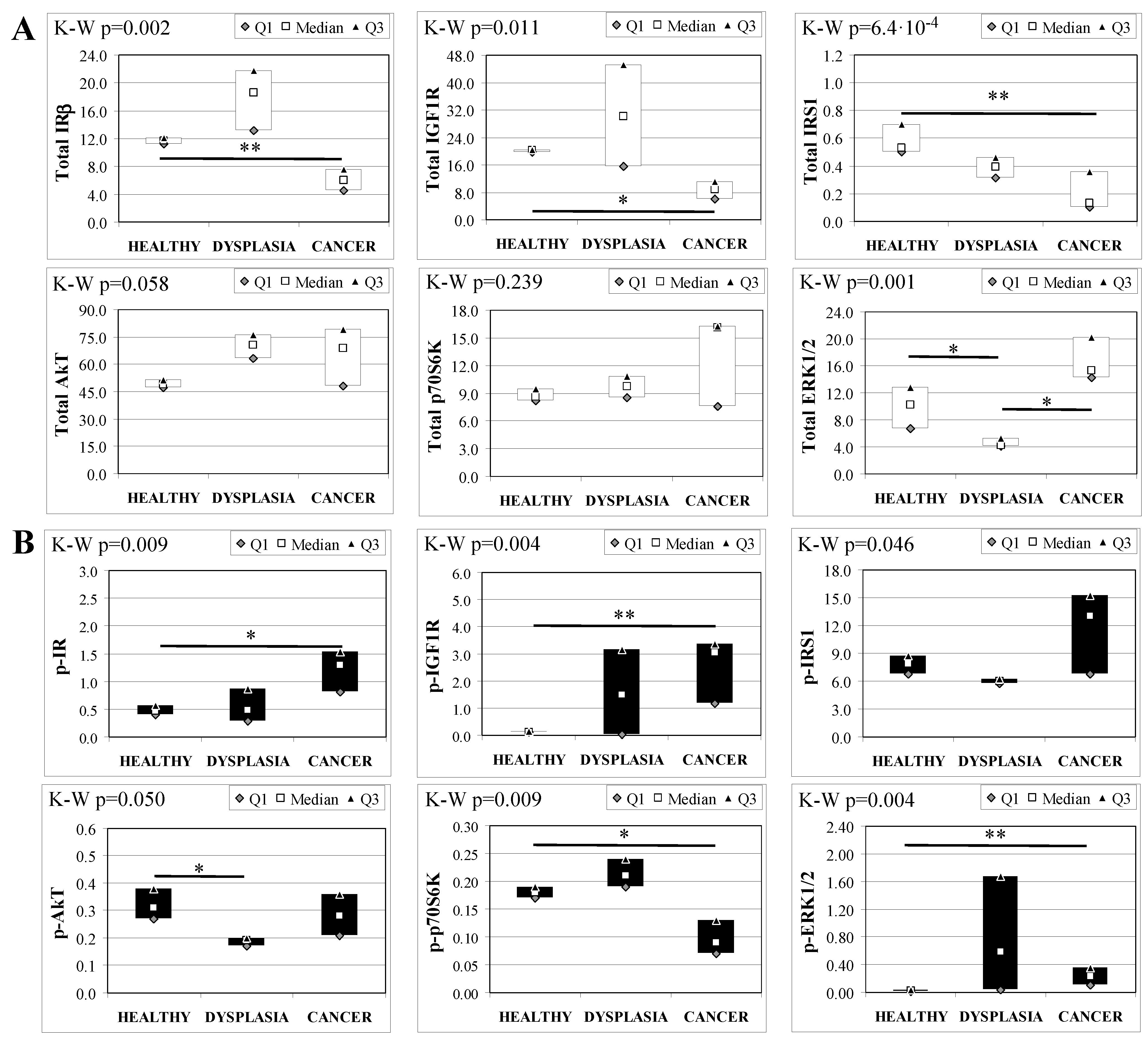
2.7. Insulin Signaling in Operated Mice Esophageal Tissue
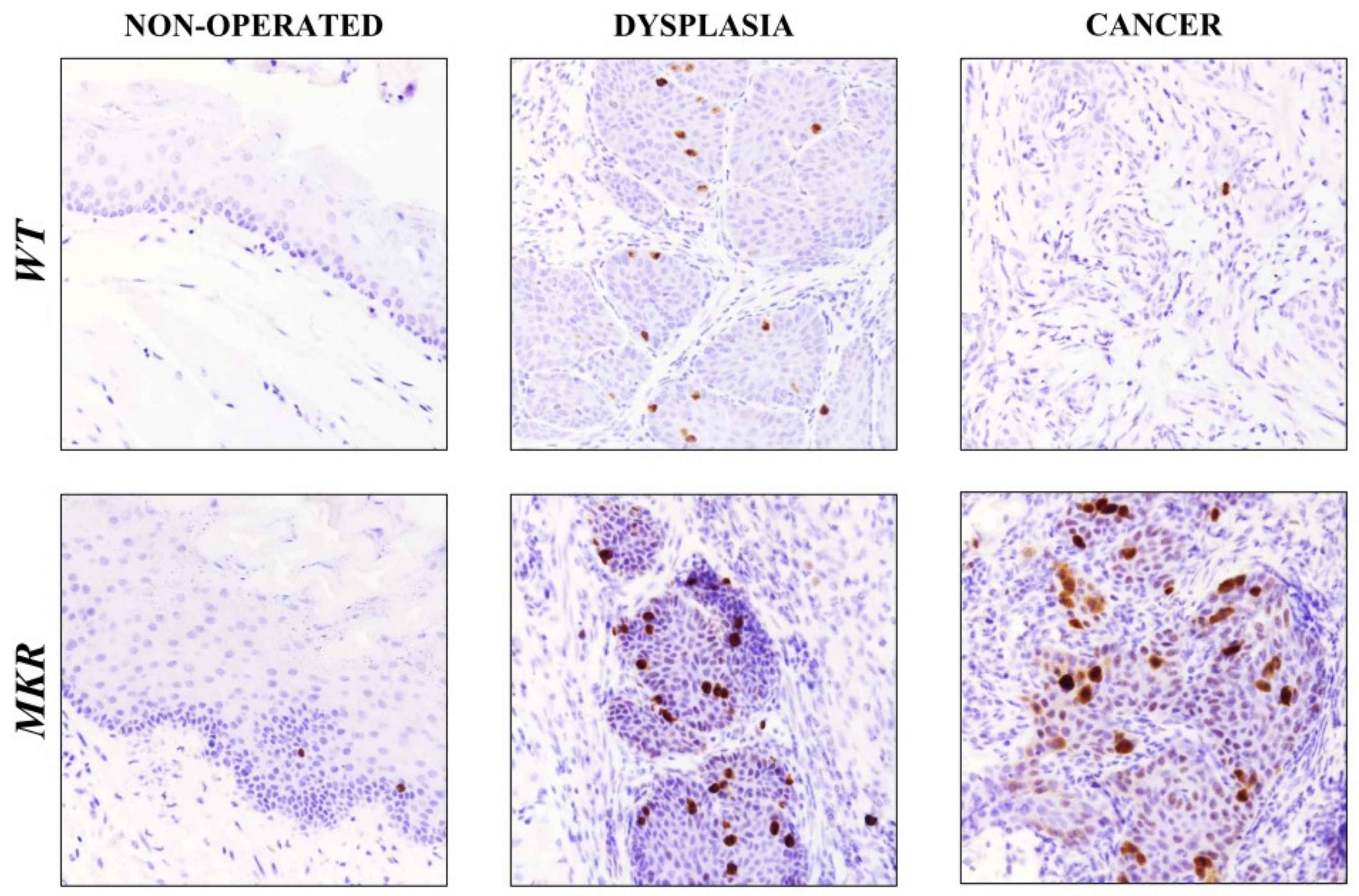
2.8. Ki-67 Nuclear Protein Expression on Esophageal Lesions
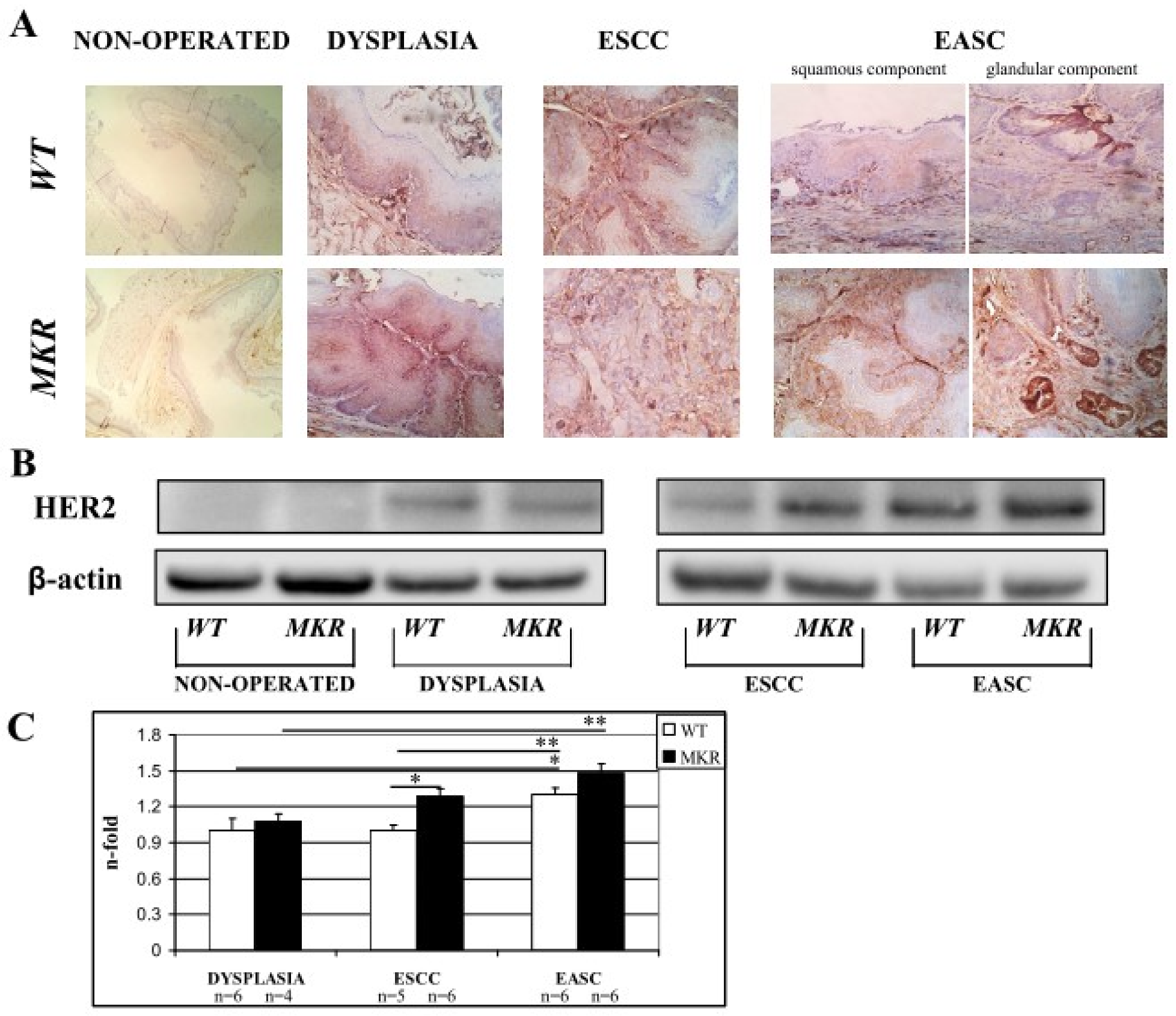
2.9. HER-2 Expression in Cancer Progression
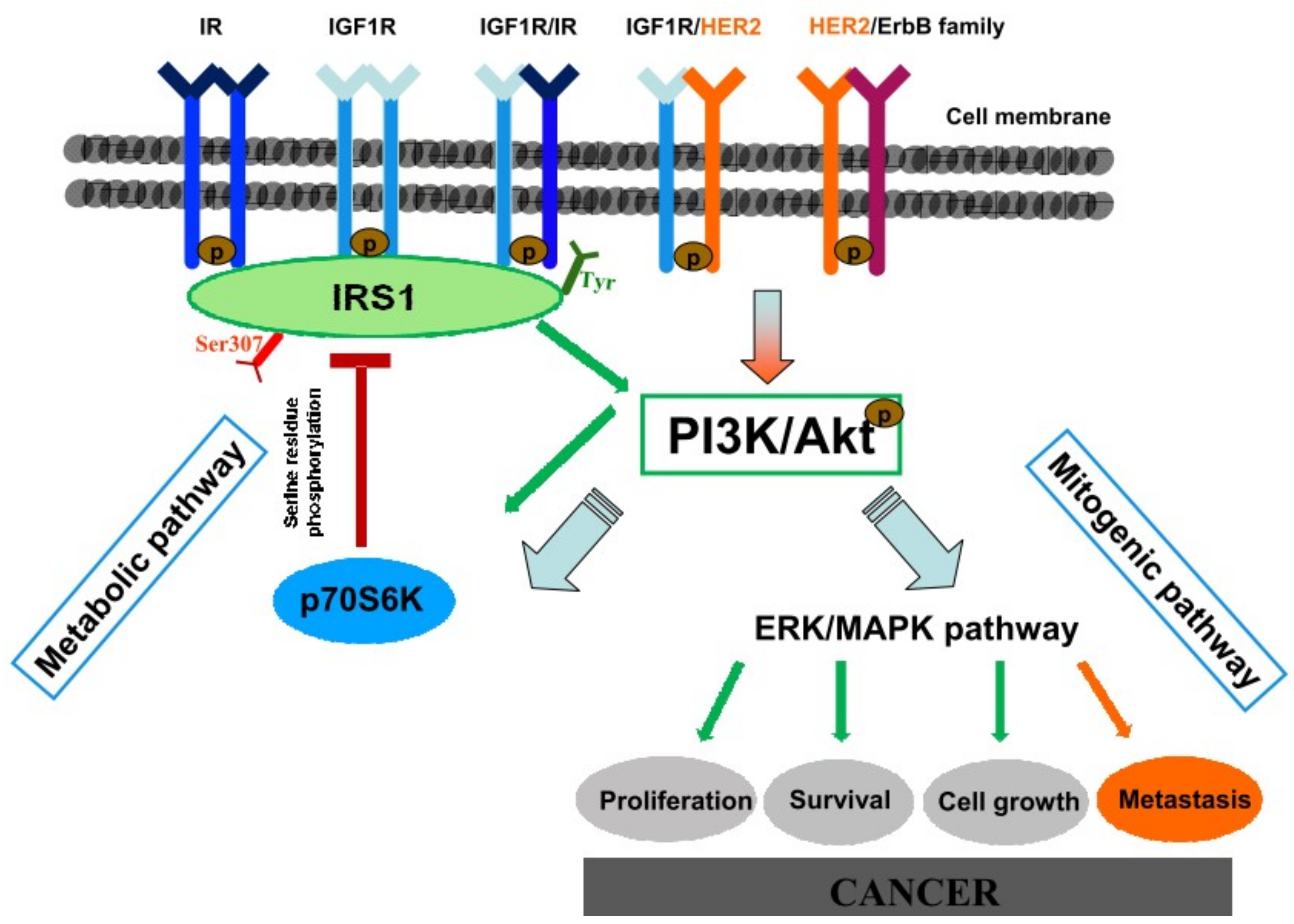
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Surgical Procedure
4.3. Tissues Collection
4.4. Histological and Immunohistochemical Analysis
4.5. Serum Analysis
4.6. Esophageal Tissue Analysis
4.7. Immunoblotting
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| EAC | Esophageal Adenocarcinoma |
| ERK | Extracellular signal-Regulated Kinase |
| ESCC | Esophageal Squamous Cell Carcinoma |
| GERD | Gastro-Esophageal Reflux Disease |
| HER2 | Human Epidermal growth factor receptor 2 |
| IGF1R | Insulin-like growth factor 1 receptor |
| IL-6 | Interleukin 6 |
| IR | Insulin receptor |
| MAPK | Mitogen-Activated Protein Kinase |
| pS | Phosphorylated serine residue |
| pY | Phosphorylated tyrosine residue |
| Thr | Threonine residue |
| Tyr | Tyrosine residue |
References
- Edgren, G.; Adami, H.O.; Weiderpass, E.; Nyrén, O. A global assessment of the oesophageal adenocarcinoma epidemic. Gut 2013, 62, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Hvid-Jensen, F.; Pedersen, L.; Drewes, A.M.; Sørensen, H.T.; Funch-Jensen, P. Incidence of adenocarcinoma among patients with Barrett’s esophagus. N. Engl. J. Med. 2011, 365, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, T.; Ohta, T.; Fujimura, T.; Ninomiya, I.; Fushida, S.; Hattori, T.; Miwa, K. Duodenal juice stimulates oesophageal stem cells to induce Barrett’s oesophagus and oesophageal adenocarcinoma in rats. Oncol. Rep. 2006, 15, 1469–1475. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Singh, S.; Sharma, A.N.; Murad, M.H.; Buttar, N.S.; El-Serag, H.B.; Katzka, D.A.; Iyer, P.G. Central adiposity is associated with increased risk of esophageal inflammation, metaplasia, and adenocarcinoma: A systematic review and meta-analysis. Clin. Gastroenterol. Hepatol. 2013, 11, 1399–1412. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [PubMed]
- Fierz, Y.; Novosyadlyy, R.; Vijayakumar, A.; Yakar, S.; LeRoith, D. Insulin-Sensitizing Therapy Attenuates Type 2 Diabetes–Mediated Mammary Tumor Progression. Diabetes 2010, 59, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Teng, J.A.; Wu, S.G.; Chen, J.X.; Li, Q.; Peng, F.; Zhu, Z.; Qin, J.; He, Z.Y. The Activation of ERK1/2 and JNK MAPK Signaling by Insulin/IGF-1 Is Responsible for the Development of Colon Cancer with Type 2 Diabetes Mellitus. PLoS ONE 2016, 11, e0149822. [Google Scholar] [CrossRef] [PubMed]
- Novosyadlyy, R.; Lann, D.E.; Vijayakumar, A.; Rowzee, A.; Lazzarino, D.A.; Fierz, Y.; Carboni, J.M.; Gottardis, M.M.; Pennisi, P.A.; Molinolo, A.A.; et al. Insulin-mediated acceleration of breast cancer development and progression in a non-obese model of type 2 diabetes. Cancer Res. 2010, 70, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Ning, Z.; Yuanyuan, Z.; Hang, Y.G.; Jian, H.; Xiaodong, C. Metformin and lung cancer risk of patients with type 2 diabetes mellitus: A meta-analysis. Biomed. Rep. 2015, 3, 235–241. [Google Scholar] [CrossRef][Green Version]
- Di Sebastiano, K.M.; Pinthus, J.H.; Duivenvoorden, W.C.M.; Mourtzakis, M. Glucose impairments and insulin resistance in prostate cancer: The role of obesity, nutrition and exercise. Obes. Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Esteva, F.J. Cross-talk between the ErbB/HER family and the type I insulin-like growth factor receptor signaling pathway in breast cancer. J. Mammary Gland Biol. Neoplasia 2008, 13, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Christopoulos, P.F.; Corthay, A.; Koutsilieris, M. Aiming for the Insulin-like Growth Factor-1 system in breast cancer therapeutics. Cancer Treat. Rev. 2018, 63, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Denduluri, S.K.; Idowu, O.; Wang, Z.; Liao, Z.; Yan, Z.; Mohammed, M.K.; Ye, J.; Wei, Q.; Wang, J.; Zhao, L.; et al. Insulin-like growth factor (IGF) signaling in tumorigenesis and the development of cancer drug resistance. Genes Dis. 2015, 2, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, A.; Allister, E.M.; Wheeler, M.B. MK-626, a dipeptidyl peptidase-4 inhibitor, does not improve the hyperglycemia or hyperinsulinemia of nonobese diabetic MKR mice. Can. J. Physiol. Pharmacol. 2012, 90, 663–668. [Google Scholar] [CrossRef] [PubMed]
- LeRoith, D.; Werner, H.; Beitner-Johnson, D.; Roberts, C.T., Jr. Molecular and cellular aspects of the insulin-like growth factor I receptor. Endocr. Rev. 1995, 16, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Zick, Y. Ser/Thr phosphorylation of IRS proteins: A molecular basis for insulin resistance. Sci. STKE 2005, 2005, pe4. [Google Scholar] [CrossRef] [PubMed]
- Zick, Y. Insulin resistance: A phosphorylation-based uncoupling of insulin signaling. Trends Cell Biol. 2001, 11, 437–441. [Google Scholar] [CrossRef]
- Arcidiacono, D.; Antonello, A.; Fassan, M.; Nucci, D.; Morbin, T.; Agostini, M.; Nitti, D.; Rugge, M.; Alberti, A.; Battaglia, G.; et al. Insulin promotes HER2 signaling activation during Barrett’s Esophagus carcinogenesis. Dig. Liver Dis. 2017, 49, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Cobleigh, M.A.; Vogel, C.L.; Tripathy, D.; Robert, N.J.; Scholl, S.; Fehrenbacher, L.; Wolter, J.M.; Paton, V.; Shak, S.; Lieberman, G.; et al. Multinational study of the efficacy and safety of humanized anti-HER2 monoclonal antibody in women who have HER2-overexpressing metastatic breast cancer that has progressed after chemotherapy for metastatic disease. J. Clin. Oncol. 1999, 17, 2639–2648. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.Z.; Chen, Y.; Wang, J.J. Overexpression of Grb2/HER2 signaling in Chinese gastric cancer: Their relationship with clinicopathological parameters and prognostic significance. J. Cancer Res. Clin. Oncol. 2009, 135, 1331–1339. [Google Scholar] [CrossRef] [PubMed]
- Scheel, A.H.; Penault-Llorca, F.; Hanna, W.; Baretton, G.; Middel, P.; Burchhardt, J.; Hofmann, M.; Jasani, B.; Rüschoff, J. Physical basis of the ‘magnification rule’ for standardized Immunohistochemical scoring of HER2 in breast and gastric cancer. Diagn. Pathol. 2018, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Gowryshankar, A.; Nagaraja, V.; Eslick, G.D. HER2 status in Barrett’s esophagus & esophageal cancer: A meta analysis. J. Gastrointest. Oncol. 2014, 5, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, V.; Shaw, N.; Morey, A.L.; Cox, M.R.; Eslick, G.D. HER2 expression in oesophageal carcinoma and Barrett’s oesophagus associated adenocarcinoma: An Australian study. Eur. J. Surg. Oncol. 2016, 42, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Estimating the world cancer burden: Globocan 2000. Int. J. Cancer 2001, 94, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.H.; Mukaisho, K.; Ling, Z.Q.; Shimomura, A.; Sugihara, H.; Hattori, T. Association between duodenal contents reflux and squamous cell carcinoma—Establishment of an esophageal cancer cell line derived from the metastatic tumor in a rat reflux model. Anticancer Res. 2007, 27, 175–181. [Google Scholar] [PubMed]
- Nozoe, T.; Korenaga, D.; Kabashima, A.; Sugimachi, K. Smoking-related increase of O(6)-methylguanine-DNA methyltransferase expression in squamous cell carcinoma of the esophagus. Cancer Lett. 2002, 184, 49–55. [Google Scholar] [CrossRef]
- Sami, S.S.; Iyer, P.G. Recent Advances in Screening for Barrett’s Esophagus. Curr. Treat. Options Gastroenterol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Dulai, G.S.; Guha, S.; Kahn, K.L.; Gornbein, J.; Weinstein, W.M. Preoperative prevalence of Barrett’s esophagus in esophageal adenocarcinoma: A systematic review. Gastroenterology 2002, 122, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Runge, T.M.; Abrams, J.A.; Shaheen, N.J. Epidemiology of Barrett’s Esophagus and Esophageal Adenocarcinoma. Gastroenterol. Clin. N. Am. 2015, 44, 203–231. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Juergens, S.; Zhang, X.; Rezaei, D.; Yu, C.; Strauch, E.D.; Wang, J.Y.; Cheng, E.; Meyer, F.; Wang, D.H.; et al. Deoxycholic acid causes DNA damage while inducing apoptotic resistance through NF-κB activation in benign Barrett’s epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G278–G286. [Google Scholar] [CrossRef] [PubMed]
- Miwa, K.; Sahara, H.; Segawa, M.; Kinami, S.; Sato, T.; Miyazaki, I.; Hattori, T. Reflux of duodenal or gastro-duodenal contents induces esophageal carcinoma in rats. Int. J. Cancer 1996, 67, 269–274. [Google Scholar] [CrossRef]
- Kumagai, H.; Mukaisho, K.; Sugihara, H.; Bamba, M.; Miyashita, T.; Miwa, K.; Hattori, T. Cell kinetic study on histogenesis of Barrett’s esophagus using rat reflux model. Scand. J. Gastroenterol. 2003, 38, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.H. From reflux esophagitis to Barrett’s esophagus and esophageal adenocarcinoma. World J. Gastroenterol. 2015, 21, 5210–5219. [Google Scholar] [CrossRef] [PubMed]
- Le May, C.; Chu, K.; Hu, M.; Ortega, C.S.; Simpson, E.R.; Korach, K.S.; Tsai, M.J.; Mauvais-Jarvis, F. Estrogens protect pancreatic beta-cells from apoptosis and prevent insulin-deficient diabetes mellitus in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 9232–9237. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.; Liu, J.P.; Robertson, E.J.; Efstratiadis, A. Role of insulin-like growth factors in embryonic and postnatal growth. Cell 1993, 75, 73–82. [Google Scholar] [CrossRef]
- Gallagher, E.J.; LeRoith, D. Minireview: IGF, Insulin, and Cancer. Endocrinology 2011, 152, 2546–2551. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, D.R. Metabolic actions of insulin-like growth factor-I in normal physiology and diabetes. Endocrinol. Metab. Clin. N. Am. 2012, 41, 425–443. [Google Scholar] [CrossRef] [PubMed]
- Boucher, J.; Tseng, Y.H.; Kahn, C.R. Insulin and insulin-like growth factor-1 receptors act as ligand-specific amplitude modulators of a common pathway regulating gene transcription. J. Biol. Chem. 2010, 285, 17235–17245. [Google Scholar] [CrossRef] [PubMed]
- Baserga, R.; Peruzzi, F.; Reiss, K. The IGF-1 receptor in cancer biology. Int. J. Cancer 2003, 107, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Guerard, M.; Robin, T.; Perron, P.; Hatat, A.S.; David-Boudet, L.; Vanwonterghem, L.; Busser, B.; Coll, J.L.; Lantuejoul, S.; Eymin, B.; et al. Nuclear translocation of IGF1R by intracellular amphiregulin contributes to the resistance of lung tumour cells to EGFR-TKI. Cancer Lett. 2018, 420, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Schnarr, B.; Strunz, K.; Ohsam, J.; Benner, A.; Wacker, J.; Mayer, D. Down-regulation of insulin-like growth factor-I receptor and insulin receptor substrate-1 expression in advanced human breast cancer. Int. J. Cancer 2000, 89, 506–513. [Google Scholar] [CrossRef]
- Kuklinski, A.; Kamocki, Z.; Koda, M.; Piotrowski, Z.; Sulkowski, S.; Lesniewicz, R.; Pawlak, K.; Mysliwiec, P.; Kedra, B. IGF-IR in patients with advanced colorectal cancer in correlation with certain clinico-morphological factors: Initial report. Oncol. Lett. 2011, 2, 1155–1159. [Google Scholar] [CrossRef] [PubMed]
- Realdon, S.; Dassie, E.; Fassan, M.; Dall’Olmo, L.; Hatem, G.; Buda, A.; Arcidiacono, D.; Diamantis, G.; Zhang, H.; Greene, M.I.; et al. In vivo molecular imaging of HER2 expression in a rat model of Barrett’s esophagus adenocarcinoma. Dis. Esophagus 2015, 28, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Dassie, E.; Arcidiacono, D.; Wasiak, I.; Damiano, N.; Dall’Olmo, L.; Giacometti, C.; Facchin, S.; Cassaro, M.; Guido, E.; De Lazzari, F.; et al. Detection of fluorescent organic nanoparticles by confocal laser endomicroscopy in a rat model of Barrett’s esophageal adenocarcinoma. Int. J. Nanomed. 2015, 10, 6811–6823. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Lesion | |||
|---|---|---|---|---|
| No Lesion | Esophagitis | Dysplasia | Cancer | |
| WT (n = 38) | 2 (5.3) | 0 (0.0) | 24 (63.1) | 12 (31.6) |
| MKR (n = 35) | 1 (2.9) | 4 (11.4) | 4 (11.4) | 26 (74.3) |
| Group | Gender | No Lesion | Esophagitis | LGD | HGD | ESCC | EASC |
|---|---|---|---|---|---|---|---|
| WT (n = 38) | FEMALE (n = 20) | 1 (5.0) | 0 (0.0) | 11 (55.0) | 2 (10.0) | 4 (20.0) | 2 (10.0) |
| MALE (n = 18) | 1 (5.5) | 0 (0.0) | 5 (27.8) | 6 (33.4) | 1 (5.5) | 5 (27.8) | |
| MKR (n = 35) | FEMALE (n = 18) | 0 (0.0) | 4 (22.2) | 2 (11.1) | 2 (11.1) | 6 (33.4) | 4 (22.2) |
| MALE (n = 17) | 1 (5.9) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 7 (41.2) | 9 (52.9) |
| WT Females | Non-Operated n = 8 | Dysplasia n = 13 | Cancer n = 6 | K–W Test |
| Glucose (mg/dL) | 177.5 (147.3; 192.3) | 196.0 (125.0; 254.0) | 235.5 (225.3; 291.5) | 0.179 |
| Insulin (pg/mL) | 423.4 (374.2; 462.2) | 376.5 (360.0; 390.0) | 443.5 (420.0; 741.8) | 0.215 |
| C-Peptide (pg/mL) | 323.0 (295.2; 423.9) | 313.3 (240.1; 405.8) | 439.0 (346.9; 522.5) | 0.334 |
| Leptin (pg/mL) | 2345.8 (1998.7; 2912.6) | 1928.0 (1515.7; 2540.0) | 1784.5 (1640.1; 1962.0) | 0.103 |
| IL-6 (pg/mL) | 23.2 (3.1; 31.9) | 29.0 (1.4; 55.2) | 59.8 (21.8; 74.3) | 0.292 |
| WT Males | Non-Operated n = 8 | Dysplasia n = 11 | Cancer n = 6 | K–W Test |
| Glucose (mg/dL) | 191.0 (166.3; 208.3) | 209.0 (175.3; 252.8) | 222.0 (157.3; 242.5) | 0.469 |
| Insulin (pg/mL) | 838.7 (689.6; 860.4) | 784.0 (467.5; 973.1) | 1406.4 (1033.8; 1912.6) | 0.094 |
| C-Peptide (pg/mL) | 761.7 (407.5; 922.5) | 661.2 (334.9; 755.4) | 850.2 (814.3; 1060.2) | 0.210 |
| Leptin (pg/mL) | 4902.2 (3801.4; 6749.4) | 1564.8 (1336.5; 2811.2) | 1104.1 (894.0; 1415.5) | 0.011 |
| IL-6 (pg/mL) | 24.9 (13.4; 28.3) | 29.0 (11.1; 40.7) | 65.8 (37.4; 162.7) | 0.121 |
| MKR Females | Non-Operated n = 8 | Dysplasia n = 4 | Cancer n = 10 | K–W Test |
| Glucose (mg/dL) | 188.0 (169.0; 206.8) | 258.0 (220.3; 302.0) | 219.5 (201.3; 241.3) | 0.102 |
| Insulin (pg/mL) | 2713.9 (2266.0; 5179.1) | 4884.8 (4595.4; 5084.0) | 3160.0 (2160.8; 3517.8) | 0.055 |
| C-Peptide (pg/mL) | 1498.9 (1182.9;1729.7) | 1711.2 (1496.1;1926.5) | 1534.4 (1074.7;2560.9) | 0.919 |
| Leptin (pg/mL) | 1548.9 (968.1;1975.2) | 1323.0 (910.8;1565.1) | 921.1 (842.0;1123.9) | 0.192 |
| IL-6 (pg/mL) | 20.5 (14.3;29.6) | 28.7 (17.2;36.8) | 40.5 (11.2;57.8) | 0.390 |
| MKR Males | Non-Operated n = 8 | Cancer n = 16 | M–W Test | |
| Glucose (mg/dL) | 207.5 (189.3; 237.0) | 171.0 (151.3; 199.5) | 0.037 | |
| Insulin (pg/mL) | 10780.8 (8810.5; 12018.1) | 8043.4 (5268.8; 10373.4) | 0.126 | |
| C-Peptide (pg/mL) | 5238.0 (4263.1; 5635.3) | 2168.2 (1383.7; 3413.6) | 0.043 | |
| Leptin (pg/mL) | 1344.1 (1182.7; 1524.9) | 808.2 (640.4; 1077.6) | 0.014 | |
| IL-6 (pg/mL) | 17.7 (12.1; 23.6) | 73.6 (54.8; 152.7) | 0.009 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arcidiacono, D.; Dedja, A.; Giacometti, C.; Fassan, M.; Nucci, D.; Francia, S.; Fabris, F.; Zaramella, A.; Gallagher, E.J.; Cassaro, M.; et al. Hyperinsulinemia Promotes Esophageal Cancer Development in a Surgically-Induced Duodeno-Esophageal Reflux Murine Model. Int. J. Mol. Sci. 2018, 19, 1198. https://doi.org/10.3390/ijms19041198
Arcidiacono D, Dedja A, Giacometti C, Fassan M, Nucci D, Francia S, Fabris F, Zaramella A, Gallagher EJ, Cassaro M, et al. Hyperinsulinemia Promotes Esophageal Cancer Development in a Surgically-Induced Duodeno-Esophageal Reflux Murine Model. International Journal of Molecular Sciences. 2018; 19(4):1198. https://doi.org/10.3390/ijms19041198
Chicago/Turabian StyleArcidiacono, Diletta, Arben Dedja, Cinzia Giacometti, Matteo Fassan, Daniele Nucci, Simona Francia, Federico Fabris, Alice Zaramella, Emily J. Gallagher, Mauro Cassaro, and et al. 2018. "Hyperinsulinemia Promotes Esophageal Cancer Development in a Surgically-Induced Duodeno-Esophageal Reflux Murine Model" International Journal of Molecular Sciences 19, no. 4: 1198. https://doi.org/10.3390/ijms19041198
APA StyleArcidiacono, D., Dedja, A., Giacometti, C., Fassan, M., Nucci, D., Francia, S., Fabris, F., Zaramella, A., Gallagher, E. J., Cassaro, M., Rugge, M., LeRoith, D., Alberti, A., & Realdon, S. (2018). Hyperinsulinemia Promotes Esophageal Cancer Development in a Surgically-Induced Duodeno-Esophageal Reflux Murine Model. International Journal of Molecular Sciences, 19(4), 1198. https://doi.org/10.3390/ijms19041198