Global and Complement Gene-Specific DNA Methylation in Grass Carp after Grass Carp Reovirus (GCRV) Infection

 ,
,
Abstract
:1. Introduction
2. Results
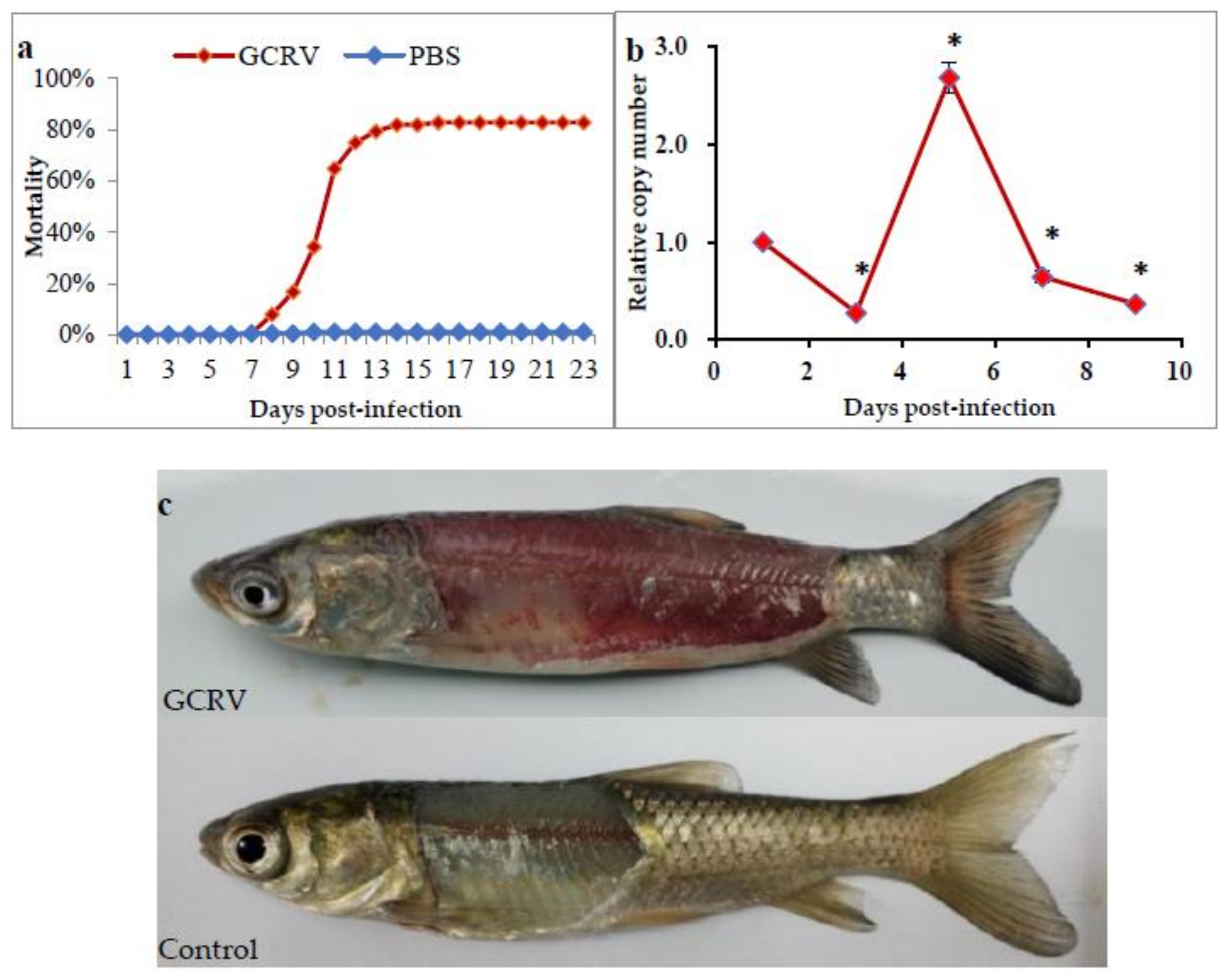
2.1. Grass Carp after GCRV Infection
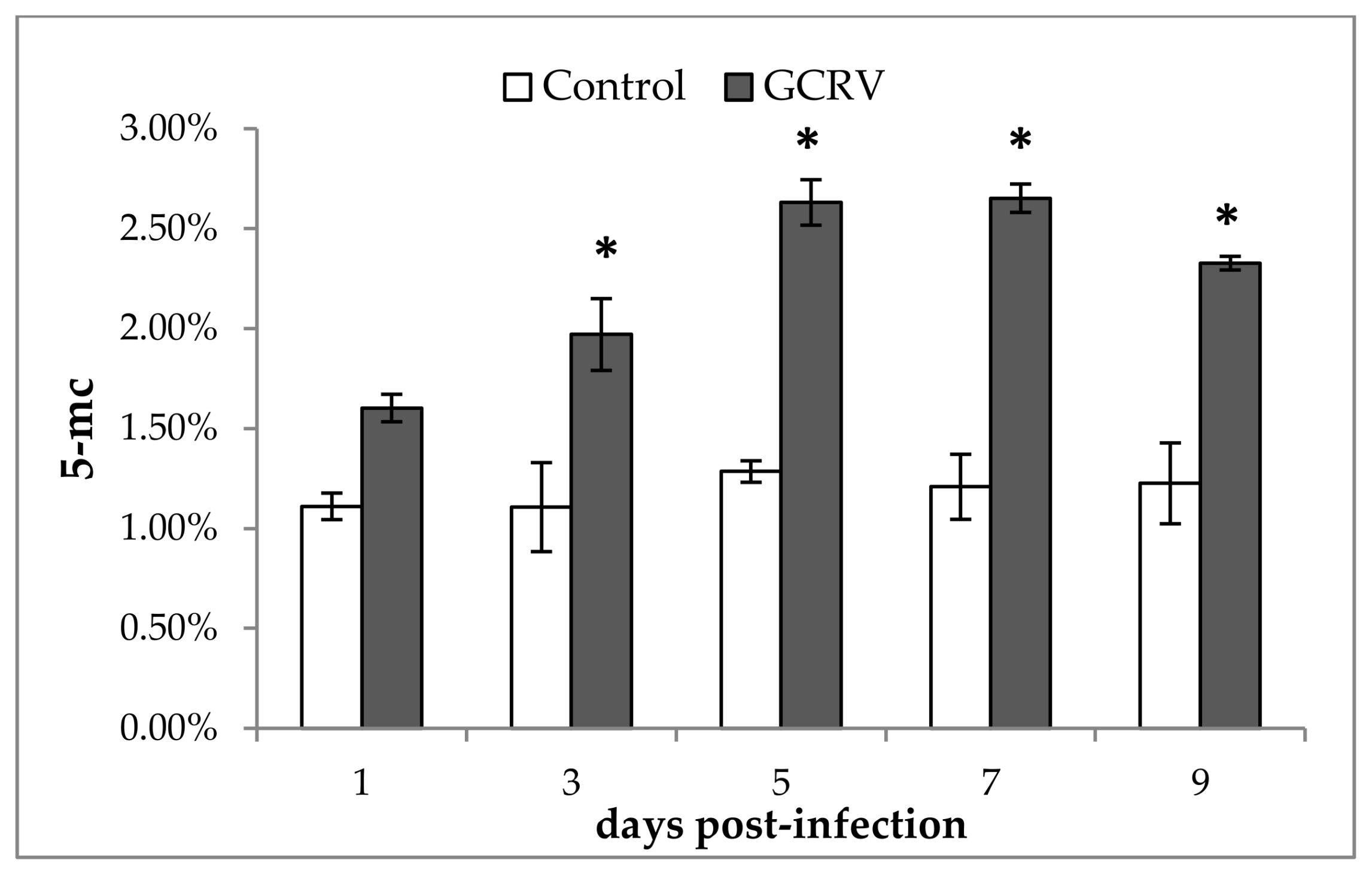
2.2. Global DNA Methylation Changes after GCRV Infection
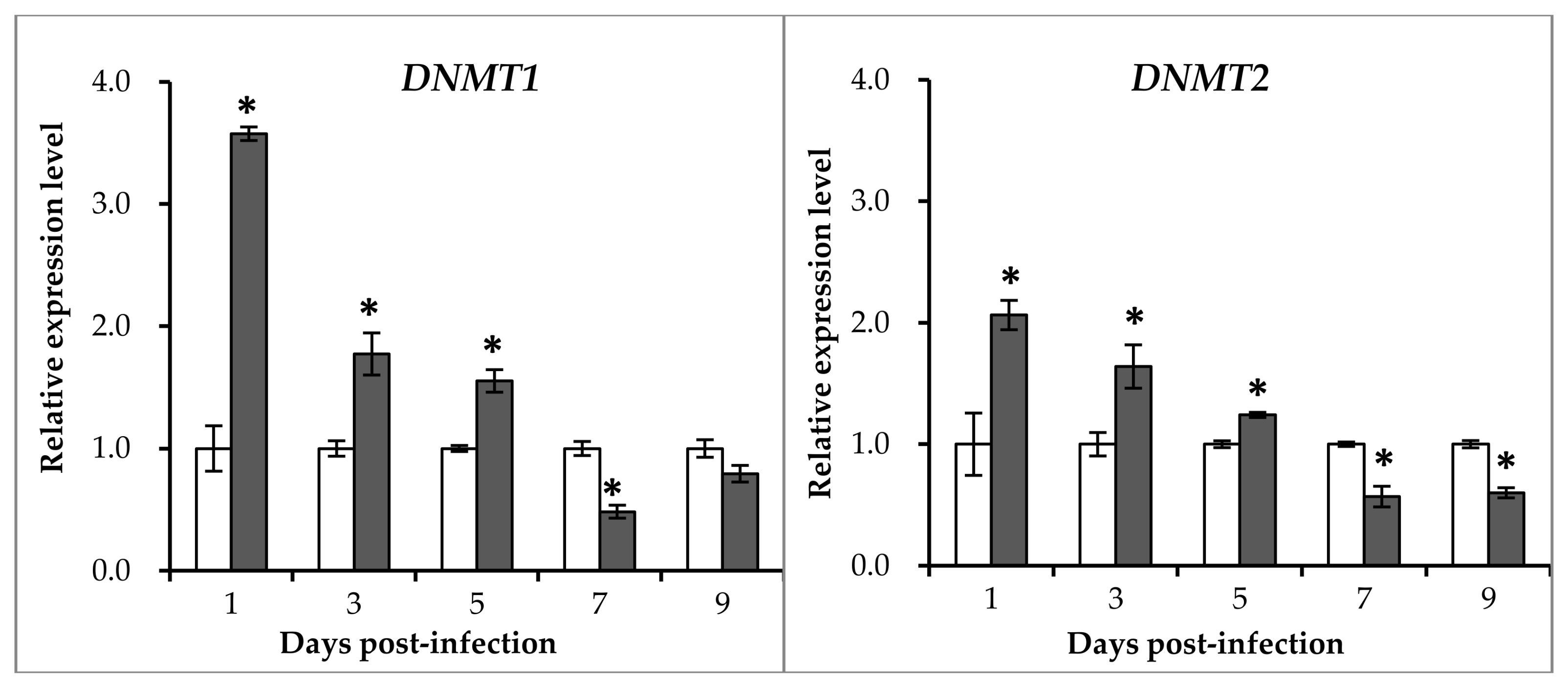
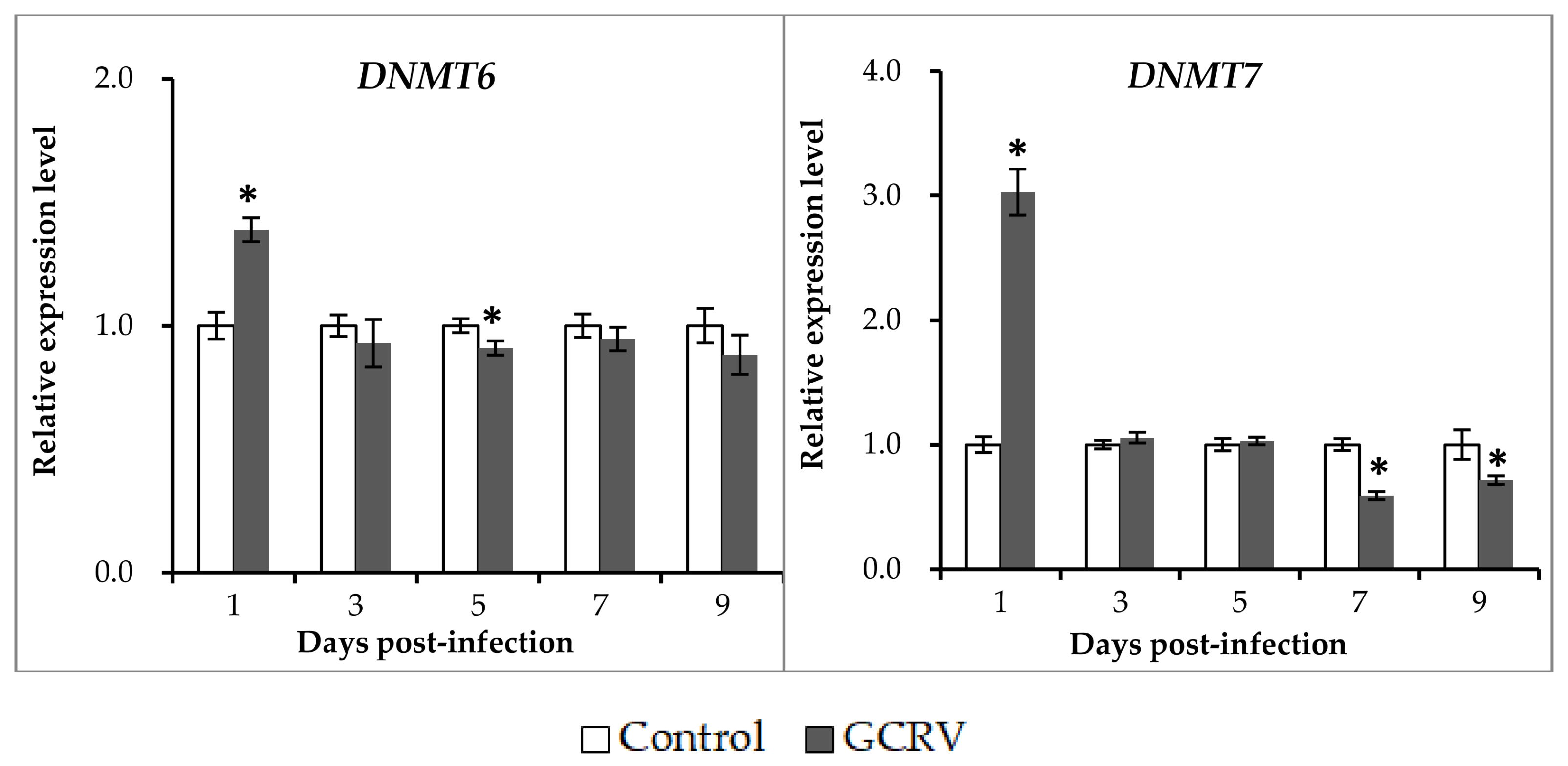
2.3. Expression Patterns of DNA Methyltransferase Genes after GCRV Infection
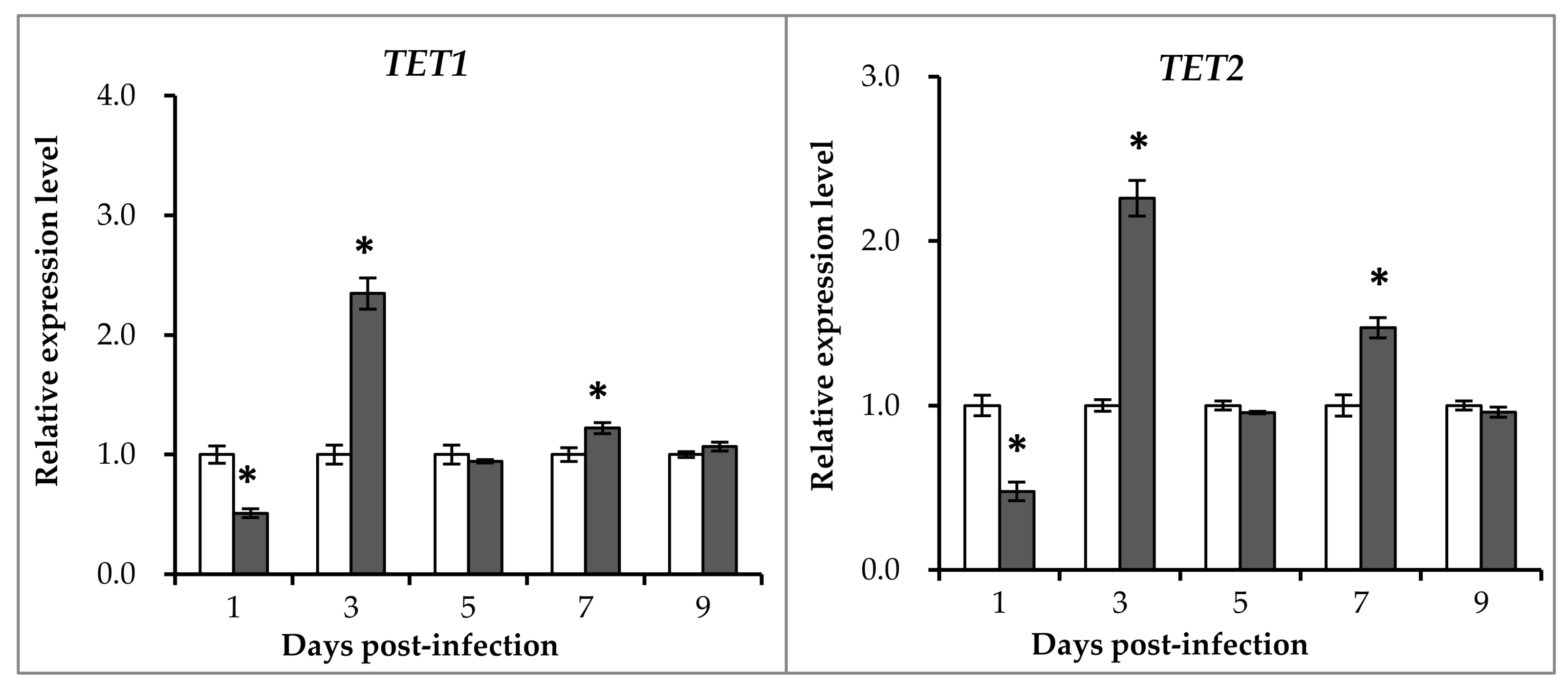
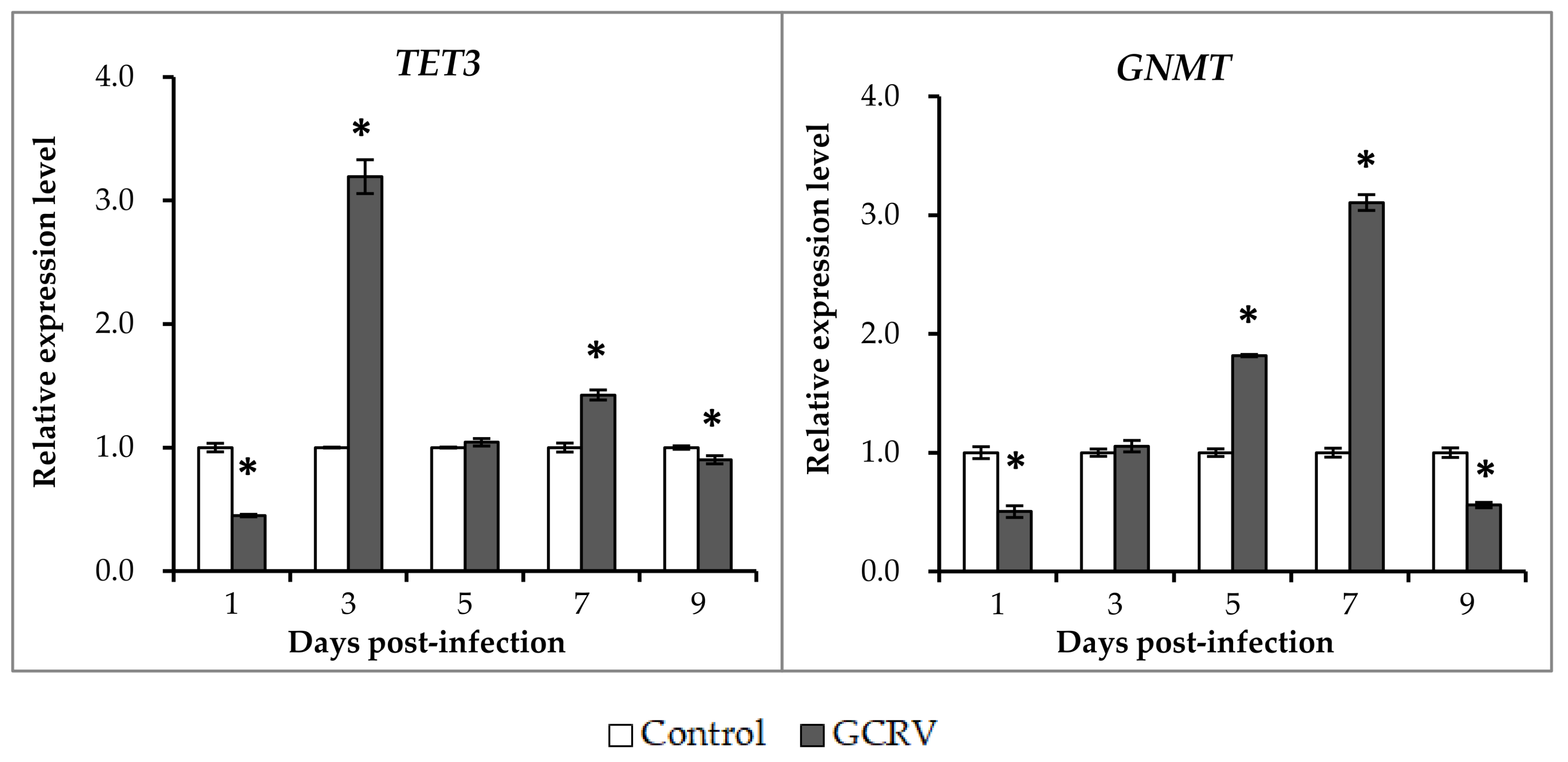
2.4. Expression Patterns of Ten-Eleven Translocation Proteins and Glycine N-methyltransferase after GCRV Exposure
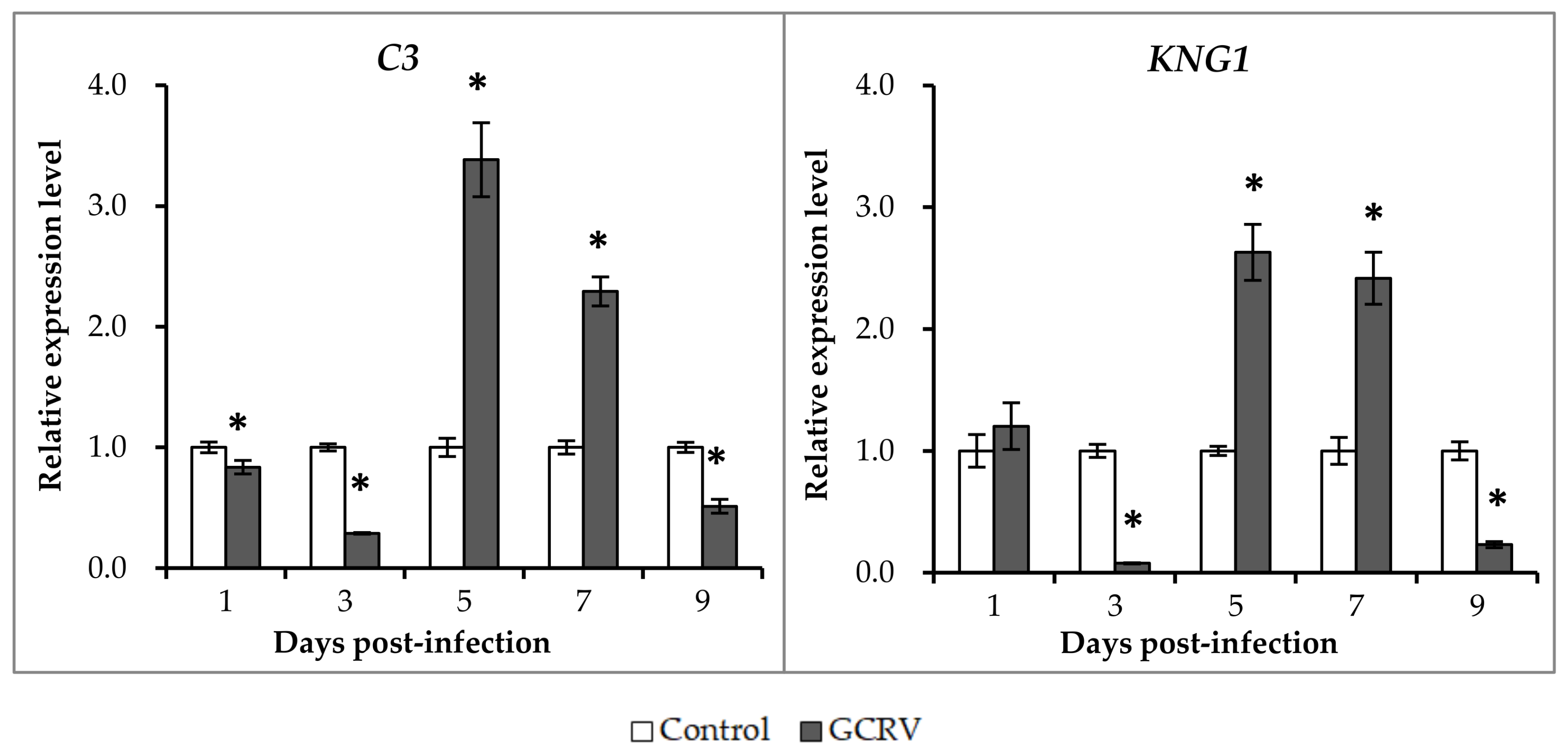
2.5. Expression Patterns of Representative Complement Genes after GCRV Infection
2.6. Methylation Changes in the 5′ flanking Region of C3 and KNG1 after GCRV Infection
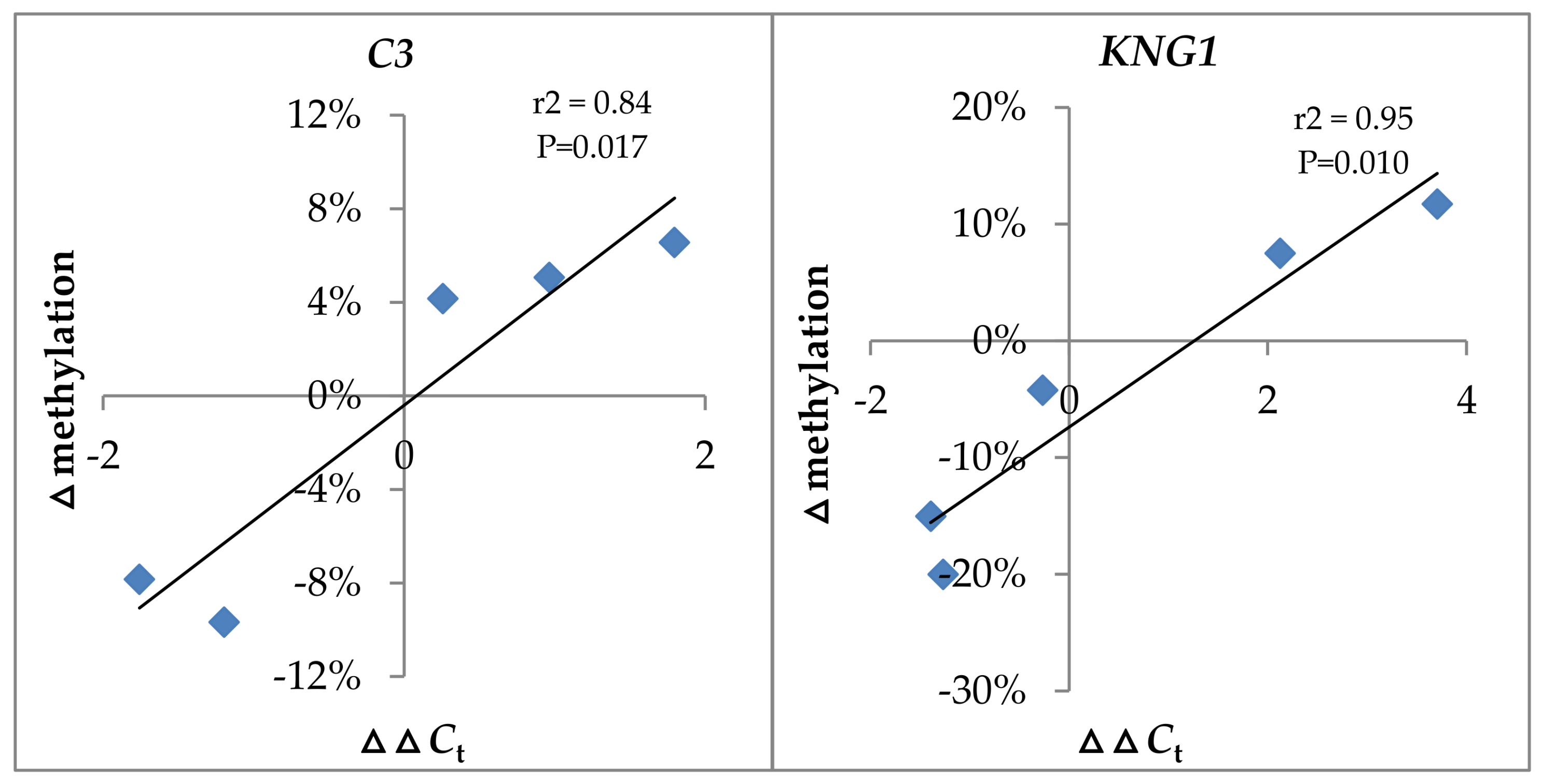
2.7. The Relationship between Expression Level and DNA Methylation Level of C3 and KNG1 after GCRV Infection
3. Discussion
4. Materials and Methods
4.1. Ethical Procedures
4.2. Experimental Animals, Virus Exposure, and Sample Collection
4.3. DNA Isolation and Global DNA Methylation Measurement
4.4. Bisulfite Treatment and Sequencing
4.5. RNA Isolation and Quantitative Real-Time PCR (RT-qPCR)
4.6. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| C3 | complement component 3 |
| CpG | cytosine-phosphor-guanine |
| DNMT | DNA methyltransferase |
| GCRV | grass carp reovirus |
| GNMT | glycine N-methyltransferase |
| KNG1 | kininogen-1 |
| LSD | least significant difference |
| MDV | Marek’s disease virus |
| TETs | ten-eleven translocation proteins |
| TIS | translation initiation site |
| TSS | transcriptional start site |
| SAM | S-adenosyl-l-methionine |
| SPSS | Statistical Product and Service Solutions |
| 5hmC | 5-hydroxymethylcytosine |
| 5-mC | 5-methylcytosine |
References
- Food and Agriculture Organization (FAO). Fishery and Aquaculture Statistics Yearbook; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016. [Google Scholar]
- Wang, Q.; Zeng, W.; Liu, C.; Zhang, C.; Wang, Y.; Shi, C.; Wu, S. Complete genome sequence of reovirus isolated from grass carp, indicating different genotypes of GCRV in China. J. Virol. 2012, 86, 12466. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.; Su, J. Insights into the antiviral immunity against grass carp (Ctenopharyngodon idella) reovirus (GCRV) in grass carp. J. Immunol. Res. 2015, 2015, 670437. [Google Scholar] [CrossRef] [PubMed]
- Varriale, A. DNA methylation, epigenetics, and evolution in vertebrates: Facts and challenges. Int. J. Evol. Biol. 2014, 2014, 475981. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.P. CpG-rich islands and the function of DNA methylation. Nature 1986, 321, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Caiafa, P.; Zampieri, M. DNA methylation and chromatin structure: The puzzling CpG islands. J. Cell. Biochem. 2005, 94, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Hellmann, I.; Stadler, M.B.; Ramos, L.; Pääbo, S.; Rebhan, M.; Schübeler, D. Distribution, silencing potential and evolutionary impact of promoter DNA methylation in the human genome. Nat. Genet. 2007, 39, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Meissner, A.; Mikkelsen, T.S.; Gu, H.; Wernig, M.; Hanna, J.; Sivachenko, A.; Zhang, X.; Bernstein, B.E.; Nusbaum, C.; Jaffe, D.B.; et al. Genome-scale DNA methylation maps of pluripotent and differentiated cells. Nature 2008, 454, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Hinoue, T.; Weisenberger, D.J.; Lange, C.P.; Shen, H.; Byun, H.M.; Van Den Berg, D.; Malik, S.; Pan, F.; Noushmehr, H.; van Dijk, C.M.; et al. Genome-scale analysis of aberrant DNA methylation in colorectal cancer. Genome Res. 2012, 22, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yuan, C.; Chen, S.; Zheng, Y.; Zhang, Y.; Gao, J.; Wang, Z. Global and cyp19a1a gene specific DNA methylation in gonads of adult rare minnow Gobiocypris rarus under bisphenol A exposure. Aquat. Toxicol. 2014, 156, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Wan, Q.; Su, J.; Su, J. DNA methylation of CiRIG-I gene notably relates to the resistance against GCRV and negatively-regulates mRNA expression in grass carp, Ctenopharyngodon idella. Immunobiology 2016, 221, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Su, J.; Wan, Q.; Su, J. CpA/CpG methylation of CiMDA5 possesses tight association with the resistance against GCRV and negatively regulates mRNA expression in grass carp, Ctenopharyngodon idella. Dev. Comp. Immunol. 2015, 48, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; He, F.; Wen, H.; Li, J.; Si, Y.; Liu, M.; Huang, Y.; Meng, L. Low salinity affects cellularity, DNA methylation, and mRNA expression of igf1 in the liver of half smooth tongue sole (Cynoglossus semilaevis). Fish Physiol. Biochem. 2017, 43, 1587–1602. [Google Scholar] [CrossRef] [PubMed]
- Hermann, A.; Gowher, H.; Jeltsch, A. Biochemistry and biology of mammalian DNA methyltransferases. Cell. Mol. Life Sci. 2004, 61, 2571–2587. [Google Scholar] [CrossRef] [PubMed]
- Goll, M.G.; Bestor, T.H. Eukaryotic cytosine methyltransferases. Annu. Rev. Biochem. 2005, 74, 481–514. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.; Nadauld, L.D.; Chidester, S.; Manos, E.J.; James, S.R.; Karpf, A.R.; Cairns, B.R.; Jones, D.A. Zebrafish Dnmt1 and Suv39h1 regulate organ-specific terminal differentiation during development. Mol. Cell. Biol. 2006, 26, 7077–7085. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Pu, M.T.; Hirasawa, R.; Li, B.Z.; Huang, Y.N.; Zeng, R.; Jing, N.H.; Chen, T.; Li, E.; Sasaki, H.; et al. Synergistic function of DNA methyltransferases Dnmt3a and Dnmt3b in the methylation of Oct4 and Nanog. Mol. Cell. Biol. 2007, 27, 8748–8759. [Google Scholar] [CrossRef] [PubMed]
- Campos, C.; Valente, L.M. Fernandes JM. Molecular evolution of zebrafish dnmt3 genes and thermal plasticity of their expression during embryonic development. Gene 2012, 500, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Dong, W.; Thornton, C.; Willett, K.L. Benzo[a]pyrene effects on glycine N-methyltransferase mRNA expression and enzyme activity in Fundulus heteroclitus embryos. Aquat. Toxicol. 2010, 98, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; D’Alessio, A.C.; Taranova, O.V.; Hong, K.; Sowers, L.C.; Zhang, Y. Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification. Nature 2010, 466, 1129–1133. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhang, Y. Mechanisms and functions of Tet protein-mediated 5-methylcytosine oxidation. Genes Dev. 2011, 25, 2436–2452. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Zhang, R.P.; Wan, F.; Guo, D.Y.; Wang, P.; Xiang, L.X.; Shao, J.Z. TET2 plays an essential role in erythropoiesis by regulating lineage-specificgenes via DNA oxidative demethylation in a zebrafish model. Mol. Cell. Biol. 2014, 34, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Fan, Y.D.; Zeng, L.B.; Ma, J. Prokaryotic expression and immunoassay of grass carp reovirus capsid VP6 protein. Acta Virol. 2013, 57, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Jian, J.C.; Wang, Y.; Yan, X.Y.; Ding, Y.; Wu, Z.H.; Lu, Y.S. Molecular cloning and prokaryotic expression of vp5 gene of grass carp reovirus strain GCRV096. Virus Genes 2013, 47, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.L.; Zhang, L.F.; Fang, Z.Z.; Xu, L.P.; Zhang, M.; Wang, N.; Jiang, Y.L.; Lin, X.M. Detection of grass carp reovirus (GCRV) with monoclonal antibodies. Arch. Virol. 2014, 159, 649–655. [Google Scholar]
- Zeng, W.; Wang, Y.; Liang, H.; Liu, C.; Song, X.; Shi, C.; Wu, S.; Wang, Q. A one-step duplex rRT-PCR assay for the simultaneous detection of grass carp reovirus genotypes I and II. J. Virol. Methods 2014, 210C, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Liu, G.L.; Gong, Y.X.; Ling, F.; Wang, G.X. Protective immunity of grass carp immunized with DNA vaccine encoding the vp7 gene of grass carp reovirus using carbon nanotubes as a carrier molecule. Fish Shellfish Immunol. 2015, 42, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Song, L.; Wang, H.; Xu, X.; Wang, T.; Lu, L. Proteomic analysis of cellular protein expression profiles in response to grass carp reovirus infection. Fish Shellfish Immunol. 2015, 44, 515–524. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhang, A.; Pei, Y.; Chu, P.; Li, Y.; Huang, R.; Liao, L.; Zhu, Z.; Wang, Y. Differences in responses of grass carp to different types of grass carp reovirus(GCRV) and the mechanism of hemorrhage revealed by transcriptome sequencing. BMC Genom. 2017, 18, 452. [Google Scholar] [CrossRef] [PubMed]
- Bird, A. Perceptions of epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Egger, G.; Liang, G.; Aparicio, A.; Jones, P.A. Epigenetics in human disease and prospects for epigenetic therapy. Nature 2004, 429, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Serman, A.; Vlahović, M.; Serman, L.; Bulić-Jakus, F. DNA methylation as a regulatory mechanism for gene expression in mammals. Coll. Antropol. 2006, 30, 665–671. [Google Scholar] [PubMed]
- Lyles, D.S. Cytopathogenesis and inhibition of host gene expression by RNA viruses. Microbiol. Mol. Biol. Rev. 2000, 64, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Pulgar, R.; Hödar, C.; Travisany, D.; Zuñiga, A.; Domínguez, C.; Maass, A.; González, M.; Cambiazo, V. Transcriptional response of Atlantic salmon families to Piscirickettsia salmonis infection highlights the relevance of the iron-deprivation defence system. BMC Genom. 2015, 16, 495. [Google Scholar] [CrossRef] [PubMed]
- Gale, M., Jr.; Tan, S.L.; Katze, M.G. Translational control of viral gene expression in eukaryotes. Microbiol. Mol. Biol. Rev. 2000, 64, 239–280. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Shin, Y.S.; Kang, B.W.; Kim, J.G.; Won, K.J.; Lieberman, P.M.; Cho, H.; Kang, H. DNA hypermethylation induced by Epstein-Barr virus in the development of Epstein-Barr virus-associated gastric carcinoma. Arch. Pharm. Res. 2017, 40, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Meng, J.L.; Gao, Y.; Zhang, J.Y.; Niu, S.L.; Yu, X.Z.; Li, Y.B.; Guan, Y.T.; Sun, B.X.; Zhao, Z.H. Changes in methylation of genomic DNA from chicken immune organs in response to H5N1 influenza virus infection. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, V.R.; Jarmasz, J.S.; Murugeshan, N.; Del Bigio, M.R.; Rastegar, M.; Davie, J.R. DNA modifications: Function and applications in normal and disease States. Biology (Basel) 2014, 3, 670–723. [Google Scholar] [CrossRef] [PubMed]
- Takata, Y.; Huang, Y.; Komoto, J.; Yamada, T.; Konishi, K.; Ogawa, H.; Gomi, T.; Fujioka, M.; Takusagawa, F. Catalytic mechanism of glycine N-methyltransferase. Biochemistry 2003, 42, 8394–8402. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Zhan, F.; VanderKraats, N.D.; Hiken, J.F.; Edwards, J.R.; Zhang, H.; Zhao, K.; Song, J. DNMT gene expression and methylome in Marek’s disease resistant and susceptible chickens prior to and following infection by MDV. Epigenetics 2013, 8, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Jiang, Z.; Xia, Y.; Lou, P.; Chen, L.; Wang, H.; Bai, L.; Xie, Y.; Liu, Y.; Li, W.; et al. Genome-wide DNA methylation changes in skeletal muscle between young and middle-aged pigs. BMC Genom. 2014, 15, 653. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Zhang, A.; Chu, P.; Li, Y.; Huang, R.; Liao, L.; Zhu, Z.; Wang, Y. Deep Illumina sequencing reveals conserved and novel microRNAs in grass carp in response to grass carp reovirus infection. BMC Genom. 2017, 18, 195. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.R.; Li, Y.G.; Zeng, W.W.; Wang, Y.Y.; Wang, Q.; Wu, S.Q. Pathogenicity and tissue distribution of grass carp reovirus after intraperitoneal administration. Virol. J. 2014, 11, 178. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, Y.; Zhang, Y.; Ning, Z.; Li, Y.; Zhao, Q.; Lu, H.; Huang, R.; Xia, X.; Feng, Q.; et al. The draft genome of the grass carp (Ctenopharyngodon idellus) provides genomic insights into its evolution and vegetarian diet adaptation. Nat. Genet. 2015, 47, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Li, L.C.; Dahiya, R. MethPrimer: Designing primers for methylation PCRs. Bioinformatics 2002, 18, 27–31. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′ to 3′) | Usage |
|---|---|---|
| GCRV-F | GGCGGCTGCTATGATACCAGTAGAC | GCRV detection |
| GCRV-R | GTCGCTTCAGATCAATCACCAAATC | |
| C3-BSP-F | TTAGAGGGTTATTATGTTGAGTATTT | BSP PCR of C3 |
| C3-BSP-R | AAAACTCATAAAACCATTCAACTC | |
| C3-BSP-outer-R | AACAATATAAATTTTTCAAAAAATTC | |
| KNG1-BSP-F | TGTAAAGGTTTTTGTGAAGGTTTTT | BSP PCR of KNG1 |
| KNG1-BSP-R | CCACTAAACTCTATCCTCCCCTATAATA | |
| KNG1-BSP-outer-R | TCTCTACTCTTCTAAAAACCCAACAA | |
| Qdnmt1-F | CCACCGAAATGTGCCGACTG | QPCR of dnmt1 |
| Qdnmt1-R | GGTGCCCATGCTTGTCATACAC | |
| Qdnmt2-F | AAGTCTCAAAGCCCTTCACATTCTC | QPCR of dnmt2 |
| Qdnmt2-R | ACTACCCCACTCCTGCCAAAAAC | |
| Qdnmt6-F | CAACCGCCACAGCCTCAAACT | QPCR of dnmt6 |
| Qdnmt6-R | CCTCGGCCATCTCGTACTCTGAC | |
| Qdnmt7-F | GAGAAGCAAGTGGGCTCCTACG | QPCR of dnmt7 |
| Qdnmt7-R | CAGCGATGGAGGTCGTAAGAGTT | |
| Qtet1-F | AGGCGTGTGAACATCAGGTGG | QPCR of tet1 |
| Qtet1-R | AACCGAAAAACGGGGCATC | |
| Qtet2-F | CCTAACTCCAAAATAGACGGCAC | QPCR of tet2 |
| Qtet2-R | GATGCGGGGTCATTGGTTTA | |
| Qtet3-F | GCCGCATATCCCTGGTCTTCTA | QPCR of tet3 |
| Qtet3-R | TCGTGTCATCACACCGTCTCGT | |
| Qgnmt-F | AGTCTGAATGCCCTCTGGTGG | QPCR of gnmt |
| Qgnmt-R | ACCAGACTTCAAAGGGACCAAA | |
| QC3-F | ATGGTTCGCAAACACTCCTCAG | QPCR of C3 |
| QC3-R | ACAGGTACTTGGCTTCTATGTCAACT | |
| Qkng1-F | TGAAGGTGCTCAACTGGCTCT | QPCR of kng1 |
| Qkng1-R | AGTAACTCTTGACCTGCTTCTGTAAAC | |
| Q-β-actin-F | AGCCATCCTTCTTGGGTATG | QPCR of β-actin |
| Q-β-actin-R | GGTGGGGCGATGATCTTGAT | |
| Q-GCRV-F | AGCGCAGCAGGCAATTACTATCT | QPCR of GCRV |
| Q-GCRV-R | ATCTGCTGGTAATGCGGAACG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, L.; He, L.; Luo, L.; Li, Y.; Liao, L.; Huang, R.; Zhu, Z.; Wang, Y. Global and Complement Gene-Specific DNA Methylation in Grass Carp after Grass Carp Reovirus (GCRV) Infection. Int. J. Mol. Sci. 2018, 19, 1110. https://doi.org/10.3390/ijms19041110
Xiong L, He L, Luo L, Li Y, Liao L, Huang R, Zhu Z, Wang Y. Global and Complement Gene-Specific DNA Methylation in Grass Carp after Grass Carp Reovirus (GCRV) Infection. International Journal of Molecular Sciences. 2018; 19(4):1110. https://doi.org/10.3390/ijms19041110
Chicago/Turabian StyleXiong, Lv, Libo He, Lifei Luo, Yongming Li, Lanjie Liao, Rong Huang, Zuoyan Zhu, and Yaping Wang. 2018. "Global and Complement Gene-Specific DNA Methylation in Grass Carp after Grass Carp Reovirus (GCRV) Infection" International Journal of Molecular Sciences 19, no. 4: 1110. https://doi.org/10.3390/ijms19041110
APA StyleXiong, L., He, L., Luo, L., Li, Y., Liao, L., Huang, R., Zhu, Z., & Wang, Y. (2018). Global and Complement Gene-Specific DNA Methylation in Grass Carp after Grass Carp Reovirus (GCRV) Infection. International Journal of Molecular Sciences, 19(4), 1110. https://doi.org/10.3390/ijms19041110