ARHGAP15 in Human Breast Carcinoma: A Potent Tumor Suppressor Regulated by Androgens

 ,
,
Abstract
:1. Introduction
2. Results
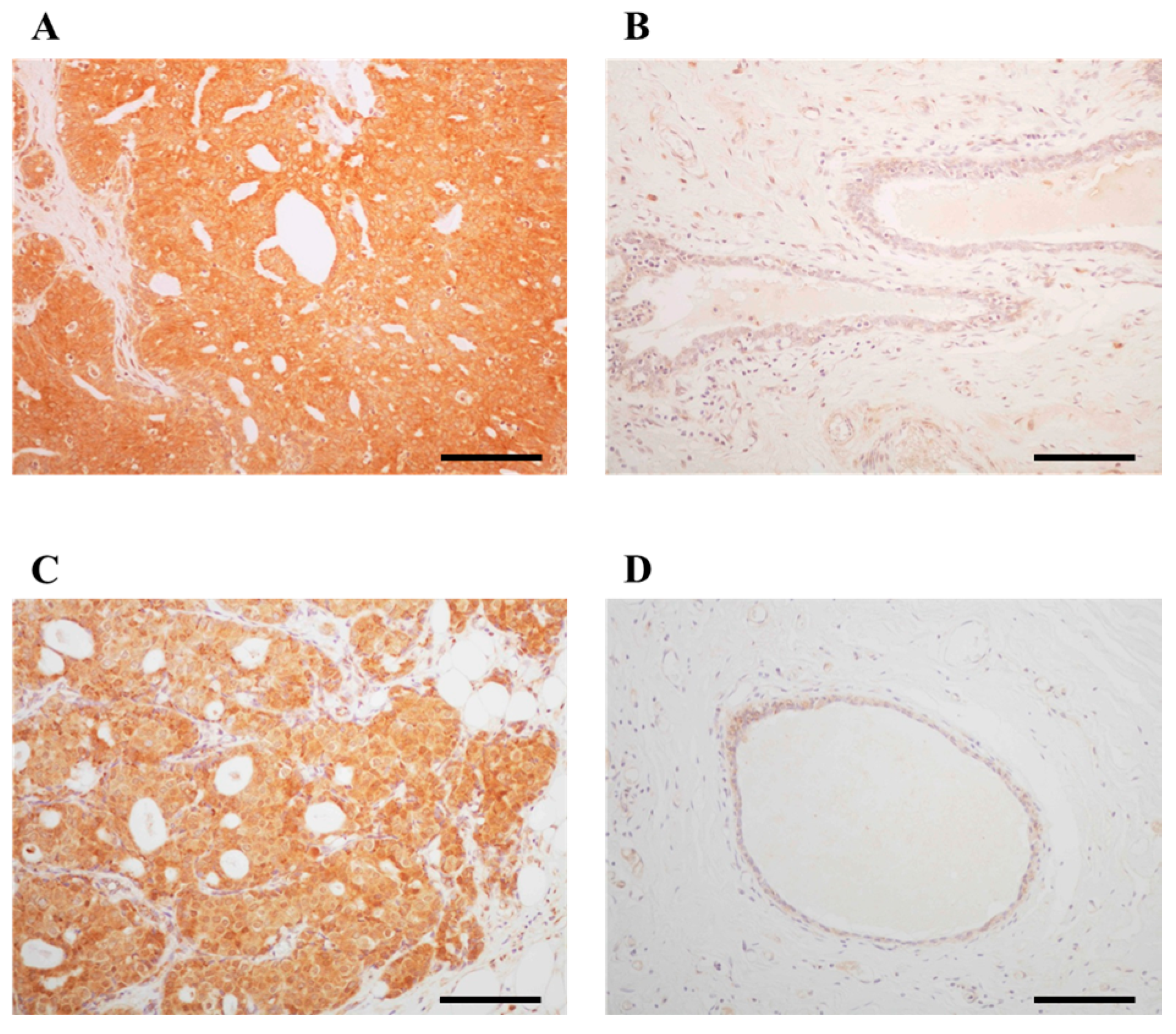
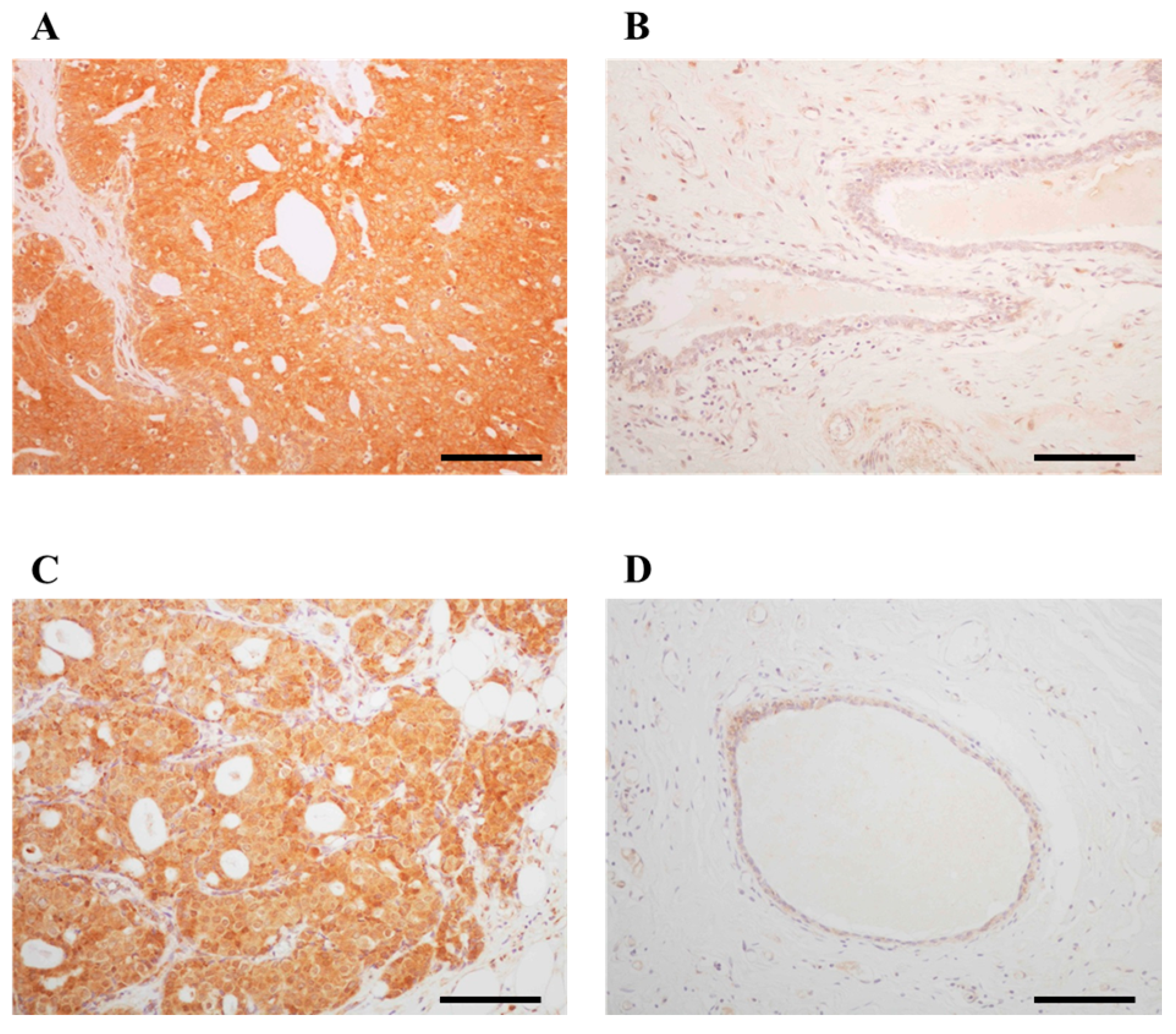
2.1. Immunolocalization of ARHGAP15 and Rac1 in Human Breast Carcinoma Tissues
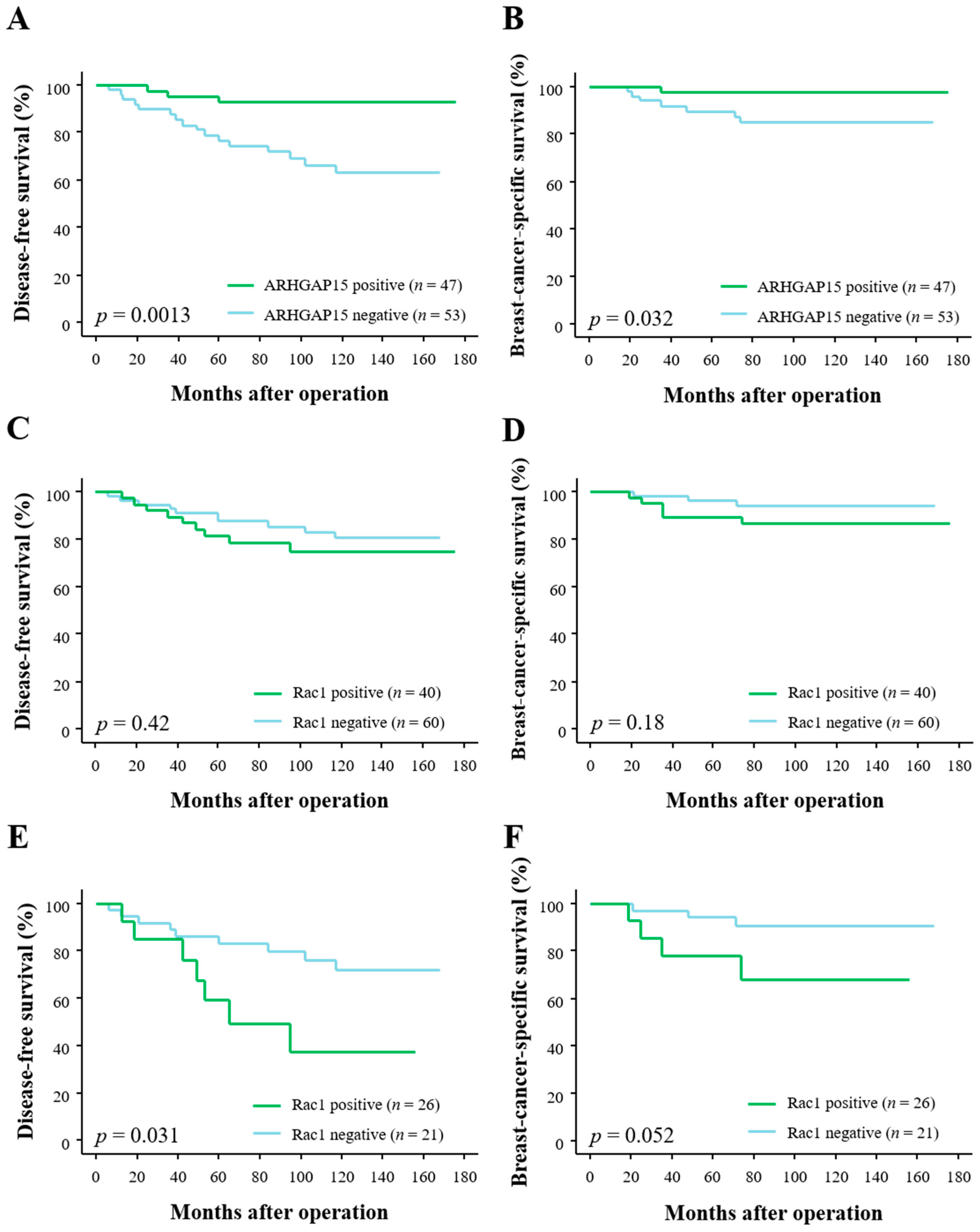
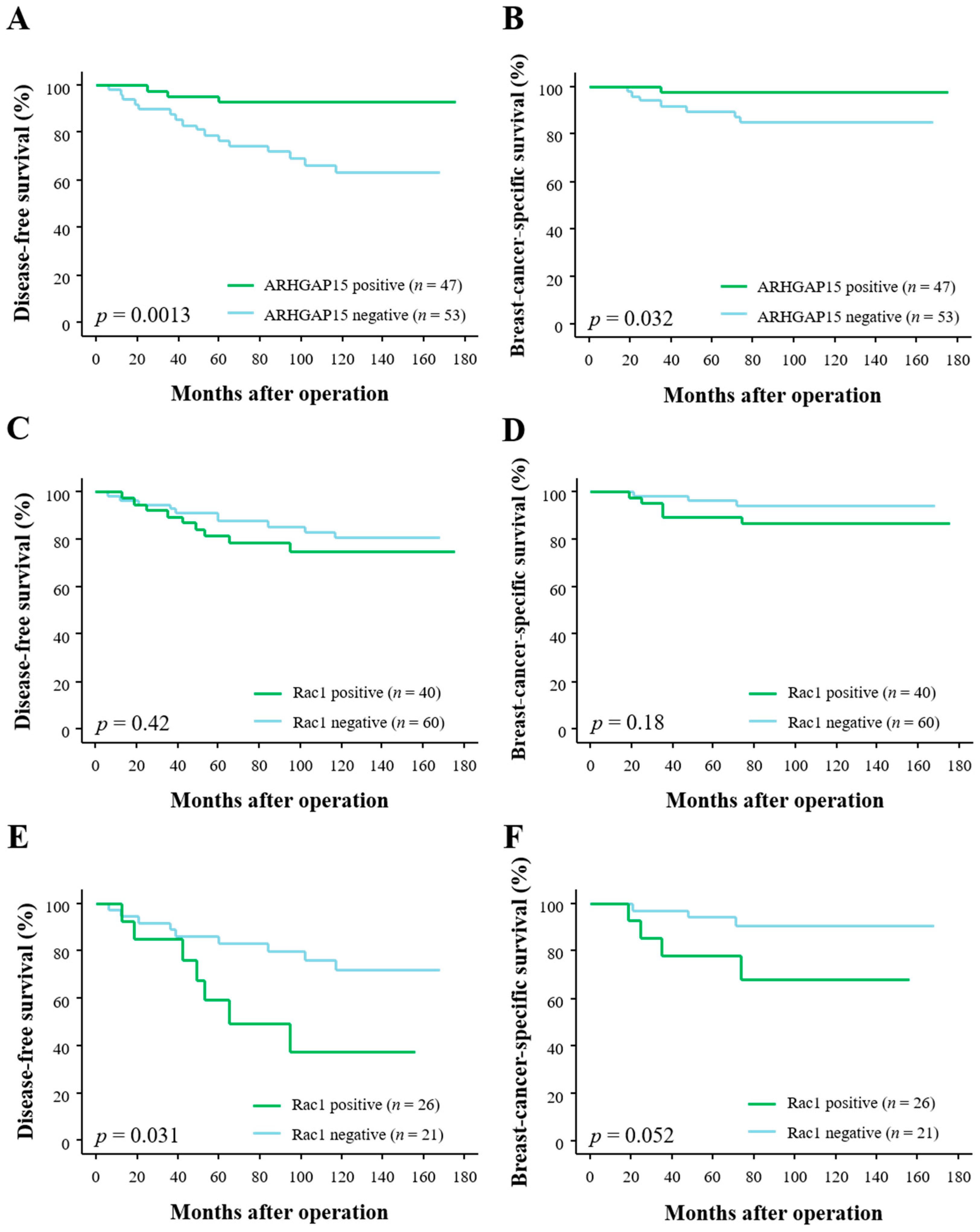
2.2. Correlation between ARHGAP15, Rac1 Immunoreactivity, and Clinical Outcome of Breast Carcinoma Patients
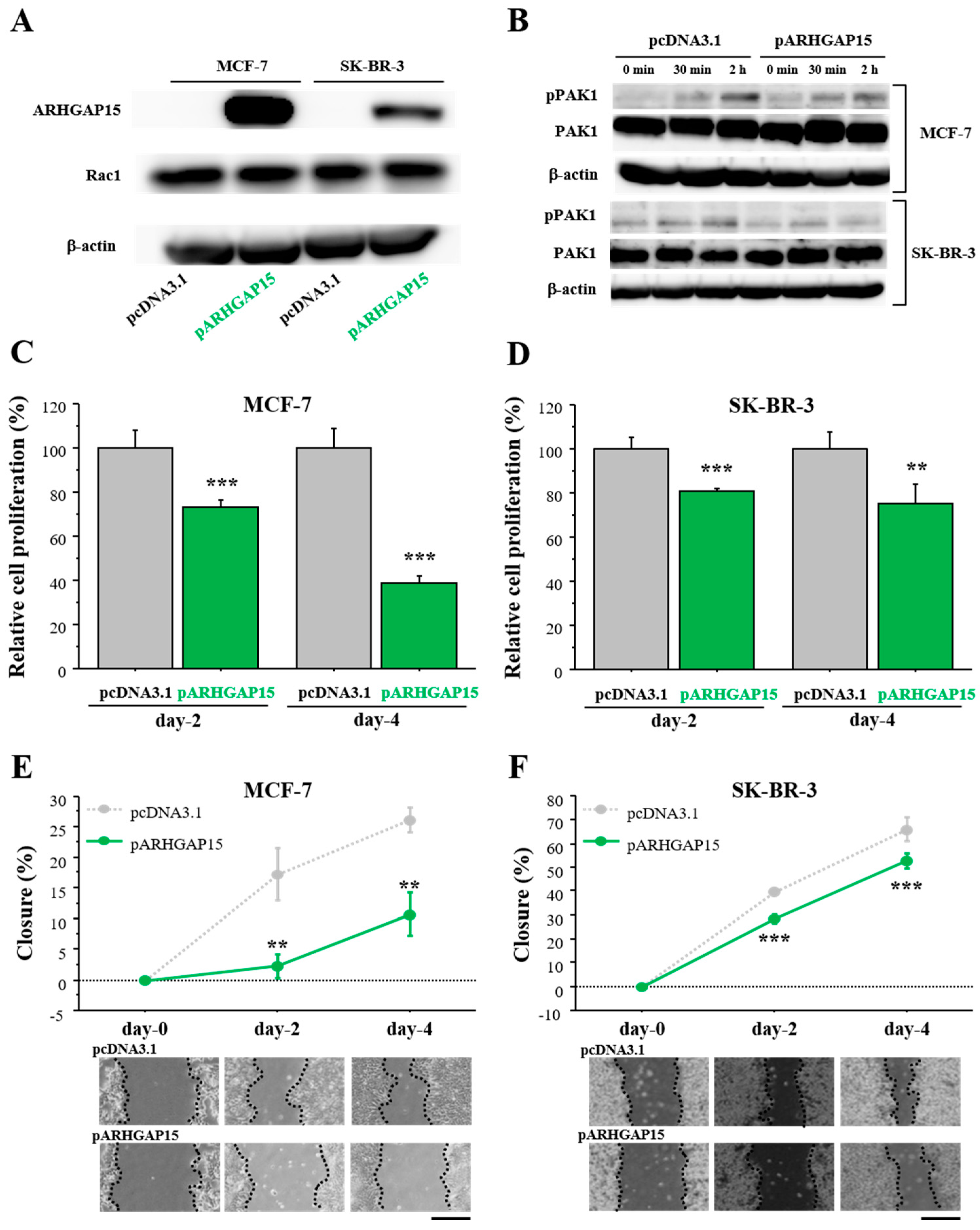
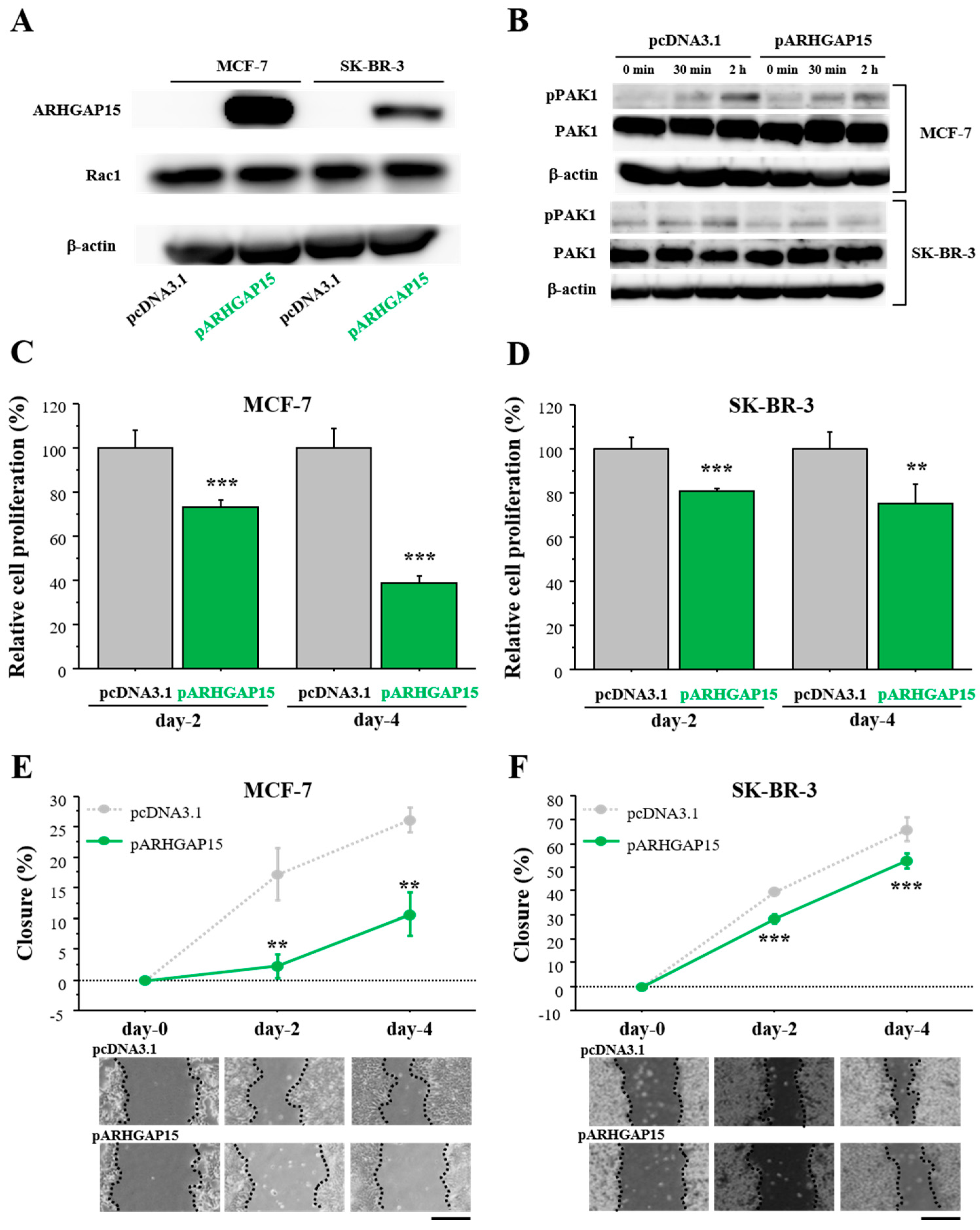
2.3. Effects of ARHGAP15 on Rac1 Activation, Proliferation, and Migration of Breast Carcinoma Cells
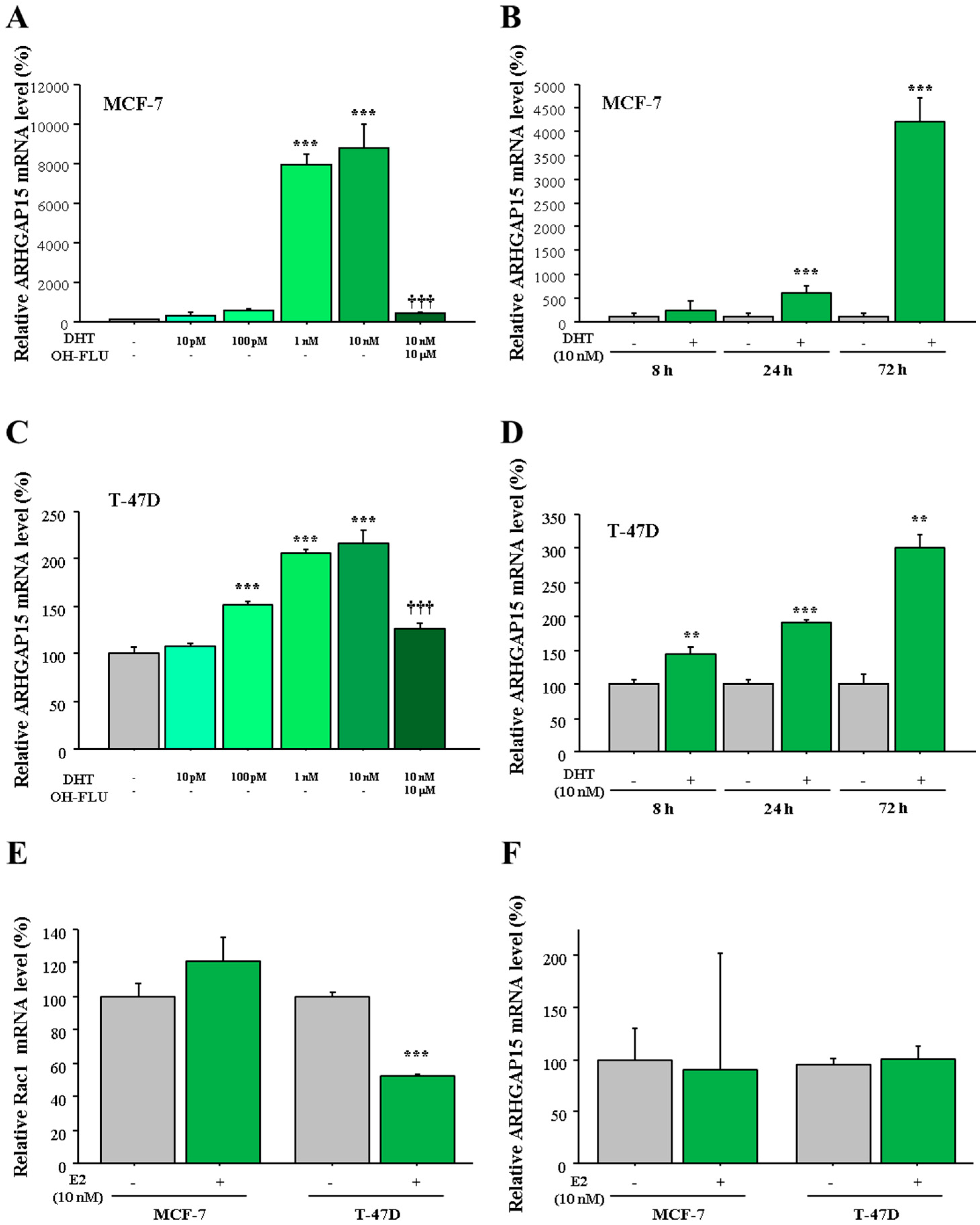
2.4. Regulation of ARHGAP15 and RAC1 Expression by Sex Steroids in Human Breast Carcinomas
3. Discussion
4. Materials and Methods
4.1. Patients and Tissues
4.2. Immunohistochemistry
4.3. Scoring of Immunoreactivity
4.4. Cell Lines and Chemicals
4.5. Real-Time RT-PCR
4.6. Immunoblotting
4.7. Plasmid Construction and Transfection
4.8. Cell Proliferation Assay and Wound Healing Assay
4.9. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AR | Androgen receptor |
| ARHGAP15 | Rho GTPase activating protein |
| DHT | Dihydrotestosterone |
| Cdc42 | Cell division cycle 42 |
| ER | Estrogen receptor |
| E2 | Estradiol |
| HER2 | Human epidermal growth factor receptor 2 |
| HRG | Heregulin-β1 |
| LI | Labeling index |
| OH-FLU | Hydroxyflutamide |
| PAK1 | Phosphorylation of p21 (RAC)-activated kinase 1 |
| PR | Progesterone receptor |
| P-Rex1 | phosphatidylinositol 3,4,5-triphosphate-dependent Rac exchanger 1 |
| pT | Pathological T factor |
| Rac1 | Ras-related C3 botulinum toxin substrate 1 |
| Rho | Ras-related homolog |
| RhoBTB | Rho related BTB domain containing |
| Tiam1 | T-cell lymphoma invasion and metastasis 1 |
| Wrch | Wnt responsive Cdc42 homolog |
| Vav3 | vav guanine nucleotide exchange factor 3 |
References
- Manser, E. Small GTPases take the stage. Dev. Cell 2002, 3, 323–328. [Google Scholar] [CrossRef]
- Wertheimer, E.; Gutierrez-Uzquiza, A.; Rosemblit, C.; Lopez-Haber, C.; Sosa, M.S.; Kazanietz, M.G. Rac signaling in breast cancer: A tale of GEFs and GAPs. Cell. Signal. 2012, 24, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Jansen, S.; Gosens, R.; Wieland, T.; Schmidt, M. Paving the Rho in cancer metastasis: Rho GTPases and beyond. Pharmacol. Ther. 2018, 183, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R.; Sauzeau, V.; Berenjeno, I.M. GTP-binding proteins of the Rho/Rac family: Regulation, effectors and functions in vivo. Bioessays 2007, 29, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Dovas, A.; Couchman, J.R. RhoGDI: Multiple functions in the regulation of Rho family GTPase activities. Biochem. J. 2005, 390, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dokmanovic, M.; Hirsch, D.S.; Shen, Y.; Wu, W.J. Rac1 contributes to trastuzumab resistance of breast cancer cells: Rac1 as a potential therapeutic target for the treatment of trastuzumab-resistant breast cancer. Mol. Cancer Ther. 2009, 8, 1557–1569. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.; Seachrist, D.D.; DeLeon-Rodriguez, C.M.; Lozada, K.L.; Miedler, J.; Abdul-Karim, F.W.; Keri, R.A. HER2/ErbB2-induced breast cancer cell migration and invasion require p120 catenin activation of Rac1 and Cdc42. J. Biol. Chem. 2010, 285, 29491–29501. [Google Scholar] [CrossRef] [PubMed]
- Seoh, M.L.; Ng, C.H.; Yong, J.; Lim, L.; Leung, T. ArhGAP15, a novel human RacGAP protein with GTPase binding property. FEBS Lett. 2003, 539, 131–137. [Google Scholar] [CrossRef]
- Takagi, K.; Miki, Y.; Nagasaki, S.; Hirakawa, H.; Onodera, Y.; Akahira, J.; Ishida, T.; Watanabe, M.; Kimijima, I.; Hayashi, S.; et al. Increased intratumoral androgens in human breast carcinoma following aromatase inhibitor exemestane treatment. Endocr. Relat. Cancer 2010, 17, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zhang, B.; Wang, C.; Fu, T.; Li, L.; Wu, Q.; Cai, Y.; Wang, J. Forkhead box P3 regulates ARHGAP15 expression and affects migration of glioma cells through the Rac1 signaling pathway. Cancer Sci. 2017, 108, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Sosa, M.S.; Lopez-Haber, C.; Yang, C.; Wang, H.; Lemmon, M.A.; Busillo, J.M.; Luo, J.; Benovic, J.L.; Klein-Szanto, A.; Yagi, H.; et al. Identification of the Rac-GEF P-Rex1 as an essential mediator of ErbB signaling in breast cancer. Mol. Cell 2010, 40, 877–892. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, A.; Castoria, G.; Di Domenico, M.; de Falco, A.; Bilancio, A.; Lombardi, M.; Barone, M.V.; Ametrano, D.; Zannini, M.S.; Abbondanza, C.; et al. Steroid-induced androgen receptor-oestradiol receptor β-Src complex triggers prostate cancer cell proliferation. EMBO J. 2000, 19, 5406–5417. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Xiong, J.; Liu, G.; Wu, J.; Wen, L.; Zhang, Q.; Zhang, C. High expression of Rac1 is correlated with partial reversed cell polarity and poor prognosis in invasive ductal carcinoma of the breast. Tumour Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- McHenry, P.R.; Sears, J.C.; Herrick, M.P.; Chang, P.; Heckman-Stoddard, B.M.; Rybarczyk, M.; Chodosh, L.A.; Gunther, E.J.; Hilsenbeck, S.G.; Rosen, J.M.; et al. P190B RhoGAP has pro-tumorigenic functions during MMTV-Neu mammary tumorigenesis and metastasis. Breast Cancer Res. 2010, 12, R73. [Google Scholar] [CrossRef] [PubMed]
- Pliarchopoulou, K.; Kalogeras, K.T.; Kronenwett, R.; Wirtz, R.M.; Eleftheraki, A.G.; Batistatou, A.; Bobos, M.; Soupos, N.; Polychronidou, G.; Gogas, H.; et al. Prognostic significance of RACGAP1 mRNA expression in high-risk early breast cancer: A study in primary tumors of breast cancer patients participating in a randomized Hellenic Cooperative Oncology Group trial. Cancer Chemother. Pharmacol. 2013, 71, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Imaoka, H.; Toiyama, Y.; Saigusa, S.; Kawamura, M.; Kawamoto, A.; Okugawa, Y.; Hiro, J.; Tanaka, K.; Inoue, Y.; Mohri, Y.; et al. RacGAP1 expression, increasing tumor malignant potential, as a predictive biomarker for lymph node metastasis and poor prognosis in colorectal cancer. Carcinogenesis 2015, 6, 46–54. [Google Scholar] [CrossRef]
- Keely, P.J.; Westwick, J.K.; Whitehead, I.P.; Der, C.J.; Parise, L.V. Cdc42 and Rac1 induce integrin-mediated cell motility and invasiveness through PI(3)K. Nature 1997, 390, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Shields, S.K.; Chakraborty, C. Enhanced intrinsic migration of aggressive breast cancer cells by inhibition of Rac1 GTPase. Biochem. Biophys. Res. Commun. 2006, 351, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Marui, N.; Sakai, T.; Morii, N.; Kozaki, S.; Ikai, K.; Imamura, S.; Narumiya, S. ADP-ribosylation of the rhoA gene product by botulinum C3 exoenzyme causes Swiss 3T3 cells to accumulate in the G1 phase of the cell cycle. Oncogene 1993, 8, 1449–1455. [Google Scholar] [PubMed]
- Olson, M.F.; Ashworth, A.; Hall, A. An essential role for Rho, Rac, and Cdc42 GTPases in cell cycle progression through G1. Science 1995, 269, 1270–1272. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.A.; Sethi, R.; Doanes, A.M.; Johnson, T.M.; Pracyk, J.B.; Kirby, M.; Irani, K.; Goldschmidt-Clermont, P.J.; Finkel, T. Rac1 is required for cell proliferation and G2/M progression. Biochem. J. 1997, 326, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Zhang, Y.; Rosado, L.A.R.; Chen, J.; Khan, T.; Moon, S.Y.; Zhang, B. Blockade of Rac1 activity induces G1 cell cycle arrest or apoptosis in breast cancer cells through downregulation of cyclin D1, survivin, and X-linked inhibitor of apoptosis protein. Mol. Cancer Ther. 2010, 9, 1657–1668. [Google Scholar] [CrossRef] [PubMed]
- Gastonguay, A.; Berg, T.; Hauser, A.D.; Schuld, N.; Lorimer, E.; Williams, C.L. The role of Rac1 in the regulation of NF-κB activity, cell proliferation, and cell migration in non-small cell lung carcinoma. Cancer Biol. Ther. 2012, 13, 647–656. [Google Scholar] [CrossRef] [PubMed]
- De Azambuja, E.; Cardoso, F.; de Castro, G., Jr.; Colozza, M.; Mano, M.S.; Durbecq, V.; Sotiriou, C.; Larsimont, D.; Piccart-Gebhart, M.J.; Paesmans, M. Ki-67 as prognostic marker in early breast cancer: A meta-analysis of published studies involving 12,155 patients. Br. J. Cancer 2007, 96, 1504–1513. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miki, Y.; Takagi, K.; Hirakawa, H.; Moriya, T.; Ohuchi, N.; Sasano, H. Androgens in human breast carcinoma. Med. Mol. Morphol. 2010, 43, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Miki, Y.; Ishida, T.; Sasano, H.; Suzuki, T. The interplay of endocrine therapy, steroid pathways and therapeutic resistance: Importance of androgen in breast carcinoma. Mol. Cell. Endocrinol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Merlo, A.; Casalini, P.; Carcangiu, M.L.; Malventano, C.; Triulzi, T.; Mènard, S.; Tagliabue, E.; Balsari, A. FOXP3 expression and overall survival in breast cancer. J. Clin. Oncol. 2009, 27, 1746–1752. [Google Scholar] [CrossRef] [PubMed]
- Douglass, S.; Meeson, A.P.; Overbeck-Zubrzycka, D.; Brain, J.G.; Bennett, M.R.; Lamb, C.A.; Lennard, T.W.; Browell, D.; Ali, S.; Kirby, J.A. Breast cancer metastasis: Demonstration that FOXP3 regulates CXCR4 expression and the response to CXCL12. J. Pathol. 2014, 234, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Laufs, U.; Adam, O.; Strehlow, K.; Wassmann, S.; Konkol, C.; Laufs, K.; Schmidt, W.; Böhm, M.; Nickenig, G. Down-regulation of Rac-1 GTPase by Estrogen. J. Biol. Chem. 2003, 278, 5956–5962. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miki, Y.; Nakamura, Y.; Moriya, T.; Ito, K.; Ohuchi, N.; Sasano, H. Sex steroid-producing enzymes in human breast cancer. Endocr. Relat. Cancer 2005, 12, 701–720. [Google Scholar] [CrossRef] [PubMed]
- Hammond, M.E.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M.; et al. American Society of Clinical Oncology/College of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer. Arch. Pathol. Lab. Med. 2010, 134, e48–e72. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ARHGAP15 Immunoreactivity | p Value | ||
|---|---|---|---|
| Negative (n = 53) | Positive (n = 47) | ||
| Age | 59 (36–81) | 56 (31–80) | 0.65 |
| Menopausal status | |||
| Premenopausal | 16 | 14 | |
| Postmenopausal | 37 | 33 | 0.97 |
| Stage | |||
| I | 17 | 17 | |
| II | 27 | 23 | |
| III | 9 | 7 | 0.90 |
| pT | |||
| pT1 | 21 | 21 | |
| pT2–4 | 32 | 26 | 0.61 |
| Lymph node metastasis | |||
| Negative | 31 | 30 | |
| Positive | 22 | 17 | 0.58 |
| Histological grade | |||
| 1 (well) | 9 | 15 | |
| 2 (intermediate) | 26 | 21 | |
| 3 (poor) | 18 | 11 | 0.19 |
| ER status | |||
| Negative | 9 | 10 | |
| Positive | 44 | 37 | 0.59 |
| PR status | |||
| Negative | 13 | 14 | |
| Positive | 40 | 33 | 0.55 |
| AR LI (%) * | 14.5 (0–86) | 28 (0–89) | 0.033 |
| HER2 status | |||
| Negative | 45 | 37 | |
| Positive | 8 | 10 | 0.42 |
| Ki-67 LI (%) * | 17 (0–82) | 14 (0–54) | 0.42 |
| Rac1 | |||
| Negative | 39 | 21 | |
| Positive | 14 | 26 | 0.003 |
| Rac1 Immunoreactivity | p Value | ||
|---|---|---|---|
| Negative (n = 60) | Positive (n = 40) | ||
| Age | 56.5 (31–81) | 59 (37–80) | 0.33 |
| Menopausal status | |||
| Premenopausal | 22 | 8 | |
| Postmenopausal | 38 | 32 | 0.074 |
| Stage | |||
| I | 23 | 11 | |
| II | 30 | 20 | |
| III | 7 | 9 | 0.27 |
| pT | |||
| pT1 | 29 | 13 | |
| pT2–4 | 31 | 27 | 0.12 |
| Lymph node metastasis | |||
| Negative | 39 | 22 | |
| Positive | 21 | 18 | 0.088 |
| Histological grade | |||
| 1 (well) | 14 | 10 | |
| 2 (intermediate) | 33 | 14 | |
| 3 (poor) | 13 | 16 | 0.19 |
| ER status | |||
| Negative | 7 | 12 | |
| Positive | 53 | 28 | 0.022 |
| PR status | |||
| Negative | 10 | 17 | |
| Positive | 50 | 23 | 0.0044 |
| AR LI (%) * | 21.5 (0–89) | 17 (0–83) | 0.87 |
| HER2 status | |||
| Negative | 51 | 31 | |
| Positive | 9 | 9 | 0.34 |
| Ki-67 LI (%) * | 14 (0–73) | 18.5 (0–82) | 0.14 |
| Parameter | Univariate | Multivariate | |
|---|---|---|---|
| p Value | p Value | Relative Risk (95% CI) | |
| Age (31–81) * | 0.77 | ||
| Menopausal status (Post-/Pre-) | 0.62 | ||
| pT (2–4/1) | 0.077 | 0.17 | |
| Lymph node metastasis (positive/negative) | 0.025 | 0.0087 | 3.8 (1.4–12) |
| Histological grade (3/1,2) | 0.89 | ||
| ER status (negative/positive) | 0.89 | ||
| PR status (negative/positive) | 0.26 | ||
| AR status (negative/positive) | 0.47 | ||
| HER2 status (positive/negative) | 0.089 | 0.0085 | 4.7 (1.5–14) |
| Ki67 (≥10/<10) | 0.60 | ||
| ARHGAP15 (negative/positive) | 0.0008 | 0.0005 | 6.9 (2.2–31) |
| Rac1 (positive/negative) | 0.43 | ||
| Parameter | Univariate | Multivariate | |
|---|---|---|---|
| p Value | p Value | Relative Risk (95% CI) | |
| Age (31–81) * | 0.88 | ||
| Menopausal status (Post-/Pre-) | 0.72 | ||
| pT (2–4/1) | 0.053 | 0.37 | |
| Lymph node metastasis (positive/negative) | 0.17 | ||
| Histological grade (3/1,2) | 0.0061 | 0.042 | 4.8 (1.06–35) |
| ER status (negative/positive) | 0.18 | ||
| PR status (negative/positive) | 0.45 | ||
| AR status (negative/positive) | 0.34 | ||
| HER2 status (positive/negative) | 0.039 | 0.043 | 4.6 (1.1–20) |
| Ki67 (≥10/<10) | 0.13 | ||
| ARHGAP15 (negative/positive) | 0.023 | 0.016 | 8.5 (1.4–163) |
| Rac1 (positive/negative) | 0.19 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takagi, K.; Miki, Y.; Onodera, Y.; Ishida, T.; Watanabe, M.; Sasano, H.; Suzuki, T. ARHGAP15 in Human Breast Carcinoma: A Potent Tumor Suppressor Regulated by Androgens. Int. J. Mol. Sci. 2018, 19, 804. https://doi.org/10.3390/ijms19030804
Takagi K, Miki Y, Onodera Y, Ishida T, Watanabe M, Sasano H, Suzuki T. ARHGAP15 in Human Breast Carcinoma: A Potent Tumor Suppressor Regulated by Androgens. International Journal of Molecular Sciences. 2018; 19(3):804. https://doi.org/10.3390/ijms19030804
Chicago/Turabian StyleTakagi, Kiyoshi, Yasuhiro Miki, Yoshiaki Onodera, Takanori Ishida, Mika Watanabe, Hironobu Sasano, and Takashi Suzuki. 2018. "ARHGAP15 in Human Breast Carcinoma: A Potent Tumor Suppressor Regulated by Androgens" International Journal of Molecular Sciences 19, no. 3: 804. https://doi.org/10.3390/ijms19030804
APA StyleTakagi, K., Miki, Y., Onodera, Y., Ishida, T., Watanabe, M., Sasano, H., & Suzuki, T. (2018). ARHGAP15 in Human Breast Carcinoma: A Potent Tumor Suppressor Regulated by Androgens. International Journal of Molecular Sciences, 19(3), 804. https://doi.org/10.3390/ijms19030804