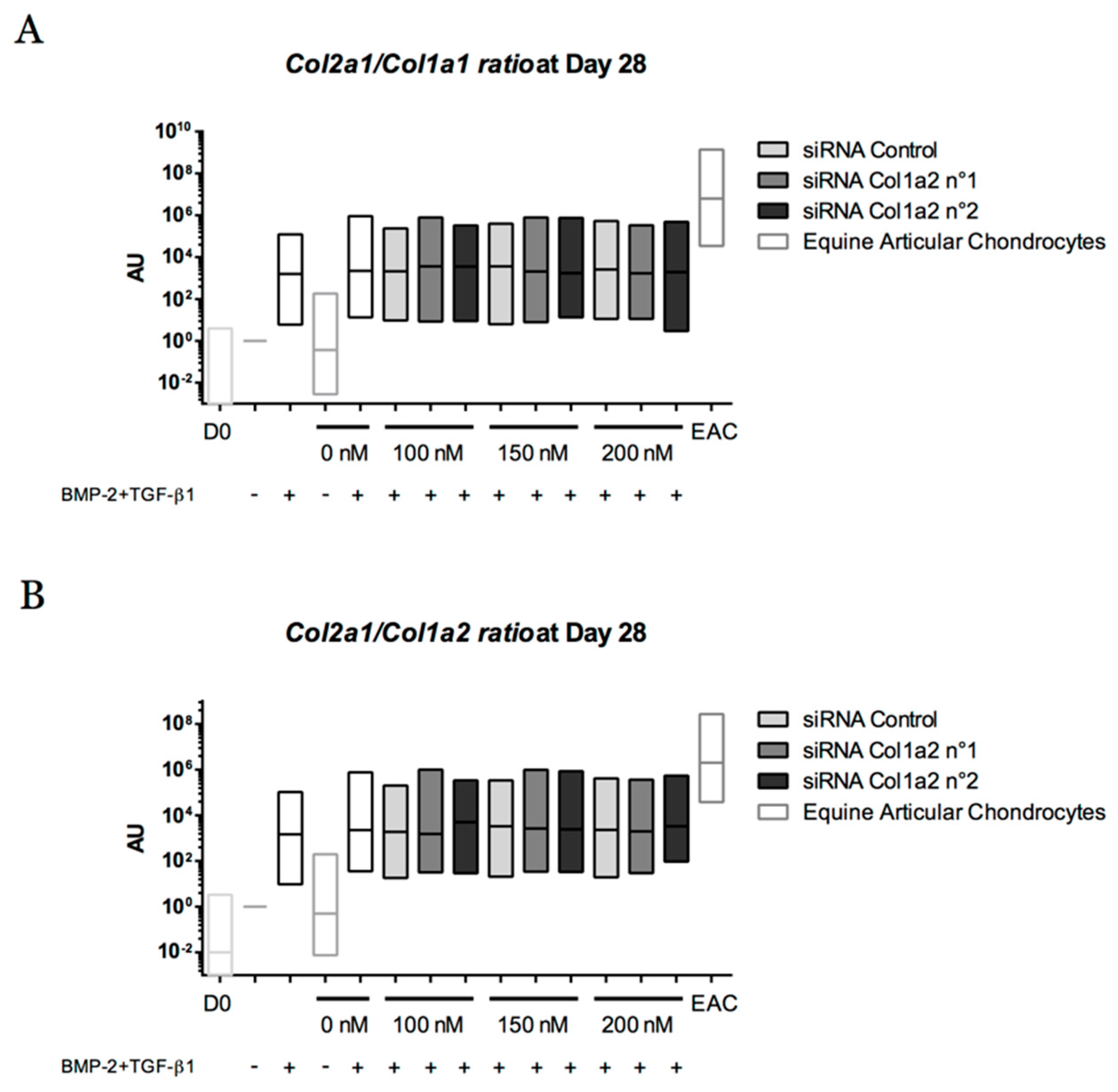
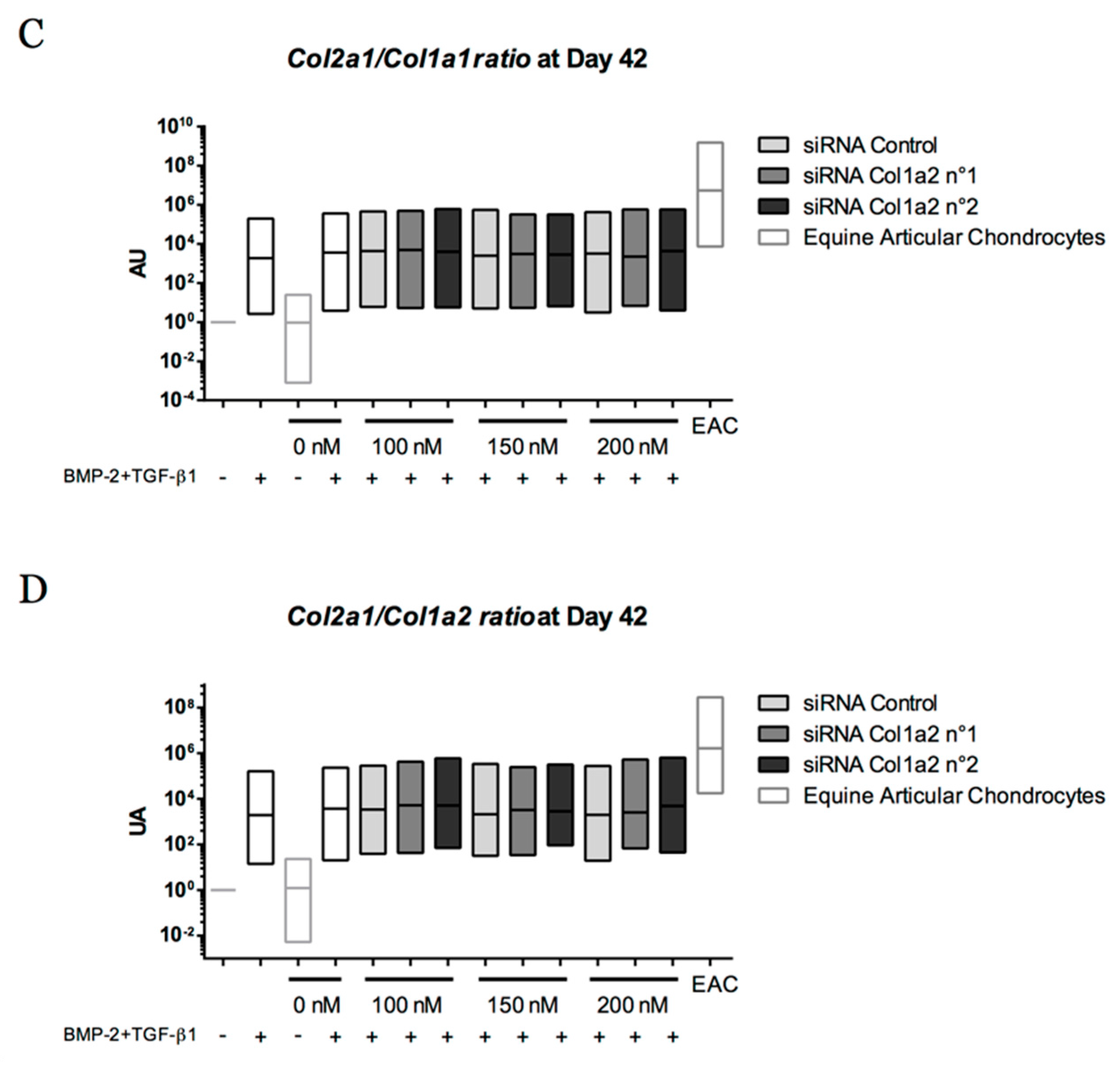
Chondrogenic Differentiation of Defined Equine Mesenchymal Stem Cells Derived from Umbilical Cord Blood for Use in Cartilage Repair Therapy

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
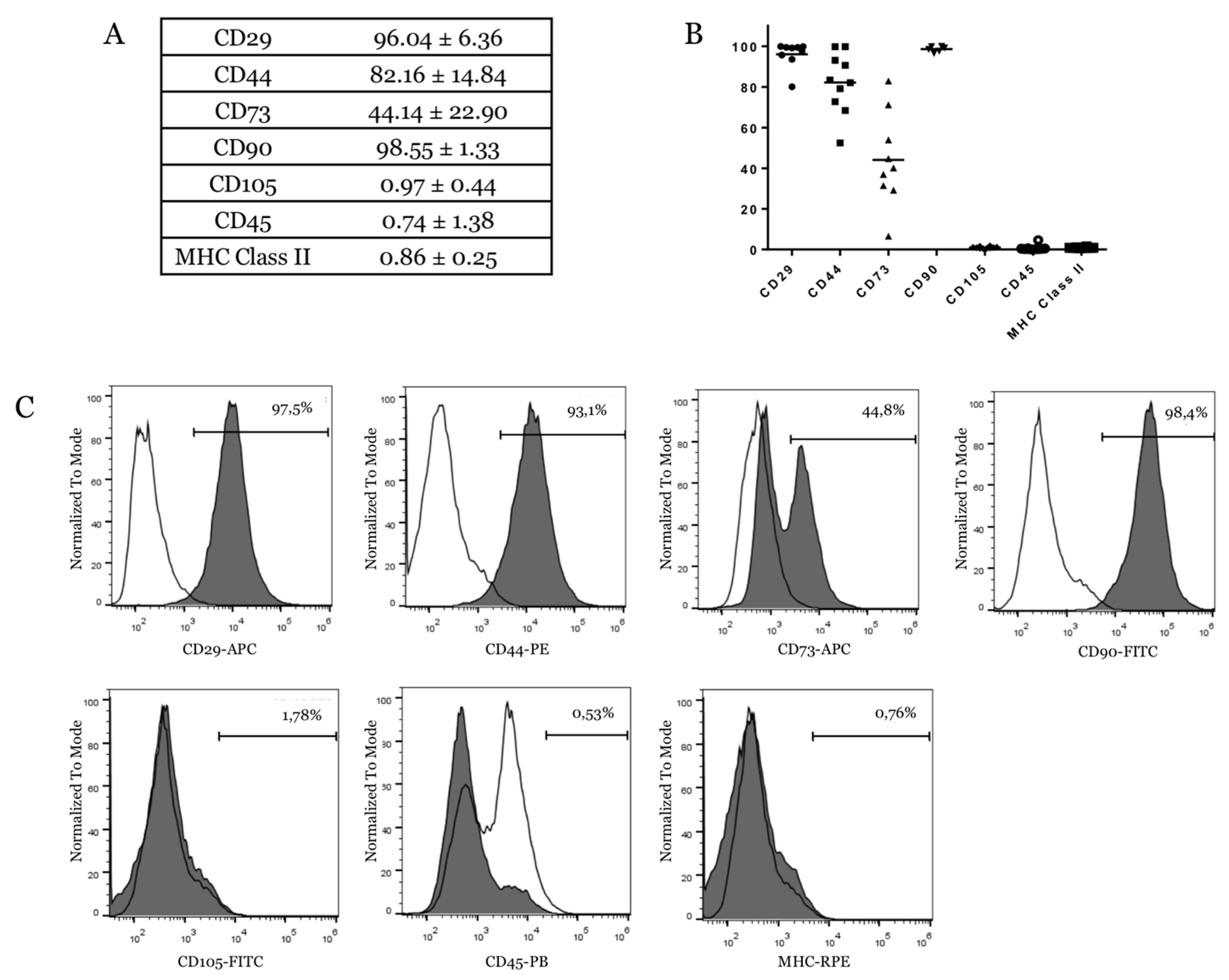{kind=link}
{kind=link}
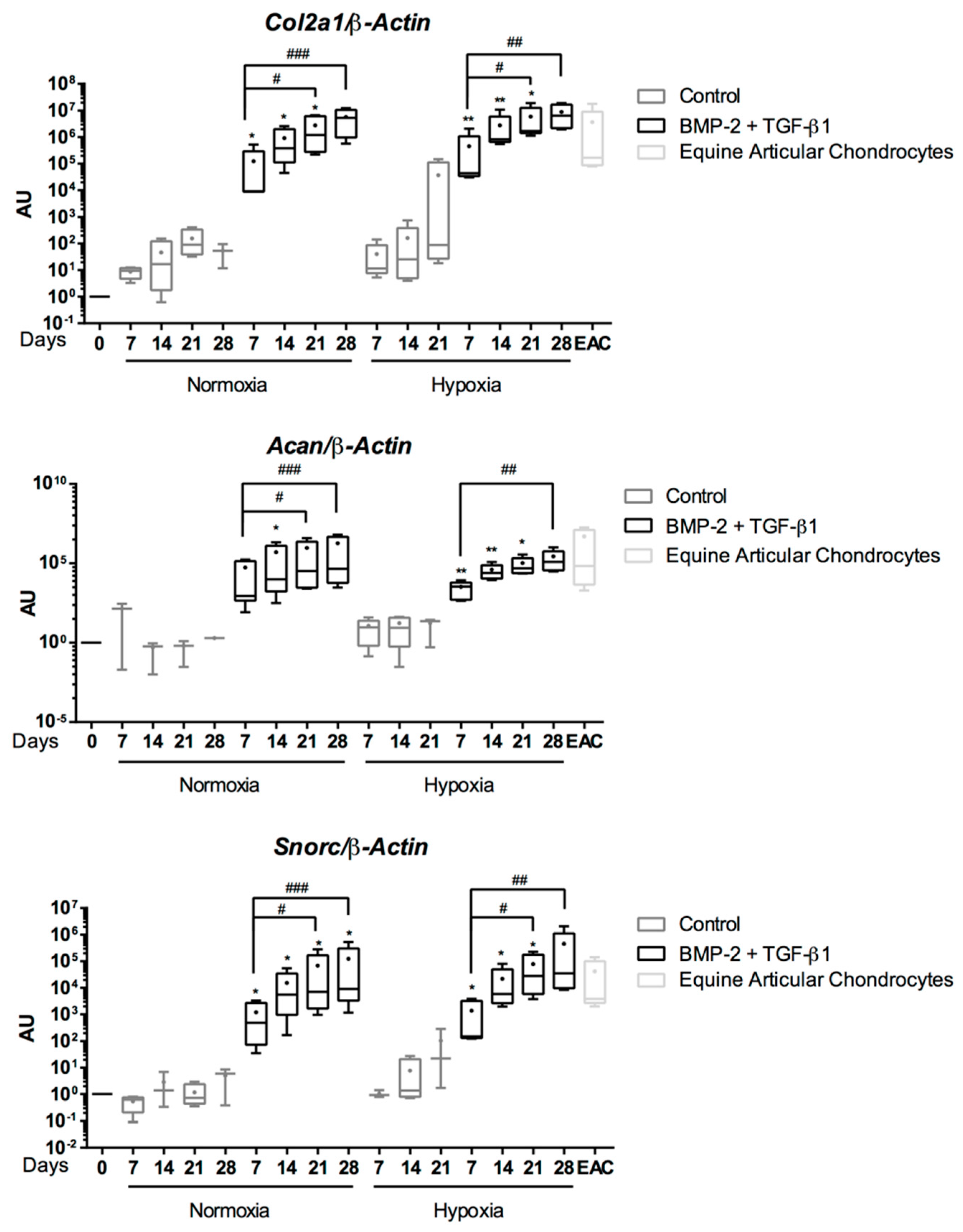
{kind=link}
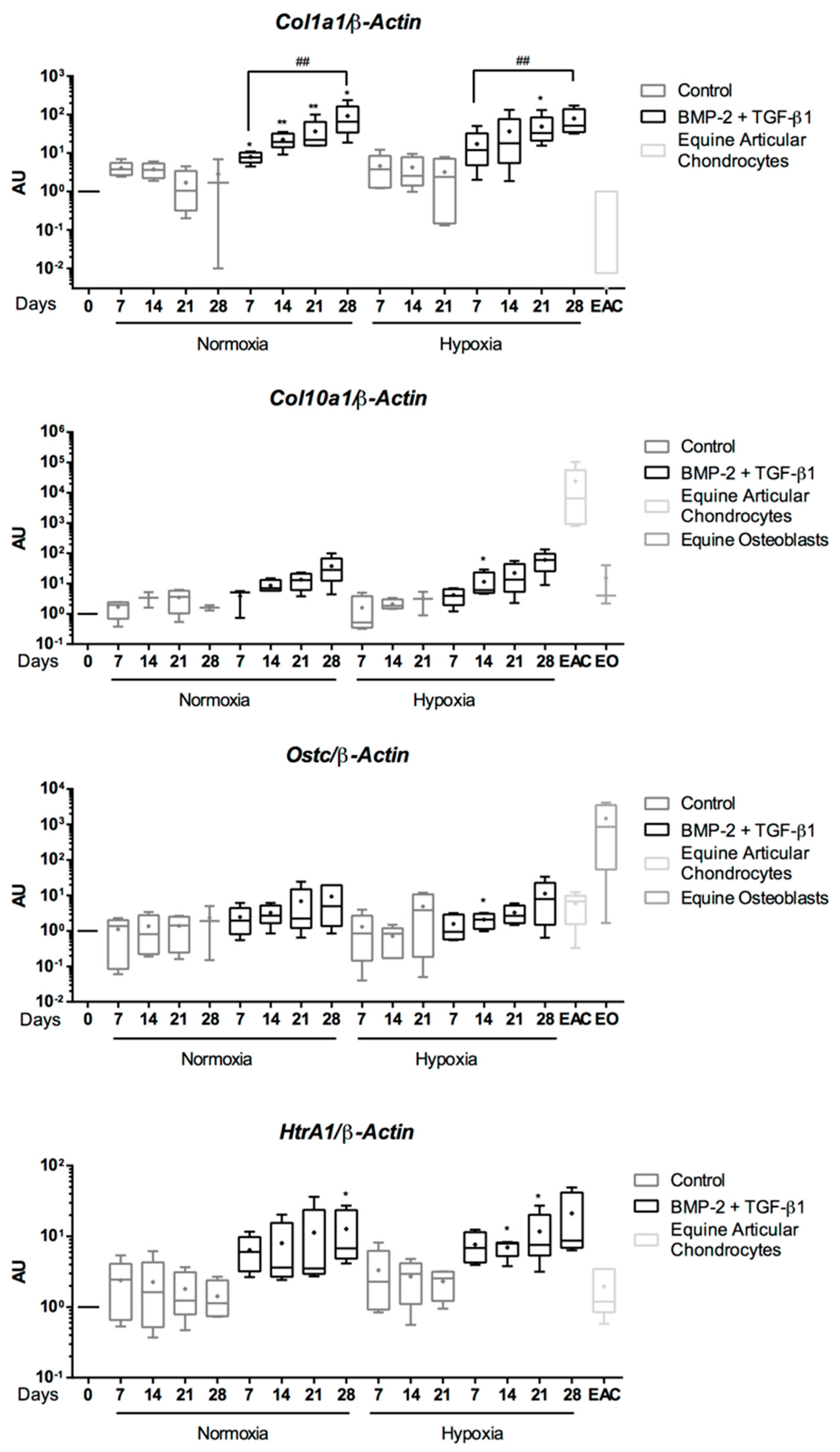
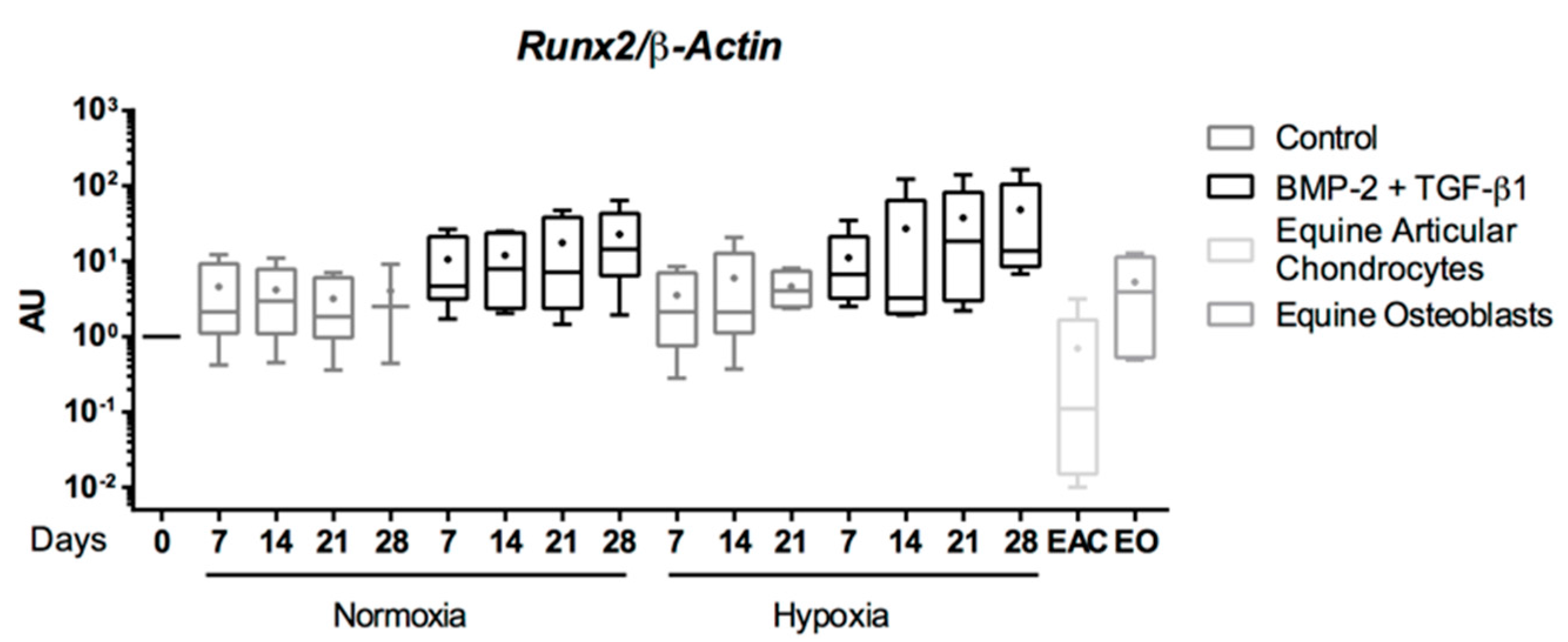{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
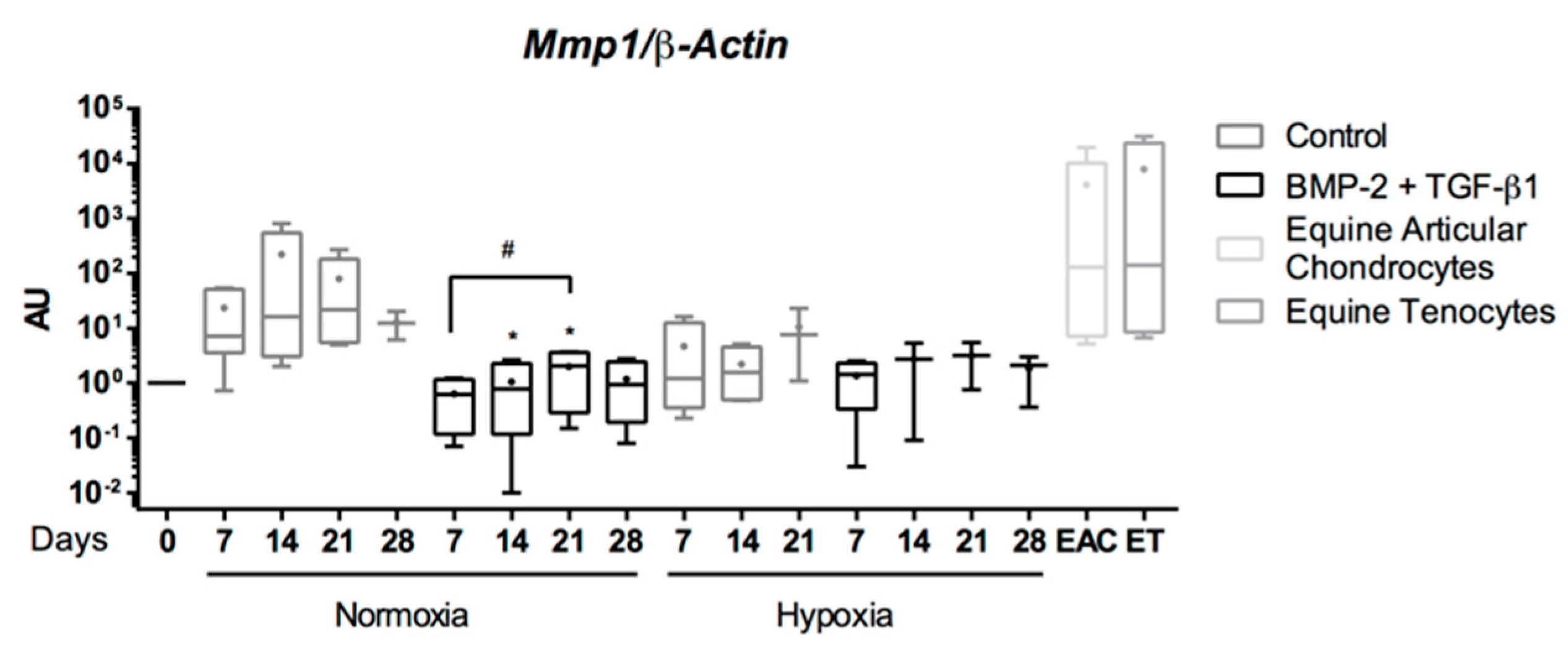
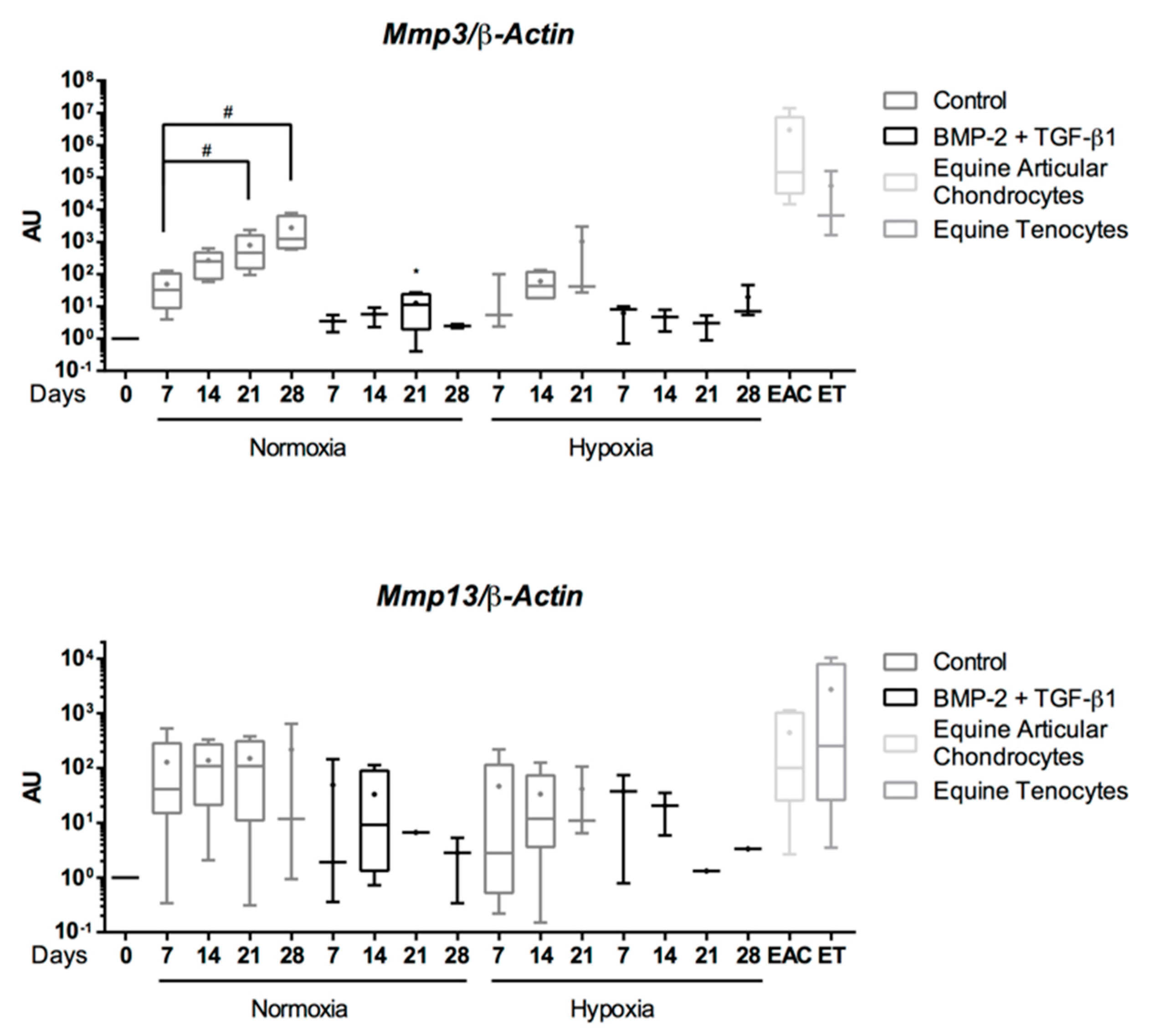
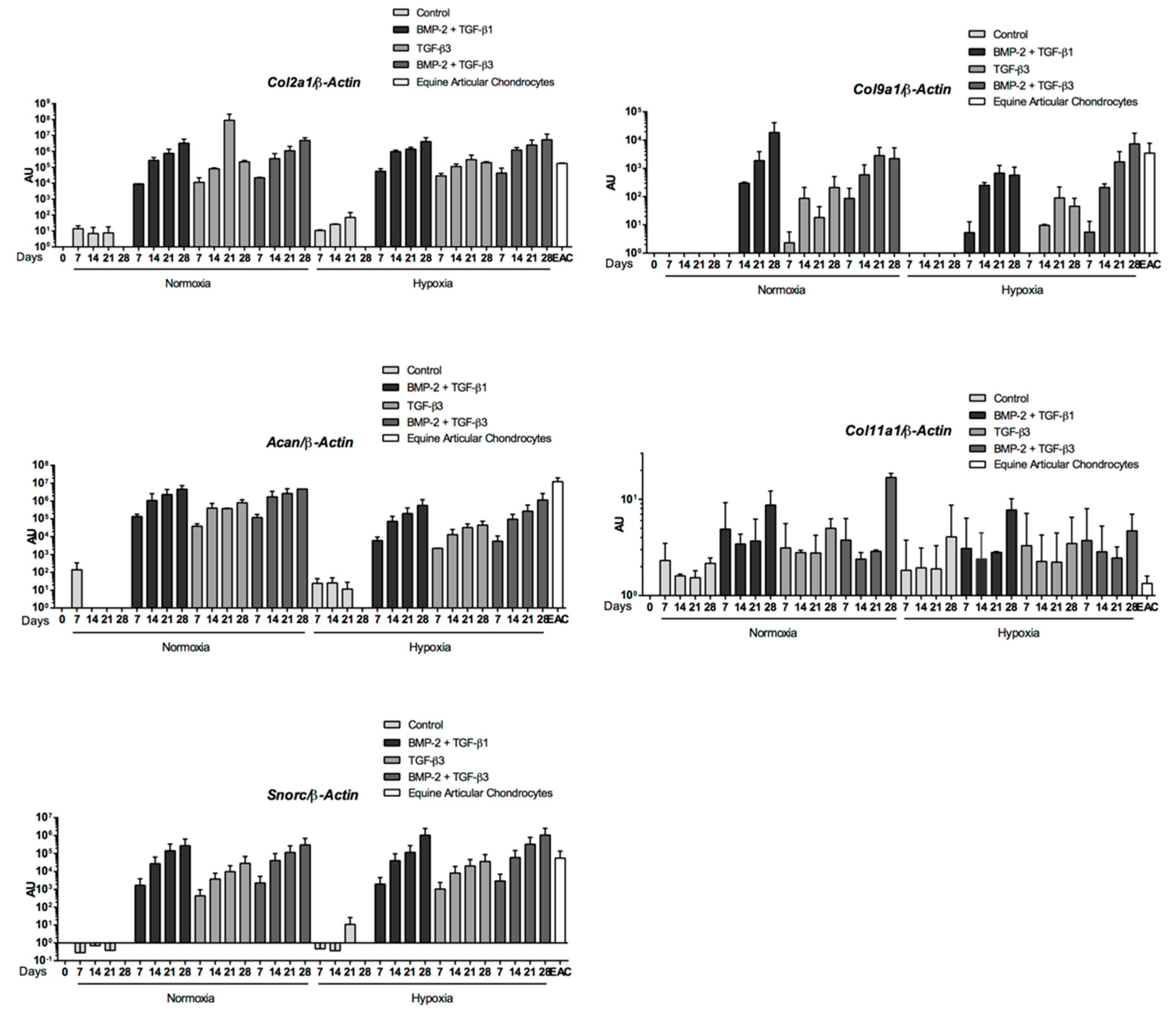
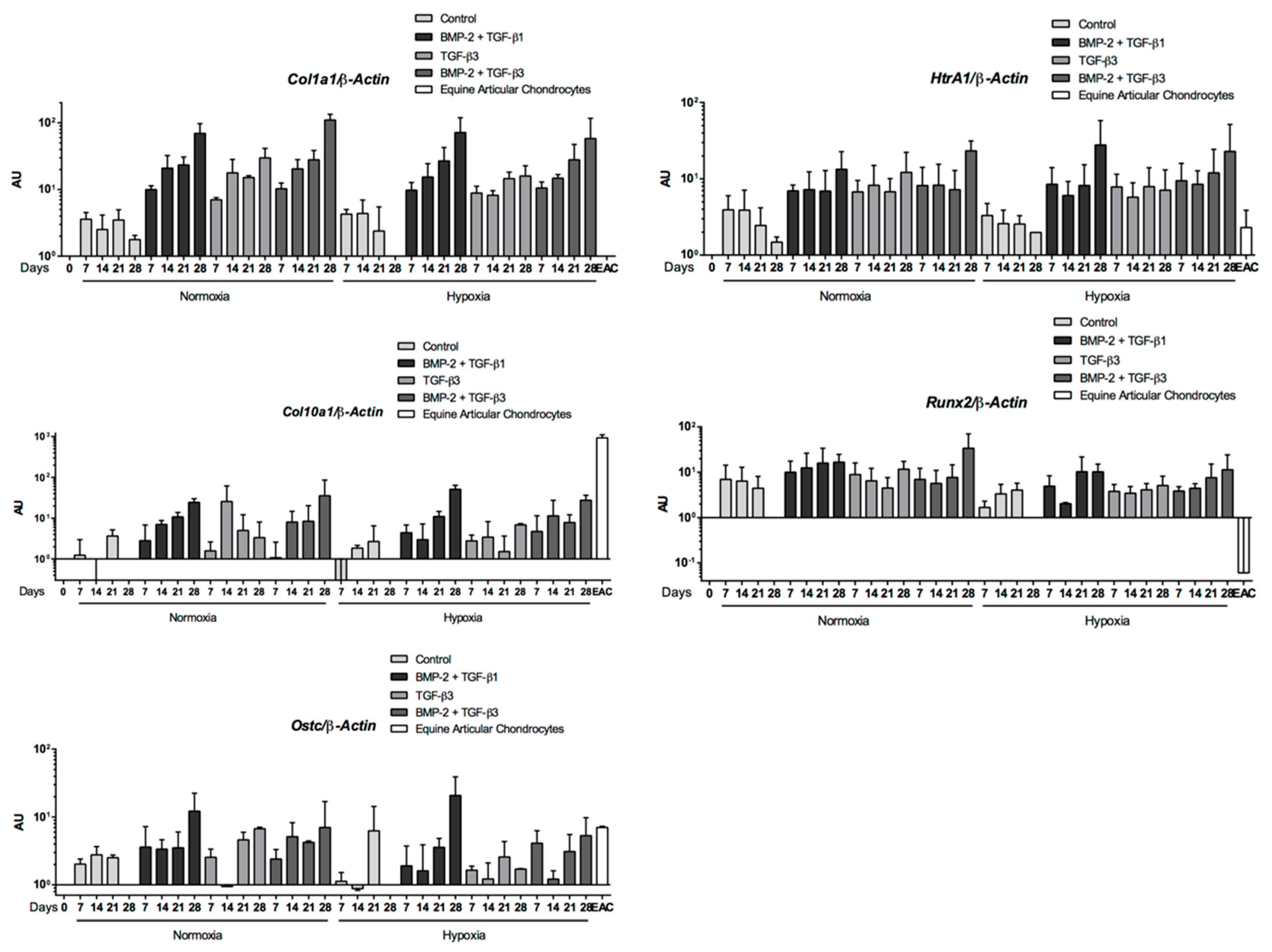
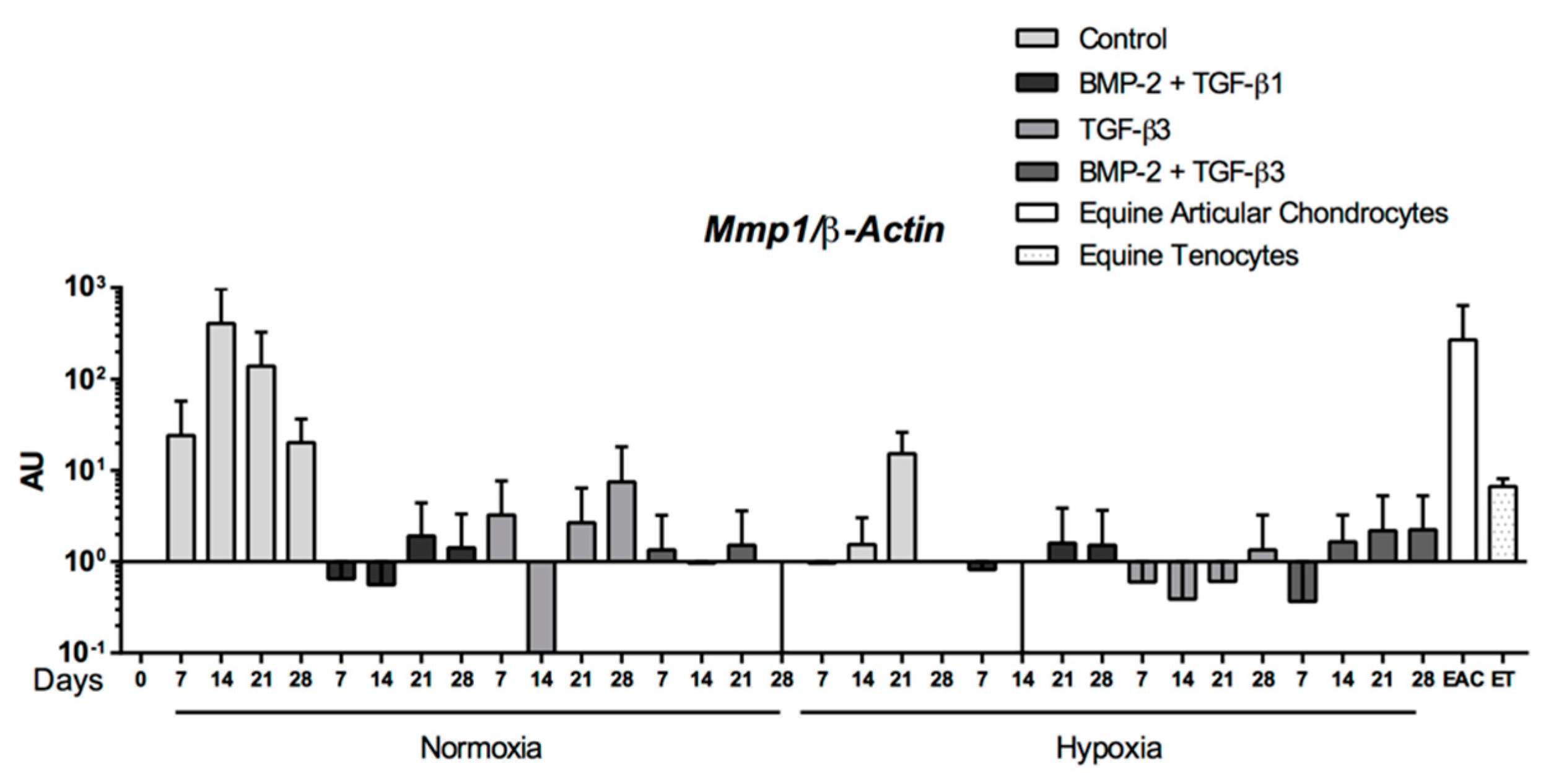
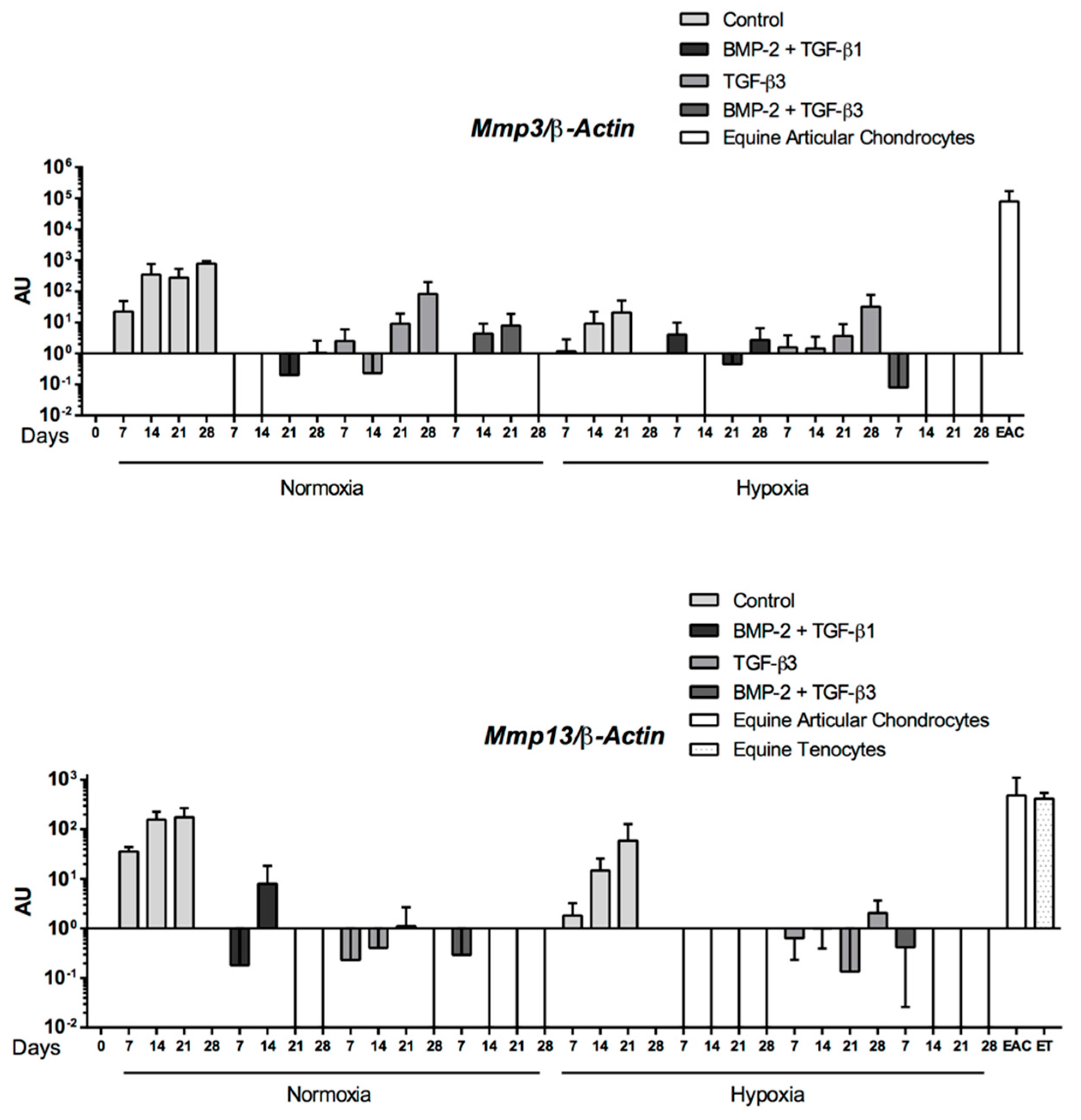
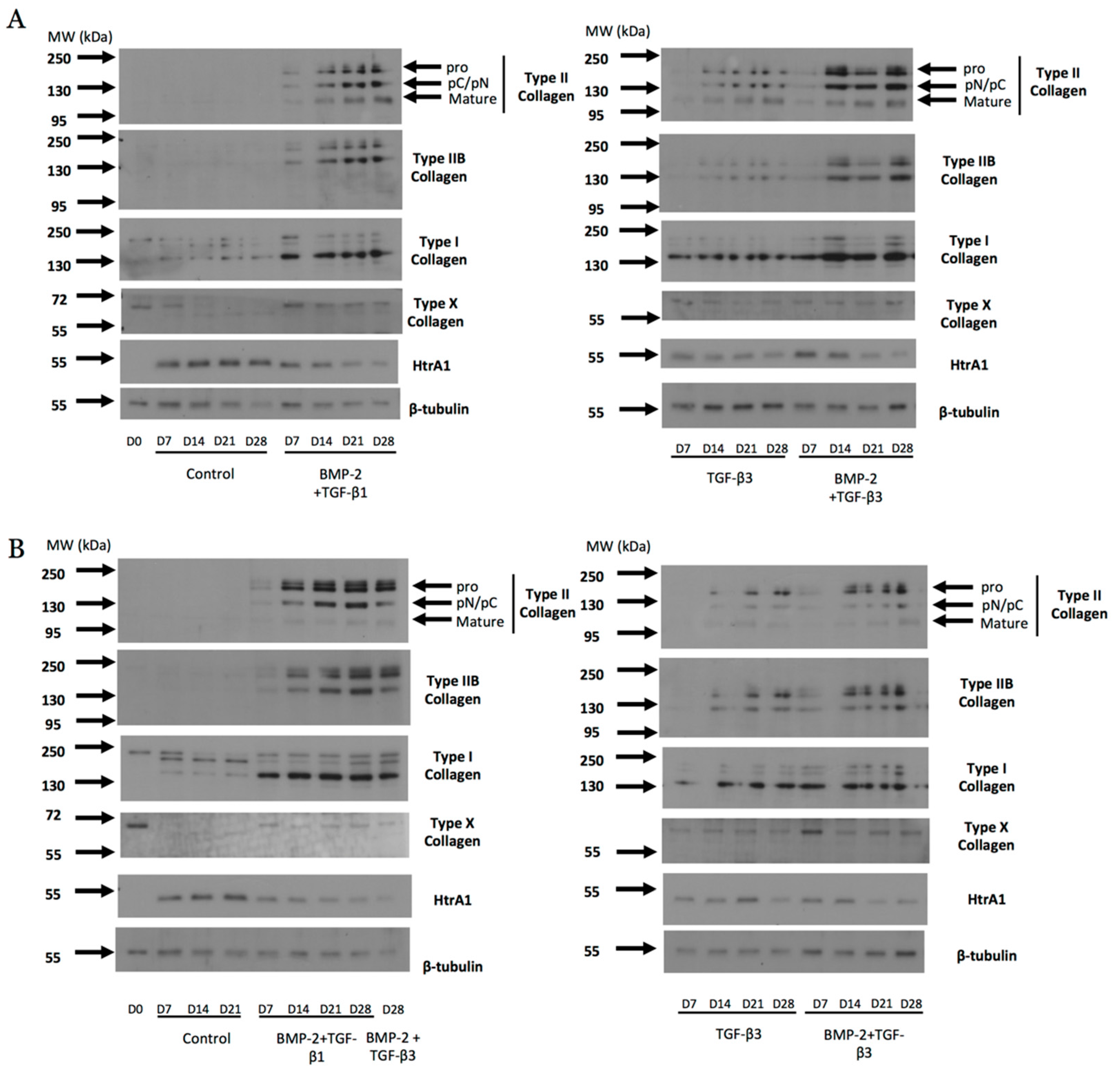
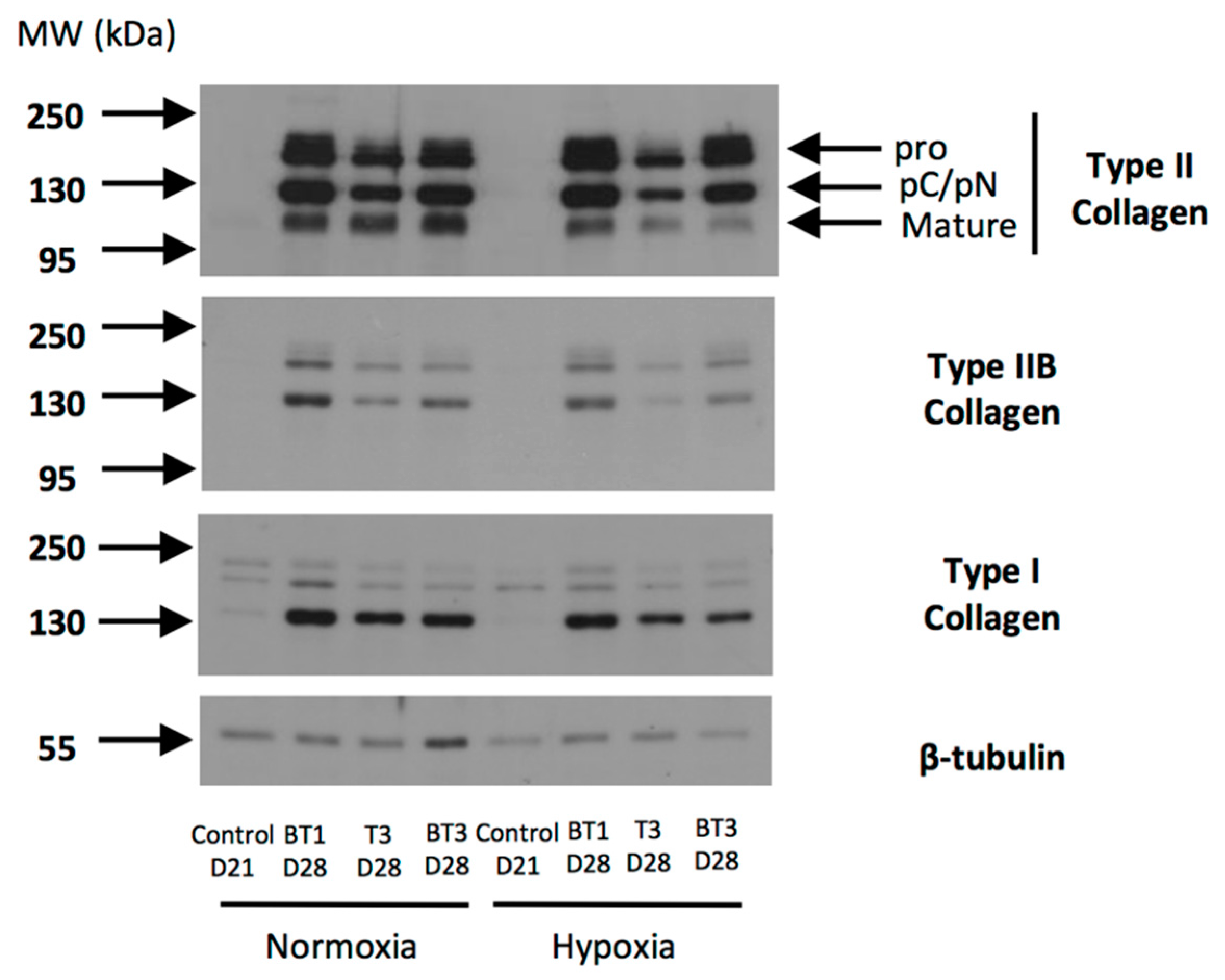
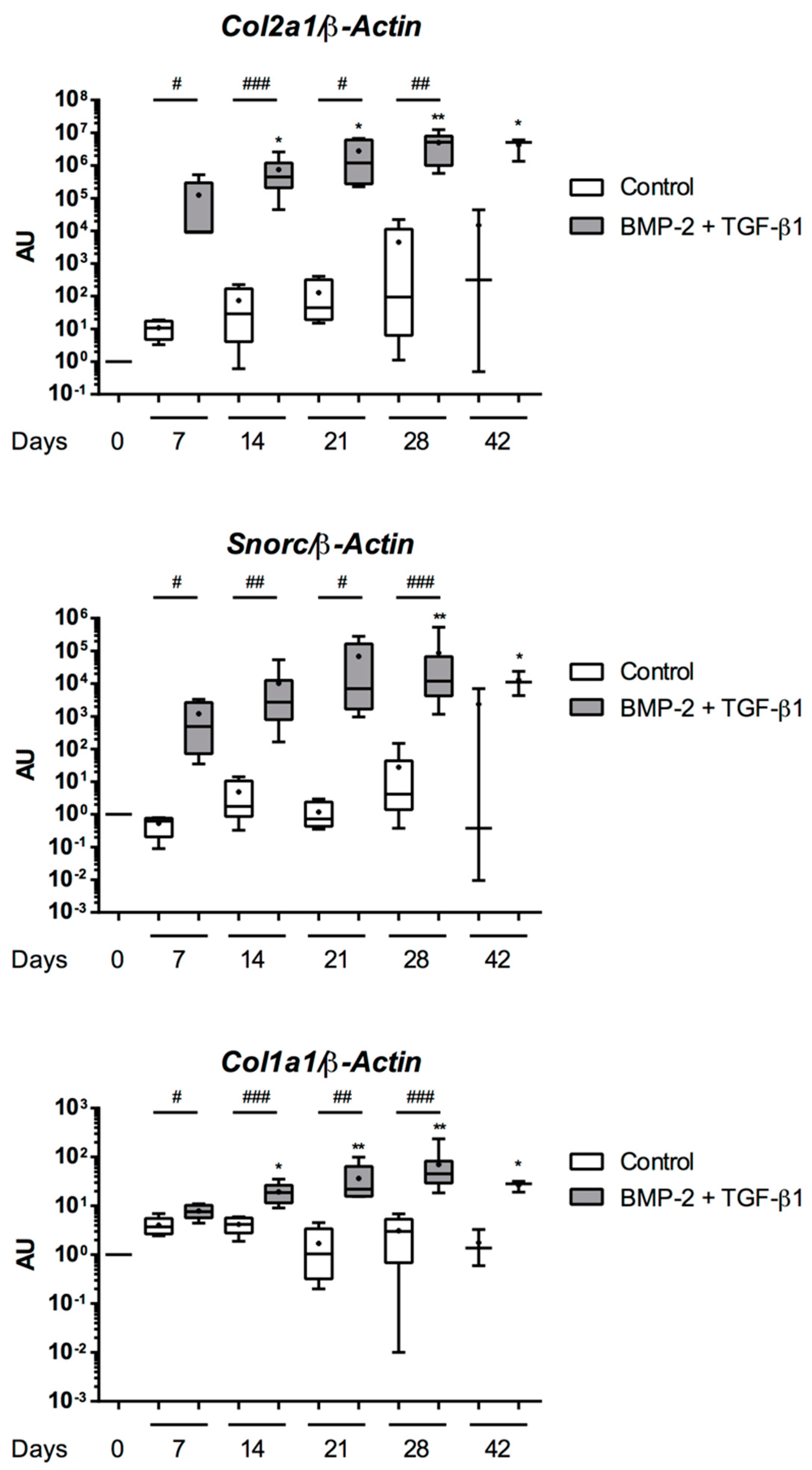
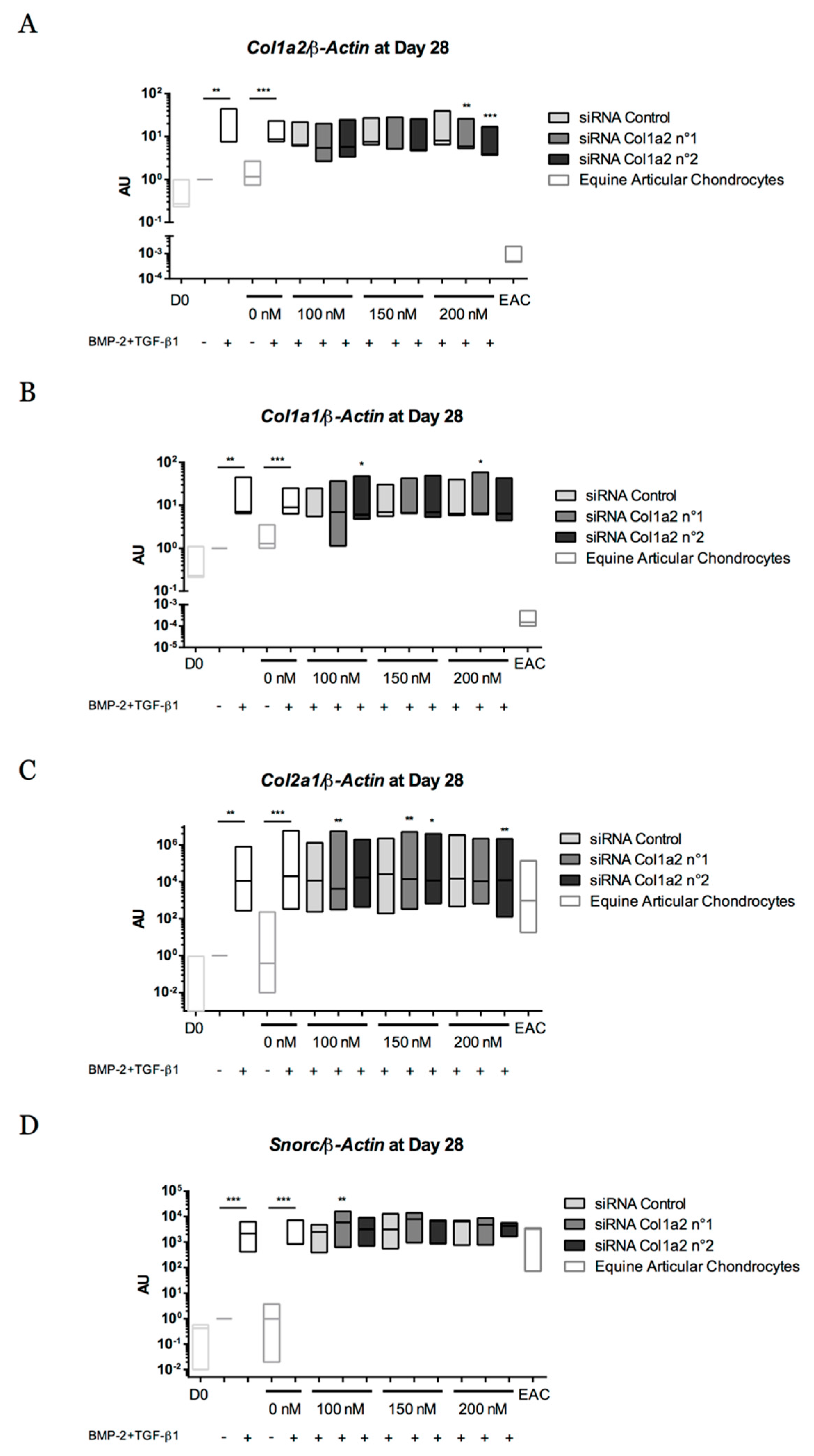
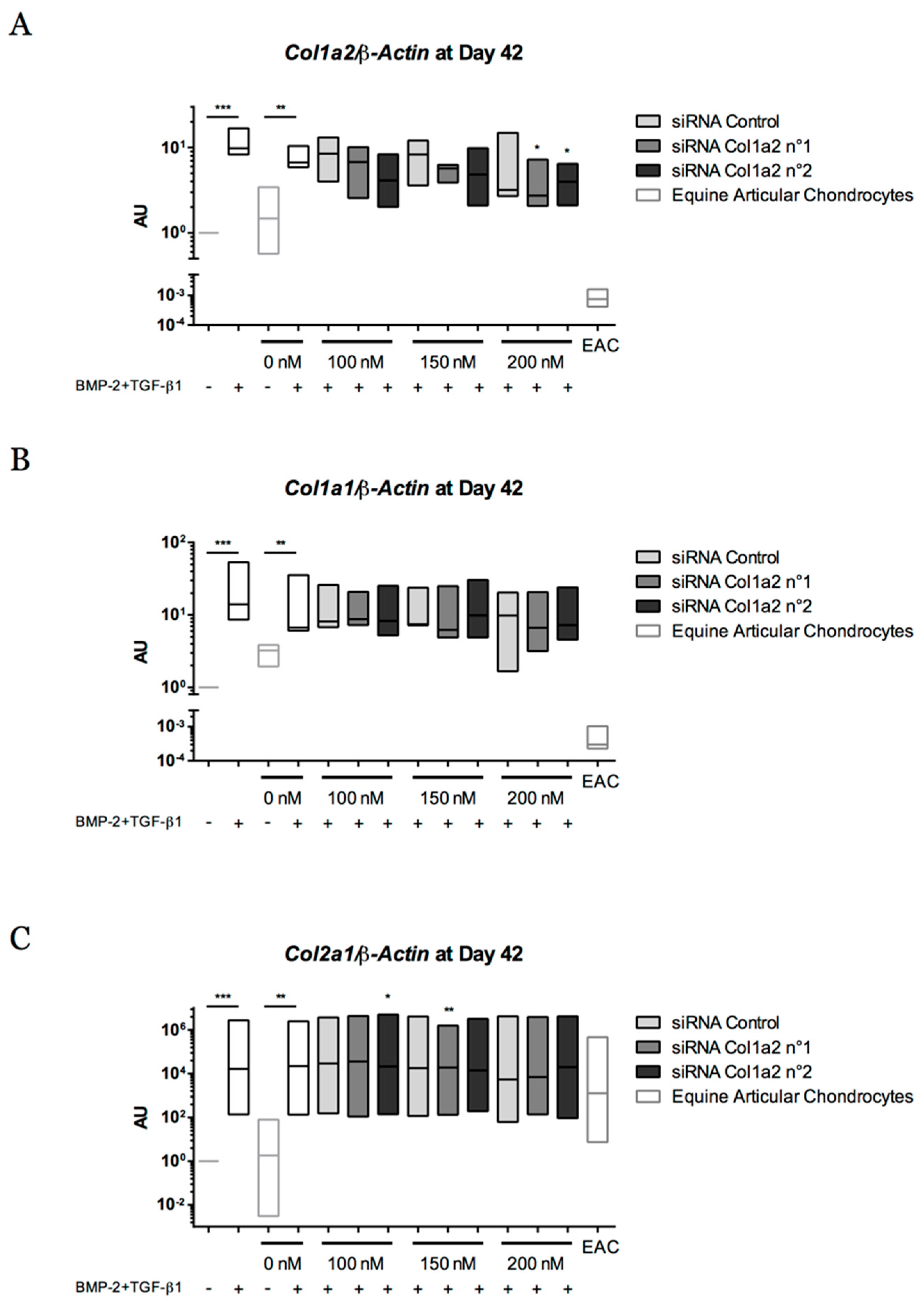
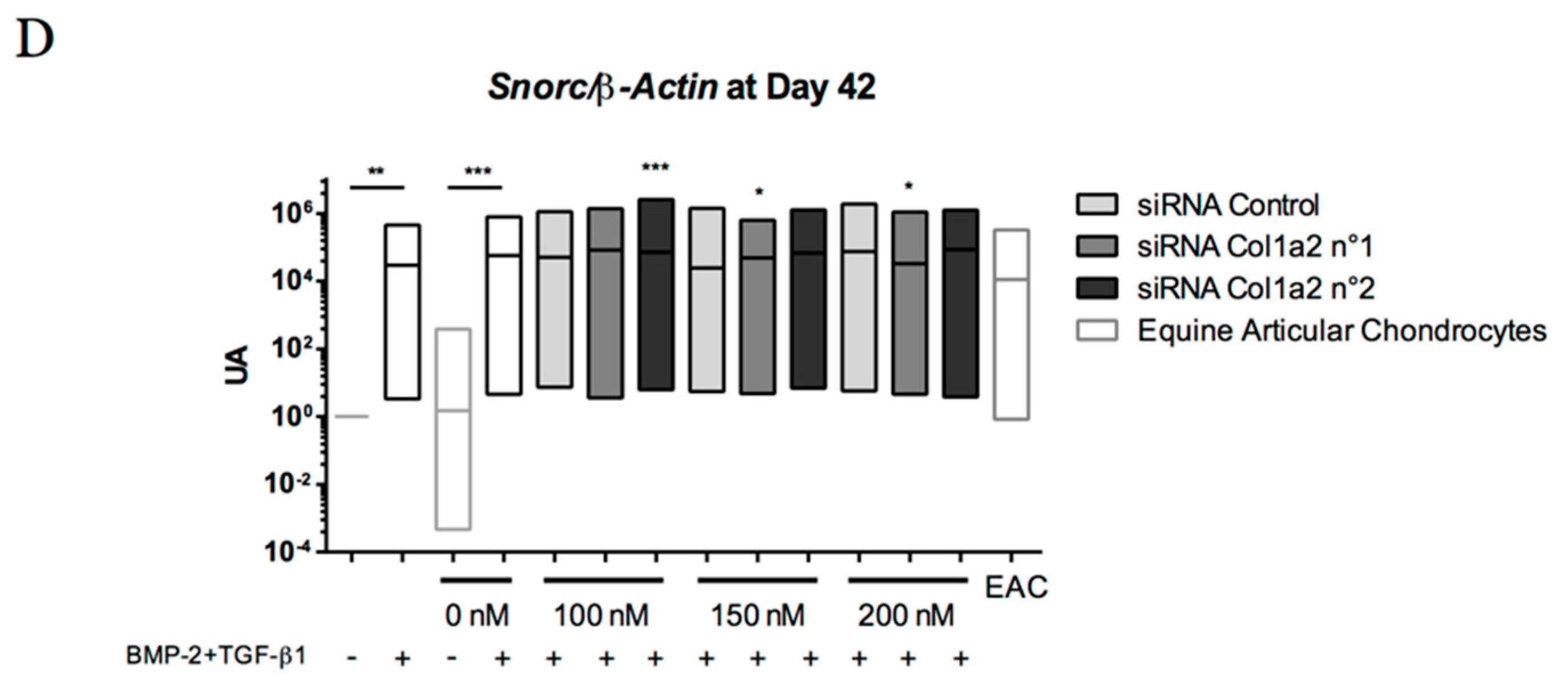
Abstract
Share and Cite
Desancé, M.; Contentin, R.; Bertoni, L.; Gomez-Leduc, T.; Branly, T.; Jacquet, S.; Betsch, J.-M.; Batho, A.; Legendre, F.; Audigié, F.; et al. Chondrogenic Differentiation of Defined Equine Mesenchymal Stem Cells Derived from Umbilical Cord Blood for Use in Cartilage Repair Therapy. Int. J. Mol. Sci. 2018, 19, 537. https://doi.org/10.3390/ijms19020537
Desancé M, Contentin R, Bertoni L, Gomez-Leduc T, Branly T, Jacquet S, Betsch J-M, Batho A, Legendre F, Audigié F, et al. Chondrogenic Differentiation of Defined Equine Mesenchymal Stem Cells Derived from Umbilical Cord Blood for Use in Cartilage Repair Therapy. International Journal of Molecular Sciences. 2018; 19(2):537. https://doi.org/10.3390/ijms19020537
Chicago/Turabian StyleDesancé, Mélanie, Romain Contentin, Lélia Bertoni, Tangni Gomez-Leduc, Thomas Branly, Sandrine Jacquet, Jean-Marc Betsch, Agnès Batho, Florence Legendre, Fabrice Audigié, and et al. 2018. "Chondrogenic Differentiation of Defined Equine Mesenchymal Stem Cells Derived from Umbilical Cord Blood for Use in Cartilage Repair Therapy" International Journal of Molecular Sciences 19, no. 2: 537. https://doi.org/10.3390/ijms19020537
APA StyleDesancé, M., Contentin, R., Bertoni, L., Gomez-Leduc, T., Branly, T., Jacquet, S., Betsch, J.-M., Batho, A., Legendre, F., Audigié, F., Galéra, P., & Demoor, M. (2018). Chondrogenic Differentiation of Defined Equine Mesenchymal Stem Cells Derived from Umbilical Cord Blood for Use in Cartilage Repair Therapy. International Journal of Molecular Sciences, 19(2), 537. https://doi.org/10.3390/ijms19020537