The Differential Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on Cardiometabolic Risk Factors: A Systematic Review

Abstract
1. Introduction
2. Results
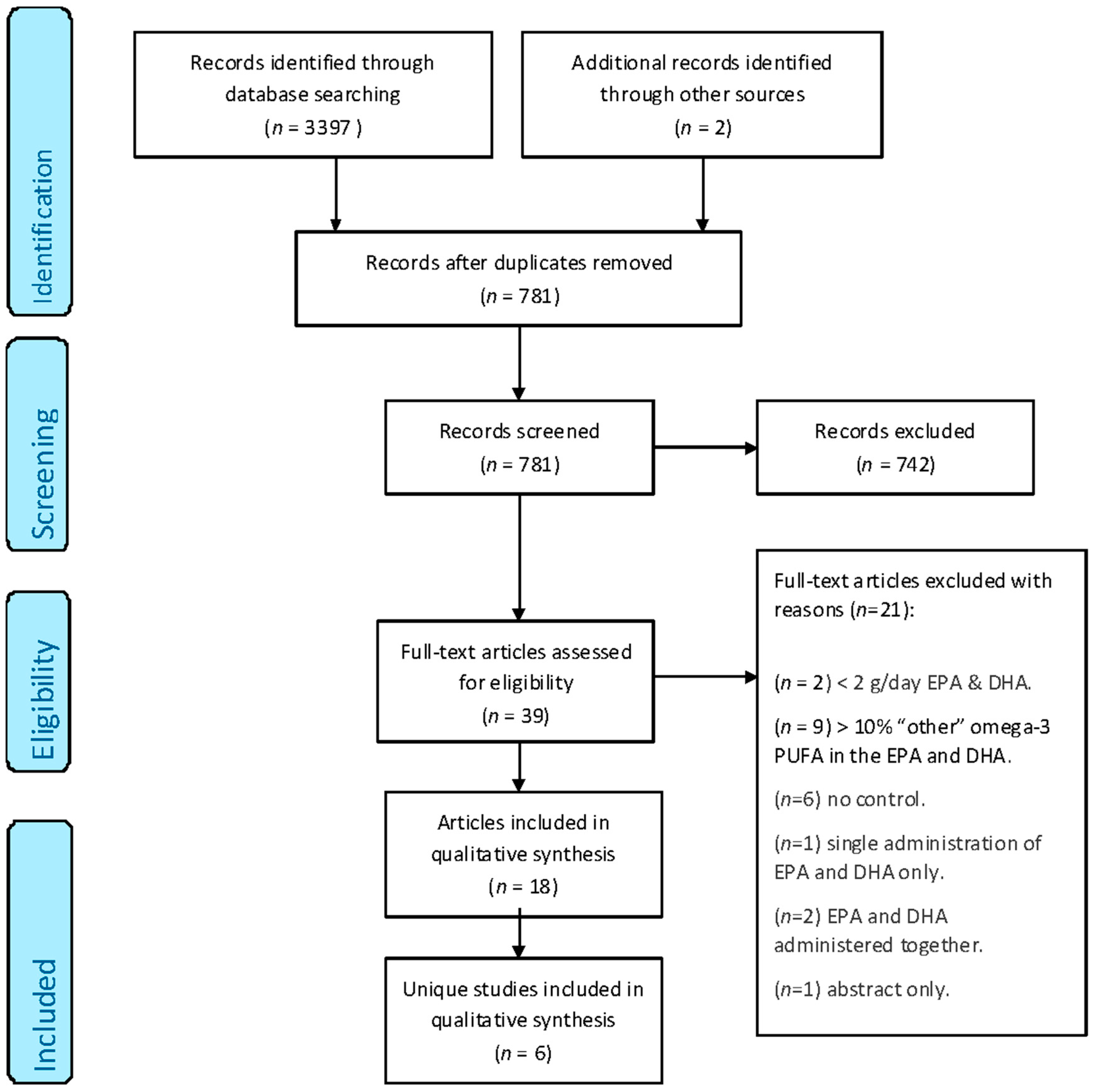
2.1. Identification of the Included Studies
2.2. Characteristics of the Included Studies
2.3. Comparative Effects of EPA and DHA on Cardiometabolic Risk Factors
2.3.1. Effect of EPA versus DHA on Blood Lipids and Lipoproteins
2.3.2. Effect of EPA versus DHA on Haemodynamics
2.3.3. Effect of EPA versus DHA on Platelet and Fibrinolytic Function
2.3.4. Effect of EPA versus DHA on Inflammatory Markers
2.3.5. Effect of EPA versus DHA on Oxidative Stress
2.3.6. Effect of EPA versus DHA on Glycaemic Control
2.4. Summary of the Effects of EPA versus DHA on Cardiometabolic Risk Factors
3. Discussion
4. Materials and Methods
4.1. Literature Search
4.2. Study Selection
4.3. Publication Bias
4.4. Data Extraction
4.5. Quality Assessment
5. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| AA | arachidonic acid |
| ALA | alpha-linolenic acid |
| ApoA-1 | apolipoprotein A-1 |
| ApoB | apolipoprotein B |
| ApoC3 | apolipoprotein C3 |
| CETP | cholesteryl ester transfer protein |
| ChREBP | carbohydrate responsive element-binding protein |
| CD14 | cluster of differentiation 14 |
| CRP | C-reactive protein |
| COX | cyclooxygenase |
| DHA | docosahexaenoic acid |
| DPA | docosapentaenoic acid |
| EPA | eicosapentaenoic acid |
| FOX-O1 | forkhead box-O transcription factor O1 |
| HbA1c | haemoglobin A1c |
| HDL | high-density lipoprotein |
| IDL | intermediate-density lipoprotein |
| IL-6 | interleukin-6 |
| IL-18 | interleukin-18 |
| LDL | low-density lipoprotein |
| PAF | platelet-activating factor |
| PGI2 | prostaglandin I2 |
| PPARA | peroxisome proliferator-activated receptor alpha |
| PUFA | polyunsaturated fatty acid |
| RCT | randomised controlled trial |
| TNF-α | tumour necrosis factor-alpha |
| TRAF3 | tumour necrosis factor receptor associated factor 3 |
| TVR | total vascular resistance |
| TXA2 | thromboxane A2 |
| TXA3 | thromboxane A3 |
| VLDL | very-low-density lipoprotein |
References
- British Nutrition Foundation. Task Force Report: Cardiovascular Disease: Diet, Nutrition and Emerging Risk Factors; Blackwell: Oxford, UK, 2005. [Google Scholar]
- Calder, P.C. N-3 fatty acids and cardiovascular disease: Evidence explained and mechanisms explored. Clin. Sci. 2004, 107, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Norell, S.E.; Ahlbom, A.; Feychting, M.; Pedersen, N.L. Fish consumption and mortality from coronary heart disease. Br. Med. J. 1986, 293, 426. [Google Scholar] [CrossRef]
- Daviglus, M.; Stamler, J.; Orencia, A.; Morris, D.; Shekelle, R.B. Fish consumption and the 30-year risk of fatal myocardial infarction. N. Engl. J. Med. 1997, 336, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Very long-chain n-3 fatty acids and human health: Fact, fiction and the future. Proc. Nutr. Soc. 2018, 52–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Harris, W.S.; Chung, M.; Lichtenstein, A.H.; Balk, E.M.; Kupelnick, B.; Jordan, H.S.; Lau, J. N-3 Fatty acids from fish or fish-oil supplements, but not alpha-linolenic acid, benefit cardiovascular disease outcomes in primary- and secondary-prevention studies: A systematic review. Am. J. Clin. Nutr. 2006, 84, 5–17. [Google Scholar] [PubMed]
- Studer, M.; Briel, M.; Leimenstoll, B.; Glass, T.R.; Bucher, H.C. Effect of different antilipidemic agents and diets on mortality: A systematic review. Arch. Intern. Med. 2005, 165, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Bucher, H.C.; Hengstler, P.; Schindler, C.; Meier, G. N-3 polyunsaturated fatty acids in coronary heart disease: A meta-analysis of randomized controlled trials. Am. J. Med. 2002, 112, 298–304. [Google Scholar] [CrossRef]
- Manuelli, M.; Della Guardia, L.; Cena, H. Enriching diet with n-3 PUFAs to help prevent cardiovascular diseases in healthy adults: Results from clinical trials. Int. J. Mol. Sci. 2017, 18, 1552. [Google Scholar] [CrossRef] [PubMed]
- AbuMweis, S.; Jew, S.; Tayyem, R.; Agraib, L. Eicosapentaenoic acid and docosahexaenoic acid containing supplements modulate risk factors for cardiovascular disease: A meta-analysis of randomised placebo-control human clinical trials. J. Hum. Nutr. Diet. 2018, 31, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Balk, E.M.; Lichtenstein, A.H.; Chung, M.; Kupelnick, B.; Chew, P.; Lau, J. Effects of omega-3 fatty acids on serum markers of cardiovascular disease risk: A systematic review. Atherosclerosis 2006, 189, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S. N-3 fatty acids and serum lipoproteins: Human studies. Am. J. Clin. Nutr. 1997, 65, 1645S–1654S. [Google Scholar] [CrossRef] [PubMed]
- Suzukawa, M.; Abbey, M.; Howe, P.; Nestel, P. Effects of fish oil fatty acids on low density lipoprotein size, oxidizability, and uptake by macrophages. J. Lipid Res. 1995, 36, 473–484. [Google Scholar] [PubMed]
- Eslick, G.D.; Howe, P.R.C.; Smith, C.; Priest, R.; Bensoussan, A. Benefits of fish oil supplementation in hyperlipidemia: A systematic review and meta-analysis. Int. J. Cardiol. 2009, 136, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Leslie, M.A.; Cohen, D.J.A.; Liddle, D.M.; Robinson, L.E.; Ma, D.W.L. A review of the effect of omega-3 polyunsaturated fatty acids on blood triacylglycerol levels in normolipidemic and borderline hyperlipidemic individuals. Lipids Health Dis. 2015, 14, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Neff, L.; Culiner, J. Algal docosahexaenoic acid affects plasma lipoprotein particle size distribution in overweight and obese adults. J. Nutr. 2011, 141, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.E.; Van Elswyk, M.; Alexander, D.D. Long-chain omega-3 fatty acids eicosapentaenoic acid and docosahexaenoic acid and blood pressure: A meta-analysis of randomized controlled trials. Am. J. Hypertens. 2014, 27, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Geelen, A.; Brouwer, I.A.; Geleijnse, J.M.; Zock, P.L.; Katan, M.B. Effect of fish oil on heart rate in humans: A meta-analysis of randomized controlled trials. Circulation 2005, 112, 1945–1952. [Google Scholar] [CrossRef] [PubMed]
- Von Schacky, C.; Fischer, S.; Weber, P.C. Long-term effects of dietary marine omega-3 fatty-acids upon plasma and cellular lipids, platelet-function, and eicosanoid formation in humans. J. Clin. Investig. 1985, 76, 1626–1631. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Huang, T.; Zheng, J.; Wu, K.; Li, D. Effect of marine-derived n-3 polyunsaturated fatty acids on C-reactive protein, interleukin 6 and tumor necrosis factor alpha: A meta-analysis. PLoS ONE 2014, 9, 1–28. [Google Scholar] [CrossRef]
- Thies, F.; Garry, J.M.C.; Yaqoob, P.; Rerkasem, K.; Williams, J.; Shearman, C.P.; Gallagher, P.J.; Calder, P.C.; Grimble, R.F. Mechanisms of disease association of n-3 polyunsaturated fatty acids with stability of atherosclerotic plaques: A randomised controlled trial. Lancet 2003, 361, 477–485. [Google Scholar] [CrossRef]
- Mori, T.; Woodman, R. The independent effects of eicosapentaenoic acid and docosahexaenoic acid on cardiovascular risk factors in humans. Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Cottin, S.C.; Sanders, T.A.; Hall, W.L. The differential effects of EPA and DHA on cardiovascular risk factors. Proc. Nutr. Soc. 2011, 70, 215–231. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.Y.; Jacobson, T.A. Effects of eicosapentaenoic acid versus docosahexaenoic acid on serum lipids: A systematic review and meta-analysis. Curr. Atheroscler. Rep. 2011, 13, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, T.A.; Glickstein, S.B.; Rowe, J.D.; Soni, P.N. Effects of eicosapentaenoic acid and docosahexaenoic acid on low-density lipoprotein cholesterol and other lipids: A review. J. Clin. Lipidol. 2012, 6, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J. (N-3) fatty acids and cardiovascular health: Are effects of EPA and DHA shared or complementary? J. Nutr. 2012, 142, 614S–625S. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Groot, R. Effects of omega-3 long chain polyunsaturated fatty acid supplementation on cardiovascular mortality: The importance of the dose of DHA. Nutrients 2017, 9, 1305. [Google Scholar] [CrossRef] [PubMed]
- Mesa, M.D.; Buckley, R.; Minihane, A.M.; Yaqoob, P. Effects of oils rich in eicosapentaenoic and docosahexaenoic acids on the oxidizability and thrombogenicity of low-density lipoprotein. Atherosclerosis 2004, 175, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Agah, S.; Shidfar, F.; Khandouzi, N.; Baghestani, A.R.; Hosseini, S. Comparison of the effects of eicosapentaenoic acid with docosahexaenoic acid on the level of serum lipoproteins in helicobacter pylori: A randomized clinical trial. Iran. Red Crescent Med. J. 2015, 17, e17652. [Google Scholar] [CrossRef] [PubMed]
- Asztalos, I.B.; Gleason, J.A.; Sever, S.; Gedik, R.; Asztalos, B.F.; Horvath, K.V.; Dansinger, M.L.; Lamon-Fava, S.; Schaefer, E.J. Effects of eicosapentaenoic acid and docosahexaenoic acid on cardiovascular disease risk factors: A randomized clinical trial. Metabolism 2016, 65, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Rontoyanni, V.G.; Hall, W.L.; Pombo-Rodrigues, S.; Appleton, A.; Chung, R.; Sanders, T.A.B. A comparison of the changes in cardiac output and systemic vascular resistance during exercise following high-fat meals containing DHA or EPA. Br. J. Nutr. 2012, 108, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Buckley, R.; Shewring, B.; Turner, R.; Yaqoob, P.; Minihane, A.M. Circulating triacylglycerol and apoE levels in response to EPA and docosahexaenoic acid supplementation in adult human subjects. Br. J. Nutr. 2004, 92, 477. [Google Scholar] [CrossRef] [PubMed]
- Olano-Martin, E.; Anil, E.; Caslake, M.J.; Packard, C.J.; Bedford, D.; Stewart, G.; Peiris, D.; Williams, C.M.; Minihane, A.M. Contribution of apolipoprotein E genotype and docosahexaenoic acid to the LDL-cholesterol response to fish oil. Atherosclerosis 2010, 209, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Rambjør, G.S.; Wlen, A.I.; Windsor, S.L.; Harris, W.S. Eicosapentaenoic acid is primarily responsible for hypotriglyceridemic effect of fish oil in humans. Lipids 1996, 31, 45–49. [Google Scholar] [CrossRef]
- Phang, M.; Sinclair, A.J.; Lincz, L.F.; Garg, M.L. Gender-specific inhibition of platelet aggregation following omega-3 fatty acid supplementation. Nutr. Metab. Cardiovasc. Dis. 2010, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Cottin, S.C.; Alsaleh, A.; Sanders, T.A.B.; Hall, W.L. Lack of effect of supplementation with EPA or DHA on platelet-monocyte aggregates and vascular function in healthy men. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Phang, M.; Scorgie, F.E.; Seldon, M.; Garg, M.L.; Lincz, L.F. Reduction of prothrombin and Factor V levels following supplementation with omega-3 fatty acids is sex dependent: A randomised controlled study. J. Nutr. Biochem. 2014, 25, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Kew, S.; Mesa, M.D.; Tricon, S.; Buckley, R.; Minihane, A.M.; Yaqoob, P. Effects of oils rich in eicosapentaenoic and docosahexaenoic acids on immune cell composition and function in healthy humans. Am. J. Clin. Nutr. 2004, 79, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Kannenberg, F.; Somoza, V.; Erbersdobler, H.F.; Wahrburg, U. Dietary alpha-linolenic acid, EPA, and DHA have differential effects on LDL fatty acid composition but similar effects on serum lipid profiles in normolipidemic humans. J. Nutr. 2009, 139, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Rassoul, F.; Boesch-Saadatmandi, C.; Richter, V.; Rimbach, G.; Erbersdobler, H.; Somoza, V.; Wahrburg, U. Effects of controlled diets enriched with alpha-linolenic acid, eicosapentaenoic acid or docosahexaenoic acid on soluble adhesion molecules and endothelin-1 concentrations in healthy volunteers. Curr. Top. Nutraceutical Res. 2007, 5, 189–195. [Google Scholar]
- Egert, S.; Fobker, M.; Andersen, G.; Somoza, V.; Erbersdobler, H.F.; Wahrburg, U. Effects of dietary alpha-linolenic acid, eicosapentaenoic acid or docosahexaenoic acid on parameters of glucose metabolism in healthy volunteers. Ann. Nutr. Metab. 2008, 53, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.-B.; Grimsgaard, S.; Nilsen, H.; Nordøy, A.; Bønaa, K.H. Effects of highly purified eicosapentaenoic acid and docosahexaenoic acid on fatty acid absorption, incorporation into serum phospholipids and postprandial triglyceridemia. Lipids 1998, 33, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Somoza, V.; Kannenberg, F.; Fobker, M.; Krome, K.; Erbersdobler, H.F.; Wahrburg, U. Influence of three rapeseed oil-rich diets, fortified with alpha-linolenic acid, eicosapentaenoic acid or docosahexaenoic acid on the composition and oxidizability of low-density lipoproteins: Results of a controlled study in healthy volunteers. Eur. J. Clin. Nutr. 2007, 61, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Von Schacky, C.; Weber, P.C. Metabolism and effects on platelet function of the purified eicosapentaenoic and docosahexaenoic acids in humans. J. Clin. Investig. 1985, 76, 2446–2450. [Google Scholar] [CrossRef] [PubMed]
- McManus, S.; Tejera, N.; Awwad, K.; Vauzour, D.; Rigby, N.; Fleming, I.; Cassidy, A.; Minihane, A. Differential effects of EPA versus DHA on postprandial vascular function and the plasma oxylipin profile in men. J. Lipid Res. 2016, 57, 1720–1727. [Google Scholar] [CrossRef] [PubMed]
- Bønaa, K.H.; Bjerve, K.S.; Nordøy, A. Docosahexaenoic and eicosapentaenoic acids in plasma phospholipids are divergently associated with high density lipoprotein in humans. Arterioscler. Thromb. 1992, 12, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Du Plooy, W.; Venter, C.; Muntingh, G.; Venter, H.; Glatthaar, I.; Smith, K. The cumulative dose response effect of eicosapentaenoic and docosahexaenoic acid on blood pressure, plasma lipid profile and diet pattern in mild to moderate essential hypertensive black patients. Prostaglandins Leukot. Essent. Fat. Acids 1992, 46, 315–321. [Google Scholar] [CrossRef]
- Grimsgaard, S.; Bonna, K.H.; Hansen, J.-B.; Nordøy, A. Highly purified eicosapentaenoic acid and docosahexaenoic acid in humans have similar triacylglycerol-lowering effects but divergent effects on serum fatty acids. Am. J. Clin. Nutr. 1997, 66, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Bao, D.; Burke, V.; Puddey, I.; Beilin, L. Docosahexaenoic acid but not eicosapentaenoic acid lowers ambulatory blood pressure and heart rate in humans. Hypertension 1999, 34, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed]
- Allaire, J.; Couture, P.; Leclerc, M.; Charest, A.; Marin, J.; Lépine, M.-C.; Talbot, D.; Tchernof, A.; Lamarche, B. A randomized, crossover, head-to-head comparison of eicosapentaenoic acid and docosahexaenoic acid supplementation to reduce inflammation markers in men and women: The Comparing EPA to DHA (ComparED) Study. Am. J. Clin. Nutr. 2016, 104, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Allaire, J.; Harris, W.S.; Vors, C.; Charest, A.; Marin, J.; Jackson, K.H.; Tchernof, A.; Couture, P.; Lamarche, B. Supplementation with high-dose docosahexaenoic acid increases the omega-3 Index more than high-dose eicosapentaenoic acid. Prostaglandins Leukot. Essent. Fat. Acids 2017, 120, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Vors, C.; Allaire, J.; Marin, J.; Lepine, M.C.; Charest, A.; Tchernof, A.; Couture, P.; Lamarche, B. Inflammatory gene expression in whole blood cells after EPA vs. DHA supplementation: Results from the ComparED study. Atherosclerosis 2017, 257, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Grimsgaard, S.; Bonaa, K.; Hansen, J.; Myhre, E. Effects of highly purified eicosapentaenoic acid and docosahexaenoic acid on hemodynamics in humans. Am. J. Clin. Nutr. 1998, 68, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Watts, G.F.; Burke, V.; Hilme, E.; Puddey, I.B.; Beilin, L.J. Differential effects of eicosapentaenoic acid and docosahexaenoic acid on vascular reactivity of the forearm microcirculation in hyperlipidaemic, overweight men. Circulation 2000, 102, 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Burke, V.; Puddey, I.B.; Watts, G.F.; O’Neal, D.N.; Best, J.D.; Beilin, L.J. Purified eicosapentaenoic and docosahexaenoic acids have differential effects on serum lipids and lipoproteins, LDL particle size, glucose, and insulin in mildly hyperlipidemic men. Am. J. Clin. Nutr. 2000, 71, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Puddey, I.B.; Burke, V.; Croft, K.D.; Dunstan, D.W.; Rivera, J.H.; Beilin, L.J. Effect of omega 3 fatty acids on oxidative stress in humans: GC-MS measurement of urinary F-2-isoprostane excretion. Redox Rep. 2000, 5, 45–46. [Google Scholar] [CrossRef] [PubMed]
- Mas, E.; Woodman, R.J.; Burke, V.; Puddey, I.B.; Beilin, L.J.; Durand, T.; Mori, T. The omega-3 fatty acids EPA and DHA decrease plasma F(2)-isoprostanes: Results from two placebo-controlled interventions. Free Radic. Res. 2010, 44, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Nestel, P.; Shige, H.; Pomeroy, S.; Cehun, M.; Abbey, M.; Raederstorff, D. The n-3 fatty acids eicosapentaenoic acid and docosahexaenoic acid increase systemic arterial compliance in humans. Am. J. Clin. Nutr. 2002, 76, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Harris, W. EPA, but not DHA, decreases mean platelet volume in normal subjects. Lipids 2002, 37, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Harris, W.S. Omega-3 fatty acid supplementation accelerates chylomicron triglyceride clearance. J. Lipid Res. 2003, 44, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Jones, P.G.; Harris, W.S. Triacylglycerol-rich lipoprotein margination: A potential surrogate for whole-body lipoprotein lipase activity and effects of eicosapentaenoic and docosahexaenoic acids. Am. J. Clin. Nutr. 2004, 80, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Woodman, R.J.; Mori, T.A.; Burke, V.; Puddey, I.B.; Watts, G.F.; Beilin, L.J. Effects of purified eicosapentaenoic and docosahexaenoic acids on glycemic control, blood pressure, and serum lipids in type 2 diabetic patients with treated hypertension. Am. J. Clin. Nutr. 2002, 76, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Woodman, R.J.; Mori, T.A.; Burke, V.; Puddey, I.B.; Barden, A.; Watts, G.F.; Beilin, L.J. Effects of purified eicosapentaenoic acid and docosahexaenoic acid on platelet, fibrinolytic and vascular function in hypertensive type 2 diabetic patients. Atherosclerosis 2003, 166, 85–93. [Google Scholar] [CrossRef]
- Woodman, R.; Mori, T.; Burke, V.; Puddey, I.; Watts, G.; Best, J.; Beilin, L. Docosahexaenoic acid but not eicosapentaenoic acid increases LDL particle size in treated hypertensive type 2 diabetic patients. Diabetes Care 2003, 26, 253. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Woodman, R.J.; Burke, V.; Puddey, I.B.; Croft, K.D.; Beilin, L.J. Effect of eicosapentaenoic acid and docosahexaenoic acid on oxidative stress and inflammatory markers in treated-hypertensive type 2 diabetic subjects. Free Radic. Biol. Med. 2003, 35, 772–781. [Google Scholar] [CrossRef]
- Higgins, J.; Green, S. (Eds.) Cochrane Handbook for Systematic Reviews of Interventions Version 5.1.0; The Cochrane Collaboration: London, UK, 2011. [Google Scholar]
- Calder, P.C. Functional roles of fatty acids and their effects on human health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S. The omega-3 index as a risk factor for coronary heart disease. Am. J. Clin. Nutr. 2008, 87, 1997S–2002S. [Google Scholar] [PubMed]
- Harris, W.S.; Sands, S.A.; Windsor, S.L.; Ali, H.A.; Stevens, T.L.; Magalski, A.; Porter, C.B.; Borkon, A.M. Omega-3 fatty acids in cardiac biopsies from heart transplantation patients: Correlation with erythrocytes and response to supplementation. Circulation 2004, 110, 1645–1649. [Google Scholar] [CrossRef] [PubMed]
- Block, R.C.; Harris, W.S.; Reid, K.J.; Sands, S.A.; Spertus, J.A. EPA and DHA in blood cell membranes from acute coronary syndrome patients and controls. Atherosclerosis 2008, 197, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Monge, A.; Harris, W.S.; Ortiz-Panozo, E.; Yunes, E.; Cantu-Brito, C.; Catzin-Kuhlmann, A.; Lopez-Ridaura, R.; Lajous, M. Whole blood n-3 fatty acids are inversely associated with carotid intima-media thickness in indigenous Mexican women. J. Nutr. 2016, 146, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Von Schacky, C. The Omega-3 Index: A new risk factor for death from coronary heart disease? Prev. Med. 2004, 39, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Del Gobbo, L.; Tintle, N.L. The Omega-3 Index and relative risk for coronary heart disease mortality: Estimation from 10 cohort studies. Atherosclerosis 2017, 262, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Kleber, M.E.; Delgado, G.E.; Lorkowski, S.; März, W.; von Schacky, C. Omega-3 fatty acids and mortality in patients referred for coronary angiography. The Ludwigshafen Risk and Cardiovascular Health Study. Atherosclerosis 2016, 252, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Shearer, G.C.; Savinova, O.V.; Harris, W.S. Fish oil—How does it reduce plasma triglycerides? Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2012, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M.H. Omega-3 fatty acids: New insights into the pharmacology and biology of docosahexaenoic acid, docosapentaenoic acid, and eicosapentaenoic acid. Curr. Opin. Lipidol. 2013, 24, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Allaire, J.; Vors, C.; Couture, P.; Lamarche, B. LDL particle number and size and cardiovascular risk. Curr. Opin. Lipidol. 2017, 28, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.D.; Holub, B.J. Differential eicosapentaenoic acid elevations and altered cardiovascular disease risk factor responses after supplementation with docosahexaenoic acid in postmenopausal women receiving and not receiving hormone replacement therapy. Am. J. Clin. Nutr. 2004, 79, 765–773. [Google Scholar] [CrossRef] [PubMed]
- McLennan, P.L. Cardiac physiology and clinical efficacy of dietary fish oil clarified through cellular mechanisms of omega-3 polyunsaturated fatty acids. Eur. J. Appl. Physiol. 2014, 114, 1333–1356. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Huerta, O.D.; Aguilera, C.M.; Mesa, M.D.; Gil, A. Omega-3 long-chain polyunsaturated fatty acids supplementation on inflammatory biomakers: A systematic review of randomised clinical trials. Br. J. Nutr. 2012, 107, S159–S170. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.; Petasis, N. Resolvins and protectins in inflammation resolution. Chem. Rev. 2011, 111, 5922–5943. [Google Scholar] [CrossRef] [PubMed]
- Nenseter, M.; Drevon, C. Dietary polyunsaturates and peroxidation of low density lipoprotein. Curr. Opin. Lipidol. 1996, 7, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Witztum, J.L. Atherosclerosis: The road ahead. Cell 2001, 104, 503–516. [Google Scholar] [CrossRef]
- Halvorsen, D.S.; Hansen, J.B.; Grimsgaard, S.; Bønaa, K.H.; Kierulf, P.; Nordøy, A. The effect of highly purified eicosapentaenoic and docosahexaenoic acids on monocyte phagocytosis in man. Lipids 1997, 32, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Conquer, J.A.; Holub, B.J. Dietary docosahexaenoic acid as a source of eicosapentaenoic acid in vegetarians and omnivores. Lipids 1997, 32, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Plourde, M.; Chouinard-Watkins, R.; Vandal, M.; Zhang, Y.; Lawrence, P.; Brenna, J.T.; Cunnane, S.C. Plasma incorporation, apparent retroconversion and β-oxidation of 13C-docosahexaenoic acid in the elderly. Nutr. Metab. 2011, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jadad, A.R.; Moore, R.A.; Carroll, D.; Jenkinson, C.; Reynolds, D.J.M.; Gavaghan, D.J.; McQuay, H.J. Assessing the quality of reports of randomized clinical trials: Is blinding necessary? Control. Clin. Trials 1996, 17, 1–12. [Google Scholar] [CrossRef]


{kind=link}
| Reference | Study Design & Population | Sample Size (n) | Dose of EPA (g/Day) | Duration (Weeks) | Outcomes | Jadad Score |
|---|---|---|---|---|---|---|
| Dose of DHA (g/Day) | ||||||
| Dose of Placebo (g/Day) | ||||||
| Allaire et al., 2016, 2017 [52,53], Vors et al., 2017 [54] | Double-blind randomised controlled crossover study with 9 week washout. Healthy subjects with abdominal obesity and low-grade inflammation | n = 121 (EPA) n = 123 (DHA) n = 125 (corn oil) n = 125 (total) | 2.7 (EPA)
2.7 (DHA) 3.0 (corn oil) | 10 | Inflammation markers (IL-6, IL-18, CRP, TNF-α, adiponectin)
Inflammatory gene expression (PPARA, TNFA, CD14, TRAF3, CCL2, IL-10, IL-1B, IL-1RN, NFKB, TNFRSF1A) Blood lipids (total cholesterol, LDL cholesterol, HDL cholesterol, ApoB, triglycerides) | 5 |
| Grimsgaard et al., 1997, 1998 [49,55] | Double-blind parallel RCT.
Healthy men | n = 75 (EPA) n = 72 (DHA) n = 77 (corn oil) n = 224 (total) | 3.8 (EPA) 3.6 (DHA) 4.0 (corn oil) | 7 | Blood lipids (total cholesterol, LDL cholesterol, HDL cholesterol, ApoA1, ApoB, triglycerides) Haemodynamics (blood pressure, heart rate, left ventricular function) | 5 |
| Mori et al., 1999, 2000a, 2000b, 2000c [50,56,57,58], Mas et al., 2010 [59] | Double-blind parallel RCT. Overweight mildly hyperlipidaemic men | n = 19 (EPA) n = 17 (DHA) n = 20 (olive oil) n = 56 (total) | 3.8 (EPA) 3.7 (DHA) 3.0 (olive oil) | 6 | Blood lipids (total cholesterol, LDL cholesterol, HDL cholesterol, triglycerides) Oxidative stress markers (urinary and plasma F2-isoprostanes) Glycaemic control (fasting insulin, fasting glucose) Haemodynamics (heart rate, blood pressure, endothelial function) | 3 |
| Nestel et al., 2002 [60] | Double-blind parallel RCT. Dyslipidaemic subjects | n = 12 (EPA) n = 12 (DHA) n = 14 (olive oil) n = 38 (total) | 3.0 (EPA) 2.8 (DHA) 2.8 (olive oil) | 7 | Arterial function (systemic arterial compliance) Blood lipids (Total cholesterol, LDL cholesterol, HDL cholesterol, VLDL triglyceride, total triglyceride) Haemodynamics (heart rate, blood pressure, pulse pressure, total vascular resistance) | 4 |
| Park & Harris 2002, 2003, 2004 [61,62,63] | Double-blind parallel RCT with 4 week run-in (olive oil) followed by 4 week wash-out. Healthy subjects | n = 11 (EPA) n = 11 (DHA) n = 11 (safflower oil) n = 33 (total) | 3.8 (EPA) 3.8 (DHA) 4.0 (safflower oil) | 4 | Blood lipids (total cholesterol, HDL cholesterol, LDL cholesterol, VLDL cholesterol, triglycerides, plasma phospholipids, chylomicron triglycerides, chylomicron size, ApoB-48, ApoB-100, margination volume) Platelet function (mean platelet volume) | 3 |
| Woodman et al., 2002, 2003a, 2003b [64,65,66], Mori et al., 2003 [67], Mas et al., 2010 [59] | Double-blind parallel RCT. Treated hypertensive Type 2 diabetics | n = 17 (EPA) n = 18 (DHA) n = 16 (olive oil) n = 51 (total) | 3.8 (EPA) 3.7 (DHA) 3.0 (olive oil) | 6 | Oxidative stress markers (urinary and plasma F2-isoprostanes) Blood lipids (total cholesterol, LDL cholesterol, HDL cholesterol, triglycerides, LDL particle size) Haemodynamics (blood pressure) Glycaemic control (fasting glucose, glycated haemoglobin, fasting insulin, C-peptide, insulin sensitivity) Inflammation markers (TNF-α, CRP, IL-6) Platelet, fibrinolytic and vascular function (collagen and PAF-stimulated platelet aggregation, collagen-stimulated thromboxane release, plasma tPA & PAI-1 antigen, von Willebrand factor, P-selectin, brachial artery dilatation) | 3 |
| Study | Selection Bias | Performance Bias | Detection Bias | Attrition Bias | Reporting Bias | Other Bias |
|---|---|---|---|---|---|---|
| Allaire et al., 2016, 2017 [52,53], Vors et al., 2017 [54] | ||||||
| Grimsgaard et al., 1997, 1998 [49,55] | ||||||
| Mori et al., 1999, 2000a, 2000b, 2000c [50,56,57,58] 1, Mas et al., 2010 [59] 1 | ||||||
| Nestel et al., 2002 [60] 2 | ||||||
| Park & Harris 2002, 2003, 2004 [61,62,63] 1 | ||||||
| Woodman et al., 2002, 2003a, 2003b [64,65,66] 1, Mori et al., 2003 [67] 1, Mas et al., 2010 [59] 1 |
| Study | Population | Control | Effect of EPA vs. Control on Blood Lipids and Lipoproteins | Effect of DHA vs. Control on Blood Lipids and Lipoproteins | Effect of EPA vs. DHA on Blood Lipids and Lipoproteins |
|---|---|---|---|---|---|
| Allaire et al., 2016 [52] | Healthy subjects with abdominal obesity and low-grade inflammation | Corn oil | ↓ Triglycerides (−12%,
p < 0.0001) ↑ LDL cholesterol (+2%, p = 0.046) | ↓ Triglycerides (−13%,
p < 0.0001) ↑ Total cholesterol (+4%, p = 0.001) ↑ LDL cholesterol (+7%, p < 0.0001) ↑ HDL cholesterol (+8%, p < 0.0001) ↓ Cholesterol/HDL cholesterol ratio (−3%, p < 0.001) ↑ ApoB (+5%, p = 0.02) | Compared to EPA, DHA resulted in a greater: ↓ Triglycerides (p = 0.005) ↑ Total cholesterol (p < 0.001) ↑ LDL cholesterol (p = 0.04)—more so in men than women (p = 0.046) ↑ HDL cholesterol (p < 0.0001) ↓ Cholesterol/HDL cholesterol ratio (p = 0.006) |
| Grimsgaard et al., 1997 [49] | Healthy men | Corn oil | ↓ Triglycerides (−21%,
p = 0.0001) ↓ Total cholesterol (−0.15 ± 0.55 mmol/L, p < 0.05) ↓ ApoA-1 (−0.04 ± 0.10 g/L, p < 0.001) ↓ ApoB (−0.03 ± 0.11 g/L, p < 0.05) ↑ HDL:ApoA-1 (+0.04 ± 0.08, p = 0.0001) ↓ Total:HDL cholesterol (−0.13 ± 0.47, p = 0.007) | ↓ Triglycerides (−26%,
p = 0.0001) ↑ HDL cholesterol (+0.06 ± 0.13 mmol/L, p < 0.001) ↑ HDL:ApoA-1 (+0.04 ± 0.07, p < 0.001) ↓ Total/HDL cholesterol (−0.19 ± 0.52, p < 0.01) | Compared to EPA, DHA resulted in greater: ↑ HDL cholesterol (p = 0.009) Non-statistically significant lowering of triglycerides (p = 0.14) |
| Mori et al., 2000b [57] | Overweight mildly hyperlipidaemic men | Olive oil | ↓ Triglycerides (−18%,
p = 0.012) ↓ HDL3 cholesterol (−7%, p = 0.032) No significant difference in total cholesterol | ↓ Triglycerides (−20%,
p = 0.003) ↑ LDL cholesterol (+8%, p = 0.019) ↑ LDL particle size (+0.25 ± 0.08 nm, p = 0.002) ↑ HDL2 cholesterol (+29%, p = 0.004) No significant difference in total cholesterol | N/A |
| Nestel et al., 2002 [60] | Dyslipidaemic subjects | Olive oil | ↓ Total triglycerides (p = 0.026) ↓ VLDL triglycerides (p = 0.006) No significant difference in total or LDL cholesterol | ↓ Total triglycerides (p = 0.026) ↓ VLDL triglycerides (p = 0.006) No significant difference in total or LDL cholesterol | No significant difference between EPA and DHA |
| Park & Harris 2003, Park et al., 2004 [62,63] | Healthy subjects | Safflower oil | Results for EPA and DHA similar, so authors reported as one group: No significant effect on blood lipids (triglycerides, total, LDL-, HDL- or VLDL cholesterol) ↓ Apo B-48 (−28%, p < 0.001) ↓ Apo B-100 (−24%, p < 0.01) ↓ Chylomicron triglyceride half-lives (fed state) (p < 0.05) ↓ Chylomicron particle size (p < 0.01) ↑ Pre-heparin lipoprotein lipase (p < 0.05) ↑ Margination volumes in the fasted state (p < 0.001) ↑ Margination volumes in the fed state (DHA only; p < 0.05) | No significant difference between EPA and DHA | |
| Woodman et al., 2002, 2003b [64,66] | Hypertensive-treated Type 2 diabetics | Olive oil | ↓ Triglycerides (−19%,
p = 0.022) ↑ HDL2 cholesterol (+16%, p = 0.026) ↓ HDL3 cholesterol (−11%, p = 0.026) No significant difference in total, LDL- or HDL cholesterol | ↓ Triglycerides (−15%,
p = 0.022) ↑ HDL2 cholesterol (+12%, p = 0.05) ↑ LDL particle size (+0.26 ± 0.10 nm, p = 0.02) No significant difference in total, LDL- or HDL cholesterol | N/A |
| Study | Population | Control | Effect of EPA vs. Control on Haemodynamics | Effect of DHA vs. Control on Haemodynamics | Effect of EPA vs. DHA on Haemodynamics |
|---|---|---|---|---|---|
| Grimsgaard et al., 1998 [55] | Healthy men | Corn oil | ↑ Heart rate (increased 1.9 bpm, p = 0.04) Improved left ventricular diastolic filling No significant effect on blood pressure | ↓ Heart rate (decreased 2.2 bpm, p = 0.006) Improved left ventricular diastolic filling No significant effect on blood pressure | Compared to EPA, DHA resulted in: ↓ Heart rate (p = 0.0001) |
| Mori et al., 1999, 2000a [50,56] | Overweight mildly hyperlipidaemic men | Olive oil | No significant effect on blood pressure. Small nonsignificant rise in heart rate. | ↓ 24 h (5.8/3.3 mm Hg) and daytime (3.5/2.0 mm Hg) ambulatory systolic and diastolic blood pressure (p < 0.05) ↓ 24 h (decreased 3.5 bpm), daytime (decreased 3.7 bpm), nighttime (decreased 2.8 bpm) ambulatory heart rate (p = 0.001) Increased vasodilator responses and attenuation of constrictor responses in forearm blood flow | N/A |
| Nestel et al., 2002 [60] | Dyslipidaemic subjects | Olive oil | ↑ Systemic arterial compliance (+36%, p = 0.028) Nonsignificant lowering of pulse pressure and vascular resistance. No significant difference in heart rate, blood pressure, pulse pressure, or total vascular resistance. | ↑ Systemic arterial compliance (+27%, p = 0.091) Nonsignificant lowering of pulse pressure and vascular resistance. No significant difference in heart rate, blood pressure, pulse pressure or total vascular resistance. | No significant difference between EPA and DHA |
| Woodman et al., 2002, 2003a [64,65] | Hypertensive-treated Type 2 diabetics | Olive oil | No significant difference in blood pressure Nonsignificant decrease in 24 h heart rate No significant difference in vascular function | No significant difference in blood pressure Non-significant decrease in 24 h heart rate No significant difference in vascular function | N/A |
| Study | Population | Control | Effect of EPA vs. Control on Platelet and Fibrinolytic Function | Effect of DHA vs. Control on Platelet and Fibrinolytic Function | Effect of EPA vs. DHA on Platelet and Fibrinolytic Function |
|---|---|---|---|---|---|
| Park & Harris 2002 [61] | Healthy subjects | Safflower oil | ↓ Mean platelet volume ↓ Platelet count | No effect | N/A |
| Woodman et al., 2003 [65] | Hypertensive-treated Type 2 diabetics | Olive oil | Platelet function: No effect on collagen-stimulated platelet aggregation or platelet-derived TXB2 Fibrinolytic function: No effect on PAI-1 antigen, tPA antigen, von Willebrand factor, or P-selectin. | Platelet function: ↓ Collagen-stimulated platelet aggregation (−17%, p = 0.054) ↓ Platelet-derived TXB2 (−19%, p = 0.03) No effect on PAF-stimulated platelet aggregation. Fibrinolytic function: No effect on PAI-1 antigen, tPA antigen, von Willebrand factor, or P-selectin. | N/A |
| Study | Population | Control | Effect of EPA vs. Control on Inflammatory Markers | Effect of DHA vs. Control on Inflammatory Markers | Effect of EPA vs. DHA on Inflammatory Markers |
|---|---|---|---|---|---|
| Allaire et al., 2016 [52], Vors et al., 2017 [54] | Healthy subjects with abdominal obesity and low-grade inflammation | Corn oil | ↓ IL-6 (−13%, p = 0.03) No significant effect on IL-18, CRP, TNF-α or adiponectin ↓ CD14 gene expression (p = 0.008) ↑ PPARA gene expression (p = 0.003) ↑ TRAF3 gene expression (p = 0.002) | ↓ IL-6 (−12%, p = 0.01) ↓ IL-18 (−7%, p = 0.002) ↓ CRP (−8%, p = 0.02) ↓ TNF-α (−15%, p = 0.01) ↑ Adiponectin (+3%, p = 0.047) ↓ CD14 gene expression (p = 0.02) ↑ PPARA gene expression (p = 0.01) ↑ TNFA gene expression (p = 0.01) | Compared to EPA, DHA resulted in greater: ↓ IL-18 (p = 0.01) ↑ Adiponectin (<0.001) No significant difference between EPA and DHA for change in IL-6, CRP, TNF-α or gene expression |
| Mori et al., 2003 [67] | Hypertensive-treated Type 2 diabetics | Olive oil | No significant change in IL-6 and CRP Nonsignificant trend for lowered TNF-α (−19.5%, n.s.) | No significant change in IL-6 & CRP Nonsignificant trend for lowered TNF-α (−32.8%, n.s.) | N/A |
| Study | Population | Control | Effect of EPA vs. Control on Oxidative Stress | Effect of DHA vs. Control on Oxidative Stress | Effect of EPA vs. DHA on Oxidative Stress |
|---|---|---|---|---|---|
| Mori et al., 2000 [58], Mas et al., 2010 [59] | Overweight mildly hyperlipidaemic men | Olive oil | ↓ Urinary F2 isoprostanes (−27%, p < 0.0001) ↓ Plasma F2 isoprostanes (−24%, p < 0.0001) | ↓ Urinary F2 isoprostanes (−26%, p < 0.0001) ↓ Plasma F2 isoprostanes (−14%, p = 0.009) | N/A |
| Mori et al., 2003 [67] Mas et al., 2010 [59] | Hypertensive-treated Type 2 diabetics | Olive oil | ↓ Urinary F2 isoprostanes (−19%, p = 0.017) ↓ Plasma F2 isoprostanes (s19%, p = 0.039) | ↓ Urinary F2 isoprostanes (−20%, p = 0.014) ↓ Plasma F2 isoprostanes (−23%, p = 0.011) | N/A |
| Study | Population | Control | Effect of EPA vs. Control on Blood Glucose Control | Effect of DHA vs. Control on Blood Glucose Control | Effect of EPA vs. DHA on Blood Glucose Control |
|---|---|---|---|---|---|
| Mori et al., 2000b [57] | Overweight mildly hyperlipidaemic men | Olive oil | ↑ Fasting insulin (+18%, p = 0.035) Trend towards increased fasting glucose (+4%, p = 0.062) | ↑ Fasting insulin (+27%, p = 0.001) ↓ Glucose to insulin ratio (p = 0.018) | N/A |
| Woodman et al., 2002 [64] | Hypertensive-treated Type II diabetics | Olive oil | ↑ Fasting glucose (p = 0.002) No effect on glycated haemoglobin, fasting insulin, fasting C-peptide, insulin sensitivity or secretion. | ↑ Fasting glucose (p = 0.002) No effect on glycated haemoglobin, fasting insulin, fasting C-peptide, insulin sensitivity or secretion. | N/A |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Innes, J.K.; Calder, P.C. The Differential Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on Cardiometabolic Risk Factors: A Systematic Review. Int. J. Mol. Sci. 2018, 19, 532. https://doi.org/10.3390/ijms19020532
Innes JK, Calder PC. The Differential Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on Cardiometabolic Risk Factors: A Systematic Review. International Journal of Molecular Sciences. 2018; 19(2):532. https://doi.org/10.3390/ijms19020532
Chicago/Turabian StyleInnes, Jacqueline K., and Philip C. Calder. 2018. "The Differential Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on Cardiometabolic Risk Factors: A Systematic Review" International Journal of Molecular Sciences 19, no. 2: 532. https://doi.org/10.3390/ijms19020532
APA StyleInnes, J. K., & Calder, P. C. (2018). The Differential Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on Cardiometabolic Risk Factors: A Systematic Review. International Journal of Molecular Sciences, 19(2), 532. https://doi.org/10.3390/ijms19020532