Involvement of Gastrin-Releasing Peptide Receptor in the Regulation of Adipocyte Differentiation in 3T3-L1 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
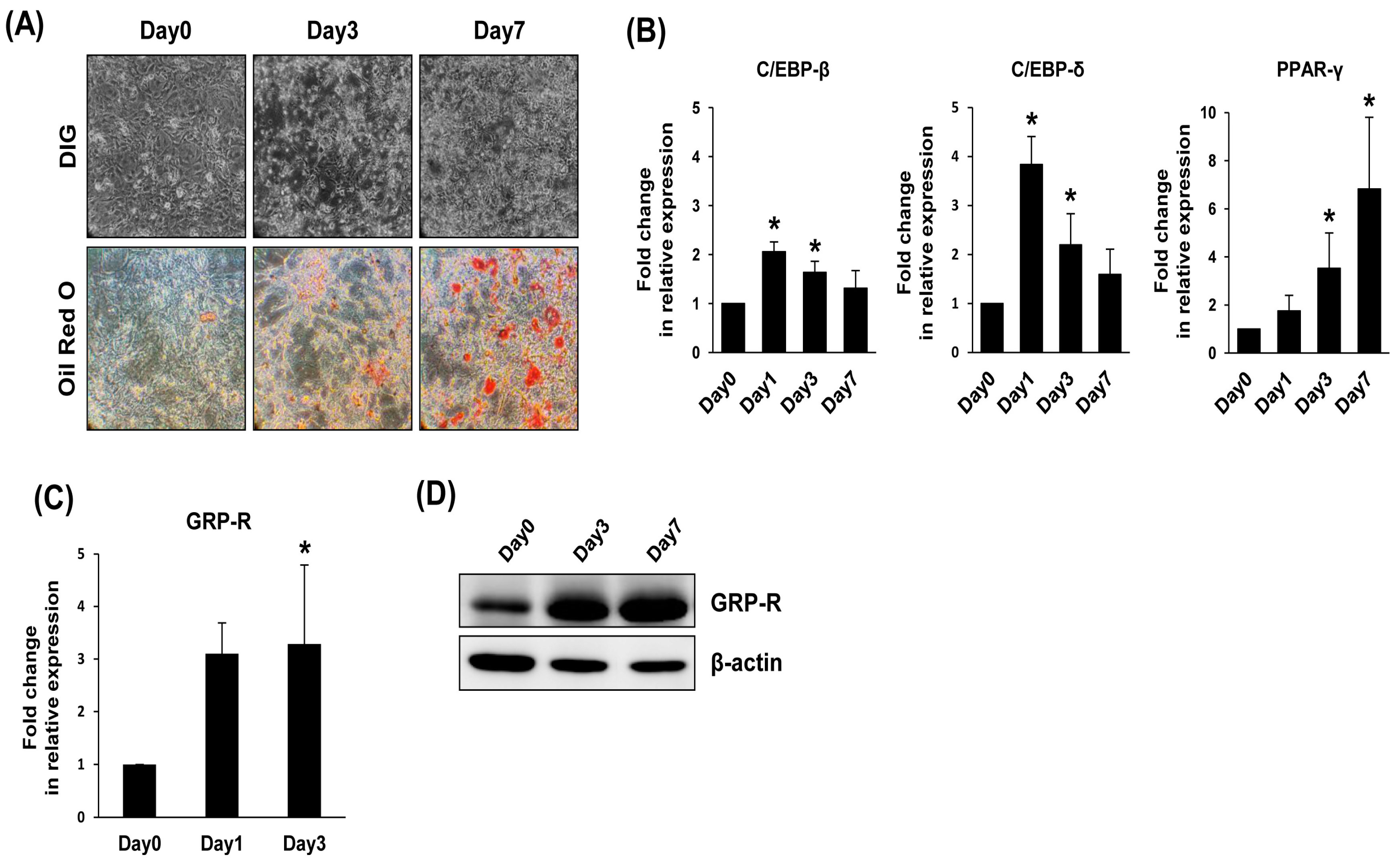
2.1. GRP-R Expression Is Increased during Adipocyte Differentiation
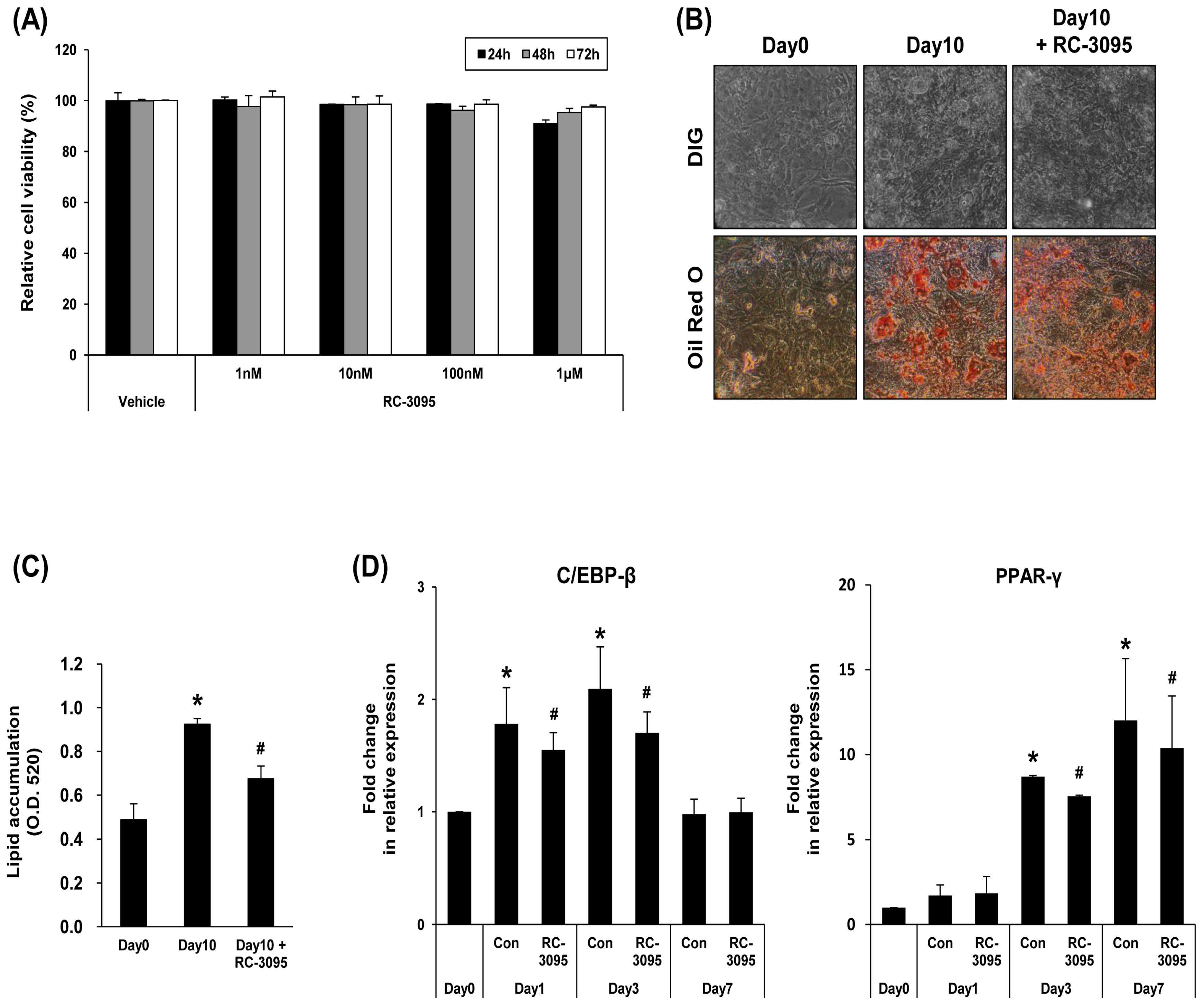
2.2. GRP-R Antagonist Modulates Adipocyte Differentiation
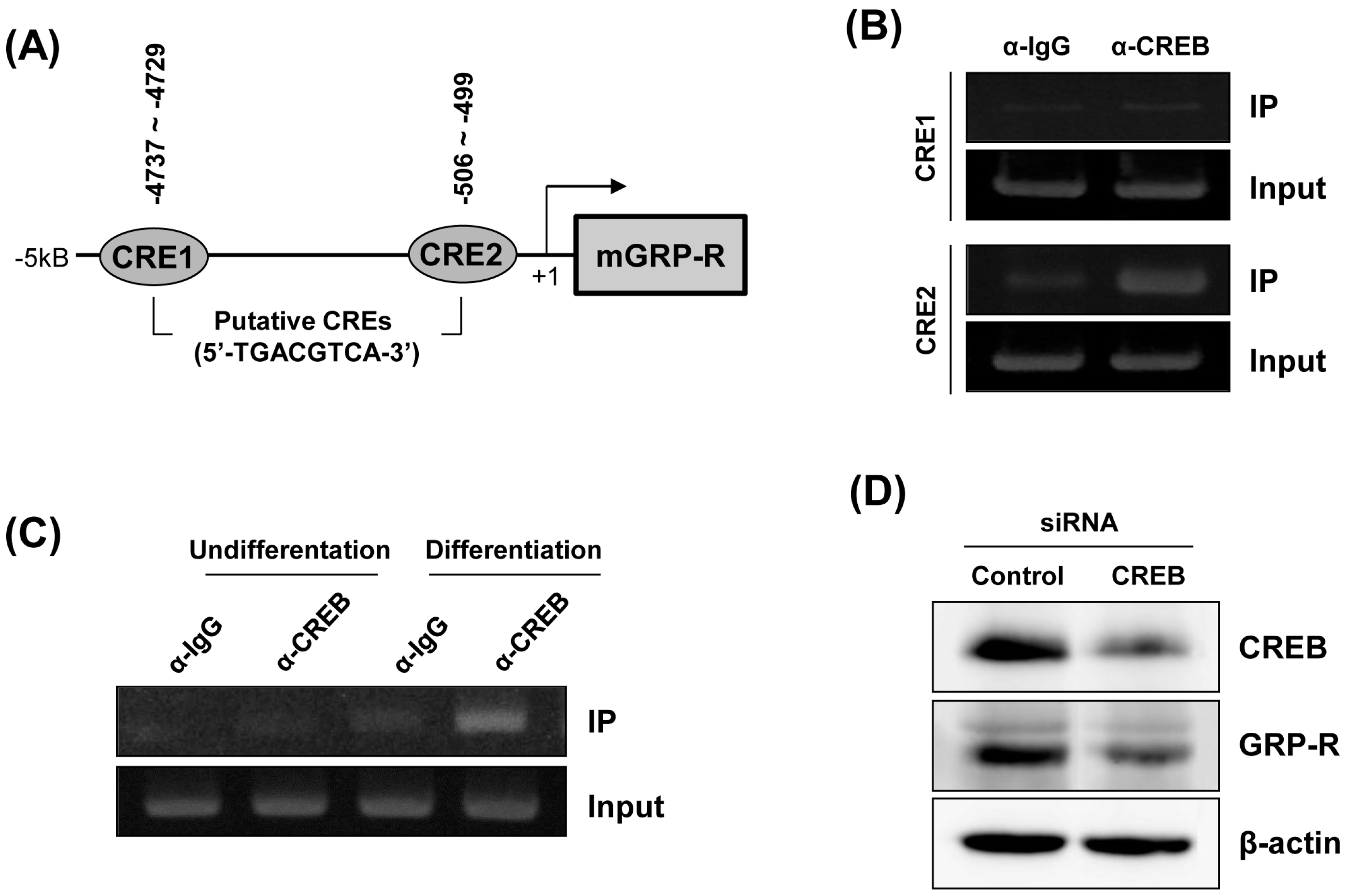
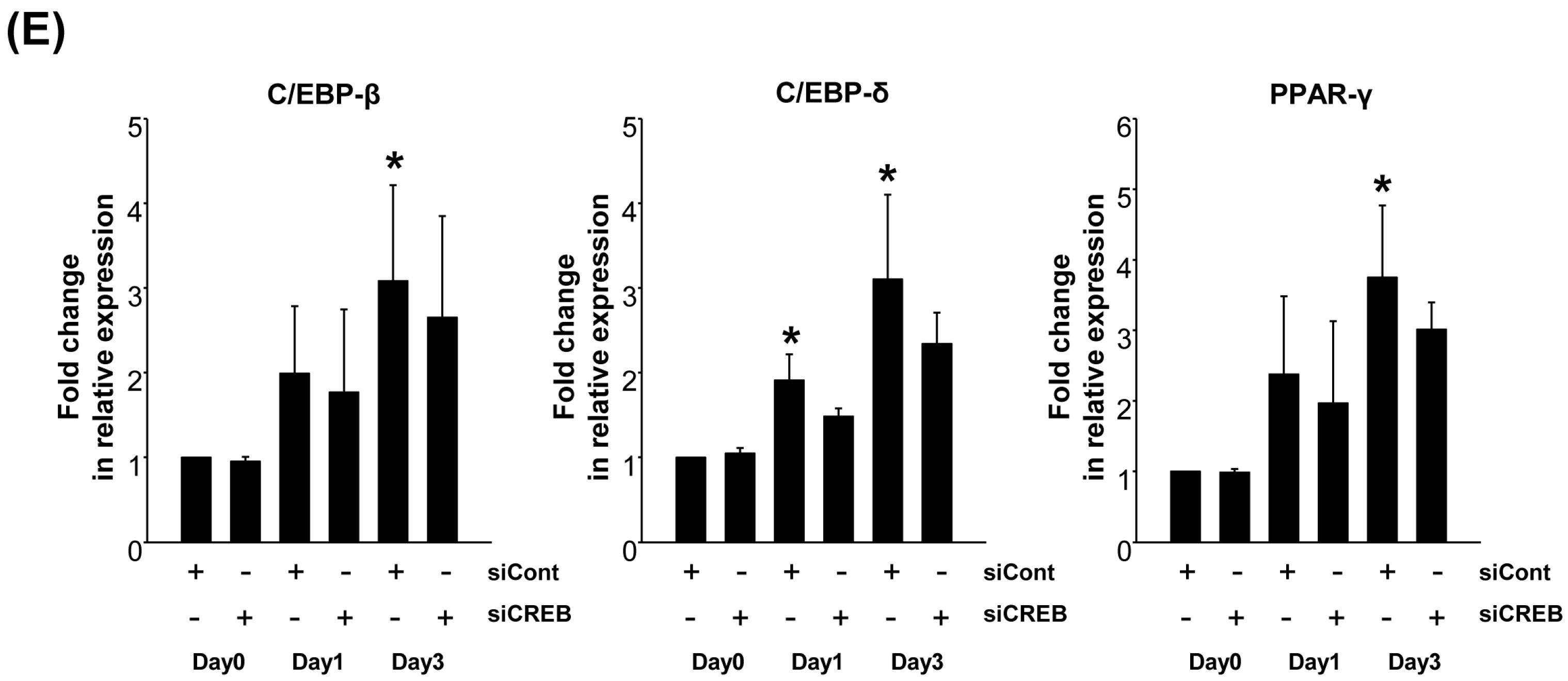
2.3. Downregulation of GRP-R by CREB Knockdown Inhibits the Adipocyte Differentiation of 3T3-L1 Cells
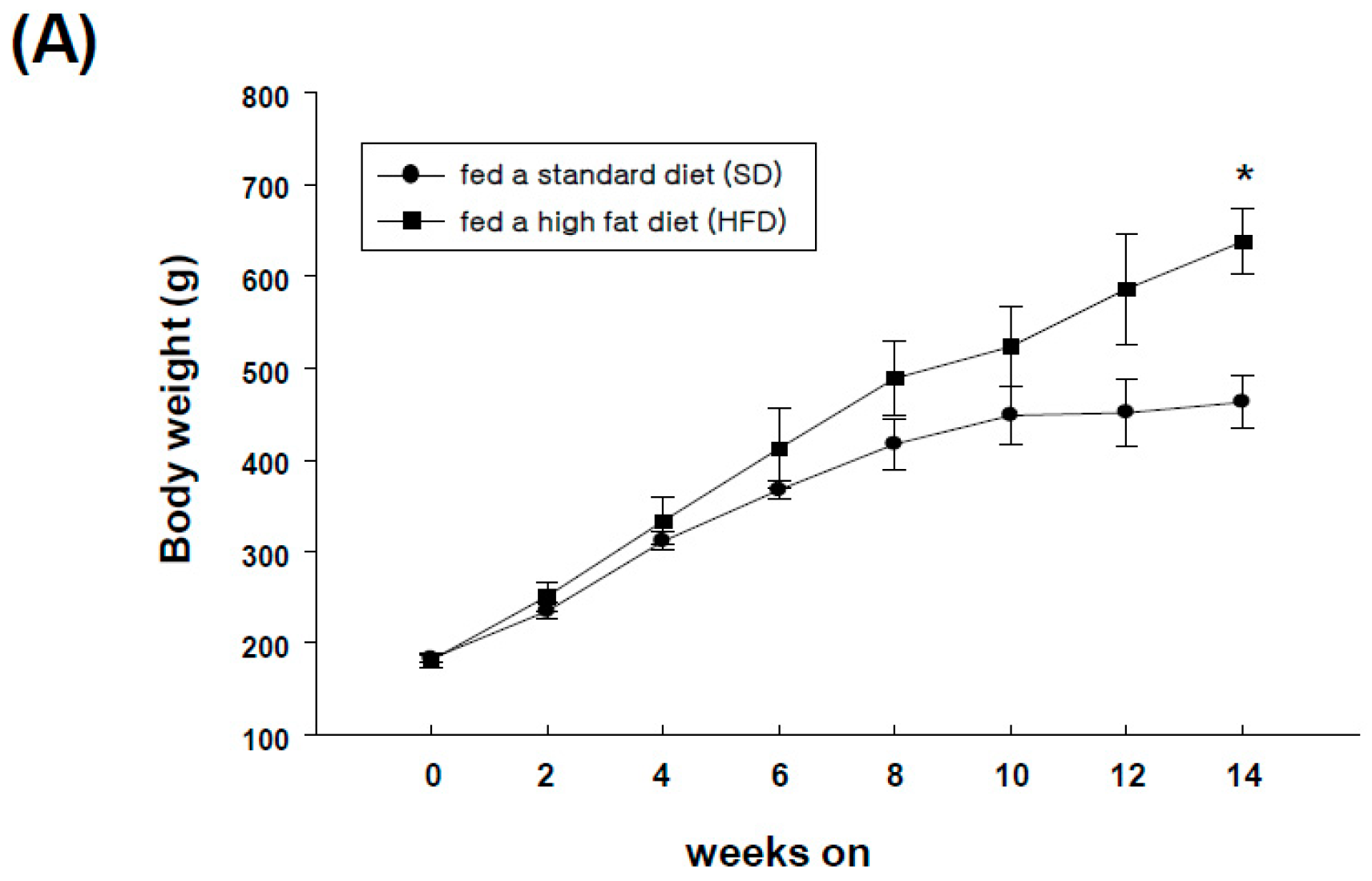
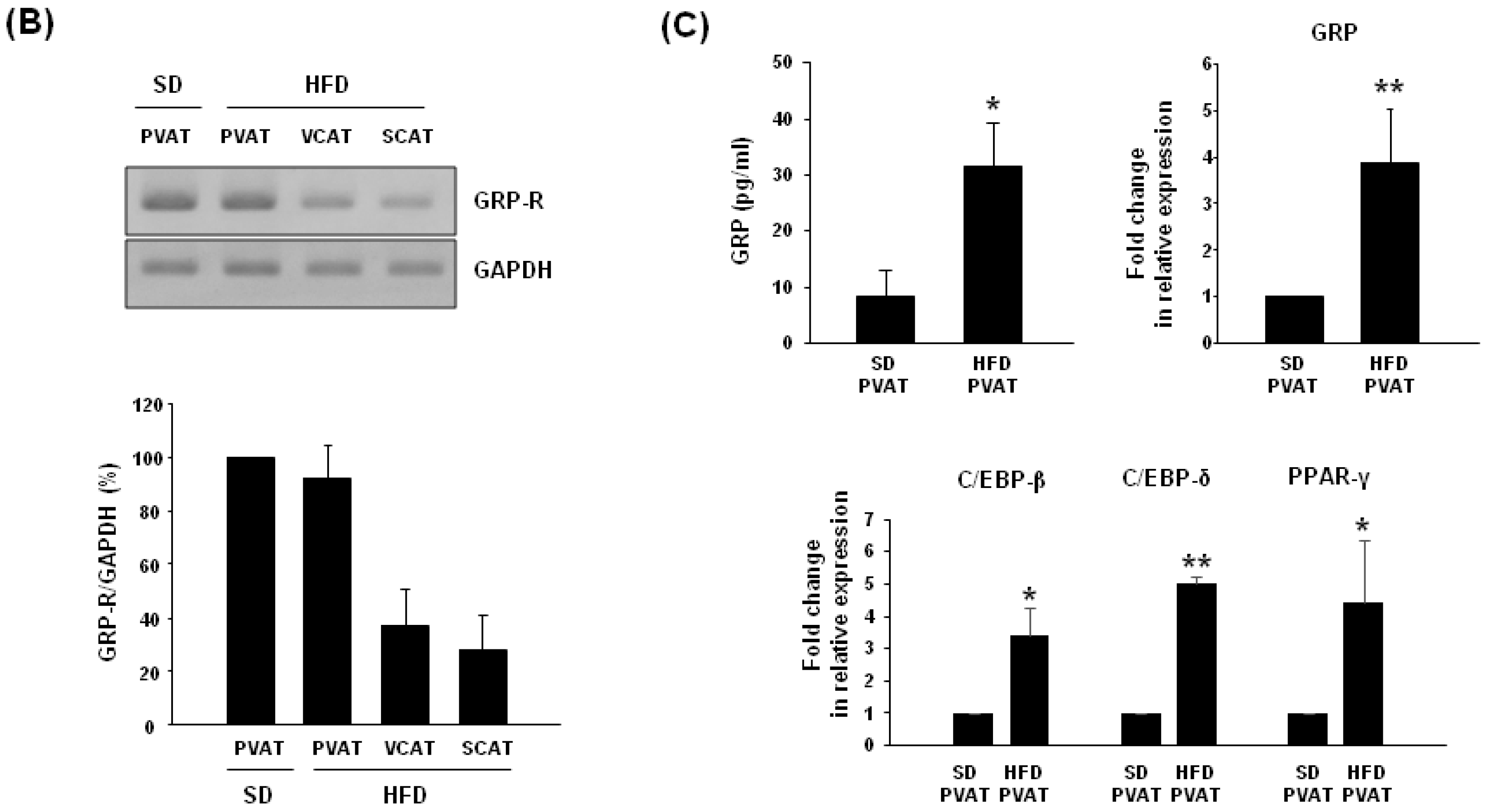
2.4. Expression of GRP-R and GRP in Adipose Tissues of High-Fat Diet-Induced Obese Rats
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Oil Red O Staining
4.4. MTT Assay
4.5. Western Blot Analysis
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. RT-PCR and Real-Time PCR Analysis
4.8. Chromatin Immunoprecipitation (ChIP) Assay
4.9. RNA Interference Experiment
4.10. High-Fat Diet (HFD)-Induced Obese Model and Experimental Protocols
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GRP | Gastrin-releasing peptide |
| GRP-R | Gastrin-releasing peptide receptor |
| CREB | Cyclic AMP response element binding protein |
| C/EBPs | CCAAT/enhancer binding proteins |
| PPAR-γ | Peroxisome proliferator-activated receptor-γ |
References
- Majumdar, I.D.; Weber, H.C. Biology of Mammalian Bombesin-Like Peptides and their Receptors. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Wang, H.; Liu, R. Recent Insights into Biological Functions of Mammalian Bombesin-Like Peptides and their Receptors. Curr. Opin. Endocrinol. Diabetes Obes. 2018, 25, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.C. Regulation and Signaling of Human Bombesin Receptors and their Biological Effects. Curr. Opin. Endocrinol. Diabetes Obes. 2009, 16, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lv, Y.; Yuan, A.; Li, Z. Gastrin-Releasing Peptide Links Stressor to Cancer Progression. J. Cancer Res. Clin. Oncol. 2010, 136, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Petronilho, F.; Danielski, L.G.; Roesler, R.; Schwartsmann, G.; Dal-Pizzol, F. Gastrin-Releasing Peptide as a Molecular Target for Inflammatory Diseases: An Update. Inflamm. Allergy Drug Targets 2013, 12, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, N.; Moody, T.W.; Igarashi, H.; Ito, T.; Jensen, R.T. Bombesin-Related Peptides and their Receptors: Recent Advances in their Role in Physiology and Disease States. Curr. Opin. Endocrinol. Diabetes Obes. 2008, 15, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Ischia, J.; Patel, O.; Shulkes, A.; Baldwin, G.S. Gastrin-Releasing Peptide: Different Forms, Different Functions. Biofactors 2009, 35, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Wang, J.; Hampton, L.L.; Weber, H.C. The Human Gastrin-Releasing Peptide Receptor Gene Structure, Its Tissue Expression and Promoter. Gene 2001, 264, 95–103. [Google Scholar] [CrossRef]
- Yang, Y.S.; Song, H.D.; Li, R.Y.; Zhou, L.B.; Zhu, Z.D.; Hu, R.M.; Han, Z.G.; Chen, J.L. The Gene Expression Profiling of Human Visceral Adipose Tissue and Its Secretory Functions. Biochem. Biophys. Res. Commun. 2003, 300, 839–846. [Google Scholar] [CrossRef]
- Hoggard, N.; Bashir, S.; Cruickshank, M.; Miller, J.D.; Speakman, J.R. Expression of Neuromedin B in Adipose Tissue and Its Regulation by Changes in Energy Balance. J. Mol. Endocrinol. 2007, 39, 199–210. [Google Scholar] [CrossRef]
- Farmer, S.R. Transcriptional Control of Adipocyte Formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.G.; Grespan, R.; Pinto, L.G.; Meurer, L.; Brenol, J.C.; Roesler, R.; Schwartsmann, G.; Cunha, F.Q.; Xavier, R.M. Protective Effect of RC-3095, an Antagonist of the Gastrin-Releasing Peptide Receptor, in Experimental Arthritis. Arthritis Rheumatol. 2011, 63, 2956–2965. [Google Scholar] [CrossRef] [PubMed]
- Chinnappan, D.; Qu, X.; Xiao, D.; Ratnasari, A.; Weber, H.C. Human Gastrin-Releasing Peptide Receptor Gene Regulation Requires Transcription Factor Binding at Two Distinct CRE Sites. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G153–G162. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.C.; Jensen, R.T.; Battey, J.F. Molecular Organization of the Mouse Gastrin-Releasing Peptide Receptor Gene and Its Promoter. Gene 2000, 244, 137–149. [Google Scholar] [CrossRef]
- Qu, X.; Xiao, D.; Weber, H.C. Transcriptional Activation of the Human Gastrin-Releasing Peptide Receptor Gene in Gastrointestinal and Prostatic Epithelial Cancer Cells. J. Mol. Neurosci. 2004, 22, 75–82. [Google Scholar] [CrossRef]
- Xiao, D.; Wang, J.; Hampton, L.L.; Weber, H.C. The Human Gastrin-Releasing Peptide Receptor Gene Structure, Its Tissue Expression and Promoter. Gene 2001, 264, 95–103. [Google Scholar] [CrossRef]
- Zhang, J.W.; Klemm, D.J.; Vinson, C.; Lane, M.D. Role of CREB in Transcriptional Regulation of CCAAT/Enhancer-Binding Protein Beta Gene during Adipogenesis. J. Biol. Chem. 2004, 279, 4471–4478. [Google Scholar] [CrossRef]
- Reusch, J.E.; Colton, L.A.; Klemm, D.J. CREB Activation Induces Adipogenesis in 3T3-L1 Cells. Mol. Cell. Biol. 2000, 20, 1008–1020. [Google Scholar] [CrossRef]
- Lee, M.J.; Wu, Y.; Fried, S.K. Adipose Tissue Remodeling in Pathophysiology of Obesity. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 371–376. [Google Scholar] [CrossRef]
- Lago, F.; Dieguez, C.; Gomez-Reino, J.; Gualillo, O. Adipokines as Emerging Mediators of Immune Response and Inflammation. Nat. Clin. Pract. Rheumatol. 2007, 3, 716–724. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators Linking Adipose Tissue, Inflammation and Immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Szasz, T.; Webb, R.C. Perivascular Adipose Tissue: More than just Structural Support. Clin. Sci. 2012, 122, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rajsheker, S.; Manka, D.; Blomkalns, A.L.; Chatterjee, T.K.; Stoll, L.L.; Weintraub, N.L. Crosstalk between Perivascular Adipose Tissue and Blood Vessels. Curr. Opin. Pharmacol. 2010, 10, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Despres, J.P.; Koh, K.K. Perivascular Adipose Tissue in the Pathogenesis of Cardiovascular Disease. Atherosclerosis 2013, 230, 177–184. [Google Scholar] [CrossRef]
- Nosalski, R.; Guzik, T.J. Perivascular Adipose Tissue Inflammation in Vascular Disease. Br. J. Pharmacol. 2017, 174, 3496–3513. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Park, H.J.; Kim, Y.; Kim, H.J.; Bae, S.K.; Bae, M.K. Gastrin-Releasing Peptide Induces Monocyte Adhesion to Vascular Endothelium by Upregulating Endothelial Adhesion Molecules. Biochem. Biophys. Res. Commun. 2017, 485, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, M.K.; Kim, Y.; Bae, S.S.; Kim, H.J.; Bae, S.K.; Bae, M.K. Gastrin-Releasing Peptide Promotes the Migration of Vascular Smooth Muscle Cells through Upregulation of Matrix Metalloproteinase-2 and -9. BMB Rep. 2017, 50, 628–633. [Google Scholar] [CrossRef]
- Rittig, K.; Dolderer, J.H.; Balletshofer, B.; Machann, J.; Schick, F.; Meile, T.; Kuper, M.; Stock, U.A.; Staiger, H.; Machicao, F.; et al. The Secretion Pattern of Perivascular Fat Cells is Different from that of Subcutaneous and Visceral Fat Cells. Diabetologia 2012, 55, 1514–1525. [Google Scholar] [CrossRef]
- Szasz, T.; Bomfim, G.F.; Webb, R.C. The Influence of Perivascular Adipose Tissue on Vascular Homeostasis. Vasc. Health Risk Manag. 2013, 9, 105–116. [Google Scholar] [CrossRef]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-K.; Park, H.-J.; Kim, Y.; Bae, S.-K.; Kim, H.J.; Bae, M.-K. Involvement of Gastrin-Releasing Peptide Receptor in the Regulation of Adipocyte Differentiation in 3T3-L1 Cells. Int. J. Mol. Sci. 2018, 19, 3971. https://doi.org/10.3390/ijms19123971
Kim M-K, Park H-J, Kim Y, Bae S-K, Kim HJ, Bae M-K. Involvement of Gastrin-Releasing Peptide Receptor in the Regulation of Adipocyte Differentiation in 3T3-L1 Cells. International Journal of Molecular Sciences. 2018; 19(12):3971. https://doi.org/10.3390/ijms19123971
Chicago/Turabian StyleKim, Mi-Kyoung, Hyun-Joo Park, Yeon Kim, Soo-Kyung Bae, Hyung Joon Kim, and Moon-Kyoung Bae. 2018. "Involvement of Gastrin-Releasing Peptide Receptor in the Regulation of Adipocyte Differentiation in 3T3-L1 Cells" International Journal of Molecular Sciences 19, no. 12: 3971. https://doi.org/10.3390/ijms19123971
APA StyleKim, M.-K., Park, H.-J., Kim, Y., Bae, S.-K., Kim, H. J., & Bae, M.-K. (2018). Involvement of Gastrin-Releasing Peptide Receptor in the Regulation of Adipocyte Differentiation in 3T3-L1 Cells. International Journal of Molecular Sciences, 19(12), 3971. https://doi.org/10.3390/ijms19123971