Characterization and Expression Profiling of Neuropeptides and G-Protein-Coupled Receptors (GPCRs) for Neuropeptides in the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae)



Abstract

1. Introduction
2. Results
2.1. Sequencing and Unigene Assembly
2.2. Neuropeptide and Peptide Hormone Genes
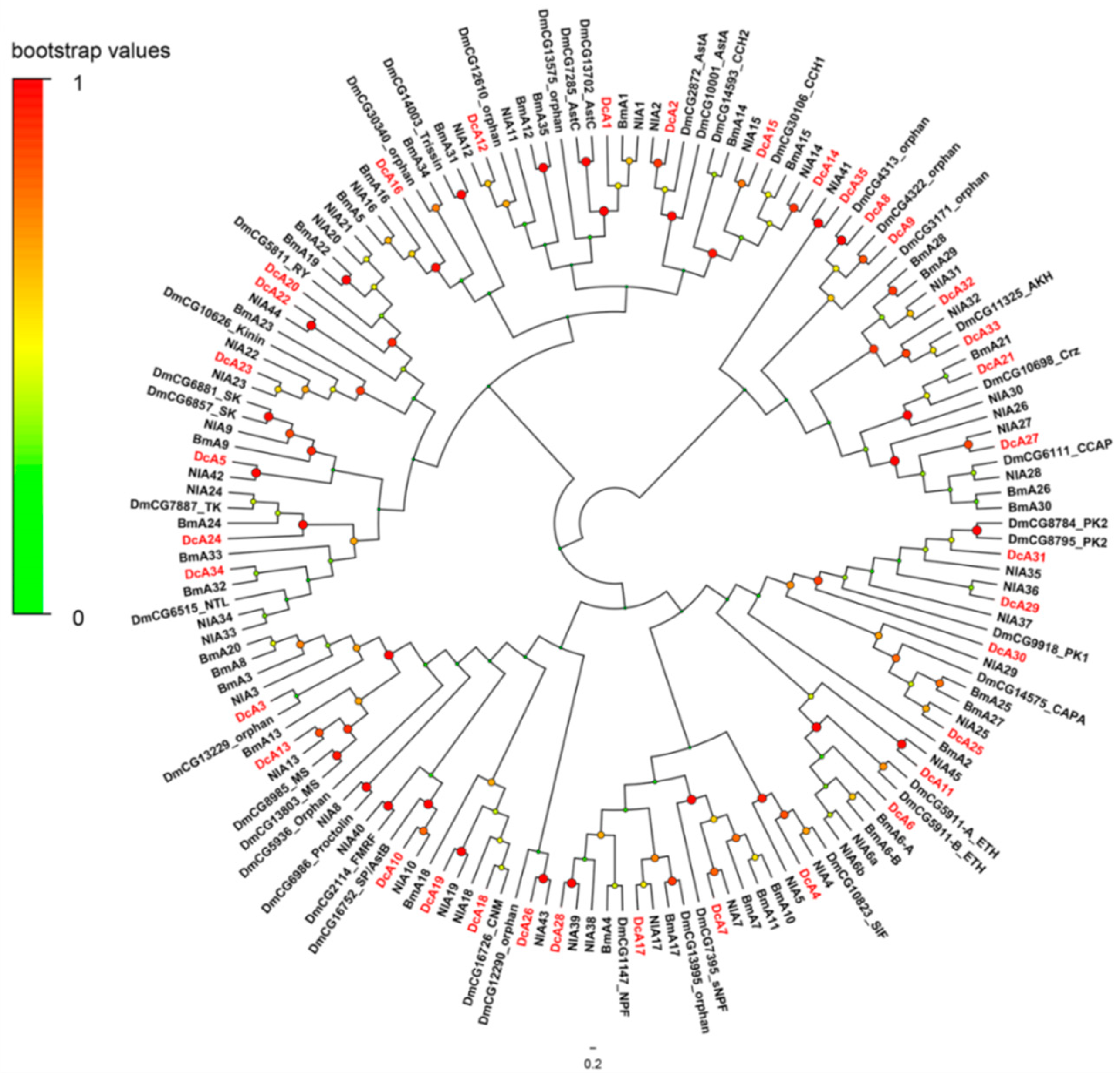
2.3. G-Protein-Coupled Receptors (GPCRs) for Neuropeptides
2.3.1. A-Family GPCRs
2.3.2. B-Family GPCRs
2.3.3. Leucine-Rich Repeat-Containing GPCRs (LGRs)
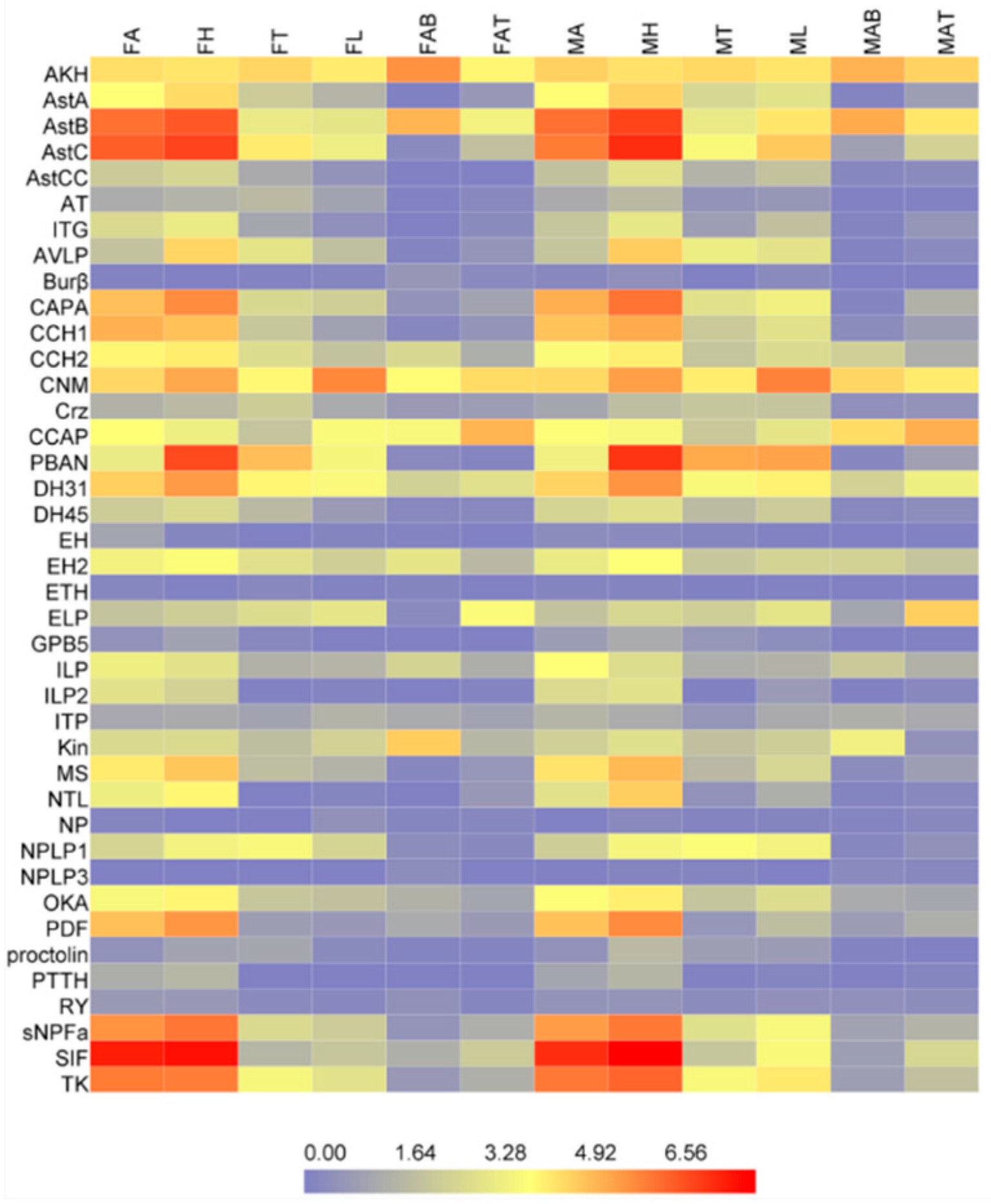
2.4. Tissue-Specific Expression Profiles of the Neuropeptides and Neuropeptide Receptors
2.5. Developmental Stages Expression Analysis by RT-qPCR
3. Discussion
3.1. Neuropeptides Involved in Ecdysis and Development
3.2. Neuropeptides Control of Metabolism
3.3. Reproductive-Related Neuropeptides
3.4. Neuropeptides in Olfaction
4. Materials and Methods
4.1. Insect Rearing and RNA Extraction
4.2. RNA-seq
4.3. Identification of the Neuropeptides and Their Putative G-Protein-Coupled Receptors in D. citri
4.4. Structure and Domain Analysis
4.5. Phylogenetic Analysis
4.6. Gene Expression Profiling of the Neuropeptides and Neuropeptide Receptors
4.7. Developmental Stages Expression Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Menn, J.J.; Borkovec, A.B. Insect neuropeptides: Potential new insect control agents. J. Agric. Food Chem. 1989, 37, 271–278. [Google Scholar] [CrossRef]
- Nässel, D.R.; Winther, Å.M.E. Drosophila neuropeptides in regulation of physiology and behavior. Prog. Neurobiol. 2010, 92, 42–104. [Google Scholar] [CrossRef] [PubMed]
- Hewes, R.S.; Taghert, P.H. Neuropeptides and neuropeptide receptors in the Drosophila melanogaster genome. Genome Res. 2001, 11, 1126–1142. [Google Scholar] [CrossRef] [PubMed]
- Fricker, L.D. Neuropeptide-processing enzymes: Applications for drug discovery. In Drug Addiction: From Basic Research to Therapy; Rapaka, R.S., Sadée, W., Eds.; Springer: New York, NY, USA, 2008; pp. 497–509. ISBN 978-0-387-76678-2. [Google Scholar]
- Zupanc, G.K.H. Peptidergic transmission: From morphological correlates to functional implications. Micron 1996, 27, 35–91. [Google Scholar] [CrossRef]
- Caers, J.; Verlinden, H.; Zels, S.; Vandersmissen, H.P.; Vuerinckx, K.; Schoofs, L. More than two decades of research on insect neuropeptide GPCRs: An overview. Front. Endocrinol. 2012, 3, 151. [Google Scholar] [CrossRef] [PubMed]
- Seinsche, A.; Dyker, H.; Lösel, P.; Backhaus, D.; Scherkenbeck, J. Effect of helicokinins and ACE inhibitors on water balance and development of Heliothis virescens larvae. J. Insect Physiol. 2000, 46, 1423–1431. [Google Scholar] [CrossRef]
- Jung, J.W.; Kim, J.H.; Pfeiffer, R.; Ahn, Y.J.; Page, T.L.; Kwon, H.W. Neuromodulation of olfactory sensitivity in the peripheral olfactory organs of the American cockroach, Periplaneta americana. PLoS ONE 2013, 8, e81361. [Google Scholar] [CrossRef] [PubMed]
- Ignell, R.; Root, C.M.; Birse, R.T.; Wang, J.W.; Nässel, D.R.; Winther, Å.M.E. Presynaptic peptidergic modulation of olfactory receptor neurons in Drosophila. Proc. Natl. Acad. Sci. USA 2009, 106, 13070–13075. [Google Scholar] [CrossRef] [PubMed]
- Verlinden, H.; Vleugels, R.; Zels, S.; Dillen, S.; Lenaerts, C.; Crabbé, K.; Spit, J.; Vanden Broeck, J. Receptors for neuronal or endocrine signalling molecules as potential targets for the control of insect pests. In Advances in Insect Physiology; Cohen, E., Ed.; Academic Press: New York, NY, USA, 2014; pp. 167–303. ISBN 0065-2806. [Google Scholar]
- Ons, S. Neuropeptides in the regulation of Rhodnius prolixus physiology. J. Insect Physiol. 2017, 97, 77–92. [Google Scholar] [CrossRef]
- Étienne, J.; Quilici, S.; Marival, D.; Franck, A. Biological control of Diaphorina citri (Hemiptera: Psyllidae) in Guadeloupe by imported Tamarixia radiata (Hymenoptera: Eulophidae). Fruits 2001, 56, 307–315. [Google Scholar] [CrossRef]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and management of Asian citrus psyllid, vector of the Huanglongbing pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed]
- Halbert, S.E.; Manjunath, K.L. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A literature review and assessment of risk in Florida. Fla. Entomol. 2004, 87, 330–353. [Google Scholar] [CrossRef]
- Manjunath, K.L.; Halbert, S.E.; Ramadugu, C.; Webb, S.; Lee, R.F. Detection of ‘Candidatus Liberibacter asiaticus’ in Diaphorina citri and its importance in the management of Citrus Huanglongbing in Florida. Phytopathology 2008, 98, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Clayson, P.J.; Kuhns, E.H.; Stelinski, L.L. Effects of buprofezin and diflubenzuron on various developmental stages of Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2012, 68, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Sétamou, M.; Rodriguez, D.; Saldana, R.; Schwarzlose, G.; Palrang, D.; Nelson, S.D. Efficacy and uptake of soil-applied imidacloprid in the control of Asian citrus psyllid and a citrus leafminer, two foliar-feeding citrus pests. J. Econ. Entomol. 2010, 103, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Mann, R.S.; Rogers, M.E.; Stelinski, L.L. Insecticide resistance in field populations of Asian citrus psyllid in Florida. Pest Manag. Sci. 2011, 67, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Roller, L.; Yamanaka, N.; Watanabe, K.; Daubnerová, I.; Žitňan, D.; Kataoka, H.; Tanaka, Y. The unique evolution of neuropeptide genes in the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Gu, G.; Teng, Z.; Wu, S.; Huang, J.; Song, Q.; Ye, G.; Fang, Q. Identification and expression profiles of neuropeptides and their G protein-coupled receptors in the rice stem borer Chilo suppressalis. Sci. Rep. 2016, 6, 28976. [Google Scholar] [CrossRef]
- Li, B.; Predel, R.; Neupert, S.; Hauser, F.; Tanaka, Y.; Cazzamali, G.; Williamson, M.; Arakane, Y.; Verleyen, P.; Schoofs, L.; et al. Genomics, transcriptomics, and peptidomics of neuropeptides and protein hormones in the red flour beetle Tribolium castaneum. Genome Res. 2007, 18, 113–122. [Google Scholar] [CrossRef]
- Hummon, A.B.; Richmond, T.A.; Verleyen, P.; Baggerman, G.; Huybrechts, J.; Ewing, M.A.; Vierstraete, E.; Rodriguez-Zas, S.L.; Schoofs, L.; Robinson, G.E.; et al. From the genome to the proteome: Uncovering peptides in the Apis brain. Science 2006, 314, 647–649. [Google Scholar] [CrossRef]
- Tanaka, Y.; Suetsugu, Y.; Yamamoto, K.; Noda, H.; Shinoda, T. Transcriptome analysis of neuropeptides and G-protein coupled receptors (GPCRs) for neuropeptides in the brown planthopper Nilaparvata lugens. Peptides 2014, 53, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Ons, S.; Sterkel, M.; Diambra, L.; Urlaub, H.; Rivera-Pomar, R. Neuropeptide precursor gene discovery in the Chagas disease vector Rhodnius prolixus. Insect Mol. Biol. 2011, 20, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Audsley, N.; Down, R.E. G protein coupled receptors as targets for next generation pesticides. Insect Biochem. Mol. Biol. 2015, 67, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Huybrechts, J.; Bonhomme, J.; Minoli, S.; Prunier-Leterme, N.; Dombrovsky, A.; Abdel-Latief, M.; Robichon, A.; Veenstra, J.A.; Tagu, D. Neuropeptide and neurohormone precursors in the pea aphid, Acyrthosiphon pisum. Insect Mol. Biol. 2010, 19, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Žitňan, D.; Galizia, C.G.; Cho, K.; Adams, M.E. A command chemical triggers an innate behavior by sequential activation of multiple peptidergic ensembles. Curr. Biol. 2006, 16, 1395–1407. [Google Scholar] [CrossRef] [PubMed]
- Zitnan, D.; Adams, M.E. Neuroendocrine regulation of ecdysis. In Insect Endocrinology; Gilbert, L.I., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 253–309. ISBN 978-0-12-384749-2. [Google Scholar]
- Kataoka, H.; Toschi, A.; Li, J.P.; Carney, R.L.; Schooley, D.A.; Kramer, S.J. Identification of an allatotropin from adult Manduca sexta. Science 1989, 243, 1481. [Google Scholar] [CrossRef] [PubMed]
- Nachman, R.J.; Moyna, G.; Williams, H.J.; Tobe, S.S.; Scott, A.I. Synthesis, biological activity, and conformational studies of insect allatostatin neuropeptide analogues incorporating turn-promoting moieties. Bioorg. Med. Chem. 1998, 6, 1379–1388. [Google Scholar] [CrossRef]
- Down, R.E.; Matthews, H.J.; Audsley, N. Effects of Manduca sexta allatostatin and an analog on the pea aphid Acyrthosiphon pisum (Hemiptera: Aphididae) and degradation by enzymes from the aphid gut. Peptides 2010, 31, 489–497. [Google Scholar] [CrossRef]
- Matthews, H.J.; Down, R.E.; Audsley, N. Effects of Manduca sexta allatostatin and an analogue on the peach-potato aphid Myzus persicae (hemiptera: Aphididae) and degradation by enzymes in the aphid gut. Arch. Insect Biochem. Physiol. 2010, 75, 139–157. [Google Scholar] [CrossRef]
- Mayoral, J.G.; Nouzova, M.; Brockhoff, A.; Goodwin, M.; Hernandez-Martinez, S.; Richter, D.; Meyerhof, W.; Noriega, F.G. Allatostatin-C receptors in mosquitoes. Peptides 2010, 31, 442–450. [Google Scholar] [CrossRef]
- Audsley, N.; Vandersmissen, H.P.; Weaver, R.; Dani, P.; Matthews, J.; Down, R.; Vuerinckx, K.; Kim, Y.; Vanden Broeck, J. Characterisation and tissue distribution of the PISCF allatostatin receptor in the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2013, 43, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wang, Z.; Jing, T. Functional characterization of ASTC (allatostatin C) and ASTCC (allatostatin double C) in Clostera anastomosis (Lepidoptera: Notodontidae). Gene 2017, 598, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Brown, M.R. Signaling and function of insulin-like peptides in insects. Annu. Rev. Entomol. 2006, 51, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Rulifson, E.J. Conserved mechanisms of glucose sensing and regulation by Drosophila corpora cardiaca cells. Nature 2004, 431, 316. [Google Scholar] [CrossRef]
- Isabel, G.; Martin, J.; Chidami, S.; Veenstra, J.A.; Rosay, P. AKH-producing neuroendocrine cell ablation decreases trehalose and induces behavioral changes in Drosophila. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R531–R538. [Google Scholar] [CrossRef] [PubMed]
- Holman, G.M.; Nachman, R.J.; Wright, M.S. Insect neuropeptides. Annu. Rev. Entomol. 1990, 35, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Holman, G.M.; Cook, B.J.; Nachman, R.J. Primary structure and synthesis of a blocked myotropic neuropeptide isolated from the cockroach, Leucophaea maderae. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1986, 85, 219–224. [Google Scholar] [CrossRef]
- Coast, G.M.; Holman, G.M.; Nachman, R.J. The diuretic activity of a series of cephalomyotropic neuropeptides, the achetakinins, on isolated Malpighian tubules of the house cricket, Acheta domesticus. J. Insect Physiol. 1990, 36, 481–488. [Google Scholar] [CrossRef]
- Hayes, T.K.; Pannabecker, T.L.; Hinckley, D.J.; Holman, G.M.; Nachman, R.J.; Petzel, D.H.; Beyenbach, K.W. Leucokinins, a new family of ion transport stimulators and inhibitors in insect Malpighian tubules. Life Sci. 1989, 44, 1259–1266. [Google Scholar] [CrossRef]
- Nachman, R.J.; Strey, A.; Isaac, E.; Pryor, N.; Lopez, J.D.; Deng, J.; Coast, G.M. Enhanced in vivo activity of peptidase-resistant analogs of the insect kinin neuropeptide family. Peptides 2002, 23, 735–745. [Google Scholar] [CrossRef]
- Nachman, R.J.; Coast, G.M.; Douat, C.; Fehrentz, J.; Kaczmarek, K.; Zabrocki, J.; Pryor, N.W.; Martinez, J. A C-terminal aldehyde insect kinin analog enhances inhibition of weight gain and induces significant mortality in Helicoverpa zea larvae. Peptides 2003, 24, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Smagghe, G.; Mahdian, K.; Zubrzak, P.; Nachman, R.J. Antifeedant activity and high mortality in the pea aphid Acyrthosiphon pisum (Hemiptera: Aphidae) induced by biostable insect kinin analogs. Peptides 2010, 31, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Verleyen, P.; Huybrechts, J.; Schoofs, L. SIFamide illustrates the rapid evolution in Arthropod neuropeptide research. Gen. Comp. Endocrinol. 2009, 162, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Terhzaz, S.; Rosay, P.; Goodwin, S.F.; Veenstra, J.A. The neuropeptide SIFamide modulates sexual behavior in Drosophila. Biochem. Biophys. Res. Commun. 2007, 352, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Stormo, G.D.; Taghert, P.H. The neuropeptide pigment-dispersing factor coordinates pacemaker interactions in the Drosophila circadian system. J. Neurosci. 2004, 24, 7951–7957. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.J.; Jan, L.Y.; Jan, Y.N. A PDF/NPF neuropeptide signaling circuitry of male Drosophila melanogaster controls rival-induced prolonged mating. Neuron 2013, 80, 1190–1205. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Lkhagva, A.; Daubnerová, I.; Chae, H.; Šimo, L.; Jung, S.; Yoon, Y.; Lee, N.; Seong, J.Y.; Žitňan, D.; et al. Natalisin, a tachykinin-like signaling system, regulates sexual activity and fecundity in insects. Proc. Natl. Acad. Sci. USA 2013, 110, E3526–E3534. [Google Scholar] [CrossRef]
- Gui, S.H.; Jiang, H.B.; Liu, X.Q.; Xu, L.; Wang, J.J. Molecular characterizations of natalisin and its roles in modulating mating in the oriental fruit fly, Bactrocera dorsalis (Hendel). Insect Mol. Biol. 2017, 26, 103–112. [Google Scholar] [CrossRef]
- Leal, W.S.; Choo, Y.M.; Xu, P.; Da Silva, C.S.B.; Ueira-Vieira, C. Differential expression of olfactory genes in the southern house mosquito and insights into unique odorant receptor gene isoforms. Proc. Natl. Acad. Sci. USA 2013, 110, 18704–18709. [Google Scholar] [CrossRef]
- Vanden Broeck, J.; Torfs, H.; Poels, J.; Poyer, W.; Swinnen, E.; Ferket, K.; Loof, A. Tachykinin-like peptides and their receptors: A review. Ann. N. Y. Acad. Sci. 1999, 897, 374–387. [Google Scholar] [CrossRef]
- Winther, Å.M.E.; Acebes, A.; Ferrús, A. Tachykinin-related peptides modulate odor perception and locomotor activity in Drosophila. Mol. Cell. Neurosci. 2006, 31, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Shankar, S.; Chua, J.Y.; Tan, K.J.; Calvert, M.E.; Weng, R.; Ng, W.C.; Mori, K.; Yew, J.Y. The neuropeptide tachykinin is essential for pheromone detection in a gustatory neural circuit. eLife 2015, 4, e6914. [Google Scholar] [CrossRef] [PubMed]
- Zdarek, J.; Nachman, R.J.; Denlinger, D.L. Parturition hormone in the tsetse Glossina morsitans: Activity in reproductive tissues from other species and response of tsetse to identified neuropeptides and other neuroactive compounds. J. Insect Physiol. 2000, 46, 213–219. [Google Scholar] [CrossRef]
- Hansen, K.K.; Hauser, F.; Williamson, M.; Weber, S.B.; Grimmelikhuijzen, C.J.P. The Drosophila genes CG14593 and CG30106 code for G-protein-coupled receptors specifically activated by the neuropeptides CCHamide-1 and CCHamide-2. Biochem. Biophys. Res. Commun. 2011, 404, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Ida, T.; Takahashi, T.; Tominaga, H.; Sato, T.; Sano, H.; Kume, K.; Ozaki, M.; Hiraguchi, T.; Shiotani, H.; Terajima, S.; et al. Isolation of the bioactive peptides CCHamide-1 and CCHamide-2 from Drosophila and their putative role in appetite regulation as ligands for G protein-coupled receptors. Front. Endocrinol. 2012, 3, 177. [Google Scholar] [CrossRef] [PubMed]
- Farhan, A.; Gulati, J.; Groβe-Wilde, E.; Vogel, H.; Hansson, B.S.; Knaden, M. The CCHamide 1 receptor modulates sensory perception and olfactory behavior in starved Drosophila. Sci. Rep. 2013, 3, 2765. [Google Scholar] [CrossRef] [PubMed]
- Berg, B.G.; Schachtner, J.; Utz, S.; Homberg, U. Distribution of neuropeptides in the primary olfactory center of the heliothine moth Heliothis virescens. Cell Tissue Res. 2006, 327, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Utz, S.; Huetteroth, W.; Wegener, C.; Kahnt, J.; Predel, R.; Schachtner, J. Direct peptide profiling of lateral cell groups of the antennal lobes of Manduca sexta reveals specific composition and changes in neuropeptide expression during development. Dev. Neurobiol. 2007, 67, 764–777. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Cserzo, M.; Eisenhaber, F.; Eisenhaber, B.; Simon, I. On filtering false positive transmembrane protein predictions. Protein Eng. 2002, 15, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, Y.; Liu, Z.; Cheng, H.; Xue, Y. HemI: A toolkit for illustrating heatmaps. PLoS ONE 2014, 9, e111988. [Google Scholar] [CrossRef]
- Xu, Y.; Gao, S.; Yang, Y.; Huang, M.; Cheng, L.; Wei, Q.; Fei, Z.; Gao, J.; Hong, B. Transcriptome sequencing and whole genome expression profiling of chrysanthemum under dehydration stress. BMC Genomics 2013, 14, 662. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, H.; Bin, S.; Chen, L.; Han, Q.; Lin, J. Antennal and abdominal transcriptomes reveal chemosensory genes in the Asian citrus psyllid, Diaphorina citri. PLoS ONE 2016, 11, e159372. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neuropeptide Name | Accession No. | Acronym | ORF (aa) | SP (aa) | Homology Search with Known Protein | ||
|---|---|---|---|---|---|---|---|
| Species | Protein ID | E-Value | |||||
| Adipokinetic hormone | MG550150 | AKH | 74 | 24 | Nilaparvata lugens | AFN26934.1 | 6.00e-17 |
| Allatostatin A | MG550151 | AstA | 181 | 21 | Rhodnius prolixus | ACX47066.1 | 2.00e-21 |
| Allatostatin B | MG550152 | AstB | 218 | 22 | Plautia stali | BAU88428.1 | 2.00e-34 |
| Allatostatin C | MG550153 | AstC | 98 | 23 | Pseudomyrmex gracilis | XP_020296756.1 | 1.00e-20 |
| Allatostatin CC | MG550154 | AstCC | 117 | 21 | Plautia stali | BAV78791.1 | 6.00e-27 |
| Allatotropin | MG550155 | AT | 152 | 28 | Nilaparvata lugens | BAO00936.1 | 1.00e-08 |
| Arginine vasopressin-like peptide | MG550156 | AVLP | 170 | 21 | Nilaparvata lugens | BAO00934.1 | 1.00e-04 |
| Bursicon beta subunit | MG550157 | Burβ | 134 | 19 | Nilaparvata lugens | BAO00938.1 | 5.90e-52 |
| Cardio acceleratory peptide 2b/Pyrokinin 1 | MG550158 | CAP2b | 189 | 29 | Nilaparvata lugens | BAO00941.1 | 1.00e-13 |
| CCHamide 1 | MG550159 | CCH1 | 165 | 26 | Nilaparvata lugens | BAO00942.1 | 8.00e-13 |
| CCHamide 2 | MG550160 | CCH2 | 169 | 28 | Bombyx mori | BAG55002.1 | 6.00e-08 |
| CNMamide | MG550161 | CNM | 165 | 20 | Plautia stali | BAV78799.1 | 9.00e-07 |
| Corazonin | MG550162 | Crz | 155 | 26 | Solenopsis invicta | XP_011172976.1 | 2.00e-03 |
| Crustacean cardioactive peptide | MG550163 | CCAP | 149 | 25 | Plautia stali | BAV78802.1 | 4.00e-42 |
| Pyrokinin 2/Pheromone biosynthesis activating neruopeptide | MG550164 | PBAN | 200 | 29 | Bemisia tabaci | XP_018895833.1 | 5.00e-20 |
| Diuretic hormone 31 | MG550165 | DH31 | 119 | 29 | Nilaparvata lugens | BAO00939.1 | 2.00e-35 |
| Diuretic hormone 45/splicing variant of CRF-DH | MG550166 | DH45 | 246 | 24 | Nilaparvata lugens | BAO00945.1 | 6.00e-15 |
| Eclosion hormone 1 | MG550167 | EH1 | 80 | 25 | Diuraphis noxia | XP_015365040.1 | 1.00e-34 |
| Eclosion hormone 2 | MG550168 | EH2 | 84 | 28 | Nilaparvata lugens | BAO00951.1 | 2.00e-19 |
| Ecdysis triggering hormone | MG550169 | ETH | 152 | 20 | Plautia stali | BAV78804.1 | 2.00e-06 |
| Elevenin-like peptide | MG550170 | ELP | 128 | 26 | Nilaparvata lugens | BAO00952.1 | 3.00e-06 |
| Glycoprotein hormone beta 5 | MG550171 | GPβ5 | 138 | 18 | Nilaparvata lugens | BAO00956.1 | 3.00e-53 |
| ITG-containing peptide | MG550172 | ITG | 216 | 21 | Agrilus planipennis | XP_018325057.1 | 2.00e-81 |
| Insulin-like peptide 1 | MG550173 | ILP1 | 136 | 25 | Nilaparvata lugens | AIY24645.1 | 1.00e-03 |
| Insulin-like peptide 2 | MG550174 | ILP2 | 147 | 20 | Nilaparvata lugens | BAO00958.1 | 4.00e-04 |
| Ion transport peptide like | MG550175 | ITPL | 124 | 26 | Harpegnathos saltator | XP_011137372.1 | 2.00e-51 |
| Kinin | MG550176 | Kin | 327 | 28 | Plautia stali | BAV78814.1 | 6.00e-17 |
| Myosuppressin | MG550177 | MS | 100 | 22 | Nilaparvata lugens | BAO00963.1 | 6.00e-32 |
| Natalisin | MG550178 | NTL | 214 | 25 | Chilo suppressalis | ALM30330.1 | 8.70e-02 |
| Neuroparsin | MG550179 | NP | 96 | 19 | Bombyx mori | BAG50366.1 | 1.00e-13 |
| Neuropeptide-like precursor 1 | MG550180 | NPLP1 | 617 | 19 | Nilaparvata lugens | BAO00966.1 | 3.00e-05 |
| neuropeptide-like precursor 3 | MG550181 | NPLP3 | 83 | 16 | Anoplophora glabripennis | XP_018572912.1 | 7.00e-03 |
| Orcokinin | MG550182 | OK | 164 | 17 | Nilaparvata lugens | BAO00969.1 | 2.00e-30 |
| Pigment dispersing factor | MG550183 | 82 | 25 | Nilaparvata lugens | BAO00970.1 | 1.00e-07 | |
| Proctolin | MG550184 | Pro | 72 | 28 | Nilaparvata lugens | BAO00972.1 | 1.20e-02 |
| Prothoracicotropic hormone | MG550185 | PTTH | 168 | 31 | Nilaparvata lugens | BAO00973.1 | 1.00e-09 |
| RYamide | MG550186 | RY | 165 | 24 | Chilo suppressalis | ALM30346.1 | 1.80e-02 |
| Short neuropeptide F | MG550187 | sNPF | 110 | 27 | Nilaparvata lugens | BAO00976.1 | 2.00e-17 |
| SIFamide | MG550188 | SIF | 75 | 22 | Tribolium castaneum | EFA07409.1 | 3.00e-17 |
| Tachykinin | MG550189 | TK | 199 | 20 | Tribolium castaneum | KYB25859.1 | 3.00e-14 |
| Receptor Name | Accession No. | ORF (aa) | TMD (No.) | Putative Identification | Species | Matched Gene (Accession No.) | E-Value |
|---|---|---|---|---|---|---|---|
| Neuropeptide receptor A1 | MG550190 | 439 | 8 | Allatostatin receptor type C | Carausius morosus | AOV81581.1 | 2e-179 |
| Neuropeptide receptor A2 | MG550191 | 392 | 7 | Allatostatin-A receptor-like | Nilaparvata lugens | XP_022192722.1 | 6e-168 |
| Neuropeptide receptor A3 | MG550192 | 434 | 7 | Neuropeptide GPCR A3 | Nilaparvata lugens | BAO01052.1 | 3e-136 |
| Neuropeptide receptor A4 | MG550193 | 542 | 7 | SIFamide receptor | Onthophagus taurus | XP_022919670.1 | 9e-162 |
| Neuropeptide receptor A5 | MG550194 | 381 | 7 | QRFP-like peptide receptor | Halyomorpha halys | XP_014285975.1 | 1e-137 |
| Neuropeptide receptor A6 | MG550195 | 1185 | 6 | Neuropeptide GPCR A6b | Nilaparvata lugens | BAO01056.1 | 7e-143 |
| Neuropeptide receptor A7 | MG550196 | 439 | 7 | Neuropeptide receptor A7 | Nilaparvata lugens | BAO01057.1 | 5e-163 |
| Neuropeptide receptor A8 | MG550197 | 520 | 7 | G-protein coupled receptor moody | Cimex lectularius | XP_014257264.1 | 0 |
| Neuropeptide receptor A9 | MG550198 | 383 | 7 | G-protein coupled receptor moody isoform X1 | Halyomorpha halys | XP_014287792.1 | 2e-152 |
| Neuropeptide receptor A10 | MG550199 | 406 | 7 | Sex peptide receptor | Bemisia tabaci | XP_018916635.1 | 0 |
| Neuropeptide receptor A11 | MG550200 | 410 | 7 | Thyrotropin-releasing hormone receptor-like | Diaphorina citri | XP_008487532.2 | 0 |
| Neuropeptide receptor A12 | MG550201 | 438 | 7 | Neuropeptide receptor | Diaphorina citri | AWT50635.1 | 0 |
| Neuropeptide receptor A13 | MG550202 | 411 | 7 | Myosuppressin receptor | Rhodnius prolixus | AGT02812.1 | 8e-146 |
| Neuropeptide receptor A14 | MG550203 | 414 | 7 | CCHamide-1 receptor | Cimex lectularius | XP_014250319.1 | 3e-173 |
| Neuropeptide receptor A15 | MG550204 | 378 | 7 | CCHamide-2 receptor | Melanaphis sacchari | XP_025191408.1 | 2e-177 |
| Neuropeptide receptor A16 | MG550205 | 439 | 7 | Neuropeptide GPCR A16 | Nilaparvata lugens | BAO01066.1 | 5e-137 |
| Neuropeptide receptor A17 | MG550206 | 362 | 5 | Neuropeptide GPCR A17 | Nilaparvata lugens | BAO01067.1 | 7e-56 |
| Neuropeptide receptor A18 | MG550207 | 432 | 7 | Neuropeptide GPCR A18 | Nilaparvata lugens | BAO01068.1 | 3e-126 |
| Neuropeptide receptor A19 | MG550208 | 460 | 7 | Neuropeptide GPCR A19 | Nilaparvata lugens | BAO01069.1 | 1e-132 |
| Neuropeptide receptor A20 | MG550209 | 426 | 7 | RYamide receptor | Zootermopsis nevadensis | XP_021924354.1 | 2e-125 |
| Neuropeptide receptor A21 | MG550210 | 405 | 7 | Corazonin receptor | Pyrrhocoris apterus | ARV86500.1 | 2e-156 |
| Neuropeptide receptor A22 | MG550211 | 461 | 7 | RYamide receptor-like | Nilaparvata lugens | XP_022187836.1 | 4e-153 |
| Neuropeptide receptor A23 | MG550212 | 448 | 7 | Neuropeptide GPCR A23 | Nilaparvata lugens | BAO01073.1 | 0 |
| Neuropeptide receptor A24 | MG550213 | 423 | 7 | Tachykinin receptor | Periplaneta americana | ARK07245.1 | 0 |
| Neuropeptide receptor A25 | MG550214 | 463 | 7 | Capa receptor-like | Bemisia tabaci | XP_018899674.1 | 8e-178 |
| Neuropeptide receptor A26 | MG550215 | 349 | 7 | Neuropeptide GPCR A43 | Nilaparvata lugens | BAO01093.1 | 6e-109 |
| Neuropeptide receptor A27 | MG550216 | 322 | 7 | Cardio acceleratory peptide receptor | Nilaparvata lugens | XP_022190770.1 | 1e-167 |
| Neuropeptide receptor A28 | MG550217 | 543 | 7 | Neuropeptide F receptor | Rhodnius prolixus | AKO62911.1 | 2e-48 |
| Neuropeptide receptor A29 | MG550218 | 383 | 7 | Pyrokinin-1 receptor | Rhodnius prolixus | AFO73269.1 | 1e-154 |
| Neuropeptide receptor A30 | MG550219 | 552 | 7 | Pyrokinin-1 receptor | Cimex lectularius | XP_014246737.1 | 8e-122 |
| Neuropeptide receptor A31 | MG550220 | 520 | 7 | Pyrokinin-1 receptor | Cephus cinctus | XP_024938460.1 | 6e-113 |
| Neuropeptide receptor A32 | MG550221 | 493 | 6 | AKH receptor | Rhodnius prolixus | AKO62857.1 | 1e-96 |
| Neuropeptide receptor A33 | MG550222 | 365 | 7 | AKH receptor | Pseudoregma bambucicola | AKH80288.1 | 2e-151 |
| Neuropeptide receptor A34 | MG550223 | 419 | 7 | Natalisin receptor | Bactrocera dorsalis | AQM36729.1 | 7e-144 |
| Neuropeptide receptor A35 | MG550224 | 423 | 7 | Neuropeptide GPCR A41 | Nilaparvata lugens | BAO01091.1 | 2e-123 |
| Neuropeptide receptor B1 | MG550228 | 408 | 7 | Neuropeptide GPCR B1 | Nilaparvata lugens | BAO01101.1 | 4e-164 |
| Neuropeptide receptor B2 | MG550229 | 463 | 7 | PDF receptor | Nilaparvata lugens | XP_022204430.1 | 0 |
| Neuropeptide receptor B3 | MG550230 | 420 | 7 | Neuropeptide receptor B4 | Chilo suppressalis | ALM88344.1 | 4e-139 |
| Neuropeptide receptor B4 | MG550231 | 415 | 7 | Diuretic hormone receptor | Bemisia tabaci | XP_018897695.1 | 0 |
| Leucine-rich repeat G-protein-coupled receptor 1 | MG550225 | 707 | 7 | Lutropin-choriogonadotropic hormone receptor | Drosophila rhopaloa | XP_016976568.1 | 0 |
| Leucine-rich repeat G-protein-coupled receptor 2 | MG550226 | 817 | 7 | Lutropin-choriogonadotropic hormone receptor-like | Cryptotermes secundus | XP_023716984.1 | 0 |
| Leucine-rich repeat G-protein-coupled receptor 3 | MG550226 | 1011 | 7 | Neuropeptide GPCR A47 | Nilaparvata lugens | BAO01097.1 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Zhou, W.; Hameed, M.S.; Liu, J.; Zeng, X. Characterization and Expression Profiling of Neuropeptides and G-Protein-Coupled Receptors (GPCRs) for Neuropeptides in the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae). Int. J. Mol. Sci. 2018, 19, 3912. https://doi.org/10.3390/ijms19123912
Wang Z, Zhou W, Hameed MS, Liu J, Zeng X. Characterization and Expression Profiling of Neuropeptides and G-Protein-Coupled Receptors (GPCRs) for Neuropeptides in the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae). International Journal of Molecular Sciences. 2018; 19(12):3912. https://doi.org/10.3390/ijms19123912
Chicago/Turabian StyleWang, Zhengbing, Wenwu Zhou, Muhammad Salman Hameed, Jiali Liu, and Xinnian Zeng. 2018. "Characterization and Expression Profiling of Neuropeptides and G-Protein-Coupled Receptors (GPCRs) for Neuropeptides in the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae)" International Journal of Molecular Sciences 19, no. 12: 3912. https://doi.org/10.3390/ijms19123912
APA StyleWang, Z., Zhou, W., Hameed, M. S., Liu, J., & Zeng, X. (2018). Characterization and Expression Profiling of Neuropeptides and G-Protein-Coupled Receptors (GPCRs) for Neuropeptides in the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae). International Journal of Molecular Sciences, 19(12), 3912. https://doi.org/10.3390/ijms19123912