A Proteomic Approach for Understanding the Mechanisms of Delayed Corneal Wound Healing in Diabetic Keratopathy Using Diabetic Model Rat


 ,
,
Abstract
1. Introduction
2. Results
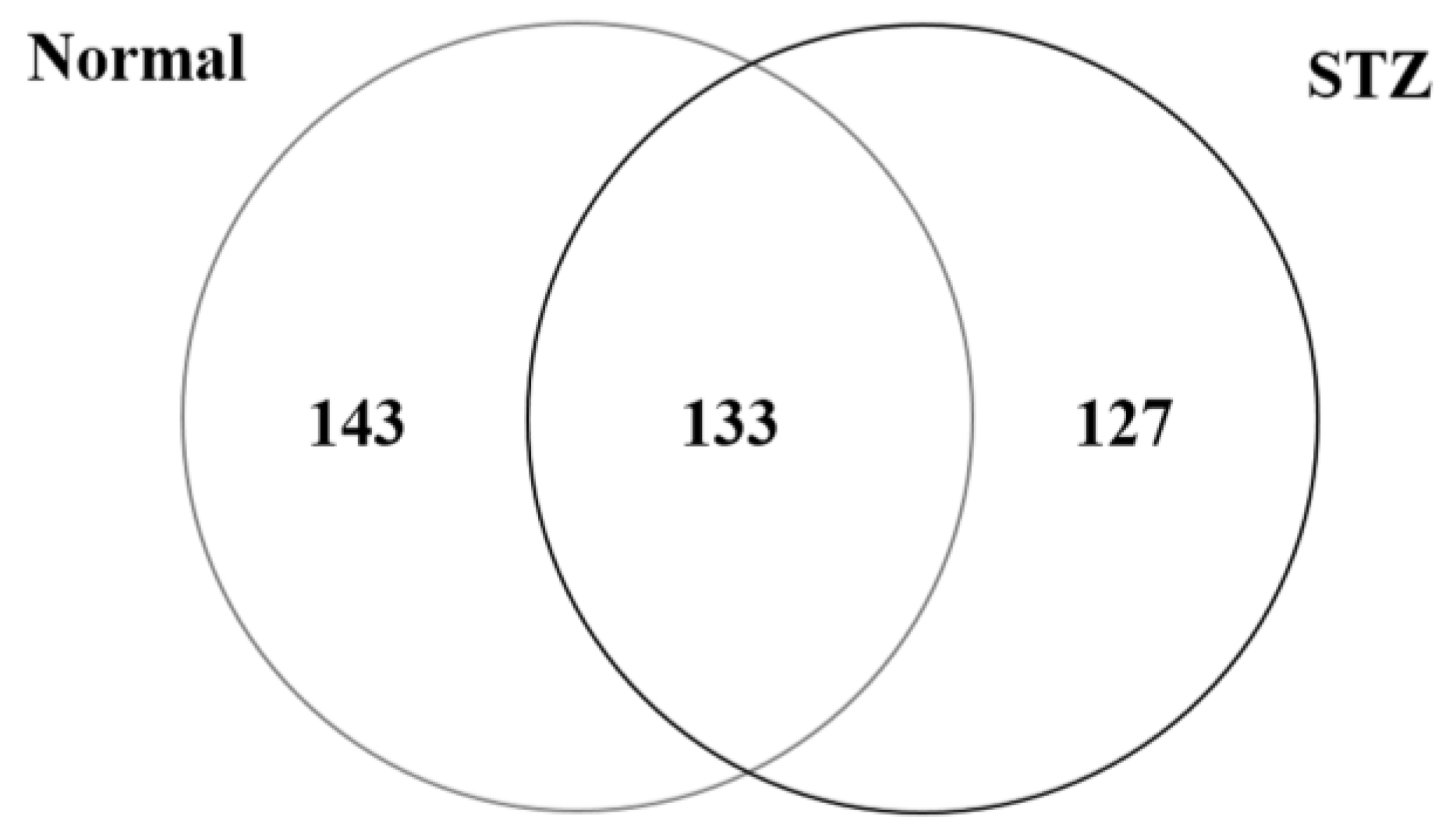
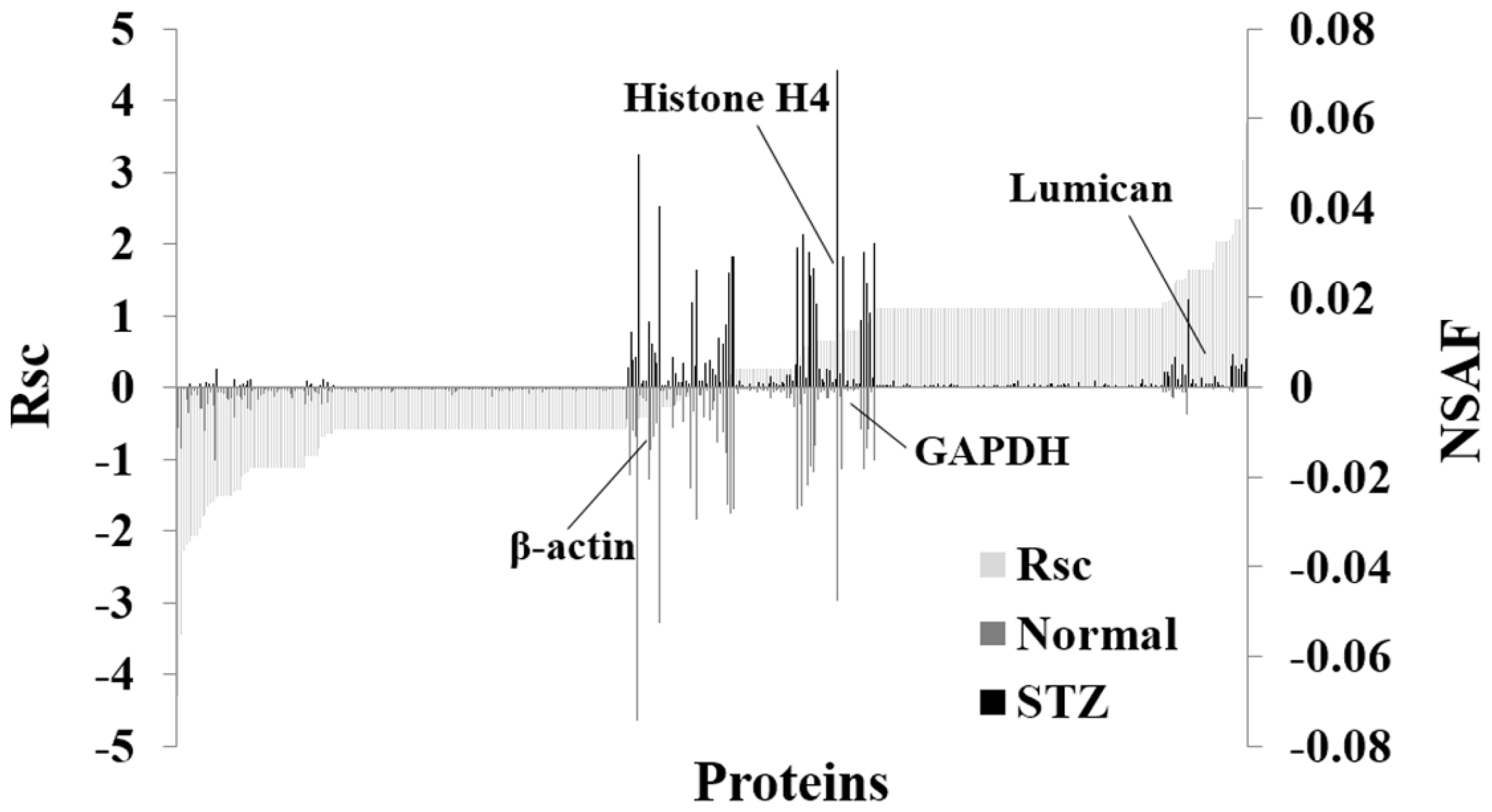
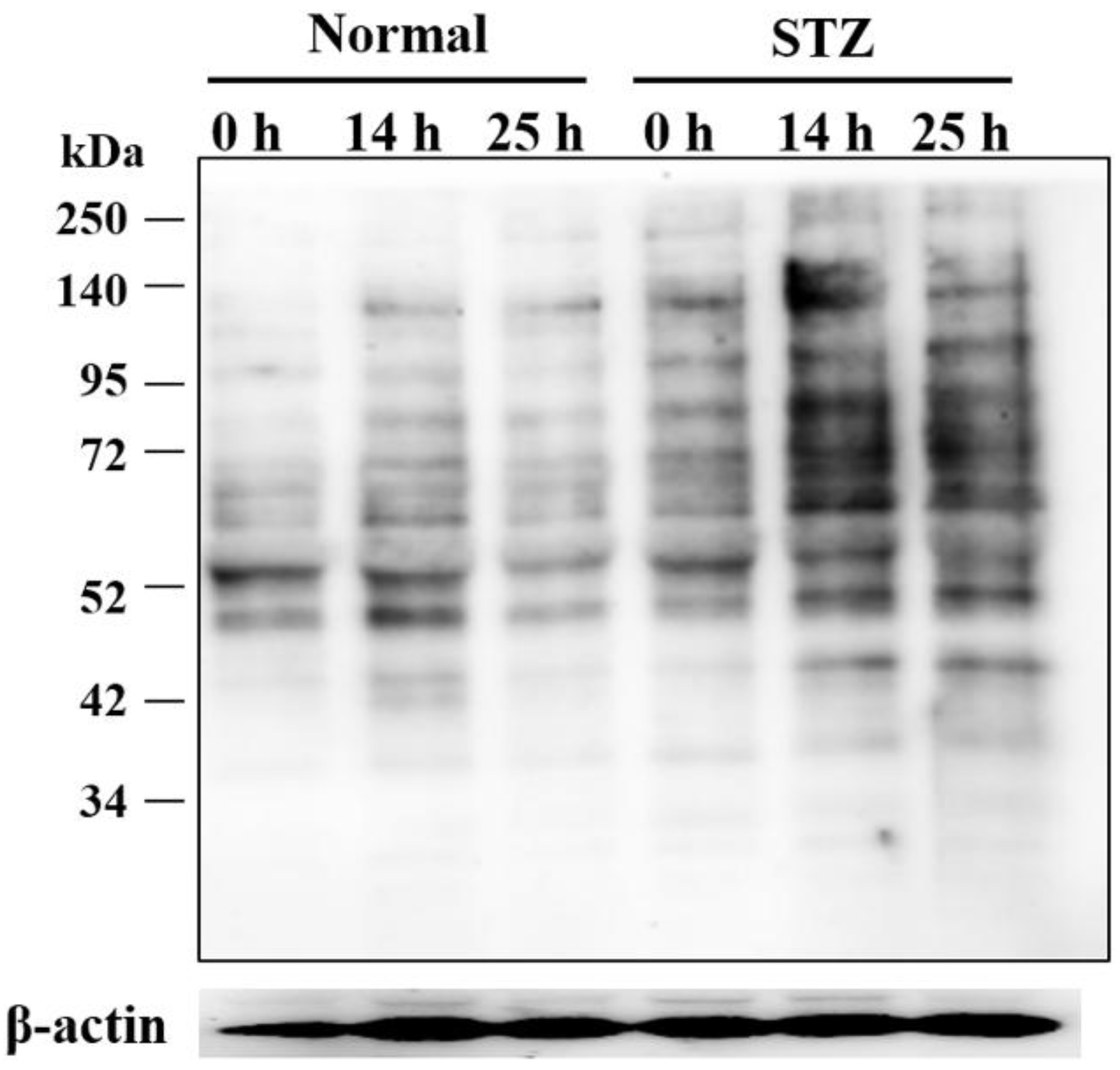
2.1. Identification and Semiquantitative Comparisons of Differentially Expressed Proteins in the Cornea of Normal and STZ Rats
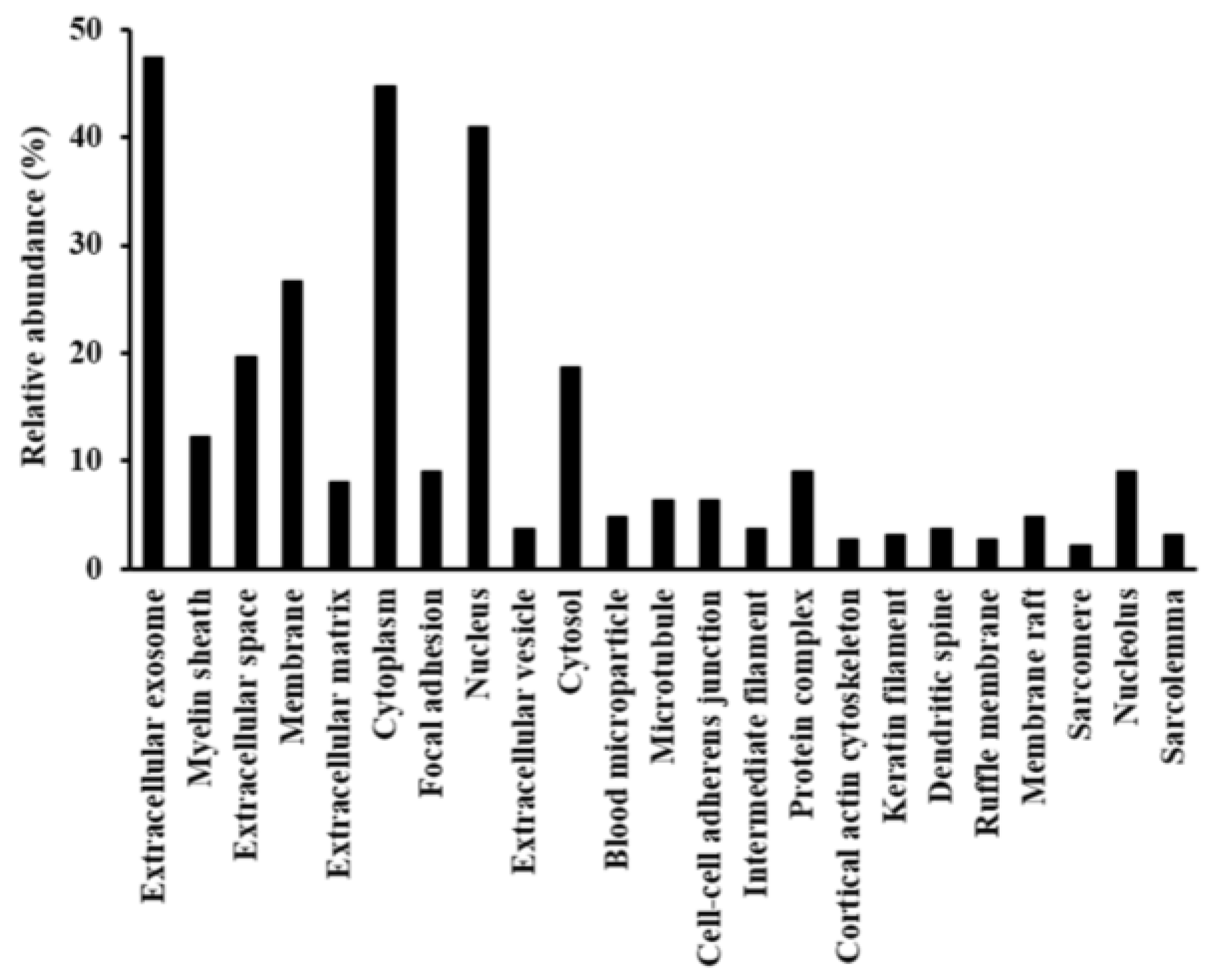
2.2. Functional Annotation of Proteins Regulated by Hyperglycemia
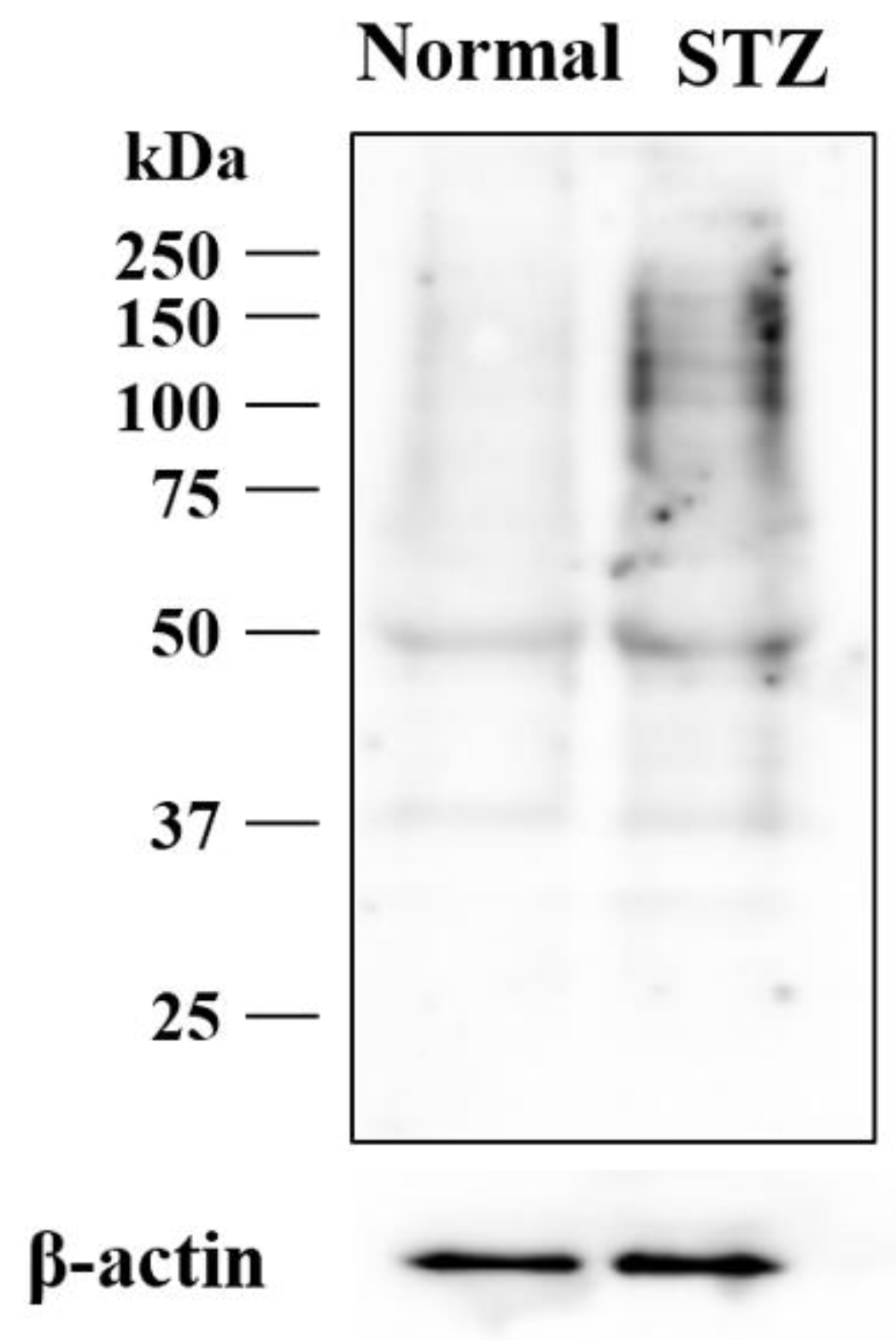
2.3. Effects of Hyperglycemia on Lumican Expression in Cornea of Rat
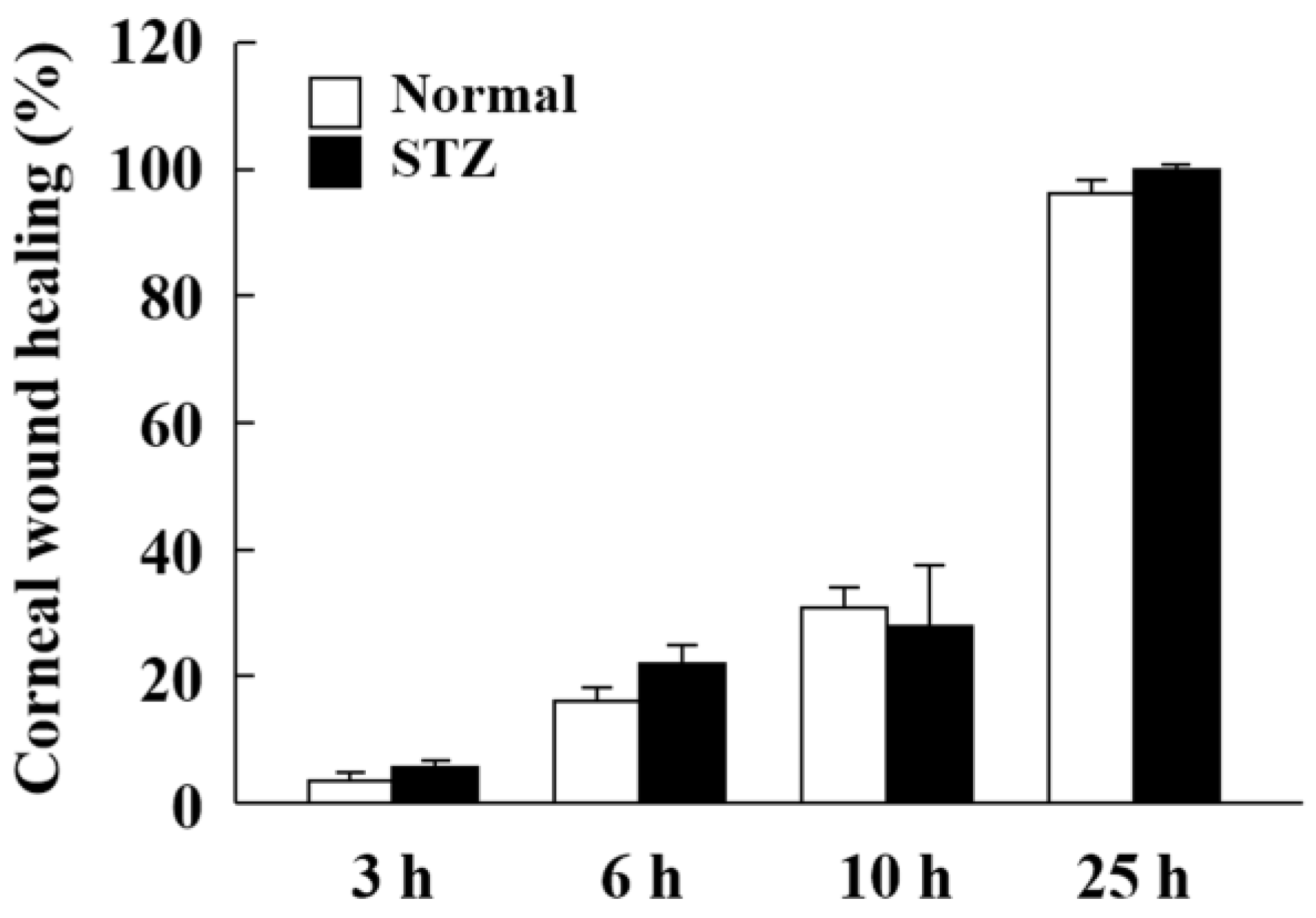
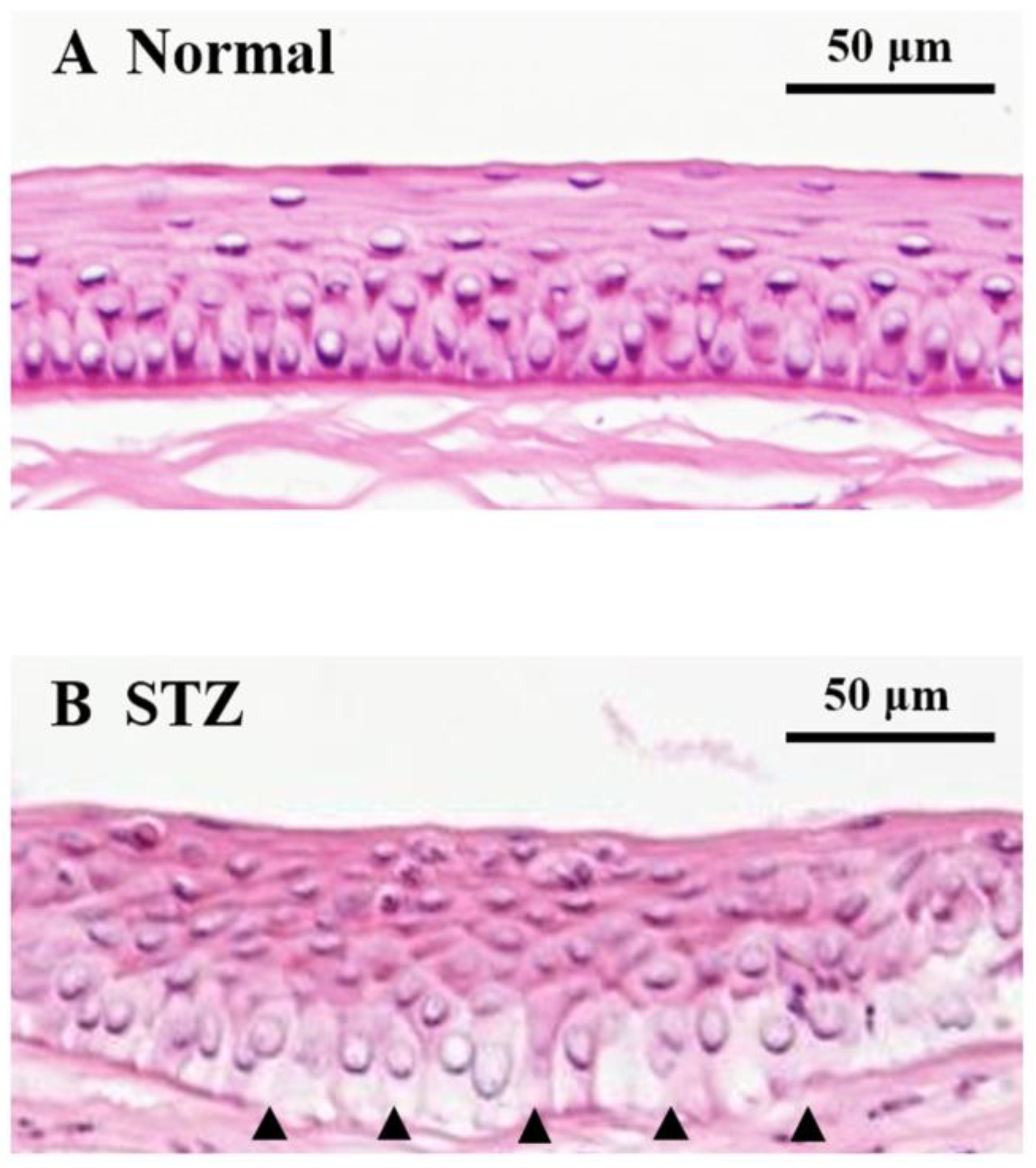
2.4. Changes in Expression of Lumican during Corneal Wound Healing in STZ Rats
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Assay of Glucose and Insulin
4.4. Tryptic Digestion of Protein Extracted from the Cornea of Normal and STZ Rats
4.5. LC-MS/MS Analysis for Protein Identification
4.6. Semiquantitative Analysis of Identified Proteins
4.7. Bioinformatics
4.8. Western Blot Analysis
4.9. In Vivo Wound Healing of the Corneal Epithelium of the Rats
4.10. Hematoxylin and Eosin Staining of the Cornea
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DM | Diabetes mellitus |
| ECM | Extracellular matrix |
| GO | Gene ontology |
| LC/MS | Liquid chromatography/mass spectroscopy |
| NSAF | Normalized spectral abundance factor |
| SLRP | Small leucine-rich proteoglycan |
| STZ | Streptozotocin |
References
- Shaw, J.E.; Sicree, R.A.; Zimmet, P.Z. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Expert Committee on the Diagnosis and Classification of Diabetes Mellitus. Report of the expert committee on the diagnosis and classification of diabetes mellitus. Diabetes Care 2003, 26, S5–S20. [Google Scholar]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Chikama, T.; Wakuta, M.; Liu, Y.; Nishida, T. Deviated mechanism of wound healing in diabetic corneas. Cornea 2007, 26, S75–S81. [Google Scholar] [CrossRef] [PubMed]
- Friend, J.; Ishii, Y.; Thoft, R.A. Corneal epithelial changes in diabetic rats. Ophthalmic Res. 1982, 14, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Azar, D.T.; Spurr-Michaud, S.J.; Tisdale, A.S.; Gipson, I.K. Altered epithelial-basement membrane interactions in diabetic corneas. Arch. Ophth. 1992, 110, 537–540. [Google Scholar] [CrossRef]
- McDermott, A.M.; Xiao, T.L.; Kern, T.S.; Murphy, C.J. Non-enzymatic glycation in corneas from normal and diabetic donors and its effects on epithelial cell attachment in vitro. Optometry 2003, 74, 443–452. [Google Scholar] [PubMed]
- Ljubimov, A.V.; Alba, S.A.; Burgeson, R.E.; Ninomiya, Y.; Sado, Y.; Sun, T.T.; Nesburn, A.B.; Kenney, M.C.; Maguen, E. Extracellular matrix changes in human corneas after radial keratotomy. Exp. Eye Res. 1998, 67, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Kabosova, A.; Kramerov, A.A.; Aoki, A.M.; Murphy, G.; Zieske, J.D.; Ljubimov, A.V. Human diabetic corneas preserve wound healing, basement membrane, integrin and MMP-10 differences from normal corneas in organ culture. Exp. Eye Res. 2003, 77, 211–217. [Google Scholar] [CrossRef]
- Zagon, I.S.; Sassani, J.W.; McLaughlin, P.J. Insulin treatment ameliorates impaired corneal reepithelialization in diabetic rats. Diabetes 2006, 55, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Wakuta, M.; Morishige, N.; Chikama, T.; Seki, K.; Nagano, T.; Nishida, T. Delayed wound closure and phenotypic changes in corneal epithelium of the spontaneously diabetic Goto-Kakizaki rat. Invest. Ophth. Vis. Sci. 2007, 48, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Okugawa, K.; Amano, S.; Oshika, T.; Takamura, E.; Egami, F.; Umizu, G.; Aikawa, K.; Kato, S. Blinking and superficial punctate keratopathy in patients with diabetes mellitus. Eye (Lond) 2005, 19, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Rehany, U.; Ishii, Y.; Lahav, M.; Rumelt, S. Ultrastructural changes in corneas of diabetic patients: An electron-microscopy study. Cornea 2000, 19, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Saini, J.S.; Khandalavla, B. Corneal epithelial fragility in diabetes mellitus. Can. J. Ophth. 1995, 30, 142–146. [Google Scholar]
- Schultz, R.O.; Van Horn, D.L.; Peters, M.A.; Klewin, K.M.; Schutten, W.H. Diabetic keratopathy. Trans. Am. Ophthal. Soc. 1981, 79, 180–199. [Google Scholar] [PubMed]
- Zieske, J.D.; Bukusoglu, G.; Gipson, I.K. Enhancement of vinculin synthesis by migrating stratified squamous epithelium. J. Cell Biol. 1989, 109, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.X.; Gipson, I.K.; Guo, Y. Differential gene expression in healing rat corneal epithelium. Invest. Ophth. Vis. Sci. 1995, 36, 1997–2007. [Google Scholar]
- Yu, F.X.; Guo, J.; Zhang, Q. Expression and distribution of adhesion molecule CD44 in healing corneal epithelia. Invest. Ophth. Vis. Sci. 1998, 39, 710–717. [Google Scholar]
- Inoue, M.; Kratz, G.; Haegerstrand, A.; Stahle-Backdahl, M. Collagenase expression is rapidly induced in wound-edge keratinocytes after acute injury in human skin, persists during healing, and stops at re-epithelialization. J. Invest. Dermatol. 1995, 104, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Saarialho-Kere, U.K.; Kovacs, S.O.; Pentland, A.P.; Olerud, J.E.; Welgus, H.G.; Parks, W.C. Cell-matrix interactions modulate interstitial collagenase expression by human keratinocytes actively involved in wound healing. J. Clin. Invest. 1993, 92, 2858–2866. [Google Scholar] [CrossRef] [PubMed]
- Hassell, J.R.; Newsome, D.A.; Krachmer, J.H.; Rodrigues, M.M. Macular corneal dystrophy: Failure to synthesize a mature keratan sulfate proteoglycan. Proc. Natl. Acad. Sci. USA 1980, 77, 3705–3709. [Google Scholar] [CrossRef] [PubMed]
- Dunlevy, J.R.; Chakravarti, S.; Gyalzen, P.; Vergnes, J.P.; Hassell, J.R. Cloning and chromosomal localization of mouse keratocan, a corneal keratan sulfate proteoglycan. Mamm. Genome 1998, 9, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Joseph, R.; Srivastava, O.P.; Pfister, R.R. Differential epithelial and stromal protein profiles in keratoconus and normal human corneas. Exp. Eye Res. 2011, 92, 282–298. [Google Scholar] [CrossRef] [PubMed]
- Nikitovic, D.; Katonis, P.; Tsatsakis, A.; Karamanos, N.K.; Tzanakakis, G.N. Lumican, a small leucine-rich proteoglycan. IUBMB Life 2008, 60, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V. Matrix proteoglycans: From molecular design to cellular function. Annu. Rev. Biochem. 1998, 67, 609–652. [Google Scholar] [CrossRef] [PubMed]
- Saika, S.; Shiraishi, A.; Liu, C.Y.; Funderburgh, J.L.; Kao, C.W.; Converse, R.L.; Kao, W.W. Role of lumican in the corneal epithelium during wound healing. J. Biol. Chem. 2000, 275, 2607–2612. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, S.; Magnuson, T.; Lass, J.H.; Jepsen, K.J.; LaMantia, C.; Carroll, H. Lumican regulates collagen fibril assembly: Skin fragility and corneal opacity in the absence of lumican. J. Cell Biol. 1998, 141, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.H.; Lai, J.Y.; Yu, S.T.; Liu, J.Y.; Yang, U.; Chen, H.C.; Yeh, L.K.; Ho, Y.J.; Chang, G.; Wang, S.F.; et al. Up-regulation of heat shock protein 70-1 (Hsp70-1) in human limbo-corneal epithelial cells cultivated on amniotic membrane: A proteomic study. J. Cell. Physiol. 2012, 227, 2030–2039. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Kudo, M.; Peng, W.X.; Naito, Z. Analysis of protein expression regulated by lumican in PANC-1 cells using shotgun proteomics. Oncol. Rep. 2013, 30, 1609–1621. [Google Scholar] [CrossRef] [PubMed]
- Meade, M.L.; Shiyanov, P.; Schlager, J.J. Enhanced detection method for corneal protein identification using shotgun proteomics. Proteome Sci. 2009, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Nagai, N.; Yamamoto, T.; Mitamura, K.; Taga, A. Proteomic profile of the lens in a streptozotocin-induced diabetic rat model using shotgun proteomics. Biomed. Rep. 2017, 7, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Grover, J.; Chen, X.N.; Korenberg, J.R.; Roughley, P.J. The human lumican gene. Organization, chromosomal location, and expression in articular cartilage. J. Biol. Chem. 1995, 270, 21942–21949. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, S.; Stallings, R.L.; SundarRaj, N.; Cornuet, P.K.; Hassell, J.R. Primary structure of human lumican (keratan sulfate proteoglycan) and localization of the gene (LUM) to chromosome 12q21.3-q22. Genomics 1995, 27, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Naito, Z. Role of the small leucine-rich proteoglycan (SLRP) family in pathological lesions and cancer cell growth. J. Nippon Med. Sch. 2005, 72, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Funderburgh, J.L. Keratan sulfate: Structure, biosynthesis, and function. Glycobiology 2000, 10, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Chikada, M.; Yokoyama, M.K.; Kurokawa, M.S.; Ando, T.; Furukawa, H.; Arito, M.; Miyairi, T.; Kato, T. Aberrant Glycosylation of Lumican in Aortic Valve Stenosis Revealed by Proteomic Analysis. Int. Heart J. 2016, 57, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Klejewski, A.; Sterzynska, K.; Wojtowicz, K.; Swierczewska, M.; Partyka, M.; Brazert, M.; Nowicki, M.; Zabel, M.; Januchowski, R. The significance of lumican expression in ovarian cancer drug-resistant cell lines. Oncotarget 2017, 8, 74466–74478. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Yamamoto, T.; Kudo, M.; Kawahara, K.; Kawamoto, M.; Nakajima, Y.; Koizumi, K.; Nakazawa, N.; Ishiwata, T.; Naito, Z. Expression and roles of lumican in lung adenocarcinoma and squamous cell carcinoma. Int. J. Oncol. 2008, 33, 1177–1185. [Google Scholar] [PubMed]
- Ishiwata, T.; Cho, K.; Kawahara, K.; Yamamoto, T.; Fujiwara, Y.; Uchida, E.; Tajiri, T.; Naito, Z. Role of lumican in cancer cells and adjacent stromal tissues in human pancreatic cancer. Oncol. Rep. 2007, 18, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Littlechild, S.L.; Young, R.D.; Caterson, B.; Yoshida, H.; Yamazaki, M.; Sakimura, K.; Quantock, A.J.; Akama, T.O. Keratan Sulfate Phenotype in the beta-1,3-N-Acetylglucosaminyltransferase-7-Null Mouse Cornea. Invest. Ophth. Vis. Sci. 2018, 59, 1641–1651. [Google Scholar] [CrossRef] [PubMed]
- Nagai, N.; Murao, T.; Ito, Y.; Okamoto, N.; Sasaki, M. Enhancing effects of sericin on corneal wound healing in rat debrided corneal epithelium. Biol. Pharm. Bull. 2009, 32, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Stephen Irudayaraj, S.; Sunil, C.; Duraipandiyan, V.; Ignacimuthu, S. Antidiabetic and antioxidant activities of Toddalia asiatica (L.) Lam. leaves in streptozotocin induced diabetic rats. J. Ethnopharmacol. 2012, 143, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Nisha, P.; Mini, S. Flavanoid rich ethyl acetate fraction of Musa paradisiaca inflorescence down-regulates the streptozotocin induced oxidative stress, hyperglycaemia and mRNA levels of selected inflammatory genes in rats. J. Funct. Foods 2013, 5, 1838–1847. [Google Scholar] [CrossRef]
- Nagai, N.; Deguchi, S.; Otake, H.; Hiramatsu, N.; Yamamoto, N. Therapeutic Effect of Cilostazol Ophthalmic Nanodispersions on Retinal Dysfunction in Streptozotocin-Induced Diabetic Rats. Int. J. Mol. Sci. 2017, 18, 1971. [Google Scholar] [CrossRef] [PubMed]
- Takaya, A.; Peng, W.X.; Ishino, K.; Kudo, M.; Yamamoto, T.; Wada, R.; Takeshita, T.; Naito, Z. Cystatin B as a potential diagnostic biomarker in ovarian clear cell carcinoma. Int. J. Oncol. 2015, 46, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, A.; Kudo, M.; Ansai, S.; Peng, W.X.; Ishino, K.; Yamamoto, T.; Wada, R.; Fuji, T.; Teduka, K.; Kawahara, K.; et al. Insulin-like growth factor 2 mRNA-binding protein-3 as a marker for distinguishing between cutaneous squamous cell carcinoma and keratoacanthoma. Int. J. Oncol. 2016, 48, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Kudo, M.; Peng, W.X.; Takata, H.; Takakura, H.; Teduka, K.; Fujii, T.; Mitamura, K.; Taga, A.; Uchida, E.; et al. Identification of aldolase A as a potential diagnostic biomarker for colorectal cancer based on proteomic analysis using formalin-fixed paraffin-embedded tissue. Tumor Biol. 2016, 37, 13595–13606. [Google Scholar] [CrossRef] [PubMed]
- Takata, H.; Kudo, M.; Yamamoto, T.; Ueda, J.; Ishino, K.; Peng, W.X.; Wadai, R.; Taniai, N.; Yoshida, H.; Uchida, E.; et al. Increased expression of PDIA3 and its association with cancer cell proliferation and poor prognosis in hepatocellular carcinoma. Oncol. Lett. 2016, 12, 4896–4904. [Google Scholar] [CrossRef] [PubMed]
- Rada, J.A.; Cornuet, P.K.; Hassell, J.R. Regulation of Corneal Collagen Fibrillogenesis Invitro by Corneal Proteoglycan (Lumican and Decorin) Core Proteins. Exp. Eye Res. 1993, 56, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Svensson, L.; Narlid, I.; Oldberg, A. Fibromodulin and lumican bind to the same region on collagen type I fibrils. FEBS Lett. 2000, 470, 178–182. [Google Scholar] [CrossRef]
- Chakravarti, S. Functions of lumican and fibromodulin: Lessons from knockout mice. Glycoconj. J. 2002, 19, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Vogel, K.G.; Paulsson, M.; Heinegard, D. Specific inhibition of type I and type II collagen fibrillogenesis by the small proteoglycan of tendon. Biochem. J. 1984, 223, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.Y.; Tu, I.H. Adhesion, phenotypic expression, and biosynthetic capacity of corneal keratocytes on surfaces coated with hyaluronic acid of different molecular weights. Acta Biomater. 2012, 8, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Du, Y.; Mann, M.M.; Funderburgh, J.L.; Wagner, W.R. Corneal stromal stem cells versus corneal fibroblasts in generating structurally appropriate corneal stromal tissue. Exp. Eye Res. 2014, 120, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Bendimerad, B. Wound Healing in Partial Denudation of Corneal Epithelium of Diabetic Rat. Kitasato Med. 1987, 17, 179–185. [Google Scholar]
- Vij, N.; Roberts, L.; Joyce, S.; Chakravarti, S. Lumican regulates corneal inflammatory responses by modulating Fas-Fas ligand signaling. Invest. Ophth. Vis. Sci. 2005, 46, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Albig, A.R.; Roy, T.G.; Becenti, D.J.; Schiemann, W.P. Transcriptome analysis of endothelial cell gene expression induced by growth on matrigel matrices: Identification and characterization of MAGP-2 and lumican as novel regulators of angiogenesis. Angiogenesis 2007, 10, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V.; Moscatello, D.K.; McQuillan, D.J.; Eichstetter, I. Decorin is a biological ligand for the epidermal growth factor receptor. J. Biol. Chem. 1999, 274, 4489–4492. [Google Scholar] [CrossRef] [PubMed]
- Moscatello, D.K.; Santra, M.; Mann, D.M.; McQuillan, D.J.; Wong, A.J.; Iozzo, R.V. Decorin suppresses tumor cell growth by activating the epidermal growth factor receptor. J. Clin. Invest. 1998, 101, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Podskochy, A.; Koulikovska, M.; Fagerholm, P.; van der Ploeg, I. Biglycan gene expression in UVR-exposed rabbit corneas. Acta Ophthalmol. Scan. 2004, 82, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Ivorra, M.D.; Paya, M.; Villar, A. A Review of Natural-Products and Plants as Potential Antidiabetic Drugs. J. Ethnopharmacol. 1989, 27, 243–275. [Google Scholar] [CrossRef]
- Arumugam, S.; Sreedhar, R.; Miyashita, S.; Karuppagounder, V.; Thandavarayan, R.A.; Giridharan, V.V.; Pitchaimani, V.; Afrin, R.; Harima, M.; Suzuki, K.; et al. Comparative evaluation of torasemide and furosemide on rats with streptozotocin-induced diabetic nephropathy. Exp. Mol. Pathol. 2014, 97, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Nagai, N.; Ito, Y.; Sasaki, H. Hyperglycemia Enhances the Production of Amyloid beta1-42 in the Lenses of Otsuka Long-Evans Tokushima Fatty Rats, a Model of Human Type 2 Diabetes. Invest. Ophth. Vis. Sci. 2016, 57, 1408–1417. [Google Scholar] [CrossRef] [PubMed]
- Bluemlein, K.; Ralser, M. Monitoring protein expression in whole-cell extracts by targeted label- and standard-free LC-MS/MS. Nat. Protoc. 2011, 6, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Old, W.M.; Meyer-Arendt, K.; Aveline-Wolf, L.; Pierce, K.G.; Mendoza, A.; Sevinsky, J.R.; Resing, K.A.; Ahn, N.G. Comparison of label-free methods for quantifying human proteins by shotgun proteomics. Mol. Cell. Proteom. 2005, 4, 1487–1502. [Google Scholar] [CrossRef] [PubMed]
- Zybailov, B.; Coleman, M.K.; Florens, L.; Washburn, M.P. Correlation of relative abundance ratios derived from peptide ion chromatograms and spectrum counting for quantitative proteomic analysis using stable isotope labeling. Anal. Chem. 2005, 77, 6218–6224. [Google Scholar] [CrossRef] [PubMed]
- Dennis, G., Jr.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, R60. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Huang da, W.; Sherman, B.T.; Zheng, X.; Yang, J.; Imamichi, T.; Stephens, R.; Lempicki, R.A. Extracting biological meaning from large gene lists with DAVID. Curr. Protoc. Bioinf. 2009, 27, 13.11.1–13.11.13. [Google Scholar] [CrossRef] [PubMed]
- Nagai, N.; Fukuoka, Y.; Ishii, M.; Otake, H.; Yamamoto, T.; Taga, A.; Okamoto, N.; Shimomura, Y. Instillation of Sericin Enhances Corneal Wound Healing through the ERK Pathway in Rat Debrided Corneal Epithelium. Int. J. Mol. Sci. 2018, 19, 1123. [Google Scholar] [CrossRef] [PubMed]
- Nagai, N.; Ito, Y.; Okamoto, N.; Shimomura, Y. Size effect of rebamipide ophthalmic nanodispersions on its therapeutic efficacy for corneal wound healing. Exp. Eye Res. 2016, 151, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Nagai, N.; Ogata, F.; Kawasaki, N.; Ito, Y.; Funakami, Y.; Okamoto, N.; Shimomura, Y. Hypercalcemia Leads to Delayed Corneal Wound Healing in Ovariectomized Rats. Biol. Pharm. Bull. 2015, 38, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Nagai, N.; Yoshioka, C.; Tanino, T.; Ito, Y.; Okamoto, N.; Shimomura, Y. Decrease in Corneal Damage due to Benzalkonium Chloride by the Addition of Mannitol into Timolol Maleate Eye Drops. J. Oleo. Sci. 2015, 64, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Terada, S.; Nishimura, T.; Sasaki, M.; Yamada, H.; Miki, M. Sericin, a protein derived from silkworms, accelerates the proliferation of several mammalian cell lines including a hybridoma. Cytotechnology 2002, 40, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Majima, K.; Marunouchi, T. A study of the proliferating activity in lens epithelium and the identification of tissue-type stem cells. Med. Mol. Morphol. 2008, 41, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Deguchi, S.; Otake, H.; Nakazawa, Y.; Hiramatsu, N.; Yamamoto, N.; Nagai, N. Ophthalmic Formulation Containing Nilvadipine Nanoparticles Prevents Retinal Dysfunction in Rats Injected with Streptozotocin. Int. J. Mol. Sci. 2017, 18, 2720. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Accession Number and Description | Fold Change (Rsc) | |
|---|---|---|---|
| 1 | P30427 | Plectin | −2.277 |
| 2 | P69897 | Tubulin beta-5 chain | −2.154 |
| 3 | Q6P9T8 | Tubulin beta-4B chain | −2.062 |
| 4 | P05197 | Elongation factor 2 | −2.062 |
| 5 | P61980 | Heterogeneous nuclear ribonucleoprotein K | −1.505 |
| 6 | P61751 | ADP-ribosylation factor 4 | −1.505 |
| 7 | P62271 | 40S ribosomal protein S18 | −1.117 |
| 8 | P45592 | Cofilin-1 | −1.117 |
| 9 | P49744 | Thrombospondin-4 | 1.111 |
| 10 | P02466 | Collagen alpha-2(I) chain | 1.111 |
| 11 | Q8CJD3 | Zymogen granule membrane protein 16 | 1.111 |
| 12 | P51886 | Lumican | 1.459 |
| 13 | Q9EQP5 | Prolargin | 1.488 |
| 14 | P47853 | Biglycan | 1.740 |
| 15 | Q01129 | Decorin | 2.0601 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, T.; Otake, H.; Hiramatsu, N.; Yamamoto, N.; Taga, A.; Nagai, N. A Proteomic Approach for Understanding the Mechanisms of Delayed Corneal Wound Healing in Diabetic Keratopathy Using Diabetic Model Rat. Int. J. Mol. Sci. 2018, 19, 3635. https://doi.org/10.3390/ijms19113635
Yamamoto T, Otake H, Hiramatsu N, Yamamoto N, Taga A, Nagai N. A Proteomic Approach for Understanding the Mechanisms of Delayed Corneal Wound Healing in Diabetic Keratopathy Using Diabetic Model Rat. International Journal of Molecular Sciences. 2018; 19(11):3635. https://doi.org/10.3390/ijms19113635
Chicago/Turabian StyleYamamoto, Tetsushi, Hiroko Otake, Noriko Hiramatsu, Naoki Yamamoto, Atsushi Taga, and Noriaki Nagai. 2018. "A Proteomic Approach for Understanding the Mechanisms of Delayed Corneal Wound Healing in Diabetic Keratopathy Using Diabetic Model Rat" International Journal of Molecular Sciences 19, no. 11: 3635. https://doi.org/10.3390/ijms19113635
APA StyleYamamoto, T., Otake, H., Hiramatsu, N., Yamamoto, N., Taga, A., & Nagai, N. (2018). A Proteomic Approach for Understanding the Mechanisms of Delayed Corneal Wound Healing in Diabetic Keratopathy Using Diabetic Model Rat. International Journal of Molecular Sciences, 19(11), 3635. https://doi.org/10.3390/ijms19113635