Integrated Analysis of Transcriptomic and Proteomics Data Reveals the Induction Effects of Rotenoid Biosynthesis of Mirabilis himalaica Caused by UV-B Radiation

 ,
,
Abstract
1. Introduction
2. Results
2.1. The Physiological Response of M. himalaica to Different Dose of UV-B Radiation
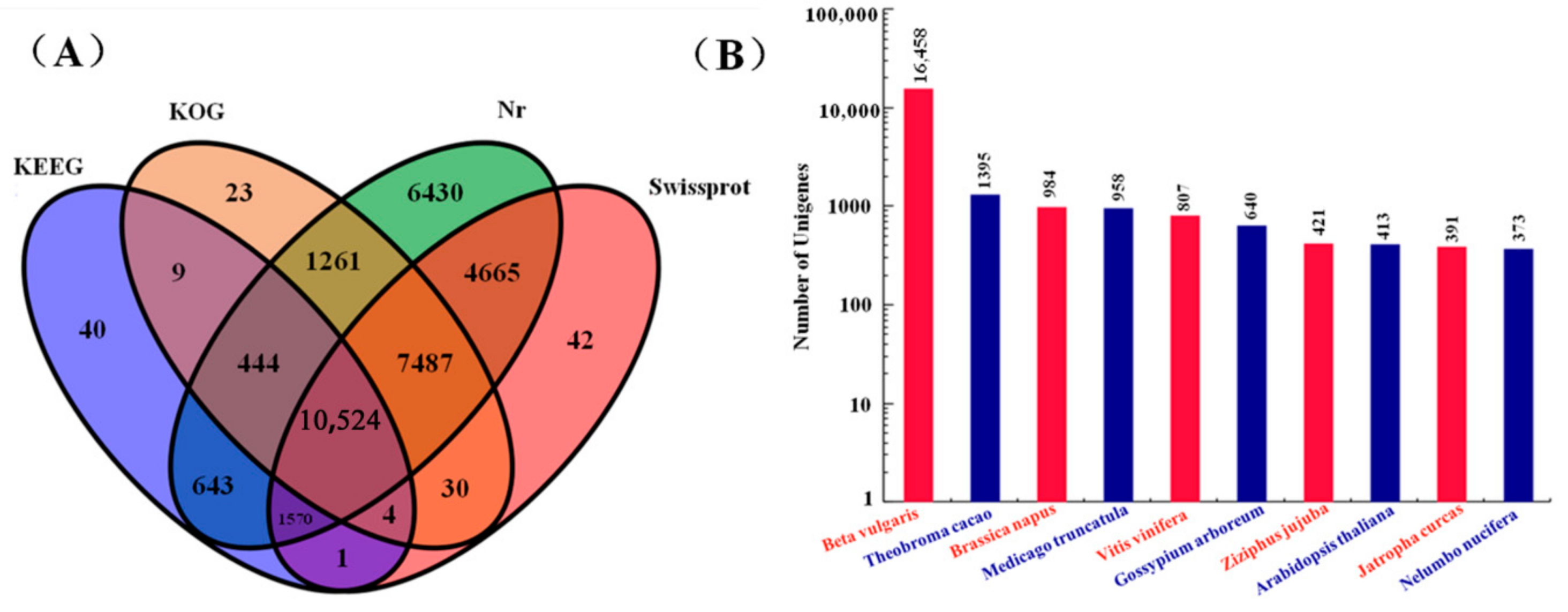
2.2. Construction and Functional Analysis of the Transcriptome Library for M. himalaica
2.3. Identification of Key Genes in Responding to UV-B Radiation by Transcriptome Analysis
2.4. Construction of Protein Library for M. himalaica and Functional Analysis
2.5. Identification of Key Proteins Related to UV-B Radiation
2.6. Combined Analysis of Critical Proteins and Genes Responed to UV-B Radiation
2.7. Quantitative Analysis of Genes Related to of M. himalaica Responses to UV-B Radiation
3. Discussion
4. Materials and Methods
4.1. Test Setup and Collection of Plant Materials
4.2. Physiological Index Determination
4.3. Library Preparation and Illumina Hiseq4000 Sequencing
4.4. De Novo Assembly and Annotation
4.5. Differential Expression Analysis and Candidate Genes Identification
4.6. qRT-PCR Analysis
4.7. Protein Extraction, Digestion and iTRAQ/TMT Labeling
4.8. Strong Cation Exchange (SCX) Fractionation and Liquid Chromatography–Tandem Mass Spectrometry (LC-MS/MS) Analysis
4.9. Database Search
4.10. Protein Functional Annotation and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DEGs | Differentially |
| DEPs | Differentially |
| KEGG | Kyoto |
| GO | Gene |
| MARS | MA-plot-based method with random sampling model |
| FDR | False discovery rate |
| CHS | Chalcone synthase |
| SOD | Superoxide dismutase |
| POD | Peroxidase |
| CAT | Catalase |
| GSH | Glutathione |
| MDA | Malondialdehyde |
| Pro | Proline |
| AUX/IAAs | Auxin-responsive protein IAA |
| GH3 | Gretchen Hagen3 |
| SAUR | Small auxin-up RNA |
| TIFY | TIFY family gene |
| EIN3 | Ethylene insensitive 3 |
| PIP5K | Phosphatidylinositol 4-phosphate 5-kinase |
| DAGK | Diacylglycerol kinase |
| PSY | phytoene synthase |
| CCD | Carotenoid cleavage dioxygenase |
| 4CL | 4-Coumarate-CoA ligase |
| PI | Phosphatidylinositol |
| IP3 | Inositol trisphosphate |
| PKC | Protein kinase C |
References
- Cai, C.P.; Wang, S.L.; Quan, H.; Luo, J.; Guan, F.C.; Lan, X.Z. On morphological dicersity in germplasm resources of Tibetan herbal medicine Mirabilis himalaica. J. Southwest China Norm. Univ. 2013, 38, 61–66. [Google Scholar]
- Lan, X.Z.; Quan, H.; Li, L.Q.; Xia, X.L.; Yin, W.L. Study on the germination properties and quality of seeds of Mirabilis himalaica. Seed 2014, 33, 6–10. [Google Scholar]
- Lu, J.; Lan, X.Z. An investigation on rare and endangered Tibetan medicinal plants in Lhasa region. China J. Chin. Mater. Med. 2013, 38, 127–132. [Google Scholar]
- Guo, L.P.; Zhang, Y.; Zhu, S.D.; Wang, G.H.; Wang, X.; Zhang, X.B.; Chen, M.L.; He, Y.L.; Han, B.X.; Chen, N.F.; et al. Good agricultural practice (GAP) of Chinese materia medica (CMM) for ten years: Achievements, problems and proposals. China J. Chin. Mater. Med. 2014, 39, 1143–1151. [Google Scholar]
- Ma, X.J.; Guo, J.; Tang, J.F.; Ma, X.H.; Ma, Y.; Dai, Z.B. Status and future of natural resource for Chinese materia medica. China J. Chin. Mater. Med. 2015, 40, 1887–1892. [Google Scholar]
- Bais, A.F.; McKenzie, R.L.; Bernhard, G.; Aucamp, P.J.; Ilyas, M.; Madronich, S.; Tourpali, K. Ozone depletion and climate change: Impacts on UV radiation. Photochem. Photobiol. Sci. 2011, 10, 182–198. [Google Scholar] [CrossRef] [PubMed]
- Phoenix, G.K.; Gwynnjones, D.; Lee, J.A.; Callaghan, T.V. Ecological importance of ambient solar ultraviolet radiation to a sub-arctic heath community. Plant Ecol. 2003, 165, 263–273. [Google Scholar] [CrossRef]
- Rakitin, V.Y.; Prudnikova, O.N.; Karyagin, V.V.; Rakitina, T.Y.; Vlasov, P.V.; Borisova, T.A.; Novikova, G.V.; Moshkov, I.E. Ethylene evolution and aba and polyamine contents in arabidopsis thaliana, during UV-B stress. Russ. J. Plant Physiol. 2008, 55, 321–327. [Google Scholar] [CrossRef]
- An, L.Z.; Feng, H.Y.; Wang, X.L. Effects of enhanced UV-B radiation on the growth of some crops. Acta Ecol. Sin. 2001, 21, 249–253. [Google Scholar]
- Kakani, V.G.; Reddy, K.R.; Zhao, D.; Mohammed, A.R. Effects of ultraviolet-b radiation on cotton (Gossypium hirsutum L.) morphology and anatomy. Ann. Bot. 2003, 91, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.A.; Bornman, J.F. UV-B radiation: From generic stressor to specific regulator. Physiol. Plant. 2012, 145, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Si, C.; Yao, X.Q.; He, X.L.; Chu, J.Z.; Ma, C.H.; Shi, X.F. Effects of enhanced UV-B radiation on biochemical traits in postharvest flowers of medicinal chrysanthemum. Photochem. Photobiol. 2015, 91, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.J.; Lin, C.P.; Mau, J.L.; Li, Y.S.; Tsai, S.Y. Effect of UV-B irradiation on physiologically active substance content and antioxidant properties of the medicinal caterpillar fungus Cordyceps militaris (ascomycetes). Int. J. Med. Mushrooms 2015, 17, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P. Influence of laser radiation on quality of folium isatidis and radix Isatidis exposed to enhanced UV-B. Appl. Laser 2005, 25, 336–338. [Google Scholar]
- Pastori, G.M.; Foyer, C.H. Common components, networks, and pathways of cross-tolerance to stress. The central role of “redox” and abscisic acid-mediated controls. Plant Physiol. 2002, 129, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Swindell, W.R.; Huebner, M.; Weber, A.P. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways. BMC Genom. 2007, 8, 125. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, K.K.; Agrawal, S.B. Ultraviolet-B induced changes in morphological, physiological and biochemical parameters of two cultivars of pea (Pisum sativum L.). Ecotoxicol. Environ. Saf. 2014, 100, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Kataria, S.; Jajoo, A.; Guruprasad, K.N. Impact of increasing Ultraviolet-B (UV-B) radiation on photosynthetic processes. J. Photochem. Photobiol. B Biol. 2014, 137, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Rizzini, L.; Favory, J.J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schafer, E.; Nagy, F.; Jenkins, G.I.; et al. Perception of UV-B by the Arabidopsis UVR8 protein. Science 2011, 332, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Hideg, E.; Jansen, M.A.; Strid, A. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.; Ma, L.J.; Bu, N.; Li, Y.Y.; Zhang, L.H. Effects of salicylic acid pre-treatment on cadmium and/or UV-B stress in soybean seedlings. Biol. Plant. 2014, 58, 195–199. [Google Scholar] [CrossRef]
- Tossi, V.; Lamattina, L.; Jenkins, G.I.; Cassia, R.O. Ultraviolet-B-induced stomatal closure in Arabidopsis is regulated by the UV resistance locus8 photoreceptor in a nitric oxide-dependent mechanism. Plant Physiol. 2014, 164, 2220–2230. [Google Scholar] [CrossRef] [PubMed]
- Cloix, C.; Kaiserli, E.; Heilmann, M.; Baxter, K.J.; Brown, B.A.; O’Hara, A.; Smith, B.O.; Christie, J.M.; Jenkins, G.I. C-terminal region of the UV-B photoreceptor UVR8 initiates signaling through interaction with the COP1 protein. Proc. Natl. Acad. Sci. USA 2012, 109, 16366–16370. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Liu, S.; Bao, W.; Xue, X.; Ma, Z.W.; Yokawa, K.; Baluška, F.; Wan, Y.L. Expression of root genes in Arabidopsis seedlings grown by standard and improved growing methods. Int. J. Mol. Sci. 2017, 18, 951. [Google Scholar] [CrossRef] [PubMed]
- Bartley, G.E.; Avena-Bustillos, R.J.; Du, W.X.; Hidalgo, M.; Cain, B.; Breksalll, A.P. Transcriptional regulation of chlorogenic acid biosynthesis in carrot root slices exposed to UV-B light. Plant Gene 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Liscovitch, M.; Cantley, L.C. Lipid second messengers. Cell 1994, 77, 329–334. [Google Scholar] [CrossRef]
- Song, Y.Q.; Yang, Q. Phospholipase in plant defence signaling transduction and development of high plants. J. Harbin Inst. Technol. 2006, 38, 243–246. [Google Scholar]
- Laxalt, A.M.; Munnik, T. Phospholipid signalling in plant defence. Curr. Opin. Plant Biol. 2002, 5, 332–338. [Google Scholar] [CrossRef]
- Chen, Y.G.; Siddhanta, A.; Austin, C.D.; Hammond, S.M.; Sung, T.C.; Frohman, M.A.; Morris, A.J.; Shields, D. Phospholipase d stimulates release of nascent secretory vesicles from the trans-golgi network. J. Cell Biol. 1997, 138, 495–504. [Google Scholar] [CrossRef]
- Chapman, K.D. Phospholipase activity during plant growth and development and in response to environmental stress. Trends Plant Sci. 1998, 3, 419–426. [Google Scholar] [CrossRef]
- Kopka, J.; Pical, C.; Gray, J.E.; Müller-Röber, B. Molecular and enzymatic characterization of three phosphoinositide-specific phospholipase c isoforms from potato. Plant Physiol. 1998, 116, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.J.; Chen, X.; Xue, H.W. Ins(1,4,5)p3 suppresses protein degradation in plant vacuoles by regulating snx-mediated protein sorting. Mol. Plant 2016, 9, 1440–1443. [Google Scholar] [CrossRef] [PubMed]
- Yong, T.; Zhao, C.Y.; Tan, S.T.; Xue, H.W. Arabidopsis type II phosphatidylinositol 4-kinase pi4kγ5 regulates auxin biosynthesis and leaf margin development through interacting with membrane-bound transcription factor anac078. PLoS Genet. 2016, 12, e1006252. [Google Scholar]
- Pappan, K.; Austin-Brown, S.; Chapman, K.D.; Wang, X. Substrate selectivities and lipid modulation of plant phospholipase D alpha, -beta, and -gamma. Arch. Biochem. Biophys. 1998, 353, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Vanhaelewyn, L.; Prinsen, E.; Van, D.S.D.; Vandenbussche, F. Hormone-controlled UV-B responses in plants. J. Exp. Bot. 2016, 67, 4469–4482. [Google Scholar] [CrossRef] [PubMed]
- Hagen, G.; Guilfoyle, T. Auxin-responsive gene expression: Genes, promoters and regulatory factors. Plant Mol. Biol. 2002, 49, 373–385. [Google Scholar] [CrossRef]
- Worley, C.K.; Zenser, N.; Ramos, J.; Rouse, D.; Leyser, O.; Theologis, A.; Callis, J. Degradation of Aux/IAA proteins is essential for normal auxin signalling. Plant J. 2000, 21, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Korasick, D.A.; Westfall, C.S.; Lee, S.G.; Nanao, M.H.; Dumas, R.; Hagen, G.; Guilfoyle, T.J.; Jez, J.M.; Strader, L.C. Molecular basis for auxin response factor protein interaction and the control of auxin response repression. Proc. Natl. Acad. Sci. USA 2014, 111, 5427–5432. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, M.; Yabe, N.; Ichikawa, T.; Yamamoto, Y.Y.; Yoshizumi, T.; Hasunuma, K.; Matsui, M. DFL1, an auxin-responsive GH3 gene homologue, negatively regulates shoot cell elongation and lateral root formation, and positively regulates the light response of hypocotyl length. Plant J. 2001, 25, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Takase, T.; Nakazawa, M.; Ishikawa, A.; Manabe, K.; Matsui, M. DFL2, a new member of the Arabidopsis GH3 gene family, is involved in red light-specific hypocotyl elongation. Plant Cell Physiol. 2003, 44, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Spartz, A.K.; Ren, H.; Park, M.Y.; Grandt, K.N.; Lee, S.H.; Murphy, A.S.; Sussman, M.R.; Overvoorde, P.J.; Gray, W.M. SAUR inhibition of PP2C-D phosphatases activates plasma membrane H+ -ATPases to promote cell expansion in Arabidopsis. Plant Cell 2014, 26, 2129–2142. [Google Scholar] [CrossRef] [PubMed]
- Tossi, V.; Lombardo, C.; Cassia, R.; Lamattina, L. Nitric oxide and flavonoids are systemically induced by UV-B in maize leaves. Plant Sci. 2012, 193, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Bandurska, H.; Cieślak, M. The interactive effect of water deficit and UV-B radiation on salicylic acid accumulation in barley roots and leaves. Environ. Exp. Bot. 2013, 94, 9–18. [Google Scholar] [CrossRef]
- Liu, X.; Chi, H.; Yue, M.; Zhang, X.F.; Li, W.J.; Jia, E.P. The regulation of exogenous jasmonic acid on UV-B stress tolerance in wheat. J. Plant Growth Regul. 2012, 31, 436–447. [Google Scholar] [CrossRef]
- Liu, X.; Chi, H.; Li, Q.; Yue, M.; Li, W.J.; Jia, E.P. Time and concentration effects of exogenous jasmonic acid on antioxidant system of wheat subjected to UV-B radiation. Genom. Appl. Biol. 2015, 34, 2748–2756. [Google Scholar]
- Katerova, Z.; Ivanov, S.; Prinsen, E.; Onckelen, H.V.; Alexieva, V.; Azmi, A. Low doses of ultraviolet-b or ultraviolet-c radiation affect phytohormones in young pea plants. Biol. Plant. 2009, 53, 365–368. [Google Scholar] [CrossRef]
- Yin, P.; Fan, H.; Hao, Q.; Yuan, X.; Wu, D.; Pang, Y.; Yan, C.; Li, W.; Wang, J.; Yan, N. Structural insights into the mechanism of abscisic acid signaling by PYL proteins. Nat. Struct. Mol. Biol. 2009, 16, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Frohnmeyer, H.; Staiger, D. Ultraviolet-B radiation-mediated responses in plants. Balancing damage and protection. Plant Physiol. 2003, 133, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Bassman, J.H. Ecosystem consequences of enhanced solar ultraviolet radiation: Secondary plant metabolites as mediators of multiple trophic interactions in terrestrial plant communities. Photochem. Photobiol. 2004, 79, 382–398. [Google Scholar] [CrossRef] [PubMed]
- Tsoyi, K.; Park, H.B.; Kim, Y.M.; Chung, J.I.; Shin, S.C.; Shim, H.J.; Lee, W.S.; Seo, H.G.; Lee, J.H.; Chang, K.C.; et al. Protective effect of anthocyanins from black soybean seed coats on UV-B-induced apoptotic cell death in vitro and in vivo. J. Agric. Food Chem. 2008, 56, 10600–10605. [Google Scholar] [CrossRef] [PubMed]
- Lavola, A.; Julkunen-Tiitto, R.; Rosa, T.M.D.L.; Lehto, T.; Aphalo, P.J. Allocation of carbon to growth and secondary metabolites in birch seedlings under UV-B radiation and CO2 exposure. Physiol. Plant. 2010, 109, 260–267. [Google Scholar] [CrossRef]
- Zhang, H.; Lv, S.H.; Zhang, W.H. Research advances on responsion molecular mechanisms and signal cascade for plants to enhanced ultraviolet-b radiation. Acta Bot. Boreali 2009, 29, 1932–1937. [Google Scholar]
- Yan, X.F.; Wang, Y.; Li, Y.M. Plant secondary metabolism and its response to environment. Acta Ecol. Sin. 2007, 27, 2554–2562. [Google Scholar]
- Gao, X.; Zhang, X.G.; Zhang, Y.H. Effects of enhanced UV-B radiation on medicinal plant photosynthetic physiology and medicinal active ingredient. Guangdong Chem. Ind. 2015, 42, 112–114. [Google Scholar]
- Khmelenina, V.N.; Rozova, N.; But, C.Y.; Mustakhimov, I.I.; Reshetnikov, A.S.; Beschastnyi, A.P.; Trotsenko, Y.A. Biosynthesis of secondary metabolites in methanotrophs: Biochemical and genetic aspects (review). Prikladnaia Biokhimiia I Mikrobiologiia 2015, 51, 140–150. [Google Scholar] [PubMed]
- Zhou, L.L.; Qi, J.J.; Li, X.E. Effect of enhanced UV-B radiation on yield and qualities of herbal plant Salvia miltiorrhiza bge. Ecol. Environ. 2008, 17, 966–970. [Google Scholar]
- Zhou, X.Y.; Cao, R.X.; Zhang, H.M.; Li, J.J.; Hei, G.G.; Wu, N.B. The response of photosynthetic physiological characters and total alkaloids of Pinellia ternate to enhanced UV-B radiation. J. Southwest Univ. 2013, 35, 166–172. [Google Scholar]
- Gu, L.; Zhang, Z.Y.; Quan, H.; Li, M.J.; Zhao, F.Y.; Xu, Y.J.; Liu, J.; Sai, M.; Zheng, W.L.; Lan, X.Z. Integrated analysis of transcriptomic and metabolomic data reveals critical metabolic pathways involved in rotenoid biosynthesis in the medicinal plant Mirabilis himalaica. Mol. Genet. Genom. 2018, 293, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.Z.; Quan, H.; Xia, X.L.; Yin, W.L.; Zheng, W.L. Molecular cloning and transgenic characterization of the genes encoding chalcone synthase and chalcone isomerase from the Tibetan herbal plant Mirabilis himalaica. Biotechnol. Appl. Biochem. 2016, 63, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Bortul, R.; Tazzari, P.L.; Billi, A.M.; Tabellini, G.; Mantovani, L.; Cappellini, A.; Grafone, T.; Martinelli, G.; Conte, R.; Martelli, A.M. Deguelin, a pi3k/akt inhibitor, enhances chemosensitivity of leukaemia cells with an active pi3k/akt pathway. Br. J. Haematol. 2005, 129, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, J.H.; Jung, K.H.; Hong, S.S. Deguelin promotes apoptosis and inhibits angiogenesis of gastric cancer. Oncol. Rep. 2010, 24, 957–963. [Google Scholar] [PubMed]
- Linghu, L.; Fan, H.X.; Hu, Y.J.; Zou, Y.L.; Yang, P.P.; Lan, X.Z.; Liao, Z.H.; Chen, M. Mirabijalone E: A novel rotenoid from Mirabilis himalaica, inhibited A549 cell growth in vitro and in vivo. J. Ethnopharmacol. 2014, 155, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.Z. Functional Characterization of the Structural Genes Involved in Rotenoid Biosynthesis and Metabolic Engineering Rotenoid Biosynthetic Pathway in Mirabilis himalaica; Beijing Forestry University: Beijing, China, 2015. [Google Scholar]
- Duthie, K.A.; Osborne, L.C.; Foster, L.J.; Abraham, N. Proteomics analysis of interleukin (IL)-7-induced signaling effectors shows selective changes in IL-7Ralpha449F knock-in T cell progenitors. Mol. Cell. Proteom. 2007, 6, 1700–1710. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.R.; Singleton, P.A.; Rowshan, A.; Gucek, M.; Cole, R.N.; Graham, D.R.; Eyk, J.E.N.; Garcia, J.G.N. Quantitative proteomics analysis of human endothelial cell membrane rafts evidence of MARCKS and MRP regulation in the sphingosine 1-phosphate-induced barrier enhancement. Mol. Cell. Proteom. 2007, 6, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.X.; Xu, Z.L.; Zhang, Y.J.; Chai, L.J.; Yi, H.L.; Deng, X.X. An integrative analysis of the transcriptome and proteome of the pulp of a spontaneous late-ripening sweet orange mutant and its wild type improves our understanding of fruit ripening in citrus. J. Exp. Bot. 2014, 65, 1651–1671. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | CK1 | CK2 | CK3 | UV-B1 | UV-B2 | UV-B3 |
|---|---|---|---|---|---|---|
| Raw reads | 50,820,034 | 65,853,560 | 47,847,484 | 58,188,756 | 57,886,202 | 43,004,860 |
| Clean reads | 49,620,668 | 64,102,640 | 46,591,316 | 56,777,882 | 56,443,466 | 42,114,956 |
| Average length (bp) | 150 | 150 | 150 | 150 | 150 | 150 |
| Raw data | 7.62 G | 9.88 G | 7.18 G | 8.73 G | 8.69 G | 6.45 G |
| Clean data | 7.18 G | 9.23 G | 6.70 G | 8.19 G | 8.15 G | 6.20 G |
| Q20 | 97.42% | 97.21% | 97.22% | 97.33% | 97.33% | 98.31% |
| GC content | 43.37% | 45.50% | 45.94% | 45.25% | 44.44% | 43.50% |
| Unigene number | 59,966 | 56,050 | 53,739 | 56,629 | 58,837 | 59,494 |
| Gene ID | log2ratio (UV-B/CK) | Description |
|---|---|---|
| Plant hormone signal transduction | ||
| Unigene0051814 | 1.35 | auxin influx carrier |
| Unigene0008506 | 2.66 | AUX/IAA |
| Unigene0031657 | 1.83 | AUX/IAA |
| Unigene0053509 | 1.76 | AUX/IAA |
| Unigene0047789 | 1.43 | AUX/IAA |
| Unigene0040950 | 1.40 | AUX/IAA |
| Unigene0059280 | 1.77 | GH3 |
| Unigene0004906 | −3.29 | SAUR |
| Unigene0061366 | −1.63 | SAUR |
| Unigene0011781 | −1.16 | SAUR |
| Unigene0054135 | 2.19 | PYL |
| Unigene0042183 | 1.76 | PYL |
| Unigene0050728 | 1.80 | TIFY |
| Unigene0021360 | 1.29 | EIN3 |
| Phosphatidylinositol signaling system | ||
| Unigene0045244 | 4.75 | PIP5K |
| Unigene0023588 | 3.20 | DAGK |
| Unigene0060974 | 11.9 | Calmodulin |
| Unigene0061365 | 11.74 | Calmodulin |
| Unigene0053452 | 10.39 | Calmodulin |
| Unigene0039588 | 1.75 | Calmodulin |
| Unigene0054615 | −1.89 | Calmodulin |
| Flavonoid biosynthesis | ||
| Unigene0055255 | 1.89 | CHS |
| Unigene0017851 | 1.52 | CHS |
| Unigene0016937 | 1.55 | 4CL |
| Carotenoid biosynthesis | ||
| Unigene0029438 | 3.17 | PSY |
| Unigene0021932 | 1.24 | CCD |
| Unigene0055629 | 4.13 | CCD |
| Unigene0040215 | 2.06 | CCD |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, L.; Zheng, W.; Li, M.; Quan, H.; Wang, J.; Wang, F.; Huang, W.; Wu, Y.; Lan, X.; Zhang, Z. Integrated Analysis of Transcriptomic and Proteomics Data Reveals the Induction Effects of Rotenoid Biosynthesis of Mirabilis himalaica Caused by UV-B Radiation. Int. J. Mol. Sci. 2018, 19, 3324. https://doi.org/10.3390/ijms19113324
Gu L, Zheng W, Li M, Quan H, Wang J, Wang F, Huang W, Wu Y, Lan X, Zhang Z. Integrated Analysis of Transcriptomic and Proteomics Data Reveals the Induction Effects of Rotenoid Biosynthesis of Mirabilis himalaica Caused by UV-B Radiation. International Journal of Molecular Sciences. 2018; 19(11):3324. https://doi.org/10.3390/ijms19113324
Chicago/Turabian StyleGu, Li, Weilie Zheng, Mingjie Li, Hong Quan, Jianming Wang, Fengji Wang, Wei Huang, Yunfang Wu, Xiaozhong Lan, and Zhongyi Zhang. 2018. "Integrated Analysis of Transcriptomic and Proteomics Data Reveals the Induction Effects of Rotenoid Biosynthesis of Mirabilis himalaica Caused by UV-B Radiation" International Journal of Molecular Sciences 19, no. 11: 3324. https://doi.org/10.3390/ijms19113324
APA StyleGu, L., Zheng, W., Li, M., Quan, H., Wang, J., Wang, F., Huang, W., Wu, Y., Lan, X., & Zhang, Z. (2018). Integrated Analysis of Transcriptomic and Proteomics Data Reveals the Induction Effects of Rotenoid Biosynthesis of Mirabilis himalaica Caused by UV-B Radiation. International Journal of Molecular Sciences, 19(11), 3324. https://doi.org/10.3390/ijms19113324