The Anticancer Properties of Cordycepin and Their Underlying Mechanisms


Abstract
1. Introduction
2. Cordyceps: A Medicinal Herb
3. An Active Constituent of Cordyceps, Cordycepin, as an Anticancer Agent
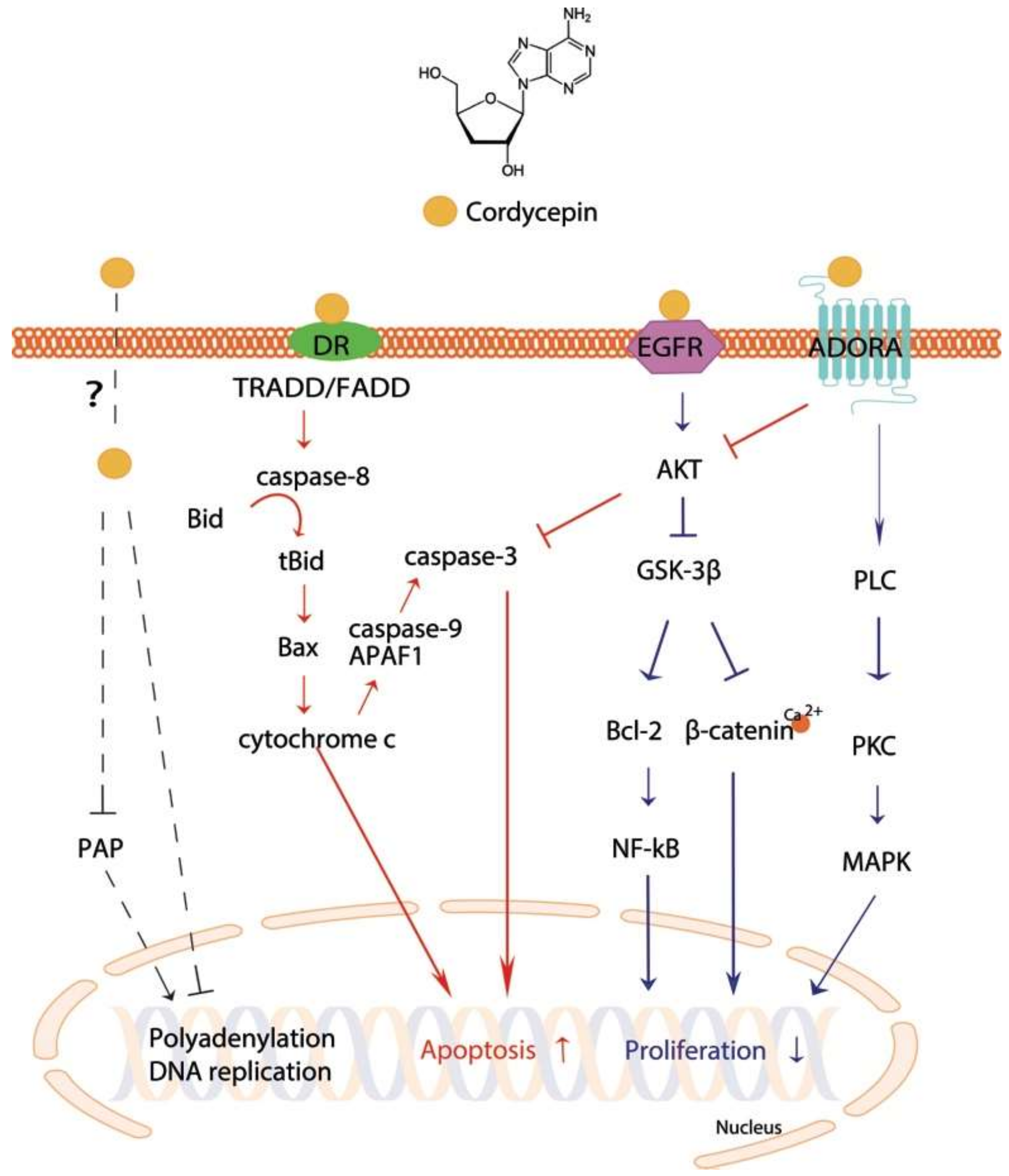
4. Major Mechanisms Underlying the Antitumor Effects of Cordycepin
4.1. Caspase-Mediated Apoptotic Induction
4.2. MAPK-Mediated Apoptotic Induction and Anti-Proliferation
4.3. Anti-Proliferation Effect via GSK-3β Pathway
4.4. Anti-Metastatic Effect Induced by Cordycepin
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADORA | Adenosine receptor |
| APAF1 | Apoptotic protease activating factor 1 |
| Caspases | Cysteine-aspartic proteases |
| cAMP | Cyclic AMP |
| CAV-1 | Caveolin-1 |
| DAG | Diacylglyceride |
| DD | Death domain |
| DR | Death receptor |
| EGFR | Epidermal growth factor receptor |
| ERK | Extracellular signal-regulated kinase |
| FADD | Fas-associated protein with death domain |
| Foxo3a | Forkhead transcription factor |
| GSK-3β | Glycogen synthase kinase 3 beta |
| IP3 | Inositol trisphosphate |
| JNK | c-Jun N-terminal kinase |
| MAPK | Mitogen-activated protein kinase |
| MMP | Matrix metalloproteinase |
| PAP | Polyadenylate polymerase |
| PIP2 | Phosphatidylinositol 4,5-biphosphate |
| PI3K | Phosphoinositide-3 kinase |
| PKA | Protein kinase A |
| PKC | Protein kinase C |
| PLC | Phospholipase C |
| TIMP | Tissue inhibitor of metalloproteinase |
| TRADD | Tumor necrosis factor receptor type 1-associated death domain protein |
References
- Thomadaki, H.; Tsiapalis, C.M.; Scorilas, A. Polyadenylate polymerase modulations in human epithelioid cervix and breast cancer cell lines, treated with etoposide or cordycepin, follow cell cycle rather than apoptosis induction. Biol. Chem. 2005, 386, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Cassileth, B.R. Alternative and complementary cancer treatments. Oncologist 1996, 1, 173–179. [Google Scholar] [PubMed]
- Jeong, J.-W.; Jin, C.-Y.; Park, C.; Hong, S.H.; Kim, G.-Y.; Jeong, Y.K.; Lee, J.-D.; Yoo, Y.H.; Choi, Y.H. Induction of apoptosis by cordycepin via reactive oxygen species generation in human leukemia cells. Toxicol. In Vitro 2011, 25, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Sharma, A.K.; Sandhu, S.S.; Kashyap, D. Cordycepin: A bioactive metabolite with therapeutic potential. Life Sci. 2013, 93, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Yue, K.; Ye, M.; Zhou, Z.; Sun, W.; Lin, X. The genus Cordyceps: A chemical and pharmacological review. J. Pharm. Pharmacol. 2013, 65, 474–493. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.B.; Gerritsma, S.; Yusah, K.M.; Mayntz, D.; Hywel-Jones, N.L.; Billen, J.; Boomsma, J.J.; Hughes, D.P. The life of a dead ant: The expression of an adaptive extended phenotype. Am. Nat. 2009, 174, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Li, Y.; Shen, Y.; Li, Q.; Wang, Q.; Feng, L. Apoptosis and inhibition of proliferation of cancer cells induced by cordycepin. Oncol. Lett. 2015, 10, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y.; Liu, J.; Xu, P.; Zhang, X.M.; Tian, Y.Y.; Xue, Y.M.; Gao, X.Y.; Liu, Y.; Wang, J.H. Synergistic effect of HMGB1 knockdown and cordycepin in the K562 human chronic myeloid leukemia cell line. Mol. Med. Rep. 2015, 12, 4462–4468. [Google Scholar] [CrossRef] [PubMed]
- Song, J.F.; Liu, C.Q.; Li, D.J.; Jin, B.Q. Optimization of cordycepin extraction from cultured Cordyceps militaris by HPLC-DAD coupled with uniform design. J. Chem. Technol. Biotechnol. 2007, 82, 1122–1126. [Google Scholar] [CrossRef]
- Wang, H.-J.; Pan, M.-C.; Chang, C.-K.; Chang, S.-W.; Hsieh, C.-W. Optimization of ultrasonic-assisted extraction of cordycepin from Cordyceps militaris using orthogonal experimental design. Molecules 2014, 19, 20808–20820. [Google Scholar] [CrossRef] [PubMed]
- Sheth, S.; Brito, R.; Mukherjea, D.; Rybak, L.P.; Ramkumar, V. Adenosine receptors: Expression, function and regulation. Int. J. Mol. Sci. 2014, 15, 2024–2052. [Google Scholar] [CrossRef] [PubMed]
- Pao, H.-Y.; Pan, B.-S.; Leu, S.-F.; Huang, B.-M. Cordycepin stimulated steroidogenesis in MA-10 mouse Leydig tumor cells through the protein kinase C pathway. J. Agric. Food Chem. 2012, 60, 4905–4913. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Shinozuka, K.; Yoshikawa, N. Anticancer and antimetastatic effects of cordycepin, an active component of Cordyceps sinensis. J. Pharm. Sci. 2015, 127, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.-L.; Ko, B.-S.; Liu, T.-A.; Liang, S.-M.; Liu, C.-C.; Lu, Y.-J.; Tzean, S.-S.; Shen, T.-L.; Liou, J.-Y. Cordycepin suppresses integrin/FAK signaling and epithelial-mesenchymal transition in hepatocellular carcinoma. Anti-Cancer Agents Med. Chem. 2014, 14, 29–34. [Google Scholar] [CrossRef]
- Li, X.; Zhou, Y.; Zhang, X.; Cao, X.; Wu, C.; Guo, P. Cordycepin stimulates autophagy in macrophages and prevents atherosclerotic plaque formation in ApoE-/-mice. Oncotarget 2017, 8, 94726. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, M.H.; Raoofi Mohseni, S.; Hojjat-Farsangi, M.; Anvari, E.; Ghalamfarsa, G.; Mohammadi, H.; Jadidi-Niaragh, F. Adenosine and adenosine receptors in the immunopathogenesis and treatment of cancer. J. Cell. Physiol. 2018, 233, 2032–2057. [Google Scholar] [CrossRef] [PubMed]
- Kuchta, R.D. Nucleotide Analogues as Probes for DNA and RNA Polymerases. Curr. Protocols Chem. Biol. 2010, 2, 111–124. [Google Scholar]
- Klenow, H. Formation of the mono-, di-and triphosphate of cordycepin in Ehrlich ascites-tumor cells in vitro. Biochim. Biophys. Acta 1963, 76, 347–353. [Google Scholar] [CrossRef]
- Holbein, S.; Wengi, A.; Decourty, L.; Freimoser, F.M.; Jacquier, A.; Dichtl, B. Cordycepin interferes with 3′ end formation in yeast independently of its potential to terminate RNA chain elongation. RNA 2009, 15, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, B.; Goldfinger, B.A.; Marmur, J. Effect of cordycepin triphosphate on the nuclear DNA-dependent RNA polymerases and poly (A) polymerase from the yeast, Saccharomyces cerevisiae. Arch. Biochem. Biophys. 1976, 172, 143–148. [Google Scholar] [CrossRef]
- Müller, W.E.; Seibert, G.; Beyer, R.; Breter, H.J.; Maidhof, A.; Zahn, R.K. Effect of cordycepin on nucleic acid metabolism in L5178Y cells and on nucleic acid-synthesizing enzyme systems. Cancer Res. 1977, 37, 3824–3833. [Google Scholar] [PubMed]
- Rottman, F.; Guarino, A.J. The inhibition of phosphoribosyl-pyrophosphate amidotransferase activity by cordycepin monophosphate. Biochim. Biophys. Acta 1964, 89, 465–472. [Google Scholar] [CrossRef]
- Thomadaki, H.; Scorilas, A.; Tsiapalis, C.M.; Havredaki, M. The role of cordycepin in cancer treatment via induction or inhibition of apoptosis: Implication of polyadenylation in a cell type specific manner. Cancer Chemother. Pharmacol. 2008, 61, 251–265. [Google Scholar] [CrossRef] [PubMed]
- Wahle, E.; Rüegsegger, U. 3′-End processing of pre-mRNA in eukaryotes. FEMS Microbiol. Rev. 1999, 23, 277–295. [Google Scholar] [CrossRef] [PubMed]
- Imesch, P.; Goerens, A.; Fink, D.; Fedier, A. MLH1-deficient HCT116 colon tumor cells exhibit resistance to the cytostatic and cytotoxic effect of the poly (A) polymerase inhibitor cordycepin (3’-deoxyadenosine) in vitro. Oncol. Lett. 2012, 3, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.Y.; Moon, A.; Duffin, R.; Barthet-Barateig, A.; Meijer, H.A.; Clemens, M.J.; de Moor, C.H. Cordycepin inhibits protein synthesis and cell adhesion through effects on signal transduction. J. Biol. Chem. 2010, 285, 2610–2621. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, S.-H.; Hueng, D.-Y.; Syu, J.-P.; Liao, C.-C.; Wu, Y.-C. Cordycepin induces apoptosis of C6 glioma cells through the adenosine 2A receptor-p53-caspase-7-PARP pathway. Chem.-Biol. Interact. 2014, 216, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.-L.; Liu, Z.-J.; Chang, Z. Cordycepin induces apoptosis in human bladder cancer cells via activation of A3 adenosine receptors. Tumor Biol. 2017, 39, 1010428317706915. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Yoshikawa, N.; Yamaguchi, Y.; Kagota, S.; Shinozuka, K.; Kunitomo, M. Antitumor effect of cordycepin (3′-deoxyadenosine) on mouse melanoma and lung carcinoma cells involves adenosine A3 receptor stimulation. Anticancer Res. 2006, 26, 43–47. [Google Scholar] [PubMed]
- Yoshikawa, N.; Yamada, S.; Takeuchi, C.; Kagota, S.; Shinozuka, K.; Kunitomo, M.; Nakamura, K. Cordycepin (3′-deoxyadenosine) inhibits the growth of B16-BL6 mouse melanoma cells through the stimulation of adenosine A 3 receptor followed by glycogen synthase kinase-3β activation and cyclin D 1 suppression. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2008, 377, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Debnath, T.; Kim, S.-K.; Lim, B.O. Anti-cancer effect and apoptosis induction of cordycepin through DR3 pathway in the human colonic cancer cell HT-29. Food Chem. Toxicol. 2013, 60, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Kim, S.O.; Kim, G.-Y.; Moon, S.-K.; Kim, W.-J.; Jeong, Y.K.; Yoo, Y.H.; Choi, Y.H. Involvement of autophagy in cordycepin-induced apoptosis in human prostate carcinoma LNCaP cells. Environ. Toxicol. Pharmacol. 2014, 38, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Y.; Lu, J.; Wang, Y.; Wang, J.; Meng, Q.; Lee, R.J.; Teng, L. Cordycepin, a Natural Antineoplastic Agent, Induces Apoptosis of Breast Cancer Cells via Caspase-dependent Pathways. Nat. Prod. Commun. 2016, 11, 63–68. [Google Scholar] [PubMed]
- Shao, L.W.; Huang, L.H.; Yan, S.; Jin, J.D.; Ren, S.Y. Cordycepin induces apoptosis in human liver cancer HepG2 cells through extrinsic and intrinsic signaling pathways. Oncol. Lett. 2016, 12, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Guo, Z.; Meng, Q.; Lu, J.; Wang, N.; Liu, H.; Liang, Q.; Quan, Y.; Wang, D.; Xie, J. Cordycepin affects multiple apoptotic pathways to mediate hepatocellular carcinoma cell death. Anti-Cancer Agents Med. Chem. 2017, 17, 143–149. [Google Scholar] [CrossRef]
- Lee, H.H.; Jeong, J.-W.; Lee, J.H.; Kim, G.-Y.; Cheong, J.; Jeong, Y.K.; Yoo, Y.H.; Choi, Y.H. Cordycepin increases sensitivity of Hep3B human hepatocellular carcinoma cells to TRAIL-mediated apoptosis by inactivating the JNK signaling pathway. Oncol. Rep. 2013, 30, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; Moon, G.-S.; Jung, K.-H.; Kim, W.-J.; Moon, S.-K. c-Jun N-terminal kinase 1 is required for cordycepin-mediated induction of G2/M cell-cycle arrest via p21WAF1 expression in human colon cancer cells. Food Chem. Toxicol. 2010, 48, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-J.; Kim, S.-K.; Choi, W.-S.; Kim, W.-J.; Moon, S.-K. Cordycepin causes p21WAF1-mediated G2/M cell-cycle arrest by regulating c-Jun N-terminal kinase activation in human bladder cancer cells. Arch. Biochem. Biophys. 2009, 490, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Shichiri, H.; Uda, A.; Yamashita, K.; Nishioka, T.; Kume, M.; Makimoto, H.; Nakagawa, T.; Hirano, T.; Hirai, M. Apoptotic effects of the extracts of cordyceps militaris via Erk phosphorylation in a renal cell carcinoma cell line. Phytother. Res. 2015, 29, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-H.; Joo, J.C.; Kim, D.J.; Jo, E.; Yoo, H.-S.; Lee, K.-B.; Park, S.J.; Jang, I.-S. Cordycepin promotes apoptosis by modulating the ERK-JNK signaling pathway via DUSP5 in renal cancer cells. Am. J. Cancer Res. 2016, 6, 1758. [Google Scholar] [PubMed]
- Joo, J.C.; Hwang, J.H.; Jo, E.; Kim, Y.-R.; Kim, D.J.; Lee, K.-B.; Park, S.J.; Jang, I.-S. Cordycepin induces apoptosis by caveolin-1-mediated JNK regulation of Foxo3a in human lung adenocarcinoma. Oncotarget 2017, 8, 12211. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Park, S.J.; Ko, W.G.; Kang, S.-M.; Lee, D.B.; Bang, J.; Park, B.-J.; Wee, C.-B.; Kim, D.J.; Jang, I.-S. Cordycepin induces human lung cancer cell apoptosis by inhibiting nitric oxide mediated ERK/Slug signaling pathway. Am. J. Cancer Res. 2017, 7, 417. [Google Scholar] [PubMed]
- Tao, X.; Ning, Y.; Zhao, X.; Pan, T. The effects of cordycepin on the cell proliferation, migration and apoptosis in human lung cancer cell lines A549 and NCI-H460. J. Pharm. Pharmacol. 2016, 68, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhong, Q.; Zhang, X.; Hu, D.; He, X.; Li, Q.; Feng, T. Effects of Cordycepin on Proliferation, Apoptosis and NF-κB Signaling Pathway in A549 Cells. Zhong Yao Cai/Zhongyaocai/J. Chin. Med. Mater. 2015, 38, 786–789. [Google Scholar]
- Choi, S.; Lim, M.-H.; Kim, K.M.; Jeon, B.H.; Song, W.O.; Kim, T.W. Cordycepin-induced apoptosis and autophagy in breast cancer cells are independent of the estrogen receptor. Toxicol. Appl. Pharmacol. 2011, 257, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Naura, A.S.; Errami, Y.; Ju, J.; Boulares, A.H. Cordycepin blocks lung injury-associated inflammation and promotes BRCA1-deficient breast cancer cell killing by effectively inhibiting PARP. Mol. Med. 2011, 17, 893. [Google Scholar] [PubMed]
- Wang, Z.; Wu, X.; Liang, Y.-N.; Wang, L.; Song, Z.-X.; Liu, J.-L.; Tang, Z.-S. Cordycepin induces apoptosis and inhibits proliferation of human lung cancer cell line H1975 via Inhibiting the Phosphorylation of EGFR. Molecules 2016, 21, 1267. [Google Scholar] [CrossRef] [PubMed]
- Noh, E.-M.; Jung, S.H.; Han, J.-H.; Chung, E.-Y.; Jung, J.-Y.; Kim, B.-S.; Lee, S.-H.; Lee, Y.-R.; Kim, J.-S. Cordycepin inhibits TPA-induced matrix metalloproteinase-9 expression by suppressing the MAPK/AP-1 pathway in MCF-7 human breast cancer cells. Int. J. Mol. Med. 2010, 25, 255–260. [Google Scholar] [PubMed]
- Tian, T.; Song, L.; Zheng, Q.; Hu, X.; Yu, R. Induction of apoptosis by Cordyceps militaris fraction in human chronic myeloid leukemia K562 cells involved with mitochondrial dysfunction. Pharm. Mag. 2014, 10, 325. [Google Scholar] [CrossRef] [PubMed]
- Ko, B.-S.; Lu, Y.-J.; Yao, W.-L.; Liu, T.-A.; Tzean, S.-S.; Shen, T.-L.; Liou, J.-Y. Cordycepin regulates GSK-3β/β-catenin signaling in human leukemia cells. PLoS ONE 2013, 8, e76320. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Ling, J.; Zhang, G.; Liu, F.; Tao, S.; Han, Z.; Chen, S.; Chen, Z.; Le, H. Cordycepin induces cell cycle arrest and apoptosis by inducing DNA damage and up-regulation of p53 in Leukemia cells. Cell Cycle 2015, 14, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.-S.; Mun, S.-W.; Kim, K.-S.; Park, S.-J.; Yoon, H.-K.; Kim, D.-H.; Park, M.-K.; Kim, C.-H.; Lee, Y.-C. Apoptotic effects of cordycepin through the extrinsic pathway and p38 MAPK activation in human glioblastoma U87MG cells. J. Microbiol. Biotechnol. 2016, 26, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, R.; Zhu, S.; Zhou, R.; Wang, L.; Du, J.; Wang, Y.; Zhou, B.; Mai, L. Cordycepin induces apoptosis and autophagy in human neuroblastoma SK-N-SH and BE (2)-M17 cells. Oncol. Lett. 2015, 9, 2541–2547. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.-S.; Kwon, H.-Y.; Kim, K.-S. Cordycepin induces apoptosis in human neuroblastoma SK-N-BE (2)-C and melanoma SK-MEL-2 cells. Indian J. Biochem. Biophys. 2012, 49, 86–91. [Google Scholar] [PubMed]
- Hsu, P.-Y.; Lin, Y.-H.; Yeh, E.-L.; Lo, H.-C.; Hsu, T.-H.; Su, C.-C. Cordycepin and a preparation from Cordyceps militaris inhibit malignant transformation and proliferation by decreasing EGFR and IL-17RA signaling in a murine oral cancer model. Oncotarget 2017, 8, 93712. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harbor Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Yang, J.; Jones, D. Mitochondrial control of apoptosis: The role of cytochrome c. Biochim. Biophys. Acta 1998, 1366, 139–149. [Google Scholar] [CrossRef]
- Gessi, S.; Merighi, S.; Sacchetto, V.; Simioni, C.; Borea, P.A. Adenosine receptors and cancer. Biochim. Biophys. Acta 2011, 1808, 1400–1412. [Google Scholar] [CrossRef] [PubMed]
- Antonioli, L.; Fornai, M.; Blandizzi, C.; Pacher, P.; Haskó, G. Adenosine signalling and the immune system: When a lot could be too much. Immunol. Lett. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; IJzerman, A.P.; Jacobson, K.A.; Klotz, K.-N.; Linden, J. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol. Rev. 2001, 53, 527–552. [Google Scholar] [PubMed]
- Ramkumar, V.; Stiles, G.; Beaven, M.; Ali, H. The A3 adenosine receptor is the unique adenosine receptor which facilitates release of allergic mediators in mast cells. J. Biol. Chem. 1993, 268, 16887–16890. [Google Scholar] [PubMed]
- Ali, H.; Müller, C.; Daly, J.W.; Beaven, M.A. Methylxanthines block antigen-induced responses in RBL-2H3 cells independently of adenosine receptors or cyclic AMP: Evidence for inhibition of antigen binding to IgE. J. Pharmacol. Exp. Ther. 1991, 258, 954–962. [Google Scholar] [PubMed]
- Waldo, G.L.; Ricks, T.K.; Hicks, S.N.; Cheever, M.L.; Kawano, T.; Tsuboi, K.; Wang, X.; Montell, C.; Kozasa, T.; Sondek, J. Kinetic scaffolding mediated by a phospholipase C–β and Gq signaling complex. Science 2010, 330, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.-A.; Morgan, F.; Watras, J.; Loew, L.M. Analysis of phosphatidylinositol-4, 5-bisphosphate signaling in cerebellar Purkinje spines. Biophys. J. 2008, 95, 1795–1812. [Google Scholar] [CrossRef] [PubMed]
- Milella, M.; Kornblau, S.M.; Estrov, Z.; Carter, B.Z.; Lapillonne, H.; Harris, D.; Konopleva, M.; Zhao, S.; Estey, E.; Andreeff, M. Therapeutic targeting of the MEK/MAPK signal transduction module in acute myeloid leukemia. J. Clin. Investig. 2001, 108, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Fresno, J.V.; Casado, E.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. PI3K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar]
- Fishman, P.; Bar-Yehuda, S.; Liang, B.T.; Jacobson, K.A. Pharmacological and therapeutic effects of A3 adenosine receptor agonists. Drug Discov. Today 2012, 17, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.R.; Wu, Y.-M.; Lonigro, R.J.; Vats, P.; Cobain, E.; Everett, J.; Cao, X.; Rabban, E.; Kumar-Sinha, C.; Raymond, V. Integrative clinical genomics of metastatic cancer. Nature 2017, 548, 297. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.-H.V.; Middleton, K.; You, L.; Sun, Y. A review of microfluidic approaches for investigating cancer extravasation during metastasis. Microsyst. Nanoeng. 2018, 4, 17104. [Google Scholar] [CrossRef]


{kind=link}
| Tumor Type | Effects | Major Mediating Signaling Pathways | Results | References |
|---|---|---|---|---|
| Liver cancer | Apoptosis induction | Cysteine-aspartic proteases (caspase) | - Increased caspase-3, caspase-8, caspase-9, FADD, and Bid - Decreased Bcl-2 | [34] |
| Apoptosis induction, Anti-proliferation | Phospholipase C (PLC) | - Increased PARP cleavage | [35,36] | |
| Colon cancer | Cell cycle arrest | c-Jun N-terminal kinases (JNK) | - G2/M-phase cell-cycle arrest - Increased JNK and p21WAF-1 expression | [37] |
| Bladder cancer | Cell cycle arrest | c-Jun N-terminal kinases (JNK) | - G2/M cell-cycle arrest - Increased cJNK | [38] |
| Renal cancer | Apoptosis induction | ERK/JNK | - Increased JNK and caspase-3 - Decreased ERK | [39,40] |
| Lung cancer | Apoptosis induction | ERK/JNK | - Increased CAV1, JNK/Foxo3a, BAX, caspase-3 cleavage, and GSK-3β - Decreased ERK | [41,42] |
| Apoptosis induction, Anti-proliferation, Anti-metastasis | Caspase | - Increased MMP-9 and caspase-3 - Decreased Bcl-2 | [43] | |
| Apoptosis induction, Anti-proliferation | NF-kB | - Increased BAX, caspase-3 cleavage - Decreased NF-kB, Bcl-2, and p65 nuclear translocation | [44] | |
| Breast cancer | Apoptosis induction | Caspase | - Increased caspase-3, caspase-9, mitochondrial translocation of BAX, and cytochrome c | [45,46,47] |
| Anti-metastasis | MAPK | - Decreased Bcl-2 | [48] | |
| Prostate cancer | Apoptosis induction | Caspase | - Increased JNK and caspase-3 - Decreased ERK | [39,40] |
| Leukemia | Apoptosis induction | Caspase | - Increased caspase-3, caspase-8, and PARP cleavage - Decreased Bcl-2 | [3,49] |
| Anti-proliferation | GSK-3𝛽 | - Decreased Akt, and β-catenine | [50] | |
| Apoptosis induction, Cell cycle arrest | MAPK | - S-phase cell-cycle arrest - Increased cytochrome c released, and caspase-9 - Decreased MAPK, p-ERK1/2, and CDK2 expression | [51] | |
| Glioblastoma | Apoptosis induction Anti-proliferation | MAPK | - Increased p38 MAPK, caspase-3,8, PARP cleavage - Decreased Akt and Bcl-2 | [52] |
| Neuroblastoma | Apoptosis induction Cell cycle arrest | Caspase | - sub-G1 phase cell-cycle arrest - Increased caspase-3 and PARP cleavage | [53,54] |
| Receptors | Downstream Targets | References | |
|---|---|---|---|
| Adenosine receptors (ADORAs) | ADORA2A | Induced apoptosis via ADORA2A receptor-p53-caspase-7-PARP pathway in C6 glioma cells. | [27] |
| ADORA3 | Inhibition of the tumor growth was antagonized by a selective adenosine A3 receptor agonist, MRS1191 1, in mouse melanoma and lung carcinoma cells via the radioligand binding assay. The involvement of adenosine A3 receptors was confirmed with the use of MRS1523 2 and MRS1220 3. | [28,29,30] | |
| Death receptors (DRs) | DR3 | Induced apoptosis of the human colon cancer cell HT-29 via the elevation of DR3, caspase-8, caspase-1, cleaved caspase-3 and cleaved PARP expression. | [31] |
| DR5 | Caused an elevation of the levels of Fas, death receptor 5 (DR5), proapoptotic BAX, and decreased anti-apoptotic Bcl-2 level. | [32] | |
| Epidermal growth factor receptor (EGFR) | Inhibited cell proliferation and induced apoptosis via the EGFR signaling pathway in the human lung cancer cell H1975. The caspase-3 and BAX protein levels were increased, while phosphorylated expression levels of EGFR, AKT, and ERK1/2 were decreased. | [47,55] | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, S.Y.; Park, S.J.; Park, Y.J. The Anticancer Properties of Cordycepin and Their Underlying Mechanisms. Int. J. Mol. Sci. 2018, 19, 3027. https://doi.org/10.3390/ijms19103027
Yoon SY, Park SJ, Park YJ. The Anticancer Properties of Cordycepin and Their Underlying Mechanisms. International Journal of Molecular Sciences. 2018; 19(10):3027. https://doi.org/10.3390/ijms19103027
Chicago/Turabian StyleYoon, So Young, Soo Jung Park, and Yoon Jung Park. 2018. "The Anticancer Properties of Cordycepin and Their Underlying Mechanisms" International Journal of Molecular Sciences 19, no. 10: 3027. https://doi.org/10.3390/ijms19103027
APA StyleYoon, S. Y., Park, S. J., & Park, Y. J. (2018). The Anticancer Properties of Cordycepin and Their Underlying Mechanisms. International Journal of Molecular Sciences, 19(10), 3027. https://doi.org/10.3390/ijms19103027