Adaptive Evolution of the Eda Gene and Scales Loss in Schizothoracine Fishes in Response to Uplift of the Tibetan Plateau

 , ,
, ,  , and
, and
Abstract

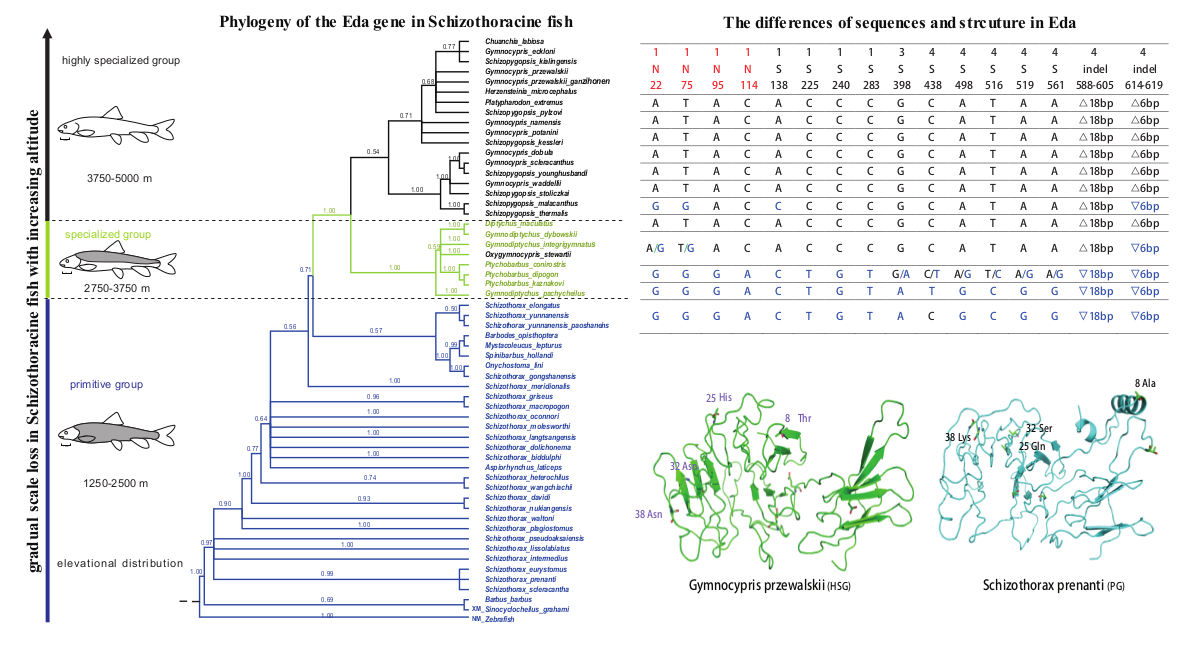{kind=link}
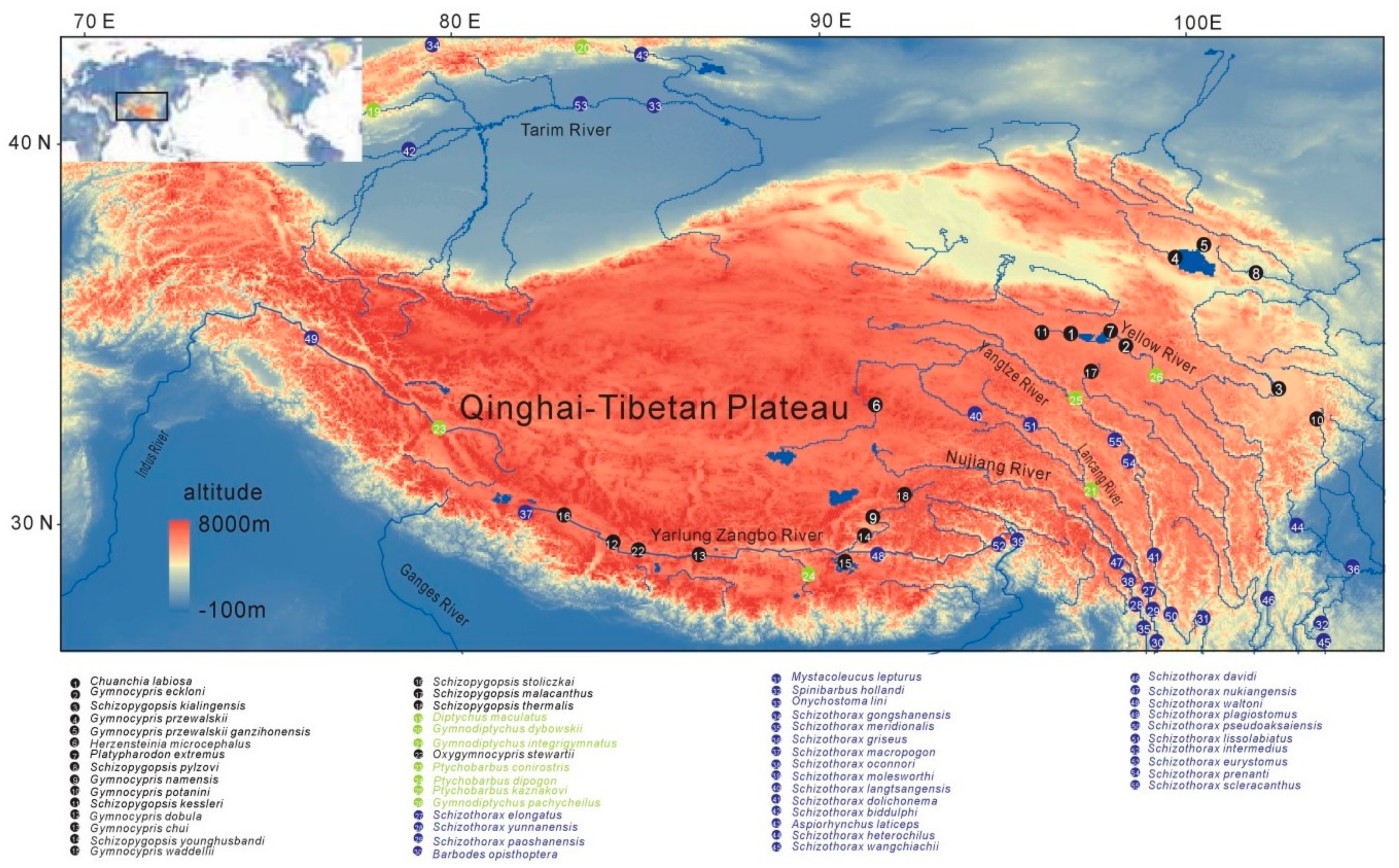{kind=link}
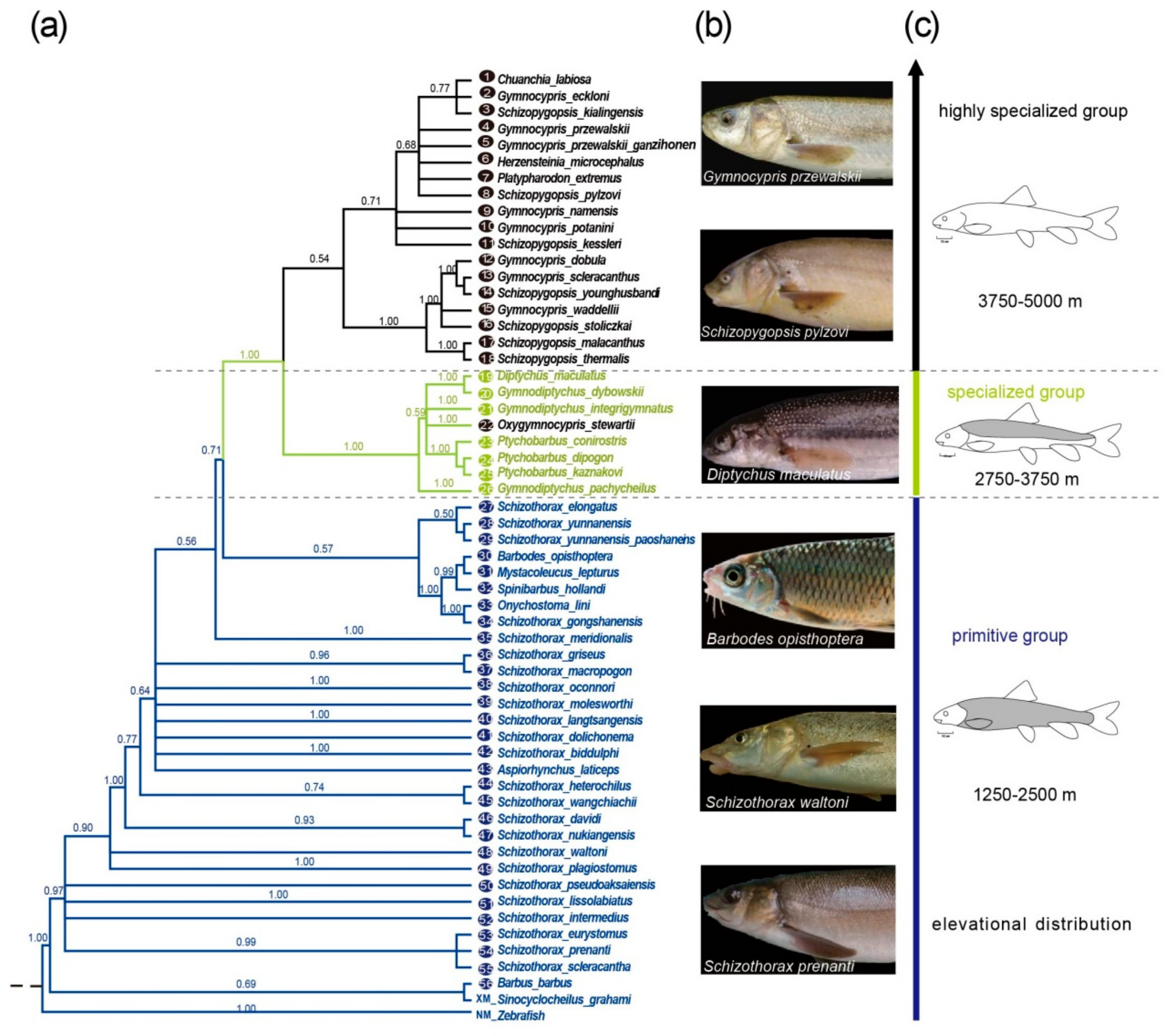{kind=link}
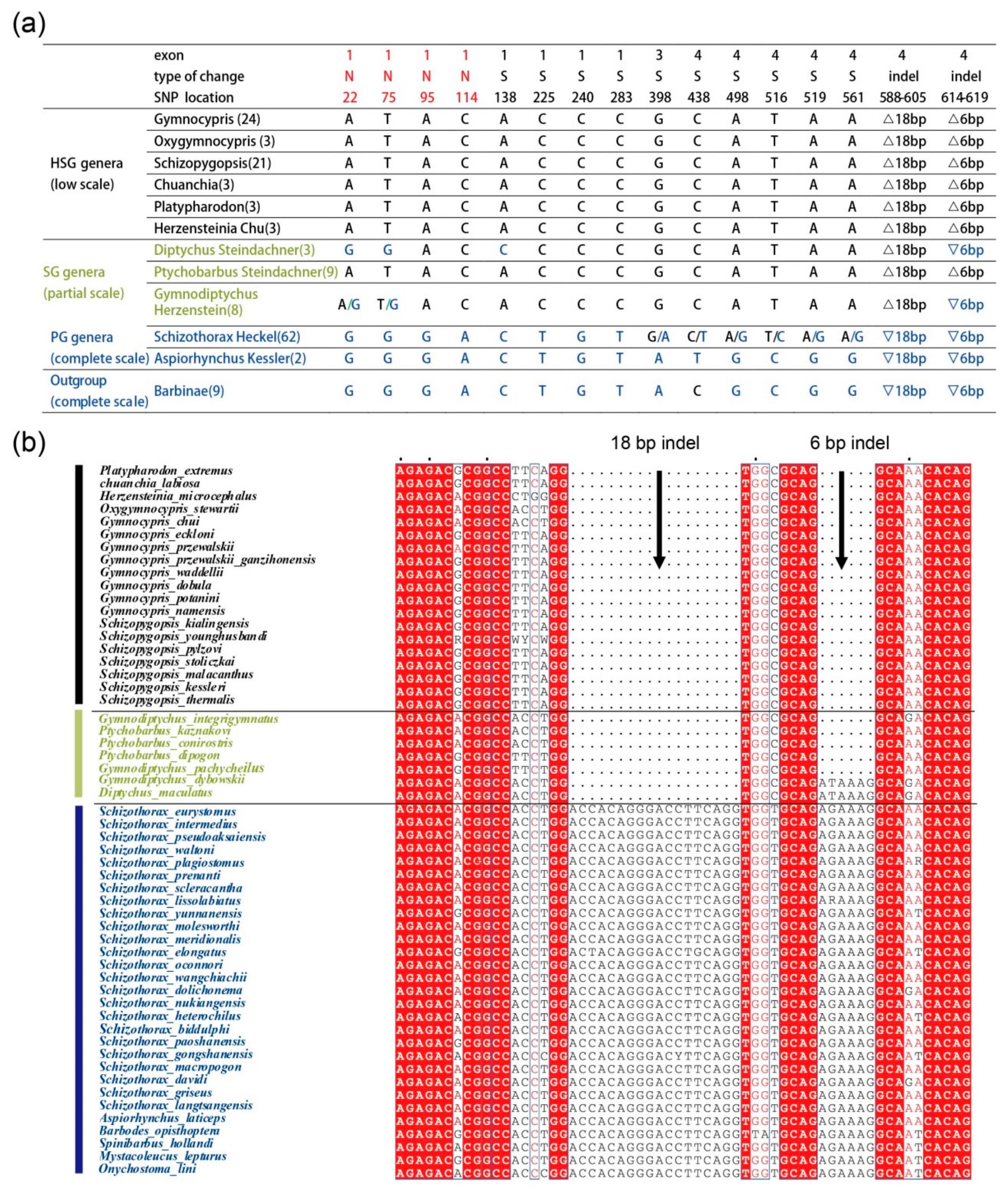{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Eda Genes of 51 Schizothoracine Species and Five Barbinae Species
2.2. Phylogenetic Reconstruction of the Eda Genes of Three Schizothoracine Groups
2.3. Multiple Sequence Alignment of the Eda Gene in Schizothoracine and Barbus Species
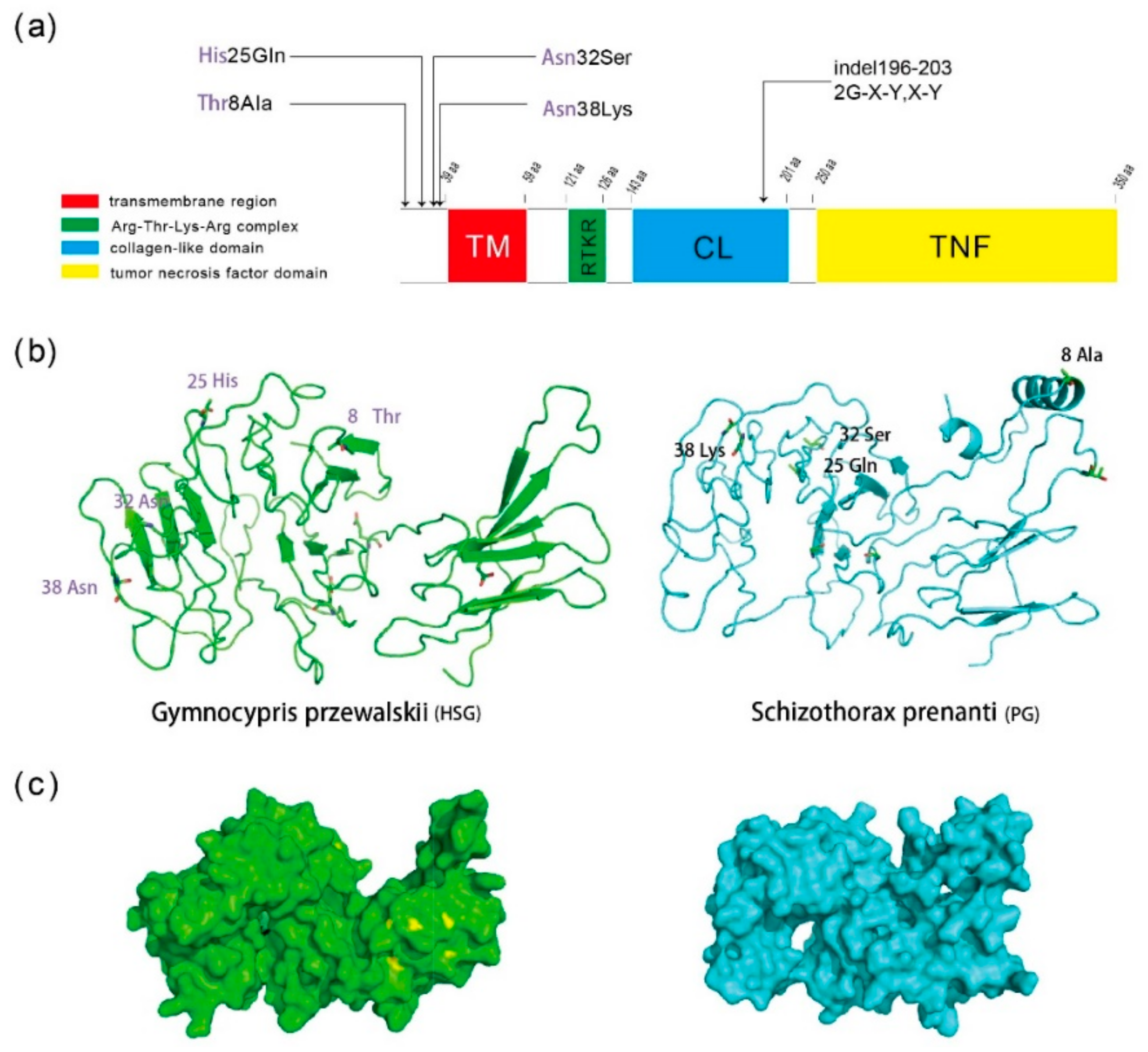
2.4. Homologous Sequence Alignment and Structure Prediction of the Eda Protein
3. Discussion
4. Materials and Methods
4.1. Fish Sampling, RNA and Genomic DNA Extraction
4.2. Eda Sequencing and Assembly Strategy
4.3. Multiple Sequence Alignments and Phylogenetic Analysis
4.4. Eda Protein Structure Analysis
Supplementary Materials
Data Accessibility
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, J.; Fang, X. Uplift of the tibetan plateau and environmental changes. Chin. Sci. Bull. 1999, 44, 2117–2124. [Google Scholar] [CrossRef]
- Li, J.J.; Wen, S.X.; Zhang, X.S.; Wang, F.B.; Zhan, B.X.; Li, B.X. A discussion on the period, amplitude and type of the uplift of the qinghai-xizang plateau. Sci. Chin. Ser. A 1979, 22, 608–616. [Google Scholar]
- Shi, Y. Uplift and Environmental Changes of Qinghai-Xizang (Tibetan) Plateau in the Late Cenozoic; Guangdong Scirence & Technology Press: Guangdong, China, 1998. [Google Scholar]
- Wen, L. Uplift of the tibetan plateau influenced the morphological evolution of animals. J. Agric. Sci. 2014, 6, 244. [Google Scholar] [CrossRef]
- Ma, X.; Lu, X.; Merila, J. Altitudinal decline of body size in a tibetan frog. J. Zool. 2009, 279, 364–371. [Google Scholar] [CrossRef]
- Jin, Y.T.; Li, J.Q.; Liu, N.F. Elevation-related variation in life history traits among Phrynocephalus lineages on the tibetan plateau: Do they follow typical squamate ecogeographic patterns? J. Zool. 2013, 290, 293–301. [Google Scholar] [CrossRef]
- Hao, Z.; Changxin, W.; Yangzom, C.; Yao, L.; Suling, J. Influence of altitude on growth curves in tibetan chicken and its hybrid. J. Chin. Agric. Univ. 2007, 12, 40. [Google Scholar]
- Zhang, H.; Wu, C.; Chamba, Y.; Ling, Y.; Li, J. Influence of altitude on weight and carcass characteristics in tibetan chicken and its crossing combination. Acta Ecol. Anim. Domastic 2007, 28, 48–52. [Google Scholar]
- Zhang, T.Z.; Nevo, E.; Tang, L.Z.; Su, J.P.; Lin, G.H. Plateau zokors on the qinghai-tibetan plateau follow bergmann’s rule latitudinally, but not altitudinally. Mamm. Biol. 2012, 77, 108–112. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Takami, Y.; Okuzaki, Y.; Sota, T. Genetic differences and phenotypic plasticity in body size between high- and low-altitude populations of the ground beetle carabus tosanus. J. Evol. Biol. 2012, 25, 1835–1842. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.X.; Chen, Y.Y.; Wu, Y.F.; Zhu, S.Q. The origin and evolution of subfamily schizothoracinae and its relationship with the uplift of the tibetan plateau. In The Era of the Uplift of the Qinghai Tibet Plateau, Amplitude and Form Issues; Science Press: Beijing, China, 1981; pp. 118–130. [Google Scholar]
- Zhu, S. The Loaches of the Subufamily Nemacheilinae in China; Jiangsu Publishing House of Science and Technology: Nanjing, China, 1989; pp. 31–38. [Google Scholar]
- Jin, Y.T.; Tian, R.R. Altitudinal variations of morphological characters of Phrynocephalus sand lizards: On the validity of bergmann’s and allen’s rules. Acta Zool. Sin. 2006, 52, 838–845. [Google Scholar]
- Liao, J.; Zhang, Z.; Liu, N. Effects of altitudinal change on the auditory bulla in Ochotona daurica (mammalia, lagomorpha). J. Zool. Syst. Evol. Res. 2007, 45, 151–154. [Google Scholar] [CrossRef]
- Liao, J.; Liu, N. Altitudinal variations of acoustic organs in anurans: A case study from china. Ital. J. Zool. 2008, 75, 125–134. [Google Scholar] [CrossRef]
- He, D.; Chen, Y. Molecular phylogeny and biogeography of the highly specialized grade schizothoracine fishes (teleostei: Cyprinidae) inferred from cytochrome b sequences. Chin. Sci. Bull. 2007, 52, 777–788. [Google Scholar] [CrossRef]
- Qi, D.L.; Chao, Y.; Guo, S.C.; Zhao, L.Y.; Li, T.P.; Wei, F.L.; Zhao, X.Q. Convergent, parallel and correlated evolution of trophic morphologies in the subfamily schizothoracinae from the qinghai-tibetan plateau. PLoS ONE 2012, 7, e34070. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Duan, Z.Y.; Peng, Z.G.; Guo, S.C.; Li, J.B.; He, S.P.; Zhao, X.Q. The youngest split in sympatric schizothoracine fish (cyprinidae) is shaped by ecological adaptations in a tibetan plateau glacier lake. Mol. Ecol. 2009, 18, 3616–3628. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Chen, Y.; Chen, Y.; Chen, Z. Molecular phylogeny of the specialized schizothoracine fishes (teleostei: Cyprinidae), with their implications for the uplift of the qinghai-tibetan plateau. Chin. Sci. Bull. 2004, 49, 39–48. [Google Scholar] [CrossRef]
- Tong, C.; Fei, T.; Zhang, C.; Zhao, K. Comprehensive transcriptomic analysis of tibetan schizothoracinae fish gymnocypris przewalskii reveals how it adapts to a high altitude aquatic life. BMC Evol. Biol. 2017, 17, 74. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Tian, F.; Zhao, K. Genomic signature of highland adaptation in fish: A case study in tibetan schizothoracinae species. BMC Genom. 2017, 18, 948. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gan, X.; Li, J.; Chen, Y.; He, S. Cyprininae phylogeny revealed independent origins of the tibetan plateau endemic polyploid cyprinids and their diversifications related to the neogene uplift of the plateau. Sci. China Life Sci. 2016, 59, 1149–1165. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, T.; Hasegawa, M.; Zhong, Y. Polyphyletic origins of schizothoracine fish (cyprinidae, osteichthyes) and adaptive evolution in their mitochondrial genomes. Genes Genet. Syst. 2014, 89, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.Y.; Schlessinger, D. EDA signaling and skin appendage development. Cell Cycle 2006, 5, 2477–2483. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk-Quintas, C.; Schneider, P. Ectodysplasin a (EDA)—EDA receptor signalling and its pharmacological modulation. Cytokine Growth Factor Rev. 2014, 25, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Cluzeau, C.; Hadj-Rabia, S.; Jambou, M.; Mansour, S.; Guigue, P.; Masmoudi, S.; Bal, E.; Chassaing, N.; Vincent, M.C.; Viot, G.; et al. Only four genes (EDA1, EDAR, EDARADD, and WNT10A) account for 90% of hypohidrotic/anhidrotic ectodermal dysplasia cases. Hum. Mutat. 2011, 32, 70–72. [Google Scholar] [CrossRef] [PubMed]
- Koguchi-Yoshioka, H.; Wataya-Kaneda, M.; Yutani, M.; Murota, H.; Nakano, H.; Sawamura, D.; Katayama, I. Atopic diathesis in hypohidrotic/anhidrotic ectodermal dysplasia. Acta Derm.-Venereol. 2015, 95, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.K.; Pispa, J.; Hartung, A.J.; Du, Y.Z.; Ezer, S.; Jenks, T.; Shimada, T.; Pekkanen, M.; Mikkola, M.L.; Ko, M.S.H.; et al. The tabby phenotype is caused by mutation in a mouse homologue of the EDA gene that reveals novel mouse and human exons and encodes a protein (ectodysplasin-A) with collagenous domains. Proc. Natl. Acad. Sci. USA 1997, 94, 13069–13074. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.P.; Rohner, N.; Schwarz, H.; Perathoner, S.; Konstantinidis, P.; Nusslein-Volhard, C. Zebrafish EDA and EDAR mutants reveal conserved and ancestral roles of ectodysplasin signaling in vertebrates. PLoS Genet. 2008, 4, e1000206. [Google Scholar] [CrossRef] [PubMed]
- Colosimo, P.F.; Hosemann, K.E.; Balabhadra, S.; Villarreal, G.; Dickson, M.; Grimwood, J.; Schmutz, J.; Myers, R.M.; Schluter, D.; Kingsley, D.M. Widespread parallel evolution in sticklebacks by repeated fixation of ectodysplasin alleles. Science 2005, 307, 1928–1933. [Google Scholar] [CrossRef] [PubMed]
- Conte, G.L.; Arnegard, M.E.; Peichel, C.L.; Schluter, D. The probability of genetic parallelism and convergence in natural populations. Proc. R. Soc. B 2012, 279, 5039–5047. [Google Scholar] [CrossRef] [PubMed]
- Elmer, K.R.; Meyer, A. Adaptation in the age of ecological genomics: Insights from parallelism and convergence. Trends Ecol. Evol. 2011, 26, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Orgogozo, V. The loci of repeated evolution: A catalog of genetic hotspots of phenotypic variation. Evol. Int. J. Org. Evol. 2013, 67, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- Stern, D.L. The genetic causes of convergent evolution. Nat. Rev. Genet. 2013, 14, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.E.; Burke, J.M.; Rieseberg, L.H. Parallel genotypic adaptation: When evolution repeats itself. Genetica 2005, 123, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.A. Ecology—Phenotypic plasticity in the interactions and evolution of species. Science 2001, 294, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Yang, X.; Tang, Z. Patterns of species diversity and phylogenetic structure of vascular plants on the qinghai-tibetan plateau. Ecol. Evol. 2013, 3, 4584–4595. [Google Scholar] [CrossRef] [PubMed]
- Bell, G. Fluctuating selection: The perpetual renewal of adaptation in variable environments. Philos. Trans. R. Soc. B 2010, 365, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chiang, T.Y.; George, M.; Liu, J.Q.; Abbott, R.J. Phylogeography of the qinghai-tibetan plateau endemic Juniperus przewalskii (cupressaceae) inferred from chloroplast DNA sequence variation. Mol. Ecol. 2005, 14, 3513–3524. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chen, X.; Bai, J.; Fang, D.; Qiu, Y.; Jiang, W.; Yuan, H.; Bian, C.; Lu, J.; He, S. The sinocyclocheilus cavefish genome provides insights into cave adaptation. BMC Biol. 2016, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Britannica, E. Britannica Concise Encyclopedia; Foreign Language Education Press: Shanghai, China, 2008; p. 7. [Google Scholar]
- Di Lullo, G.A.; Sweeney, S.M.; Korkko, J.; Ala-Kokko, L.; San Antonio, J.D. Mapping the ligand-binding sites and disease-associated mutations on the most abundant protein in the human, type I collagen. J. Boil. Chem. 2002, 277, 4223–4231. [Google Scholar] [CrossRef] [PubMed]
- Ezer, S.; Bayés, M.; Schlessinger, D.; Kere, J. Ectodysplasin is a collagenous trimeric type II membrane protein with a tumor necrosis factor-like domain and co-localizes with cytoskeletal structures at lateral and apical surfaces of cells. Hum. Mol. Genet. 1999, 8, 2079–2086. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schneider, P.; Street, S.L.; Gaide, O.; Hertig, S.; Tardivel, A.; Tschopp, J.; Runkel, L.; Alevizopoulos, K.; Ferguson, B.M.; Zonana, J. Mutations leading to X-linked hypohidrotic ectodermal dysplasia affect three major functional domains in the tumor necrosis factor family member ectodysplasin-A. J. Boil. Chem. 2001, 276, 18819–18827. [Google Scholar] [CrossRef] [PubMed]
- Swee, L.K.; Ingold-Salamin, K.; Tardivel, A.; Willen, L.; Gaide, O.; Favre, M.; Demotz, S.; Mikkola, M.; Schneider, P. Biological activity of ectodysplasin a is conditioned by its collagen and heparan sulfate proteoglycan-binding domains. J. Boil. Chem. 2009, 284, 27567–27576. [Google Scholar] [CrossRef] [PubMed]
- Bayes, M.; Hartung, A.J.; Ezer, S.; Pispa, J.; Thesleff, I.; Srivastava, A.K.; Kere, J. The anhidrotic ectodermal dysplasia gene (EDA) undergoes alternative splicing and encodes ectodysplasin-A with deletion mutations in collagenous repeats. Hum. Mol. Genet. 1998, 7, 1661–1669. [Google Scholar] [CrossRef] [PubMed]
- Paakkonen, K.; Cambiaghi, S.; Novelli, G.; Ouzts, L.V.; Penttinen, M.; Kere, J.; Srivastava, A.K. The mutation spectrum of the EDA gene in X-linked anhidrotic ectodermal dysplasia. Hum. Mutat. 2001, 17, 349. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ludwig, A.; Zhang, C.; Tong, C.; Li, G.; Tang, Y.; Peng, Z.; Zhao, K. Local adaptation of Gymnocypris przewalskii (cyprinidae) on the tibetan plateau. Sci. Rep. 2015, 5, 9780. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, D.; Michalak, I. Raxmlgui: A graphical front-end for raxml. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Alexander, N.; Woetzel, N.; Meiler, J. Bcl::Cluster: A method for clustering biological molecules coupled with visualization in the pymol molecular graphics system. In Proceedings of the 2011 IEEE 1st International Conference on Computational Advances in Bio and Medical Sciences (ICCABS), Orlando, FL, USA, 3–5 February 2011; pp. 13–18. [Google Scholar]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Tong, C.; Ludwig, A.; Tang, Y.; Liu, S.; Zhang, R.; Feng, C.; Li, G.; Peng, Z.; Zhao, K. Adaptive Evolution of the Eda Gene and Scales Loss in Schizothoracine Fishes in Response to Uplift of the Tibetan Plateau. Int. J. Mol. Sci. 2018, 19, 2953. https://doi.org/10.3390/ijms19102953
Zhang C, Tong C, Ludwig A, Tang Y, Liu S, Zhang R, Feng C, Li G, Peng Z, Zhao K. Adaptive Evolution of the Eda Gene and Scales Loss in Schizothoracine Fishes in Response to Uplift of the Tibetan Plateau. International Journal of Molecular Sciences. 2018; 19(10):2953. https://doi.org/10.3390/ijms19102953
Chicago/Turabian StyleZhang, Cunfang, Chao Tong, Arne Ludwig, Yongtao Tang, Sijia Liu, Renyi Zhang, Chenguang Feng, Guogang Li, Zuogang Peng, and Kai Zhao. 2018. "Adaptive Evolution of the Eda Gene and Scales Loss in Schizothoracine Fishes in Response to Uplift of the Tibetan Plateau" International Journal of Molecular Sciences 19, no. 10: 2953. https://doi.org/10.3390/ijms19102953
APA StyleZhang, C., Tong, C., Ludwig, A., Tang, Y., Liu, S., Zhang, R., Feng, C., Li, G., Peng, Z., & Zhao, K. (2018). Adaptive Evolution of the Eda Gene and Scales Loss in Schizothoracine Fishes in Response to Uplift of the Tibetan Plateau. International Journal of Molecular Sciences, 19(10), 2953. https://doi.org/10.3390/ijms19102953