Parathyroid Hormone (PTH) Increases Skeletal Tumour Growth and Alters Tumour Distribution in an In Vivo Model of Breast Cancer


 , ,
, ,
Abstract
1. Introduction
2. Results
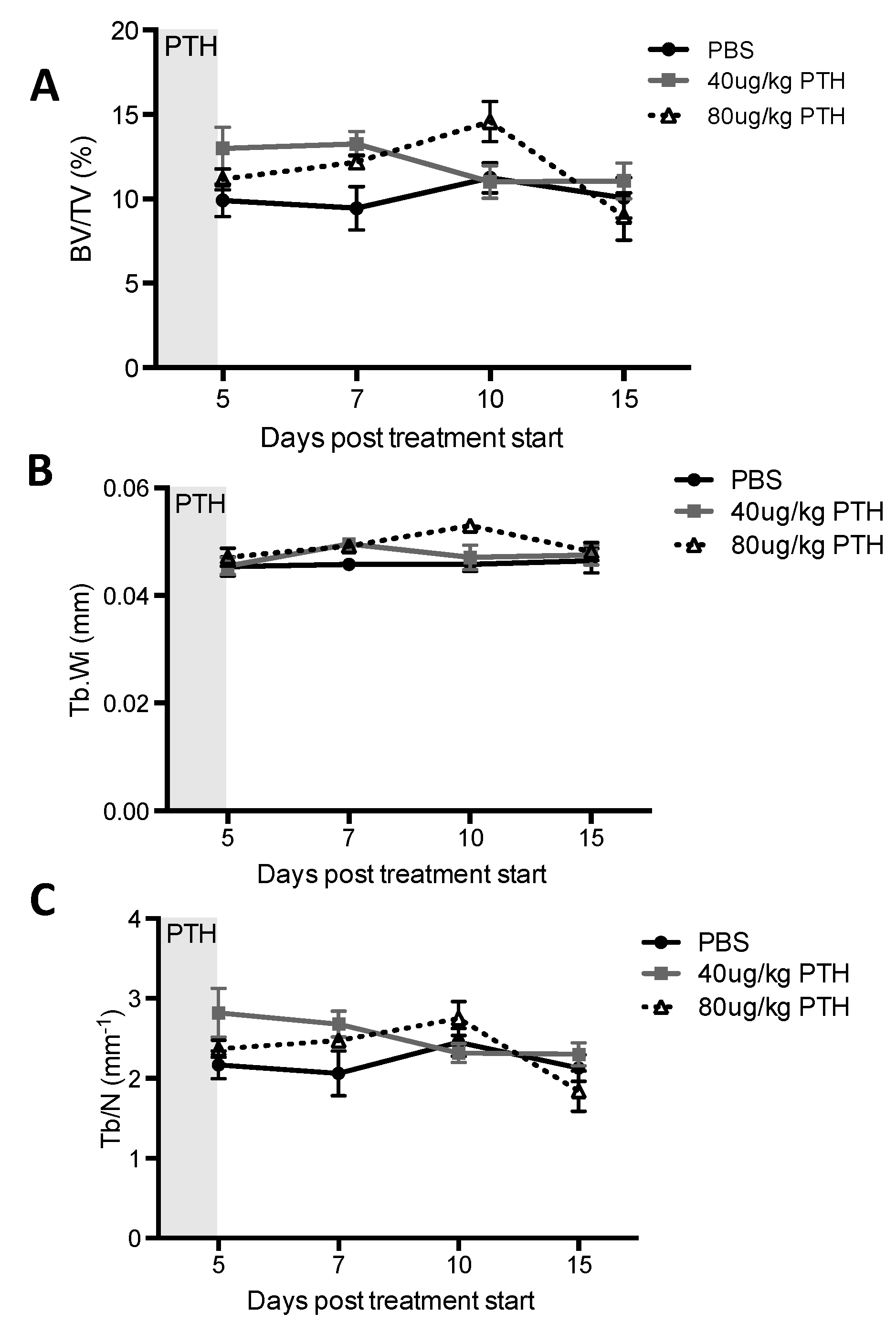
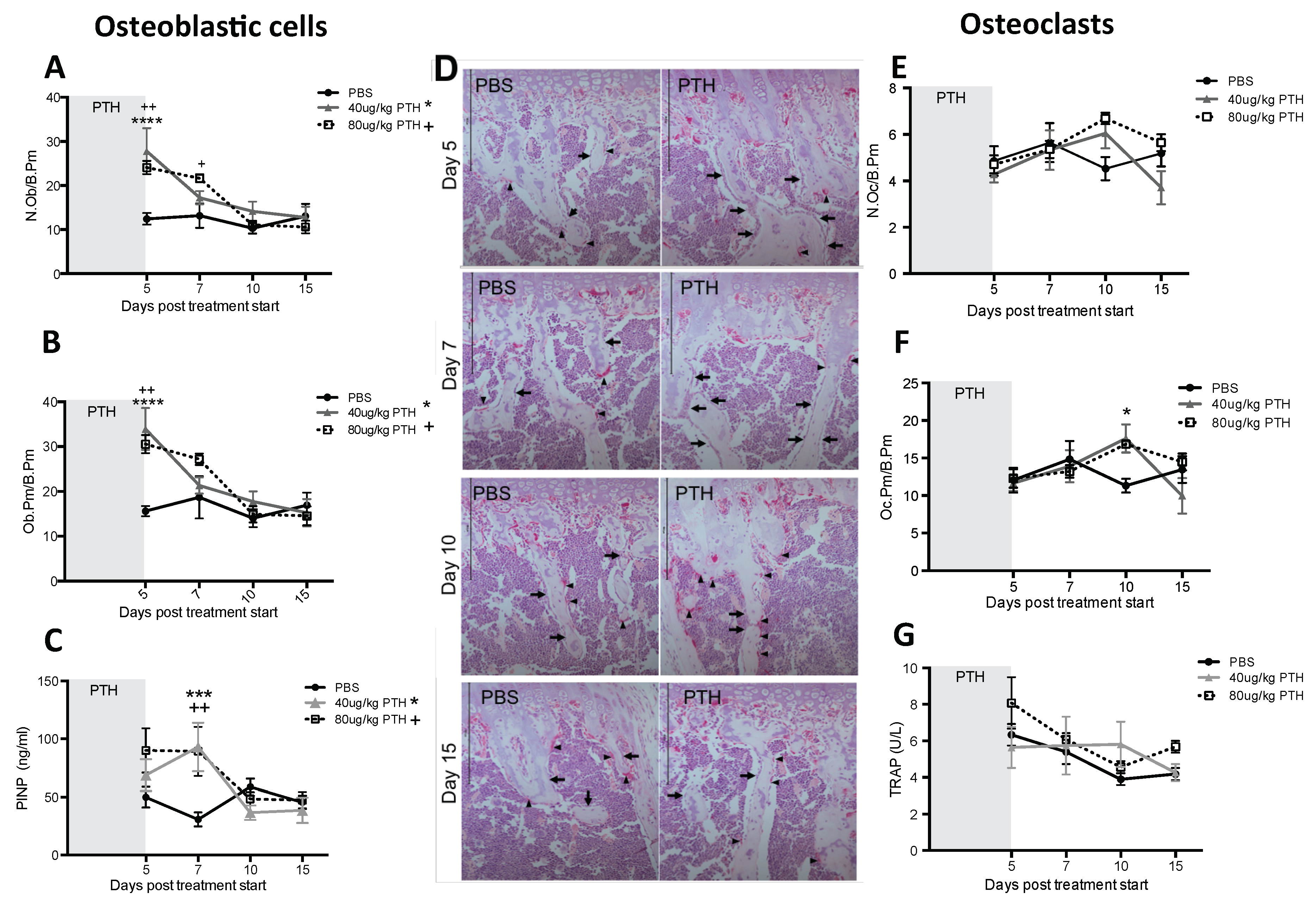
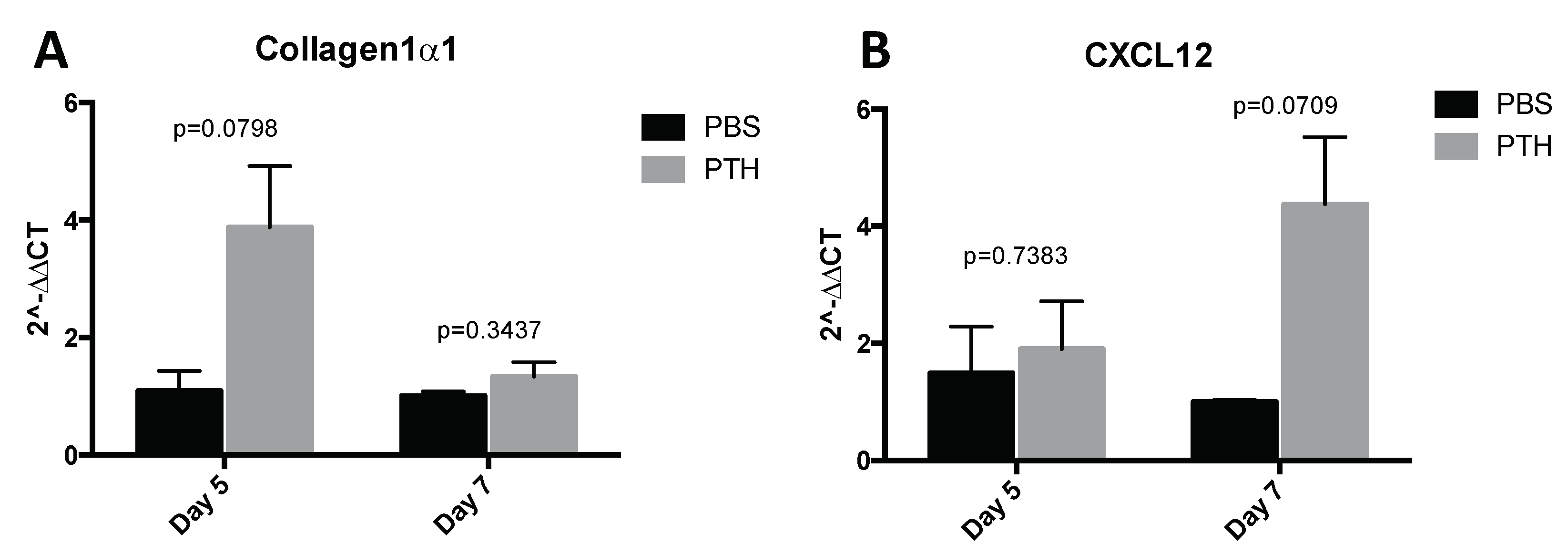
2.1. Short-Term Intermittent PTH Treatment Increased Osteoblast Numbers and Activity
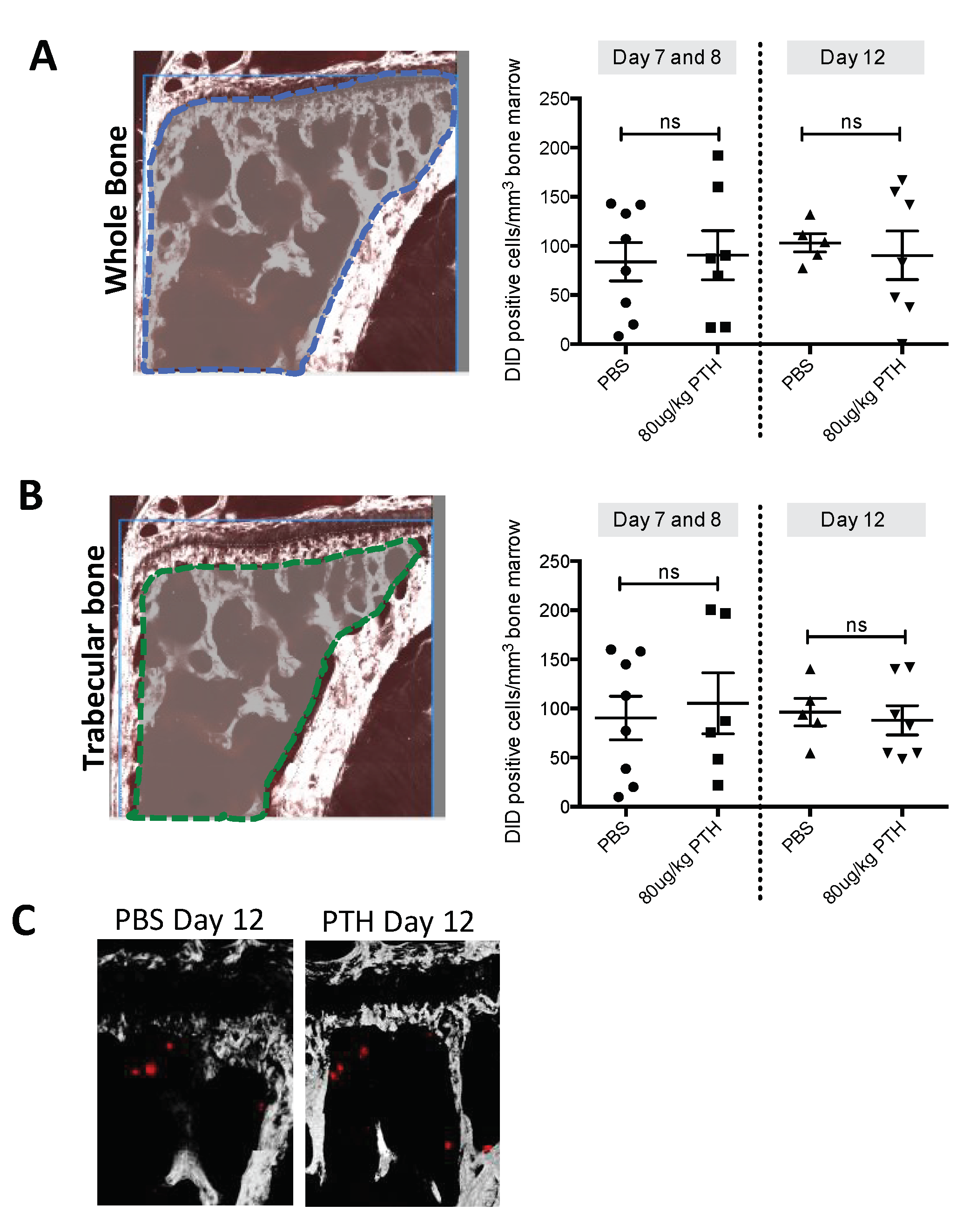
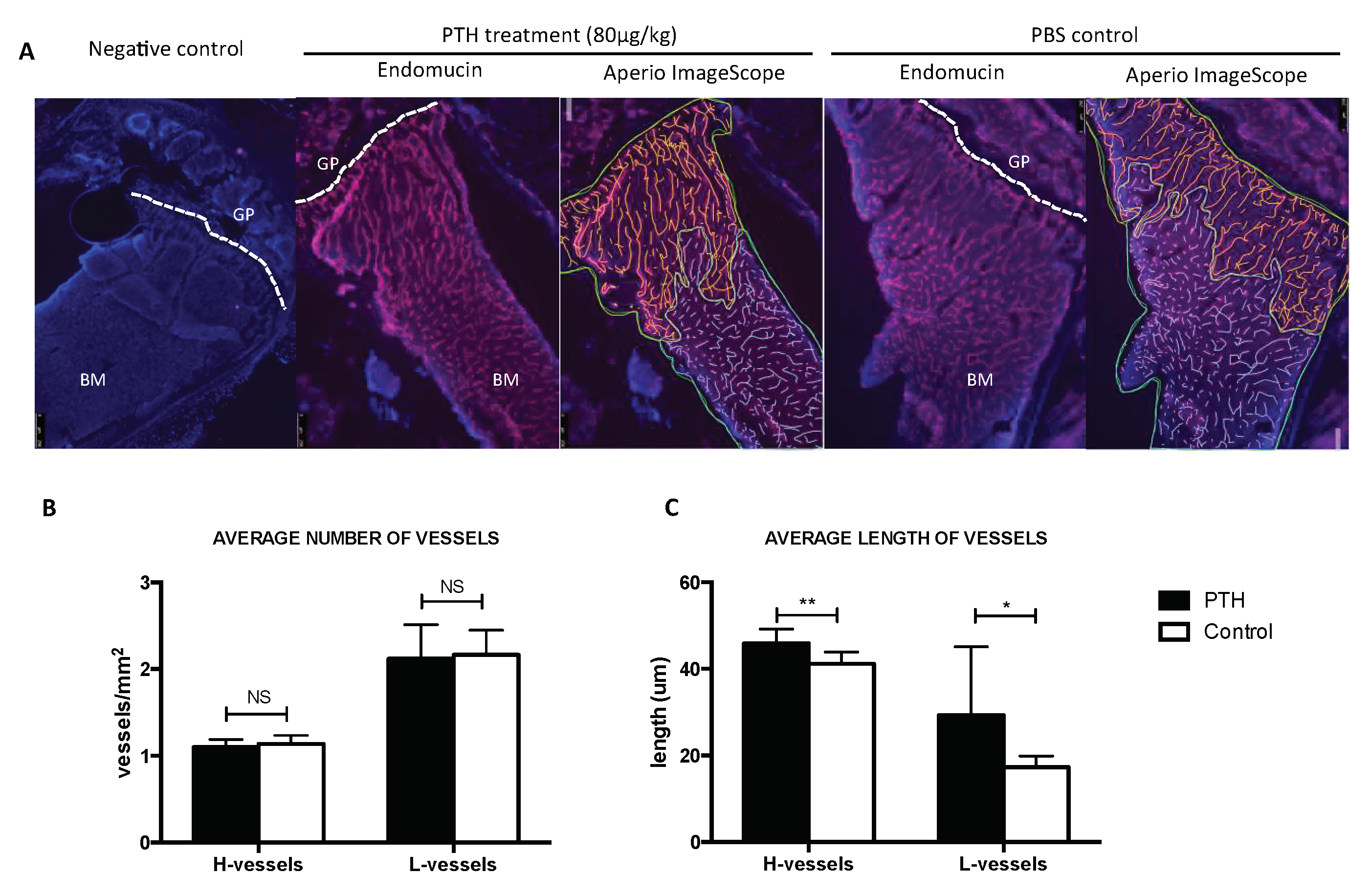
2.2. PTH Does Not Modify Tumour Cell Seeding to the Hind Limbs
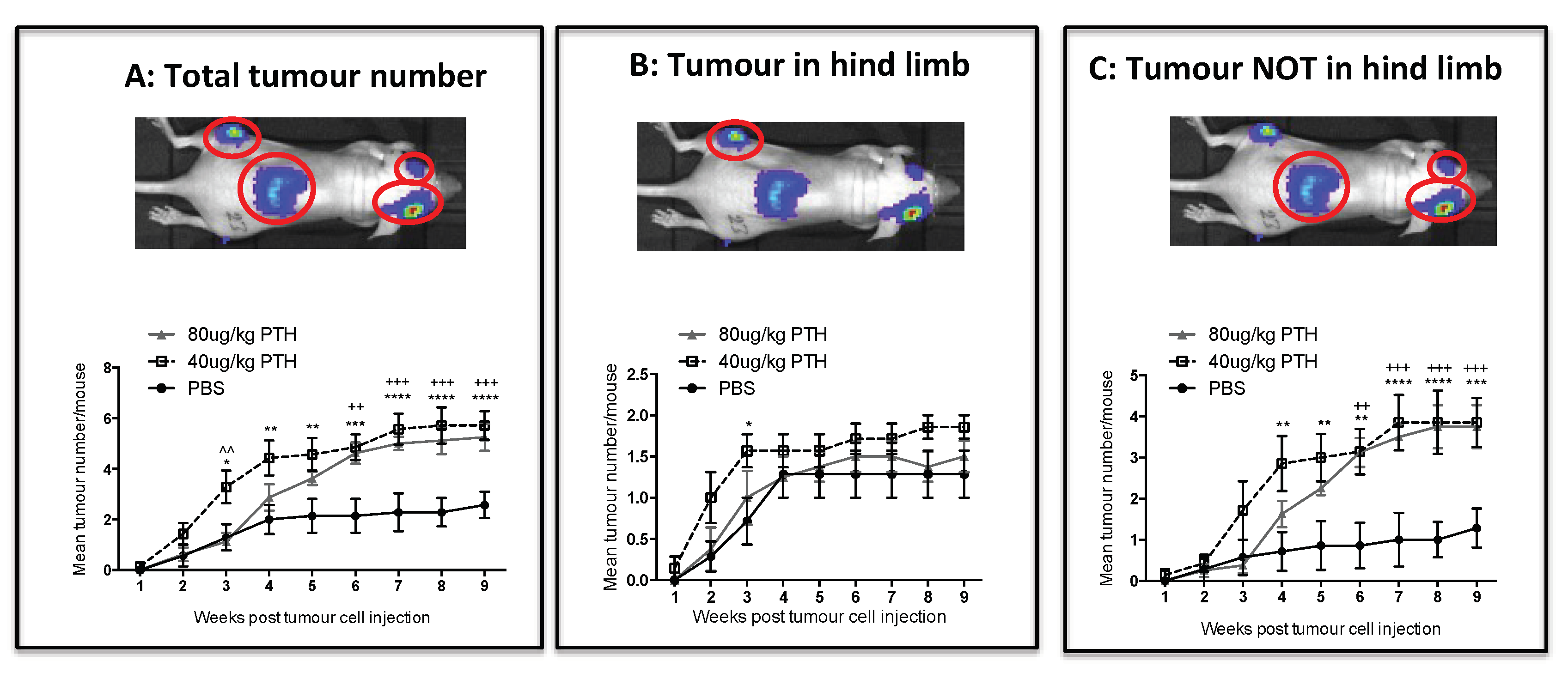
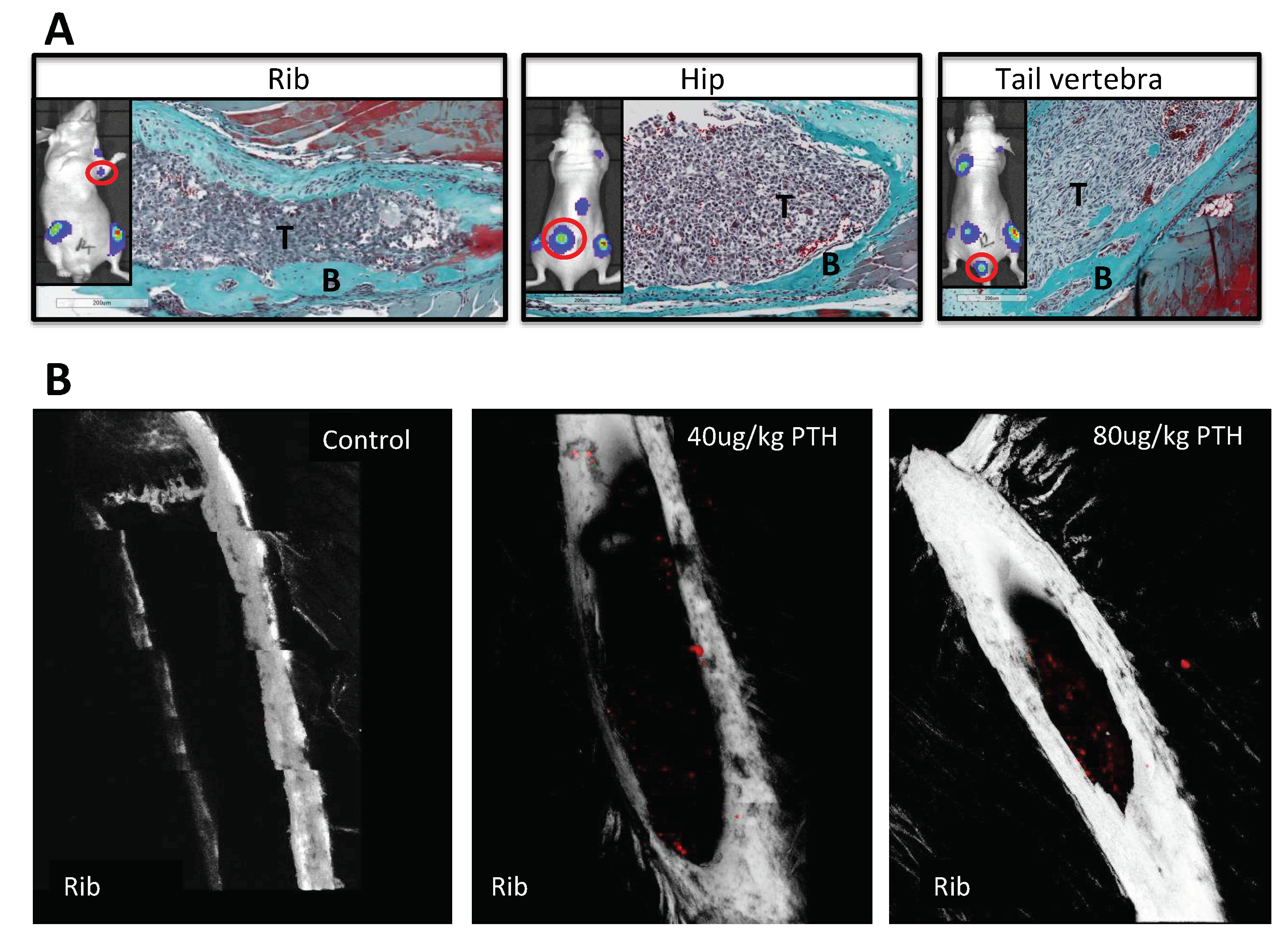
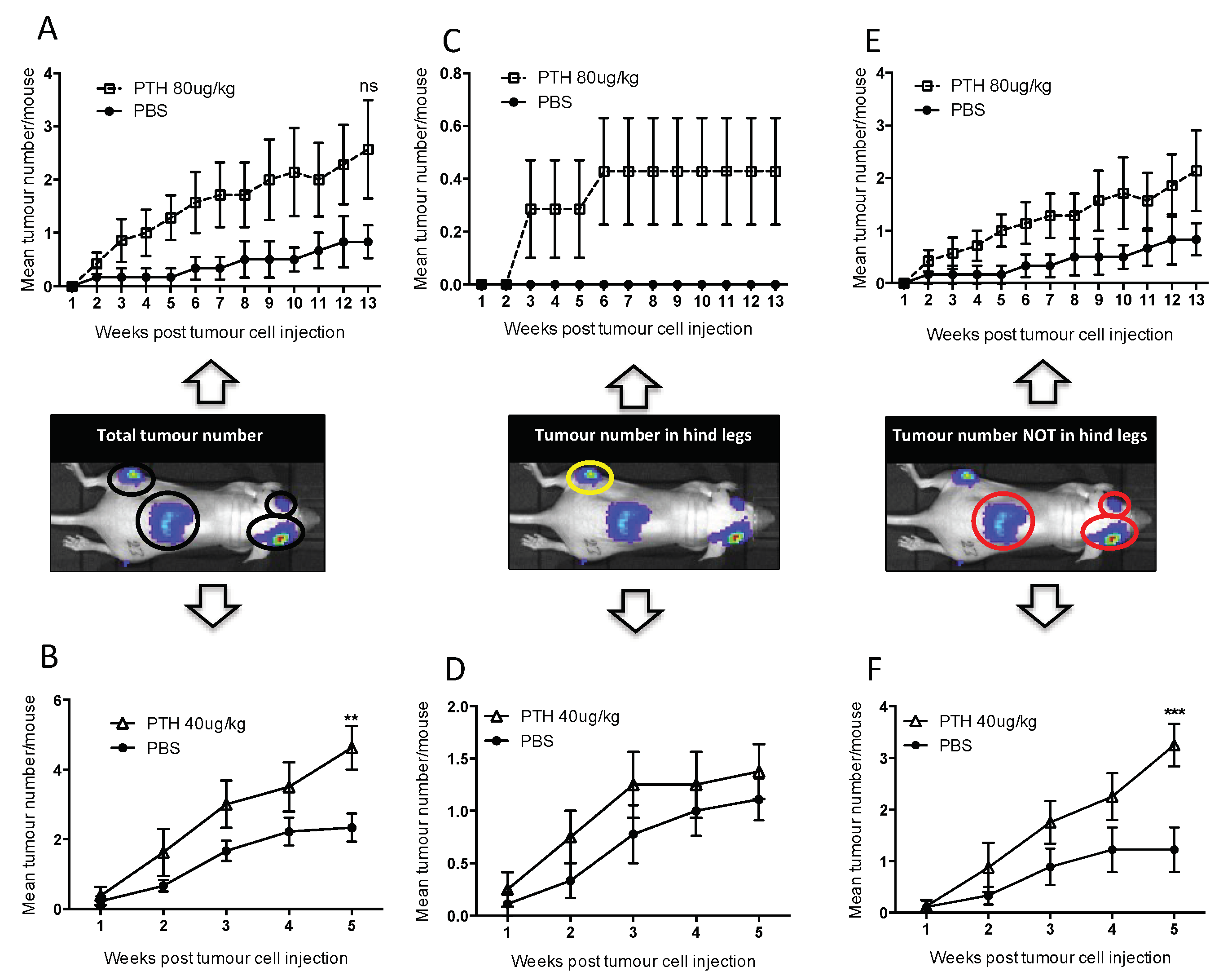
2.3. Pre-Treatment with PTH Modifies Number and Distribution of Breast Tumours in Bone
3. Discussion
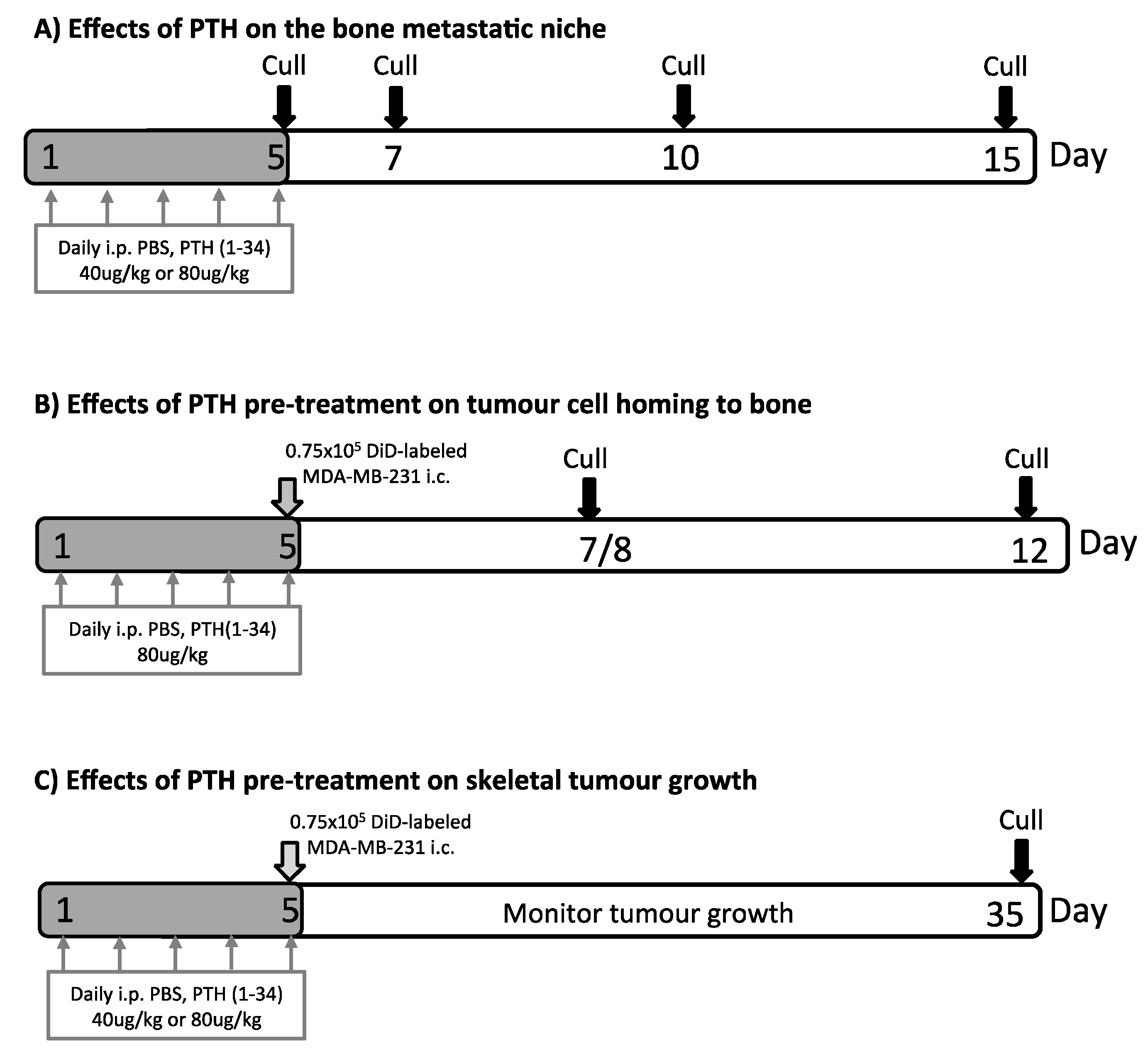
4. Materials and Methods
4.1. Cells
4.2. Animals
4.3. PTH Treatment and Intracardiac Injection of Tumour Cells
4.4. Two-Photon Microscopy
4.5. Preparation of Samples for Analyses of the Bone Microvasculature
4.6. Quantification of the Effects on the Microvasculature
4.7. Serum Bone Remodelling Markers Measured by ELISA
4.8. Quantitative Real Time PCR
4.9. Histological Quantification of Osteoclasts and Osteoblasts
4.10. Microcomputed Tomography
4.11. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DTC | Disseminated Tumour Cell |
| PTH | Parathyroid Hormone |
| Ob | Osteoblast |
| Oc | Osteoclast |
| HSC | Hematopoietic Stem Cell |
| uCT | Microcomputed Tomography |
Appendix A

Appendix B

References
- Walkington, L.; Coleman, R.E. Advances in management of bone disease in breast cancer. Bone 2011, 48, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Pantel, K.; Kang, Y. Tumor metastasis: Moving new biological insights into the clinic. Nat. Med. 2013, 19, 1450–1464. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Ghiso, J.A. Models, mechanisms and clinical evidence for cancer dormancy. Nat. Rev. Cancer 2007, 7, 834–846. [Google Scholar] [CrossRef] [PubMed]
- Ottewell, P.D.; Wang, N.; Brown, H.K.; Reeves, K.J.; Fowles, C.A.; Croucher, P.I.; Eaton, C.L.; Holen, I. Zoledronic acid has differential anti-tumour activity in the pre-and post-menopausal bone microenvironment in vivo. Clin. Cancer Res. 2014, 20, 2922–2932. [Google Scholar] [CrossRef] [PubMed]
- Ottewell, P.D.; Wang, N.; Meek, J.; Fowles, C.A.; Croucher, P.I.; Eaton, C.L.; Holen, I. Castration-induced bone loss triggers growth of disseminated prostate cancer cells in bone. Endocr. Relat. Cancer 2014, 21, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Lawson, M.A.; McDonald, M.M.; Kovacic, N.; Hua Khoo, W.; Terry, R.L.; Down, J.; Kaplan, W.; Paton-Hough, J.; Fellows, C.; Pettitt, J.A.; et al. Osteoclasts control re-activation of dormant myeloma cells by remodeling the endosteal niche. Nat. Commun. 2015, 6, 8983. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, C.; Gao, X.; Welte, T.; Muscarella, A.M.; Tian, L.; Zhao, H.; Zhao, Z.; Du, S.; Tao, J.; et al. The osteogenic niche promotes early-stage bone colonization of disseminated breast cancer cells. Cancer Cell. 2015, 27, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Ghajar, C.M.; Peinado, H.; Mori, H.; Matei, I.R.; Evason, K.J.; Brazier, H.; Almeida, D.; Koller, A.; Hajjar, K.A.; Stainier, D.Y.R.; et al. The perivascular niche regulates breast tumour dormancy. Nat. Cell. Biol. 2013, 15, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Shiozawa, Y.; Pedersen, E.A.; Havens, A.M.; Jung, Y.; Mishra, A.; Joseph, J.; Kim, J.K.; Patel, L.R.; Ying, C.; Ziegler, A.M.; et al. Human prostate cancer metastases target the hematopoietic stem cell niche to establish footholds in mouse bone marrow. J. Clin. Investig. 2011, 121, 1298–1312. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Docherty, F.E.; Brown, H.K.; Reeves, K.J.; Fowles, C.A.; Ottewell, P.D.; Dear, T.N.; Holen, I.; Croucher, P.I.; Eaton, C.L. Prostate cancer cells preferentially home to osteoblast-rich areas in the early stages of bone metastasis: Evidence from in vivo models. J. Bone Miner. Res. 2014, 29, 2688–2696. [Google Scholar] [CrossRef] [PubMed]
- Calvi, L.M.; Adams, G.B.; Weibrecht, K.W.; Weber, J.M.; Olson, D.P.; Knight, M.C.; Martin, R.P.; Schipani, E.; Divieti, P.; Bringhurst, F.R.; et al. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature 2003, 425, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Kusumbe, A.P.; Ramasamy, S.K.; Adams, R.H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 2014, 507, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Jilka, R.L. Molecular and cellular mechanisms of the anabolic effect of intermittent PTH. Bone 2007, 40, 1434–1446. [Google Scholar] [CrossRef] [PubMed]
- Jilka, R.L.; O’Brien, C.A.; Ali, A.A.; Roberson, P.K.; Weinstein, R.S.; Manolagas, S.C. Intermittent PTH stimulates periosteal bone formation by actions on post-mitotic preosteoblasts. Bone 2009, 44, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Frolik, C.A.; Black, E.C.; Cain, R.L.; Satterwhite, J.H.; Brown-Augsburger, P.L.; Sato, M.; Hock, J.M. Anabolic and catabolic bone effects of human parathyroid hormone (1-34) are predicted by duration of hormone exposure. Bone 2003, 33, 372–379. [Google Scholar] [CrossRef]
- Allen, M.R.; Hock, J.M.; Burr, D.B. Periosteum: Biology, regulation, and response to osteoporosis therapies. Bone 2004, 35, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Kutilek, S.; Zhang, C.; Shen, H.; Kodama, Y.; Srivastava, A.; Wergedal, J.; Beamer, W.; Baylink, D. Comparison of bone formation responses to parathyroid hormone (1-34), (1-31), and (2-34) in mice. Bone 2000, 27, 471–478. [Google Scholar] [CrossRef]
- Kim, S.W.; Pajevic, P.D.; Selig, M.; Barry, K.J.; Yang, J.; Shin, C.S.; Baek, W.; Kim, J.; Kronenberg, H.M. Intermittent PTH administration converts quiescent lining cells to active osteoblasts. J. Bone Miner. Res. 2012, 27, 2075–2084. [Google Scholar] [CrossRef] [PubMed]
- Ottewell, P.D.; Wang, N.; Brown, H.K.; Fowles, C.A.; Croucher, P.I.; Eaton, C.L.; Holen, I. OPG-Fc inhibits ovariectomy-induced growth of disseminated breast cancer cells in bone. Int. J. Cancer 2015, 137, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Bellido, T.; Ali, A.A.; Plotkin, L.I.; Fu, Q.; Gubrij, I.; Roberson, P.K.; Weinstein, R.S.; O’Brien, C.A.; Manolagas, S.C.; Jilka, R.L. Proteasomal degradation of Runx2 shortens parathyroid hormone-induced anti-apoptotic signaling in osteoblasts. A putative explanation for why intermittent administration is needed for bone anabolism. J. Biol. Chem. 2003, 278, 50259–50272. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, R.S.; Jilka, R.L.; Parfitt, A.M.; Manolagas, S.C. Inhibition of osteoblastogenesis and promotion of apoptosis of osteoblasts and osteocytes by glucocorticoids. Potential mechanisms of their deleterious effects on bone. J. Clin. Investig. 1998, 102, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Huber, B.C.; Grabmaier, U.; Brunner, S. Impact of parathyroid hormone on bone marrow-derived stem cell mobilization and migration. World J. Stem Cells 2014, 6, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Prisby, R.; Guignandon, A.; Vanden-Bossche, A.; Mac-Way, F.; Linossier, M.-T.; Thomas, M.; Laroche, N.; Malaval, L.; Langer, M.; Peter, Z.-A.; et al. Intermittent PTH(1-84) is osteoanabolic but not osteoangiogenic and relocates bone marrow blood vessels closer to bone-forming sites. J. Bone Miner. Res. 2011, 26, 2583–2596. [Google Scholar] [CrossRef] [PubMed]
- Adams, G.B.; Martin, R.P.; Alley, I.R.; Chabner, K.T.; Cohen, K.S.; Calvi, L.M.; Kronenberg, H.M.; Scadden, D.T. Therapeutic targeting of a stem cell niche. Nat. Biotechnol. 2007, 25, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Ballen, K.K.; Shpall, E.J.; Avigan, D.; Yeap, B.Y.; Fisher, D.C.; McDermott, K.; Dey, B.R.; Attar, E.; McAfee, S.; Konopleva, M.; et al. Phase I trial of parathyroid hormone to facilitate stem cell mobilization. Biol. Blood Marrow Transpl. 2007, 13, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Xian, L.; Crane, J.; Wen, C.; Hilton, M.; Lu, W.; Newman, P.; Cao, X. PTH Receptor Signaling in Osteoblasts Regulates Endochondral Vascularization in Maintenance of Postnatal Growth Plate. J. Bone Miner. Res. 2015, 30, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Havens, A.M.; Pedersen, E.A.; Shiozawa, Y.; Ying, C.; Jung, Y.; Sun, Y.; Neeley, C.; Wang, J.; Mehra, R.; Keller, E.T.; et al. An in vivo mouse model for human prostate cancer metastasis. Neoplasia 2008, 10, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.; Kalikin, L.M.; Mattos, A.C.; Keller, E.T.; Allen, M.J.; Pienta, K.J.; McCauley, L.K. Bone turnover mediates preferential localization of prostate cancer in the skeleton. Endocrinology 2005, 146, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- Swami, S.; Johnson, J.; Bettinson, L.A.; Kimura, T.; Zhu, H.; Albertelli, M.A. Prevention of breast cancer skeletal metastases with parathyroid hormone. JCI Insight 2017, 2, e90874. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Iida-Klein, A.; Lu, S.; Ducayen-Knowles, M.; Levine, L.; Dempster, D.; Lindsay, R. Anabolic action of parathyroid hormone on cortical and cancellous bone differs between axial and appendicular skeletal sites in mice. Bone 2003, 32, 513–520. [Google Scholar] [CrossRef]
- Iida-Klein, A.; Zhou, H.; Lu, S.S.; Levine, L.R.; Ducayen-Knowles, M.; Dempster, D.W.; Nieves, J.; Lindsay, R. Anabolic action of parathyroid hormone is skeletal site specific at the tissue and cellular levels in mice. J. Bone Miner. Res. 2002, 17, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Compston, J.E. Skeletal actions of intermittent parathyroid hormone: Effects on bone remodelling and structure. Bone 2007, 40, 1447–1452. [Google Scholar] [CrossRef] [PubMed]
- Vahle, J.L.; Sato, M.; Long, G.G.; Young, J.K.; Francis, P.C.; Engelhardt, J.A.; Westmore, M.S.; Ma, Y.L.; Nold, J.B. Skeletal changes in rats given daily subcutaneous injections of recombinant human parathyroid hormone (1-34) for 2 years and relevance to human safety. Toxicol. Pathol. 2002, 30, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Subbiah, V.; Madsen, V.S.; Raymond, A.K.; Benjamin, R.S.; Ludwig, J.A. Of mice and men: Divergent risks of teriparatide-induced osteosarcoma. Osteoporos. Int. 2010, 21, 1041–1045. [Google Scholar] [CrossRef] [PubMed]
- Suvannasankha, A.; Chirgwin, J.M. Role of bone-anabolic agents in the treatment of breast cancer bone metastases. Breast Cancer Res. 2014, 16, 484. [Google Scholar] [CrossRef] [PubMed]
- Weilbaecher, K.N.; Guise, T.A.; McCaukey, L.K. Cancer to bone: A fatal attraction. Nat. Rev. Cancer 2011, 11, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.K.; Ottewell, P.D.; Evans, C.A.; Holen, I. Location matters—Osteoblast and osteoclast distribution is modified by the presence and proximity to breast cancer cells in vivo. Clin. Exp. Metastasis 2012, 8, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, A.M. Standardized nomenclature, symbols, and units for bone histomorphometry: A 2012 update of the Report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Miner. Res. 2013, 28, 2–17. [Google Scholar] [CrossRef] [PubMed]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Müller, R. Guidelines for assessment of bone microstructure in rodents using micro–computed tomography. J. Bone Miner. Res. 2010, 25, 1468–1486. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Mouse | Hind Limbs | Adrenal Gland | Front Limbs | Ribs | Other Sites | Total no of Tumours | Tumours/Mouse |
|---|---|---|---|---|---|---|---|---|
| Control | 1 | Both | Left | -- | -- | -- | 3 | |
| 2 | Right | Right | -- | -- | -- | 2 | ||
| 3 | Right | -- | Both | -- | -- | 3 | ||
| 5 | Both | Right | Both | -- | -- | 5 | ||
| 7 | -- | -- | -- | -- | Head | 2 | ||
| 8 | Right | -- | -- | -- | -- | 1 | ||
| N = 6 | 5/6 | 3/6 | 2/6 | 0/6 | 1/6 | 16 | 2.66 | |
| 40 µg/kg PTH | 9 | Both | Right | Left | -- | Head | 5 | |
| 10 | Both | Both | Left | Right | -- | 6 | ||
| 12 | Both | Both | Left | -- | Tail, spine | 7 | ||
| 13 | Right | Both | Left | Left | Jaw | 6 | ||
| 14 | Both | -- | Left | Right | -- | 4 | ||
| 15 | Both | Right | ? | ? | Head | 4 | ||
| 16 | Both | Right | -- | Right | Head | 5 | ||
| N = 7 | 7/7 | 6/7 | 5/7 | 4/7 | 5/7 | 37 | 5.28 | |
| 80 µg/kg PTH | 17 | Both | -- | -- | Left | -- | 3 | |
| 18 | Left | Left | -- | Right | -- | 3 | ||
| 19 | Both | Both | Left | -- | Hip /pelvis | 6 | ||
| 20 | Left | Right | -- | -- | -- | 2 | ||
| 21 | Left | Left | -- | -- | -- | 2 | ||
| 22 | Left | Both | -- | Left | -- | 4 | ||
| 23 | Left | Left | -- | -- | Head | >3 | ||
| 24 | Both | Both | -- | Left | -- | 5 | ||
| N = 8 | 8/8 | 7/8 | 1/8 | 4/8 | 2/8 | 28 | 3.50 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brown, H.K.; Allocca, G.; Ottewell, P.D.; Wang, N.; Brown, N.J.; Croucher, P.I.; Eaton, C.L.; Holen, I. Parathyroid Hormone (PTH) Increases Skeletal Tumour Growth and Alters Tumour Distribution in an In Vivo Model of Breast Cancer. Int. J. Mol. Sci. 2018, 19, 2920. https://doi.org/10.3390/ijms19102920
Brown HK, Allocca G, Ottewell PD, Wang N, Brown NJ, Croucher PI, Eaton CL, Holen I. Parathyroid Hormone (PTH) Increases Skeletal Tumour Growth and Alters Tumour Distribution in an In Vivo Model of Breast Cancer. International Journal of Molecular Sciences. 2018; 19(10):2920. https://doi.org/10.3390/ijms19102920
Chicago/Turabian StyleBrown, Hannah K., Gloria Allocca, Penelope D. Ottewell, Ning Wang, Nicola J. Brown, Peter I. Croucher, Colby L. Eaton, and Ingunn Holen. 2018. "Parathyroid Hormone (PTH) Increases Skeletal Tumour Growth and Alters Tumour Distribution in an In Vivo Model of Breast Cancer" International Journal of Molecular Sciences 19, no. 10: 2920. https://doi.org/10.3390/ijms19102920
APA StyleBrown, H. K., Allocca, G., Ottewell, P. D., Wang, N., Brown, N. J., Croucher, P. I., Eaton, C. L., & Holen, I. (2018). Parathyroid Hormone (PTH) Increases Skeletal Tumour Growth and Alters Tumour Distribution in an In Vivo Model of Breast Cancer. International Journal of Molecular Sciences, 19(10), 2920. https://doi.org/10.3390/ijms19102920