Identification of Endogenous Control miRNAs for RT-qPCR in T-Cell Acute Lymphoblastic Leukemia

 , , , , ,
, , , , ,  and
and
Abstract

1. Introduction
1.1. Background
1.2. Challenges of Normalization in RT-qPCR-Based miRNA Profiling
1.3. miRNA Expression Profiling in T-Cell Acute Lymphoblastic Leukemia
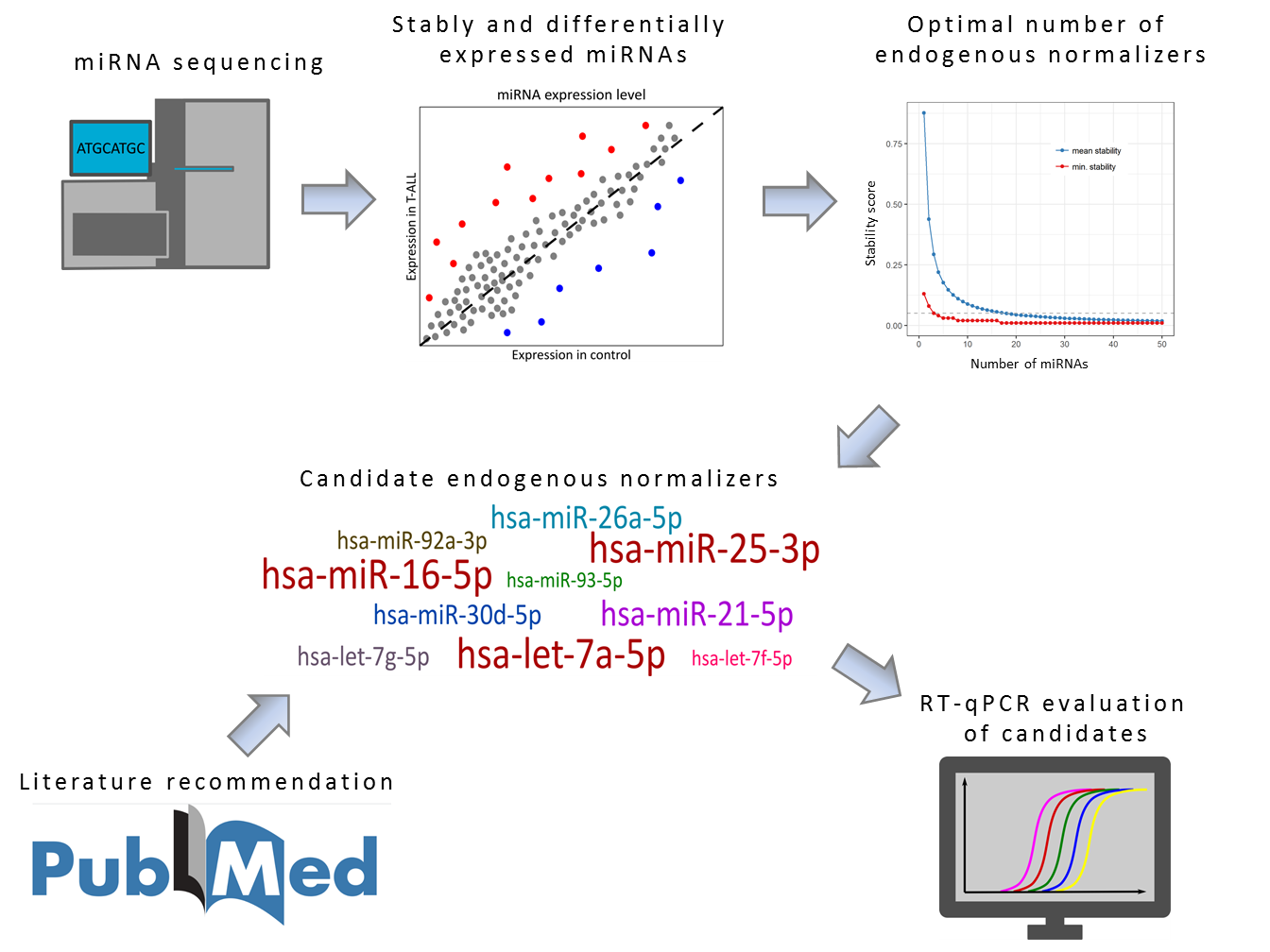
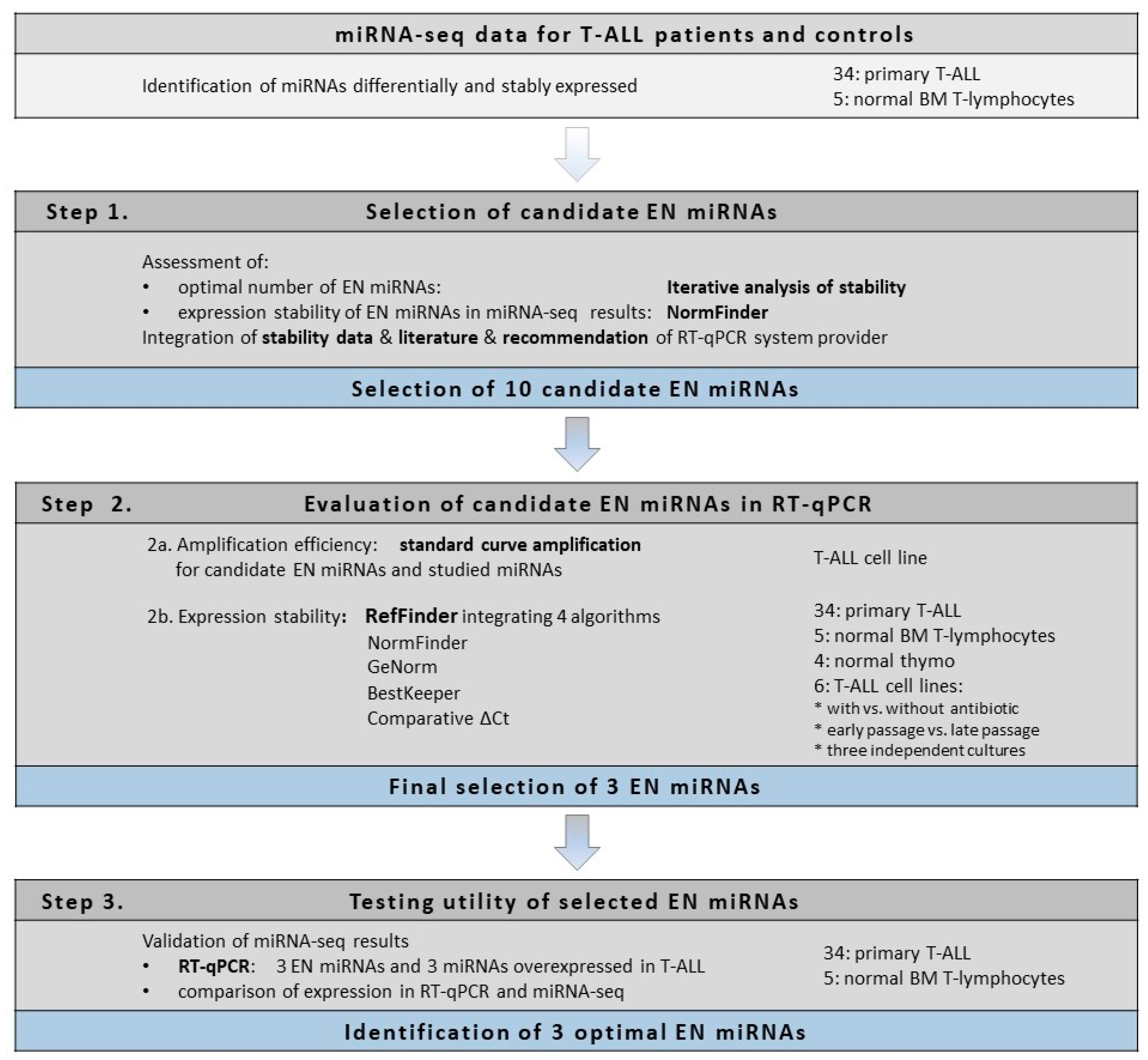
2. Results
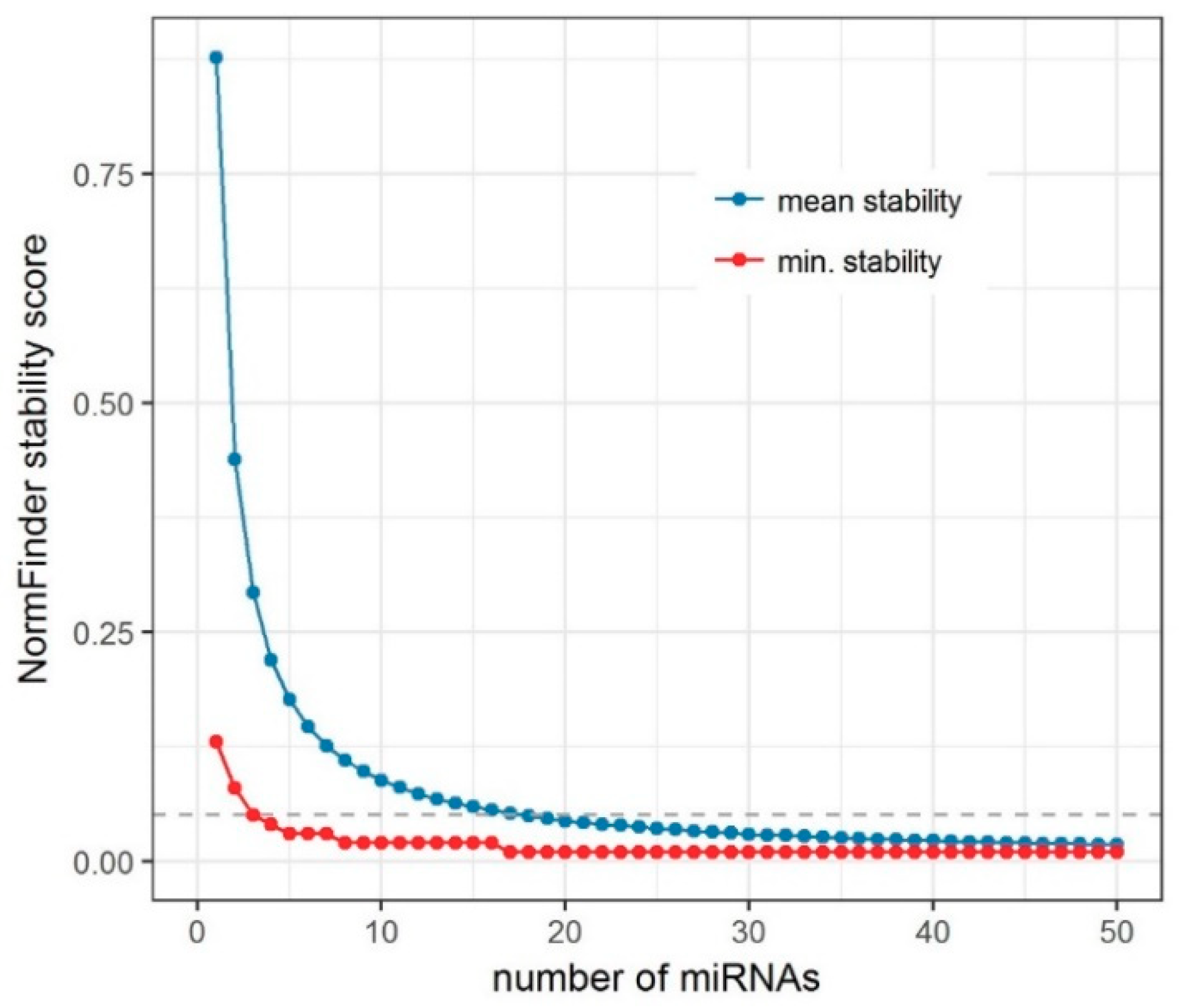
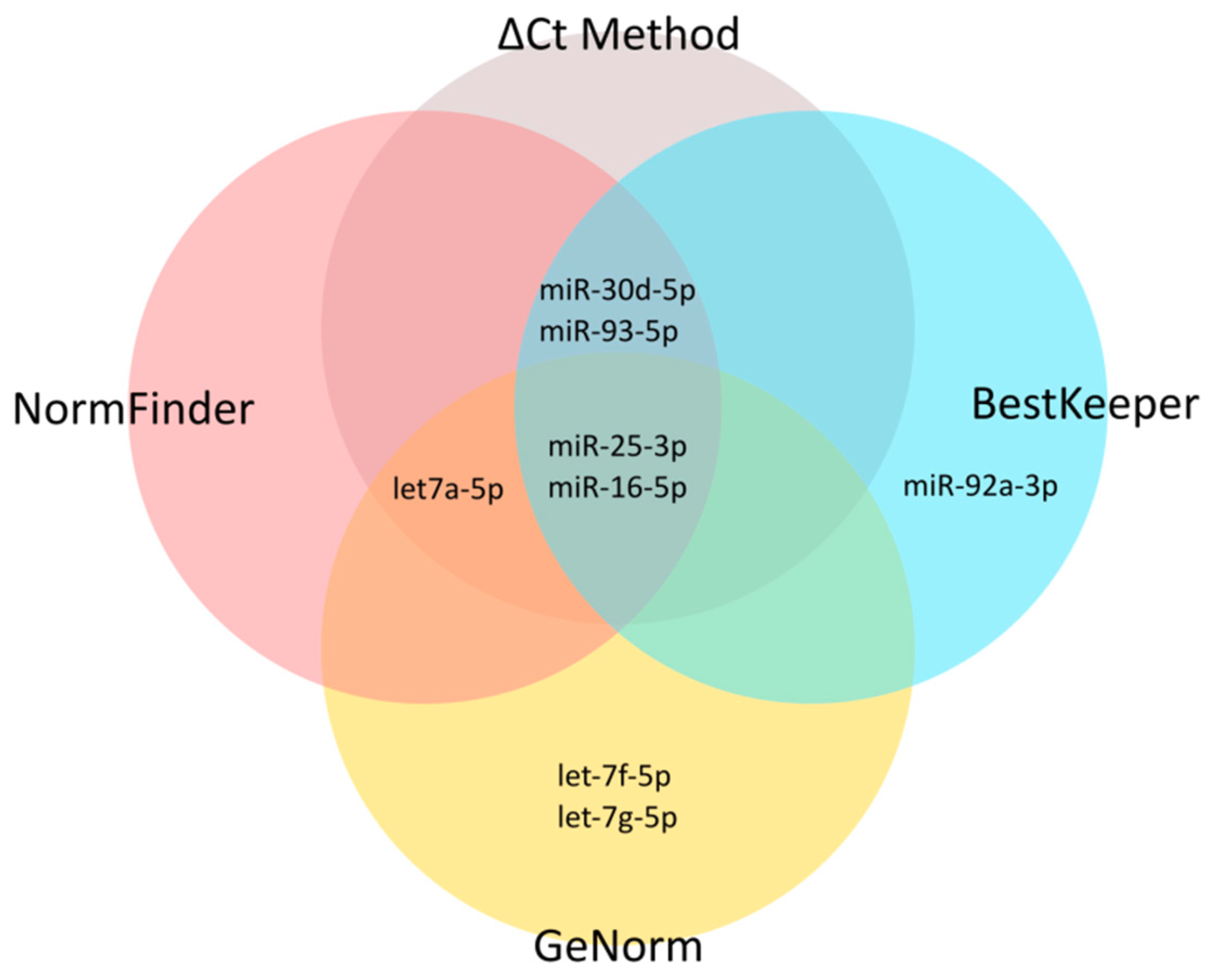
2.1. Selection of Candidate Endogenous Normalizer miRNAs (Step 1)
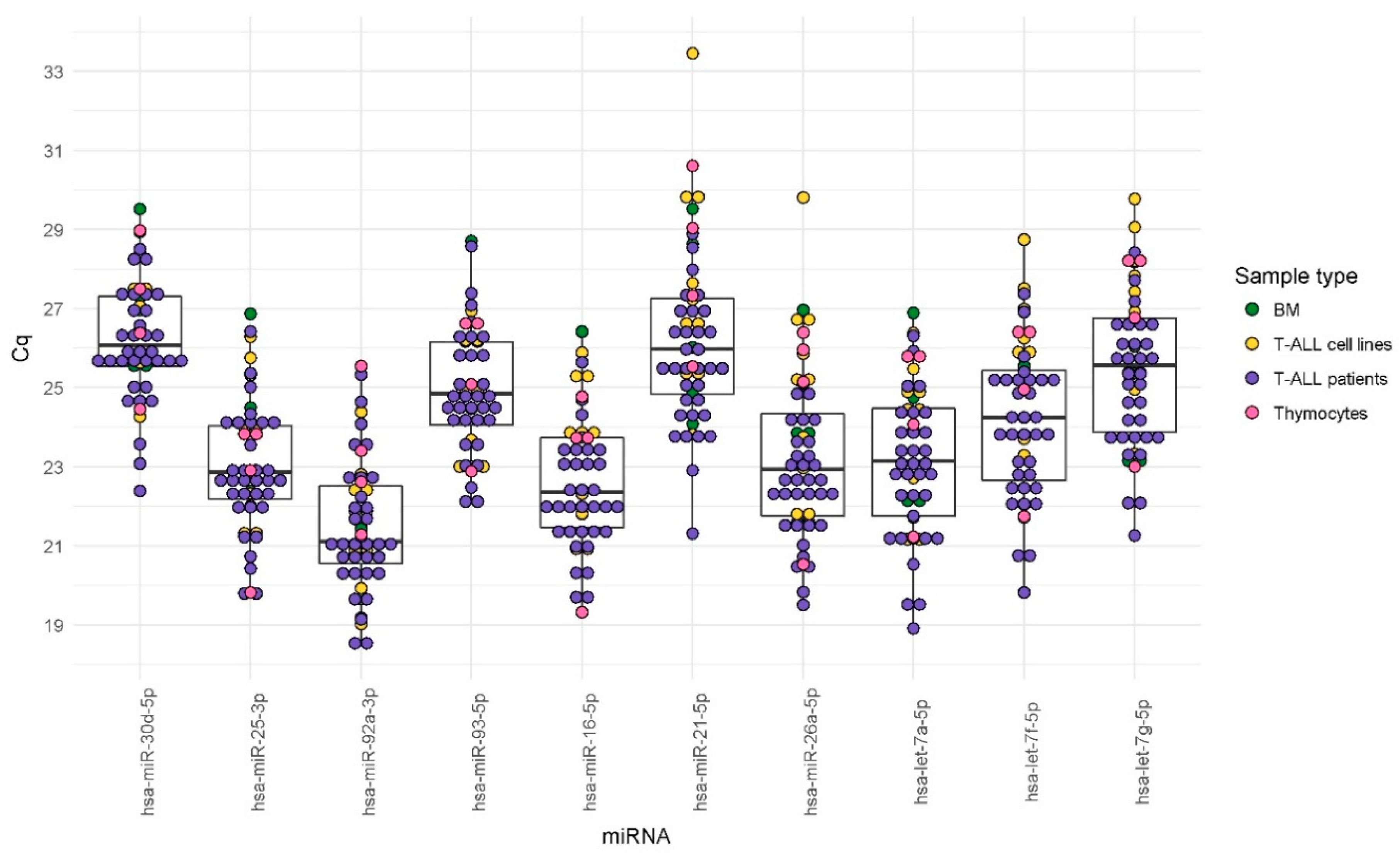
2.2. Evaluation of Candidate Endogenous Normalizer miRNAs in RT-qPCR (Step 2)
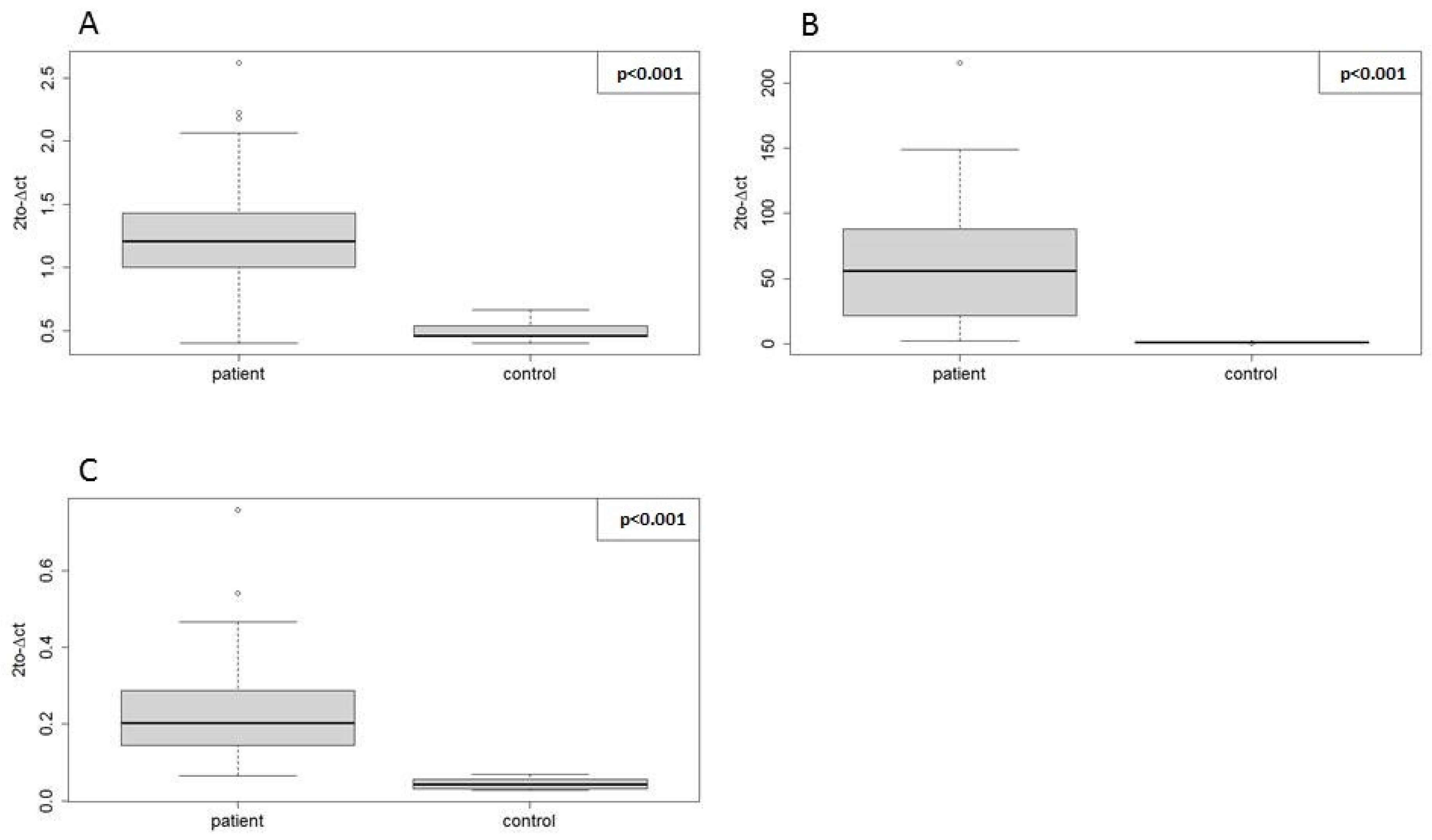
2.3. Testing the Utility of Selected Candidate Endogenous Normalizer miRNAs (Step 3)
3. Discussion
3.1. Strategy for the Identification of Optimal Endogenous Normalizer miRNAs for RT-qPCR
3.2. Number of miRNAs to Be Used as Endogenous Normalizers in RT-qPCR
3.3. Expression Stability Relative to Sample Type and Culture Conditions
3.4. RT-qPCR Validation of miRNA-Seq Results
4. Materials and Methods
4.1. Materials
4.2. miRNA-Seq
4.3. Reverse-Transcription and RT-qPCR Amplification Conditions
4.4. Amplification Efficiency
4.5. Analysis of Gene Expression
4.6. Analysis of Expression Stability
4.7. Analysis of the Similarity of Mature miRNA Sequences
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BM | bone marrow |
| Cq | quantification cycle ( nomenclature in accordance with Minimum Information for Publication of Quantitative Real-Time PCR Experiments, MIQE guidelines); Cq is equivalent to CT (threshold cycle) which is only used, when referred to ΔΔ CT method implemented in Data Assist Software version 3.01 (Thermo Fisher Scientific) or Comparative Delta CT method; these methods has retained their original names |
| EN | endogenous normalizer |
| IQR | interquartile range |
| isomiR | miRNA isoform |
| miRNA-seq | next-generation sequencing of miRNA-transcriptome |
| Q1 | first quartile |
| Q3 | third quartile |
| SD | standard deviation |
| snoRNA | small nucleolar RNA |
| snRNA | small nuclear RNA |
| T-ALL | T-cell acute lymphoblastic leukemia |
| thymo | thymocytes |
References
- MacFarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs-microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Ogata-Kawata, H.; Izumiya, M.; Kurioka, D.; Honma, Y.; Yamada, Y.; Furuta, K.; Gunji, T.; Ohta, H.; Okamoto, H.; Sonoda, H.; et al. Circulating Exosomal microRNAs as Biomarkers of Colon Cancer. PLoS ONE 2014, 9, e92921. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yan, F.; Zhao, Q.; Zhan, F.; Wang, R.; Wang, L.; Zhang, Y.; Huang, X. Circulating exosomal miR-125a-3p as a novel biomarker for early-stage colon cancer. Sci. Rep. 2017, 7, 4150. [Google Scholar] [CrossRef] [PubMed]
- Bhome, R.; Del Vecchio, F.; Lee, G.-H.; Bullock, M.D.; Primrose, J.N.; Sayan, A.E.; Mirnezami, A.H. Exosomal microRNAs (exomiRs): Small molecules with a big role in cancer. Cancer Lett. 2018, 420, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Hornick, N.I.; Huan, J.; Doron, B.; Goloviznina, N.A.; Lapidus, J.; Chang, B.H.; Kurre, P. Serum Exosome MicroRNA as a Minimally-Invasive Early Biomarker of AML. Sci. Rep. 2015, 5, 11295. [Google Scholar] [CrossRef] [PubMed]
- Elhamamsy, A.R.; El Sharkawy, M.S.; Zanaty, A.F.; Mahrous, M.A.; Mohamed, A.E.; Abushaaban, E.A. Circulating miR-92a, miR-143 and miR-342 in Plasma are Novel Potential Biomarkers for Acute Myeloid Leukemia. Int. J. Mol. Cell. Med. 2017, 6, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Manier, S.; Liu, C.-J.; Avet-Loiseau, H.; Park, J.; Shi, J.; Campigotto, F.; Salem, K.Z.; Huynh, D.; Glavey, S.V.; Rivotto, B.; et al. Prognostic role of circulating exosomal miRNAs in multiple myeloma. Blood 2017, 129, 2429–2436. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Marabita, F.; de Candia, P.; Torri, A.; Tegnér, J.; Abrignani, S.; Rossi, R.L. Normalization of circulating microRNA expression data obtained by quantitative real-time RT-PCR. Brief. Bioinform. 2016, 17, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Mavrakis, K.J.; Wolfe, A.L.; Oricchio, E.; Palomero, T.; de Keersmaecker, K.; McJunkin, K.; Zuber, J.; James, T.; Chang, K.; Khan, A.A.; et al. Genome-wide RNA-mediated interference screen identifies miR-19 targets in Notch-induced T-cell acute lymphoblastic leukaemia. Nat. Cell Biol. 2010, 12, 372. [Google Scholar] [CrossRef] [PubMed]
- Kroh, E.M.; Parkin, R.K.; Mitchell, P.S.; Tewari, M. Analysis of circulating microRNA biomarkers in plasma and serum using quantitative reverse transcription-PCR (qRT-PCR). Methods 2010, 50, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Farr, R.J.; Januszewski, A.S.; Joglekar, M.V.; Liang, H.; McAulley, A.K.; Hewitt, A.W.; Thomas, H.E.; Loudovaris, T.; Kay, T.W.H.; Jenkins, A.; et al. A comparative analysis of high-throughput platforms for validation of a circulating microRNA signature in diabetic retinopathy. Sci. Rep. 2015, 5, 10375. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 12. [Google Scholar] [CrossRef]
- Chugh, P.; Dittmer, D.P. Potential pitfalls in microRNA profiling. Wiley Interdiscip. Rev. RNA 2012, 3, 601–616. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; da Silva, A.M.; Calin, G.; Pantel, K. Data Normalization Strategies for MicroRNA Quantification. Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Gee, H.E.; Buffa, F.M.; Camps, C.; Ramachandran, A.; Leek, R.; Taylor, M.; Patil, M.; Sheldon, H.; Betts, G.; Homer, J.; et al. The small-nucleolar RNAs commonly used for microRNA normalisation correlate with tumour pathology and prognosis. Br. J. Cancer 2011, 104, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Lamba, V.; Ghodke-Puranik, Y.; Guan, W.; Lamba, J.K. Identification of suitable reference genes for hepatic microRNA quantitation. BMC Res. Notes 2014, 7, 129. [Google Scholar] [CrossRef] [PubMed]
- Serafin, A.; Foco, L.; Blankenburg, H.; Picard, A.; Zanigni, S.; Zanon, A.; Pramstaller, P.P.; Hicks, A.A.; Schwienbacher, C. Identification of a set of endogenous reference genes for miRNA expression studies in Parkinson’s disease blood samples. BMC Res. Notes 2014, 7, 715. [Google Scholar] [CrossRef] [PubMed]
- Correia, N.C.; Melao, A.; Povoa, V.; Sarmento, L.; de Cedron, M.G.; Malumbres, M.; Enguita, F.J.; Barata, J.T. microRNAs regulate TAL1 expression in T-cell acute lymphoblastic leukemia. Oncotarget 2016, 7, 8268–8281. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.; Bhatnagar, H.; Lin, A.P.; Wang, L.; Aster, J.C.; Sill, H.; Aguiar, R.C.T. A microRNA-mediated regulatory loop modulates NOTCH and MYC oncogenic signals in B- and T-cell malignancies. Leukemia 2015, 29, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.S.; Ono, M. From snoRNA to miRNA: Dual function regulatory non-coding RNAs. Biochimie 2011, 93, 1987–1992. [Google Scholar] [CrossRef] [PubMed]
- Delpu, Y.; Larrieu, D.; Gayral, M.; Arvanitis, D.; Dufresne, M.; Cordelier, P.; Torrisani, J. Noncoding RNAs: Clinical and Therapeutic Applications. In Drug Discovery in Cancer Epigenetics; Academic Press: Boston, MA, USA, 2016; Chapter 12; pp. 305–326. ISBN 978-0-12-802208-5. [Google Scholar]
- Belver, L.; Ferrando, A. The genetics and mechanisms of T cell acute lymphoblastic leukaemia. Nat. Rev. Cancer 2016, 16, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Van Vlierberghe, P.; Pieters, R.; Beverloo, H.B.; Meijerink, J.P.P. Molecular-genetic insights in paediatric T-cell acute lymphoblastic leukaemia. Br. J. Haematol. 2008, 143, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Wallaert, A.; Durinck, K.; Taghon, T.; Van Vlierberghe, P.; Speleman, F. T-ALL and thymocytes: A message of noncoding RNAs. J. Hematol. Oncol. 2017, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Drobna, M.; Szarzyńska-Zawadzka, B.; Dawidowska, M. T-cell acute lymphoblastic leukemia from miRNA perspective: Basic concepts, experimental approaches, and potential biomarkers. Blood Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Mets, E.; Van der Meulen, J.; Van Peer, G.; Boice, M.; Mestdagh, P.; Van de Walle, I.; Lammens, T.; Goossens, S.; De Moerloose, B.; Benoit, Y.; et al. MicroRNA-193b-3p acts as a tumor suppressor by targeting the MYB oncogene in T-cell acute lymphoblastic leukemia. Leukemia 2015, 29, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Correia, N.C.; Durinck, K.; Leite, A.P.; Ongenaert, M.; Rondou, P.; Speleman, F.; Enguita, F.J.; Barata, J.T. Novel TAL1 targets beyond protein-coding genes: Identification of TAL1-regulated microRNAs in T-cell acute lymphoblastic leukemia. Leukemia 2013, 27, 1603–1606. [Google Scholar] [CrossRef] [PubMed]
- Wallaert, A.; Van Loocke, W.; Hernandez, L.; Taghon, T.; Speleman, F.; Van Vlierberghe, P. Comprehensive miRNA expression profiling in human T-cell acute lymphoblastic leukemia by small RNA-sequencing. Sci. Rep. 2017, 7, 7901. [Google Scholar] [CrossRef] [PubMed]
- Schotte, D.; Moqadam, F.A.; Lange-Turenhout, E.A.M.; Chen, C.; van Ijcken, W.F.J.; Pieters, R.; den Boer, M.L. Discovery of new microRNAs by small RNAome deep sequencing in childhood acute lymphoblastic leukemia. Leukemia 2011, 25, 1389–1399. [Google Scholar] [CrossRef] [PubMed]
- Mavrakis, K.J.; Van der Meulen, J.; Wolfe, A.L.; Liu, X.P.; Mets, E.; Taghon, T.; Khan, A.A.; Setti, M.; Rondou, P.; Vandenberghe, P.; et al. A cooperative microRNA-tumor suppressor gene network in acute T-cell lymphoblastic leukemia (T-ALL). Nat. Genet. 2011, 43, 673. [Google Scholar] [CrossRef] [PubMed]
- Mets, E.; Van Peer, G.; Van der Meulen, J.; Boice, M.; Taghon, T.; Goossens, S.; Mestdagh, P.; Benoit, Y.; De Moerloose, B.; Van Roy, N.; et al. MicroRNA-128-3p is a novel oncomiR targeting PHF6 in T-cell acute lymphoblastic leukemia. Haematologica 2014, 99, 1326–1333. [Google Scholar] [CrossRef] [PubMed]
- Sanghvi, V.R.; Mavrakis, K.J.; Van der Meulen, J.; Boice, M.; Wolfe, A.L.; Carty, M.; Mohan, P.; Rondou, P.; Socci, N.D.; Benoit, Y.; et al. Characterization of a set of tumor suppressor microRNAs in T cell acute lymphoblastic leukemia. Sci. Signal. 2014, 7, ra111. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.H.; Schiavinato, J.L.; Fraguas, M.S.; Lucena-Araujo, A.R.; Haddad, R.; Araujo, A.G.; Dalmazzo, L.F.; Rego, E.M.; Covas, D.T.; Zago, M.A.; et al. Potential roles of microRNA-29a in the molecular pathophysiology of T-cell acute lymphoblastic leukemia. Cancer Sci. 2015, 106, 1264–1277. [Google Scholar] [CrossRef] [PubMed]
- Coskun, E.; von der Heide, E.K.; Schlee, C.; Kuhnl, A.; Gokbuget, N.; Hoelzer, D.; Hofmann, W.K.; Thiel, E.; Baldus, C.D. The role of microRNA-196a and microRNA-196b as ERG regulators in acute myeloid leukemia and acute T-lymphoblastic leukemia. Leuk. Res. 2011, 35, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Zhang, X.; Jia, H.; Li, D.; Zhang, B.; Zhang, H.; Hong, M.; Jiang, T.; Jiang, Q.; Lu, J.; et al. An oncogenic role of miR-142-3p in human T-cell acute lymphoblastic leukemia (T-ALL) by targeting glucocorticoid receptor-alpha and cAMP/PKA pathways. Leukemia 2012, 26, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Nemes, K.; Csoka, M.; Nagy, N.; Mark, A.; Varadi, Z.; Danko, T.; Kovacs, G.; Kopper, L.; Sebestyen, A. Expression of Certain Leukemia/Lymphoma Related microRNAs and its Correlation with Prognosis in Childhood Acute Lymphoblastic Leukemia. Pathol. Oncol. Res. 2015, 21, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Ikari, J.; Smith, L.M.; Nelson, A.J.; Iwasawa, S.; Gunji, Y.; Farid, M.; Wang, X.Q.; Basma, H.; Feghali-Bostwick, C.; Liu, X.D.; et al. Effect of culture conditions on microRNA expression in primary adult control and COPD lung fibroblasts in vitro. In Vitro Cell. Dev. Biol.-Anim. 2015, 51, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Horn, P.; Castoldi, M.; Diehlmann, A.; Bork, S.; Saffrich, R.; Benes, V.; Blake, J.; Pfister, S.; Eckstein, V.; et al. Replicative Senescence of Mesenchymal Stem Cells: A Continuous and Organized Process. PLoS ONE 2008, 3, e2213. [Google Scholar] [CrossRef] [PubMed]
- Dawidowska, M.; Szarzyńska-Zawadzka, B.; Jaksik, R.; Drobna, M.; Kosmalska, M.; Łukasz, S.; Szczepańskiand, T.; Witt, M. miRNA Profiling in Pediatric T-ALL with Use of Next-Generation Sequencing: Focus on T-ALL Pathobiology and Heterogeneity. Blood 2017, 130, 1443. [Google Scholar]
- Davoren, P.A.; McNeill, R.E.; Lowery, A.J.; Kerin, M.J.; Miller, N. Identification of suitable endogenous control genes for microRNA gene expression analysis in human breast cancer. BMC Mol. Biol. 2008, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Das, M.K.; Andreassen, R.; Haugen, T.B.; Furu, K. Identification of Endogenous Controls for Use in miRNA Quantification in Human Cancer Cell Lines. Cancer Genom. Proteom. 2016, 13, 63–68. [Google Scholar]
- Muller, V.; Gade, S.; Steinbach, B.; Loibl, S.; von Minckwitz, G.; Untch, M.; Schwedler, K.; Lubbe, K.; Schem, C.; Fasching, P.A.; et al. Changes in serum levels of miR-21, miR-210, and miR-373 in HER2-positive breast cancer patients undergoing neoadjuvant therapy: A translational research project within the Geparquinto trial. Breast Cancer Res. Treat. 2014, 147, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Song, J.N.; Bai, Z.G.; Han, W.; Zhang, J.; Meng, H.; Bi, J.T.; Ma, X.M.; Han, S.W.; Zhang, Z.T. Identification of Suitable Reference Genes for qPCR Analysis of Serum microRNA in Gastric Cancer Patients. Dig. Dis. Sci. 2012, 57, 897–904. [Google Scholar] [CrossRef] [PubMed]
- McDermott, A.M.; Kerin, M.J.; Miller, N. Identification and Validation of miRNAs as Endogenous Controls for RQ-PCR in Blood Specimens for Breast Cancer Studies. PLoS ONE 2013, 8, e83718. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.L.; Xiao, P.; Chen, D.L.; Xu, L.; Zhang, B.H. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Derveaux, S.; Vandesompele, J.; Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 2010, 50, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Henry, N.L.; Hayes, D.F. Cancer biomarkers. Mol. Oncol. 2012, 6, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Thellin, O.; Zorzi, W.; Lakaye, B.; De Borman, B.; Coumans, B.; Hennen, G.; Grisar, T.; Igout, A.; Heinen, E. Housekeeping genes as internal standards: Use and limits. J. Biotechnol. 1999, 75, 291–295. [Google Scholar] [CrossRef]
- Mestdagh, P.; Van Vlierberghe, P.; De Weer, A.; Muth, D.; Westermann, F.; Speleman, F.; Vandesompele, J. A novel and universal method for microRNA RT-qPCR data normalization. Genome Biol. 2009, 10, R64. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Peng, X.; Luo, J.; Gou, D. Identification of circulating miRNA biomarkers based on global quantitative real-time PCR profiling. J. Anim. Sci. Biotechnol. 2012, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.H.; Mestdagh, P.; Vandesompele, J.; Kerin, M.J.; Miller, N. MicroRNA expression profiling to identify and validate reference genes for relative quantification in colorectal cancer. BMC Cancer 2010, 10, 173. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, R.; Sacan, A. A novel method for the normalization of microRNA RT-PCR data. BMC Med. Genom. 2013, 6 (Suppl. 1), S14. [Google Scholar] [CrossRef]
- McIntyre, L.M.; Lopiano, K.K.; Morse, A.M.; Amin, V.; Oberg, A.L.; Young, L.J.; Nuzhdin, S.V. RNA-seq: Technical variability and sampling. BMC Genom. 2011, 12, 293. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Noguchi, S.; Kumazaki, M.; Shinohara, H.; Miki, K.; Naoe, T.; Akao, Y. Epigenetic regulation of microRNA-128a expression contributes to the apoptosis-resistance of human T-cell leukaemia Jurkat cells by modulating expression of Fas-associated protein with death domain (FADD). Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 590–602. [Google Scholar] [CrossRef] [PubMed]
- Git, A.; Dvinge, H.; Salmon-Divon, M.; Osborne, M.; Kutter, C.; Hadfield, J.; Bertone, P.; Caldas, C. Systematic comparison of microarray profiling, real-time PCR, and next-generation sequencing technologies for measuring differential microRNA expression. RNA 2010, 16, 991–1006. [Google Scholar] [CrossRef] [PubMed]
- Baras, A.S.; Mitchell, C.J.; Myers, J.R.; Gupta, S.; Weng, L.-C.; Ashton, J.M.; Cornish, T.C.; Pandey, A.; Halushka, M.K. miRge-A Multiplexed Method of Processing Small RNA-Seq Data to Determine MicroRNA Entropy. PLoS ONE 2015, 10, e0143066. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Candidate EN miRNAs | Criteria for Selection | |||
|---|---|---|---|---|
| miRNA Name | Stability in miRNA-Seq | Thermo Fisher Scientific Recommendation | Literature Data | |
| Stability Score * | Mean Read Count | |||
| hsa-let-7a-5p | 0.25 | 330,280 | – | [44] |
| hsa-miR-30d-5p | 0.25 | 98,732 | – | – |
| hsa-miR-92a-3p | 0.31 | 630,854 | Suitable endogenous control for tissue samples | – |
| hsa-miR-93-5p | 0.36 | 9838 | Suitable endogenous control | [45] |
| hsa-let-7f-5p | 0.37 | 326,028 | – | – |
| hsa-miR-25-3p | 0.45 | 108,466 | Suitable endogenous control | [45] |
| hsa-miR-26a-5p | 0.5 | 176,895 | Suitable endogenous control for breast and heart tissue | – |
| hsa-miR-21-5p | 0.5 | 116,490 | Suitable endogenous control for tissue samples | – |
| hsa-miR-16-5p | 0.52 | 5746 | Suitable endogenous control | [46,47,48] |
| hsa-let-7g-5p | 0.56 | 194,895 | Suitable endogenous control | – |
| miRNA Name | TaqMan Advanced miRNA Assay Name | Standard Curve | Amplification Efficiency (%) | |
|---|---|---|---|---|
| Slope | R2 (Correlation Coefficient) | |||
| Candidate EN miRNAs | ||||
| hsa-let-7a-5p | 478575_mir | −3.292 | 0.991 | 101 |
| hsa-miR-30d-5p | 478606_mir | −3.371 | 0.996 | 98 |
| hsa-miR-92a-3p | 477827_mir | −3.549 | 0.994 | 91 |
| hsa-miR-93-5p | 478210_mir | −3.371 | 0.996 | 98 |
| hsa-let-7f-5p | 478578_mir | −3.626 | 0.985 | 89 |
| hsa-miR-25-3p | 477994_mir | −3.533 | 0.994 | 92 |
| hsa-miR-26a-5p | 477995_mir | −3.558 | 0.997 | 91 |
| hsa-miR-21-5p | 477975_mir | −3.516 | 0.994 | 92 |
| hsa-miR-16-5p | 477860_mir | −3.582 | 0.997 | 90 |
| hsa-let-7g-5p | 478580_mir | −3.338 | 0.959 | 99 |
| Selected miRNAs Overexpressed in T-ALL vs. Control | ||||
| hsa-miR-181a-5p | 477857_mir | −3.872 | 0.966 | 81 |
| hsa-miR-128-3p | 477892_mir | −3.406 | 0.99 | 97 |
| hsa-miR-20b-5p | 477804_mir | −3.570 | 0.997 | 91 |
| miRNA | All Samples | T-ALL Samples | Normal BM T-Lymphocytes | Thymocytes | T-ALL Cell Lines | p-adj | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cq | SD | Cq | SD | Cq | SD | Cq | SD | Cq | SD | ||
| hsa-miR-92a-3p | 21.54 | 1.68 | 21.38 | 1.61 | 20.37 | 0.92 | 23.22 | 1.78 | 21.65 | 1.53 | 0.173 |
| hsa-miR-16-5p | 22.58 | 1.66 | 22.18 | 1.42 | 23.61 | 1.79 | 22.89 | 2.42 | 23.40 | 1.82 | 0.192 |
| hsa-miR-25-3p | 23.00 | 1.64 | 22.70 | 1.51 | 24.11 | 1.76 | 22.60 | 1.90 | 23.67 | 1.83 | 0.240 |
| hsa-let-7a-5p | 23.08 | 1.91 | 22.66 | 1.83 | 23.98 | 1.99 | 24.22 | 2.15 | 23.73 | 1.89 | 0.240 |
| hsa-miR-26a-5p | 23.17 | 2.07 | 22.48 | 1.42 | 23.57 | 2.21 | 24.51 | 2.70 | 24.99 | 2.50 | 0.028 |
| hsa-let-7f-5p | 24.15 | 1.96 | 23.69 | 1.77 | 24.26 | 2.07 | 24.88 | 2.20 | 25.43 | 2.15 | 0.192 |
| hsa-miR-93-5p | 24.89 | 1.55 | 24.76 | 1.54 | 26.10 | 1.64 | 25.31 | 1.77 | 24.63 | 1.47 | 0.308 |
| hsa-let-7g-5p | 25.46 | 1.95 | 24.98 | 1.72 | 25.18 | 2.12 | 26.55 | 2.46 | 26.82 | 2.02 | 0.163 |
| hsa-miR-21-5p | 26.16 | 2.20 | 25.55 | 1.61 | 26.62 | 2.36 | 28.13 | 2.19 | 27.58 | 2.80 | 0.064 |
| hsa-miR-30d-5p | 26.18 | 1.49 | 25.91 | 1.38 | 26.98 | 1.62 | 26.83 | 1.90 | 26.59 | 1.56 | 0.308 |
| miRNA Name | Comprehensive Ranking Stability Score |
|---|---|
| hsa-miR-16-5p | 2.11 |
| hsa-miR-30d-5p | 2.71 |
| hsa-miR-25-3p | 2.99 |
| hsa-let-7g-5p | 3.98 |
| hsa-let-7a-5p | 4.36 |
| hsa-miR-93-5p | 4.53 |
| hsa-let-7f-5p | 4.58 |
| hsa-miR-92a-3p | 5.66 |
| hsa-miR-21-5p | 8.74 |
| hsa-miR-26a-5p | 10 |
| miRNA Name | miRNA-Seq | RT-qPCR | ||
|---|---|---|---|---|
| Log2 Fold Change | p-Value | Log2 Fold Change | p-Value | |
| hsa-miR-128-3p | 2.814 | <0.001 | 2.373 | <0.001 |
| hsa-miR-181a-5p | 2.362 | <0.001 | 5.951 | <0.001 |
| hsa-miR-20b-5p | 3.522 | <0.001 | 1.329 | <0.001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drobna, M.; Szarzyńska-Zawadzka, B.; Daca-Roszak, P.; Kosmalska, M.; Jaksik, R.; Witt, M.; Dawidowska, M. Identification of Endogenous Control miRNAs for RT-qPCR in T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2018, 19, 2858. https://doi.org/10.3390/ijms19102858
Drobna M, Szarzyńska-Zawadzka B, Daca-Roszak P, Kosmalska M, Jaksik R, Witt M, Dawidowska M. Identification of Endogenous Control miRNAs for RT-qPCR in T-Cell Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences. 2018; 19(10):2858. https://doi.org/10.3390/ijms19102858
Chicago/Turabian StyleDrobna, Monika, Bronisława Szarzyńska-Zawadzka, Patrycja Daca-Roszak, Maria Kosmalska, Roman Jaksik, Michał Witt, and Małgorzata Dawidowska. 2018. "Identification of Endogenous Control miRNAs for RT-qPCR in T-Cell Acute Lymphoblastic Leukemia" International Journal of Molecular Sciences 19, no. 10: 2858. https://doi.org/10.3390/ijms19102858
APA StyleDrobna, M., Szarzyńska-Zawadzka, B., Daca-Roszak, P., Kosmalska, M., Jaksik, R., Witt, M., & Dawidowska, M. (2018). Identification of Endogenous Control miRNAs for RT-qPCR in T-Cell Acute Lymphoblastic Leukemia. International Journal of Molecular Sciences, 19(10), 2858. https://doi.org/10.3390/ijms19102858