Hepatitis C Virus: Viral Quasispecies and Genotypes


Abstract
:1. Background
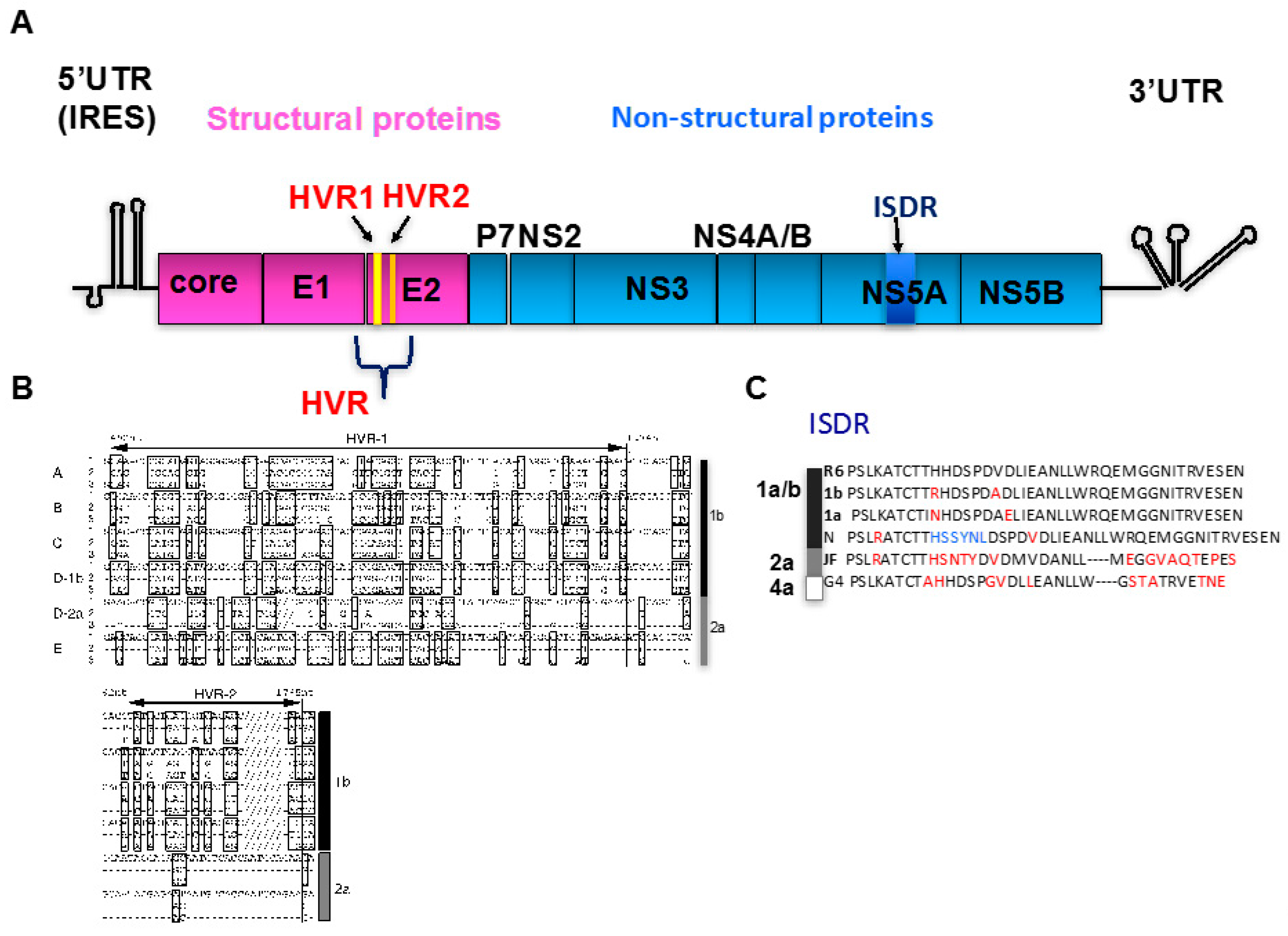
2. Viral Quasispecies
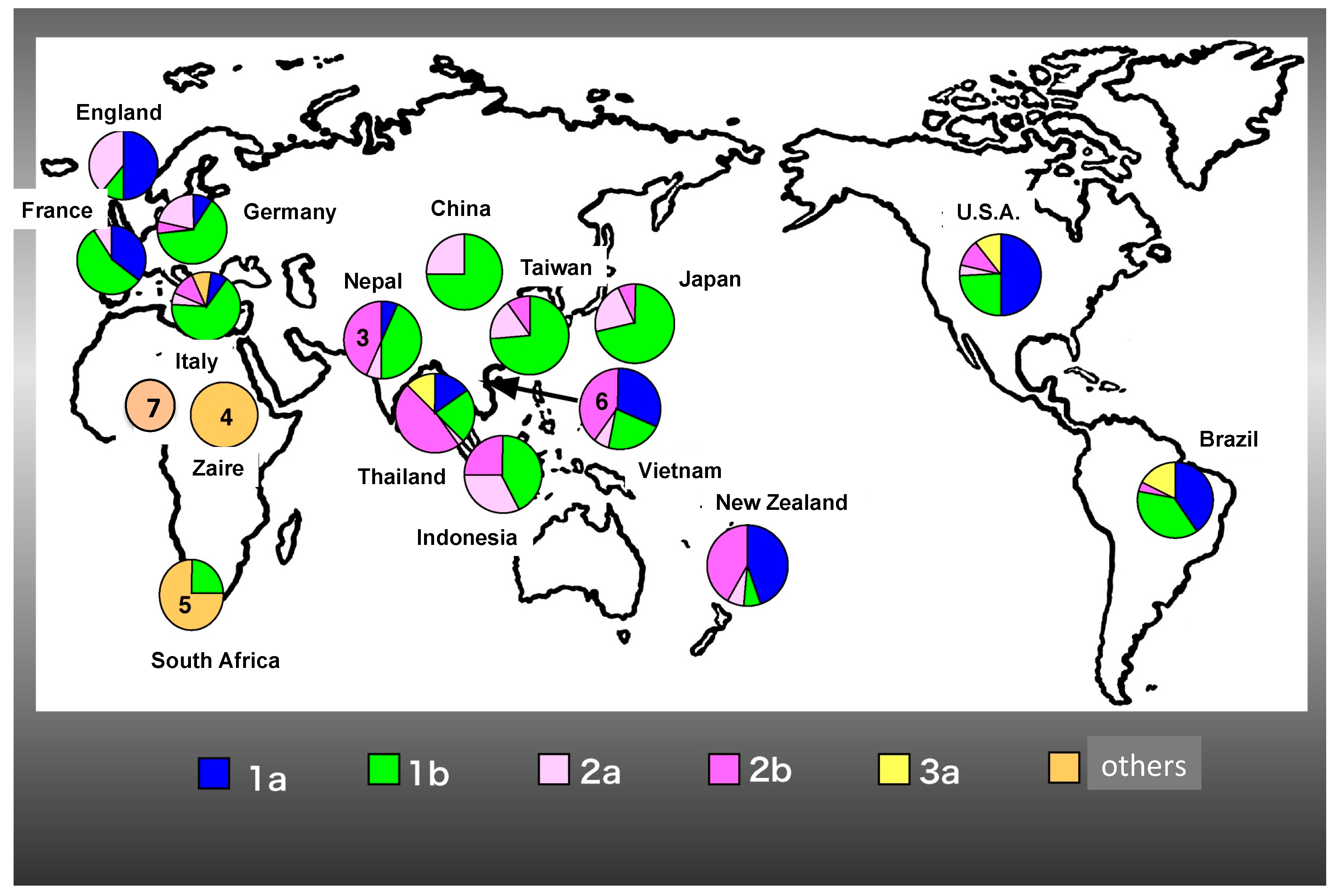
3. HCV Genotypes
4. Clinical Significance of Genotype
5. DAAs (Direct-Acting Antivirals) and Resistance Mutations
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| HCV | Hepatitis C virus |
| DAA | Direct-acting antiviral |
| HVR | Hypervariable region |
| NS | Nonstructural |
| IFN | Interferon |
| ISDR | IFN-sensitivity-determining region |
| UTR | Untranslated region |
| RBV | Ribavirin |
| SVR | Sustained virologic response |
| FDA | Food and Drug Administration |
References
- Feinstone, S.M.; Kapikian, A.Z.; Purcell, R.H.; Alter, H.J.; Holland, P.V. Transfusion-associated hepatitis not due to viral hepatitis type A or B. N. Engl. J. Med. 1975, 292, 767–770. [Google Scholar] [CrossRef] [PubMed]
- Choo, Q.L.; Kuo, G.; Weiner, A.J.; Overby, L.R.; Bradley, D.W.; Houghton, M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science 1989, 244, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kato, N.; Cho, M.J.; Shimotohno, K. A novel sequence found at the 3’ terminus of hepatitis C virus genome. Biochem. Biophys. Res. Commun. 1995, 215, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Tsukiyama-Kohara, K.; Iizuka, N.; Kohara, M.; Nomoto, A. Internal ribosome entry site within hepatitis C virus RNA. J. Virol. 1992, 66, 1476–1483. [Google Scholar] [PubMed]
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Farci, P.; Bukh, J.; Purcell, R.H. The quasispecies of hepatitis C virus and the host immune response. Springer Semin. Immunopathol. 1997, 19, 5–26. [Google Scholar] [CrossRef] [PubMed]
- Powdrill, M.H.; Tchesnokov, E.P.; Kozak, R.A.; Russell, R.S.; Martin, R.; Svarovskaia, E.S.; Mo, H.; Kouyos, R.D.; Gotte, M. Contribution of a mutational bias in hepatitis C virus replication to the genetic barrier in the development of drug resistance. Proc. Natl. Acad. Sci. USA 2011, 108, 20509–20513. [Google Scholar] [CrossRef] [PubMed]
- Weiner, A.J.; Brauer, M.J.; Rosenblatt, J.; Richman, K.H.; Tung, J.; Crawford, K.; Bonino, F.; Saracco, G.; Choo, Q.L.; Houghton, M.; et al. Variable and hypervariable domains are found in the regions of HCV corresponding to the flavivirus envelope and NS1 proteins and the pestivirus envelope glycoproteins. Virology 1991, 180, 842–848. [Google Scholar] [CrossRef]
- Weiner, A.J.; Geysen, H.M.; Christopherson, C.; Hall, J.E.; Mason, T.J.; Saracco, G.; Bonino, F.; Crawford, K.; Marion, C.D.; Crawford, K.A.; et al. Evidence for immune selection of hepatitis C virus (HCV) putative envelope glycoprotein variants: Potential role in chronic HCV infections. Proc. Natl. Acad. Sci. USA 1992, 89, 3468–3472. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Ootsuyama, Y.; Ohkoshi, S.; Nakazawa, T.; Sekiya, H.; Hijikata, M.; Shimotohno, K. Characterization of hypervariable regions in the putative envelope protein of hepatitis C virus. Biochem. Biophys. Res. Commun. 1992, 189, 119–127. [Google Scholar] [CrossRef]
- Kato, N.; Sekiya, H.; Ootsuyama, Y.; Nakazawa, T.; Hijikata, M.; Ohkoshi, S.; Shimotohno, K. Humoral immune response to hypervariable region 1 of the putative envelope glycoprotein (gp70) of hepatitis C virus. J. Virol. 1993, 67, 3923–3930. [Google Scholar] [PubMed]
- Cerny, A.; Chisari, F.V. Immunological aspects of HCV infection. Intervirology 1994, 37, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Higashi, K.; Tsukiyama-Kohara, K.; Tanaka, T.; Tanaka, E.; Kiyosawa, K.; Kohara, M. Characterization of hypervariable region in hepatitis C virus envelope protein during acute and chronic infection. Arch. Virol. 2005, 150, 883–898. [Google Scholar] [CrossRef] [PubMed]
- Tsukiyama-Kohara, K.; Tone, S.; Maruyama, I.; Inoue, K.; Katsume, A.; Nuriya, H.; Ohmori, H.; Ohkawa, J.; Taira, K.; Hoshikawa, Y.; et al. Activation of the CKI-CDK-Rb-E2F pathway in full genome hepatitis C virus-expressing cells. J. Biol. Chem. 2004, 279, 14531–14541. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Kanai, N.; Mishiro, S. Full-length nucleotide sequence of a Japanese hepatitis C virus isolate (HC-J1) with high homology to USA isolates. Nucleic Acids Res. 1992, 20, 6410. [Google Scholar] [CrossRef] [PubMed]
- Beard, M.R.; Abell, G.; Honda, M.; Carroll, A.; Gartland, M.; Clarke, B.; Suzuki, K.; Lanford, R.; Sangar, D.V.; Lemon, S.M. An infectious molecular clone of a Japanese genotype 1b hepatitis C virus. Hepatology 1999, 30, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Wakita, T.; Pietschmann, T.; Kato, T.; Date, T.; Miyamoto, M.; Zhao, Z.; Murthy, K.; Habermann, A.; Krausslich, H.G.; Mizokami, M.; et al. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nat. Med. 2005, 11, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, N.; Sakuma, I.; Asahina, Y.; Kurosaki, M.; Murakami, T.; Yamamoto, C.; Ogura, Y.; Izumi, N.; Marumo, F.; Sato, C. Mutations in the nonstructural protein 5A gene and response to interferon in patients with chronic hepatitis C virus 1b infection. N. Engl. J. Med. 1996, 334, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Castelain, S.; Khorsi, H.; Roussel, J.; Francois, C.; Jaillon, O.; Capron, D.; Penin, F.; Wychowski, C.; Meurs, E.; Duverlie, G.; et al. Variability of the nonstructural 5A protein of hepatitis C virus type 3a isolates and relation to interferon sensitivity. J. Infect. Dis. 2002, 185, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Tsukiyama-Kohara, K.; Yamaguchi, K.; Yagi, S.; Tanaka, S.; Hasegawa, A.; Ohta, Y.; Hattori, N.; Kohara, M. Significance of specific antibody assay for genotyping of hepatitis C virus. Hepatology 1994, 19, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, G.; Raffa, G.; Restuccia, T.; Pollicino, T.; Brancatelli, S.; Raimondo, G. Is investigation of hepatitis C virus NS5A gene heterogeneity a tool for predicting long-lasting response to interferon therapy in patients with HCV-1b chronic hepatitis? J. Viral Hepat. 2002, 9, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Chung, R.T.; Monto, A.; Dienstag, J.L.; Kaplan, L.M. Mutations in the NS5A region do not predict interferon-responsiveness in american patients infected with genotype 1b hepatitis C virus. J. Med. Virol. 1999, 58, 353–358. [Google Scholar] [CrossRef]
- Shen, C.; Hu, T.; Shen, L.; Gao, L.; Xie, W.; Zhang, J. Mutations in ISDR of NS5A gene influence interferon efficacy in Chinese patients with chronic hepatitis C virus genotype 1b infection. J. Gastroenterol. Hepatol. 2007, 22, 1898–1903. [Google Scholar] [CrossRef] [PubMed]
- Tsukiyama-Kohara, K.; Kohara, M.; Yamaguchi, K.; Maki, N.; Toyoshima, A.; Miki, K.; Tanaka, S.; Hattori, N.; Nomoto, A. A second group of hepatitis C viruses. Virus Genes 1991, 5, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Tsukiyama-Kohara, K.; Yamaguchi, K.; Maki, N.; Ohta, Y.; Miki, K.; Mizokami, M.; Ohba, K.; Tanaka, S.; Hattori, N.; Nomoto, A.; et al. Antigenicities of group I and II hepatitis C virus polypeptides—molecular basis of diagnosis. Virology 1993, 192, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Bukh, J.; Kuiken, C.; Muerhoff, A.S.; Rice, C.M.; Stapleton, J.T.; Simmonds, P. Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: Updated criteria and genotype assignment web resource. Hepatology 2014, 59, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Humphreys, I.; Flaxman, A.; Brown, A.; Cooke, G.S.; Pybus, O.G.; Barnes, E. Global distribution and prevalence of hepatitis C virus genotypes. Hepatology 2015, 61, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.G.; Sablon, E.; Chamberland, J.; Fournier, E.; Dandavino, R.; Tremblay, C.L. Hepatitis C virus genotype 7, a new genotype originating from central Africa. J. Clin. Microbiol. 2015, 53, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.C.; Natarajan, V.; Pandey, P.; Gupta, E.; Sharma, S.; Tripathi, R.; Kumar, M.S.; Kazim, S.N.; Sarin, S.K. Identification of Indian sub-continent as hotspot for HCV genotype 3a origin by Bayesian evolutionary reconstruction. Infect. Genet. Evol. 2014, 28, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Akkarathamrongsin, S.; Hacharoen, P.; Tangkijvanich, P.; Theamboonlers, A.; Tanaka, Y.; Mizokami, M.; Poovorawan, Y. Molecular epidemiology and genetic history of hepatitis C virus subtype 3a infection in Thailand. Intervirology 2013, 56, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Iles, J.C.; Raghwani, J.; Harrison, G.L.A.; Pepin, J.; Djoko, C.F.; Tamoufe, U.; LeBreton, M.; Schneider, B.S.; Fair, J.N.; Tshala, F.M.; et al. Phylogeography and epidemic history of hepatitis C virus genotype 4 in Africa. Virology 2014, 464–465, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yuan, M.; Lu, L.; Lu, T.; Xia, W.; Pham, V.H.; Vo, A.X.; Nguyen, M.H.; Abe, K. The genetic diversity and evolutionary history of hepatitis C virus in Vietnam. Virology 2014, 468–470, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Stelzl, E.; Haas, B.; Bauer, B.; Zhang, S.; Fiss, E.H.; Hillman, G.; Hamilton, A.T.; Mehta, R.; Heil, M.L.; Marins, E.G.; et al. First identification of a recombinant form of hepatitis C virus in Austrian patients by full-genome next generation sequencing. PLoS ONE 2017, 12, e0181273. [Google Scholar] [CrossRef] [PubMed]
- Kurbanov, F.; Tanaka, Y.; Chub, E.; Maruyama, I.; Azlarova, A.; Kamitsukasa, H.; Ohno, T.; Bonetto, S.; Moreau, I.; Fanning, L.J.; et al. Molecular epidemiology and interferon susceptibility of the natural recombinant hepatitis C virus strain RF1_2k/1b. J. Infect. Dis. 2008, 198, 1448–1456. [Google Scholar] [CrossRef] [PubMed]
- Baldo, V.; Floreani, A.; Menegon, T.; Angiolelli, G.; Trivello, R. Prevalence of antibodies against hepatitis C virus in the elderly: A seroepidemiological study in a nursing home and in an open population. The Collaborative Group. Gerontology 2000, 46, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Ezzikouri, S.; Kimura, K.; Sunagozaka, H.; Kaneko, S.; Inoue, K.; Nishimura, T.; Hishima, T.; Kohara, M.; Tsukiyama-Kohara, K. Serum DHCR24 auto-antibody as a new biomarker for progression of hepatitis C. EBioMedicine 2015, 2, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Kohara, M.; Tanaka, T.; Tsukiyama-Kohara, K.; Tanaka, S.; Mizokami, M.; Lau, J.Y.; Hattori, N. Hepatitis C virus genotypes 1 and 2 respond to interferon-alpha with different virologic kinetics. J. Infect. Dis. 1995, 172, 934–938. [Google Scholar] [PubMed]
- Orito, E.; Mizokami, M.; Mizoguchi, N.; Ohba, K.; Tohnai, M.; Yamanaka, H.; Oguri, T.; Hirashima, N.; Koide, T.; Kano, H.; et al. Hepatitis C virus serotype II responds more favorably to interferon-alpha therapy. J. Hepatol. 1994, 21, 130–132. [Google Scholar] [CrossRef]
- Lonardo, A.; Adinolfi, L.E.; Restivo, L.; Ballestri, S.; Romagnoli, D.; Baldelli, E.; Nascimbeni, F.; Loria, P. Pathogenesis and significance of hepatitis C virus steatosis: An update on survival strategy of a successful pathogen. World J. Gastroenterol. 2014, 20, 7089–7103. [Google Scholar] [CrossRef] [PubMed]
- Bechmann, L.P.; Hannivoort, R.A.; Gerken, G.; Hotamisligil, G.S.; Trauner, M.; Canbay, A. The interaction of hepatic lipid and glucose metabolism in liver diseases. J. Hepatol. 2012, 56, 952–964. [Google Scholar] [CrossRef] [PubMed]
- Moucari, R.; Asselah, T.; Cazals-Hatem, D.; Voitot, H.; Boyer, N.; Ripault, M.P.; Sobesky, R.; Martinot-Peignoux, M.; Maylin, S.; Nicolas-Chanoine, M.H.; et al. Insulin resistance in chronic hepatitis C: Association with genotypes 1 and 4, serum HCV RNA level, and liver fibrosis. Gastroenterology 2008, 134, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Patel, K.; Chuang, W.L.; Lawitz, E.J.; Rodriguez-Torres, M.; Rustgi, V.K.; Flisiak, R.; Pianko, S.; Diago, M.; Arora, S.; et al. Viral clearance is associated with improved insulin resistance in genotype 1 chronic hepatitis C but not genotype 2/3. Gut 2012, 61, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Antaki, N.; Craxi, A.; Kamal, S.; Moucari, R.; Van der Merwe, S.; Haffar, S.; Gadano, A.; Zein, N.; Lai, C.L.; Pawlotsky, J.M.; et al. The neglected hepatitis C virus genotypes 4, 5 and 6: An international consensus report. Liver Int. 2010, 30, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Hoofnagle, J.H.; Seeff, L.B. Peginterferon and ribavirin for chronic hepatitis C. N. Engl. J. Med. 2006, 355, 2444–2451. [Google Scholar] [CrossRef] [PubMed]
- Spengler, U. Direct antiviral agents (DAAs)—A new age in the treatment of hepatitis C virus infection. Pharmacol. Ther. 2017. [Google Scholar] [CrossRef] [PubMed]
- Halfon, P.; Locarnini, S. Hepatitis C virus resistance to protease inhibitors. J. Hepatol. 2011, 55, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Pilot-Matias, T.; Tripathi, R.; Cohen, D.; Gaultier, I.; Dekhtyar, T.; Lu, L.; Reisch, T.; Irvin, M.; Hopkins, T.; Pithawalla, R.; et al. In vitro and in vivo antiviral activity and resistance profile of the hepatitis C virus NS3/4A protease inhibitor ABT-450. Antimicrob. Agents Chemother. 2015, 59, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Bacon, B.R.; Gordon, S.C.; Lawitz, E.; Marcellin, P.; Vierling, J.M.; Zeuzem, S.; Poordad, F.; Goodman, Z.D.; Sings, H.L.; Boparai, N.; et al. Boceprevir for previously treated chronic HCV genotype 1 infection. N. Engl. J. Med. 2011, 364, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, I.M.; McHutchison, J.G.; Dusheiko, G.; Di Bisceglie, A.M.; Reddy, K.R.; Bzowej, N.H.; Marcellin, P.; Muir, A.J.; Ferenci, P.; Flisiak, R.; et al. Telaprevir for previously untreated chronic hepatitis C virus infection. N. Engl. J. Med. 2011, 364, 2405–2416. [Google Scholar] [CrossRef] [PubMed]
- Poordad, F.; McCone, J., Jr.; Bacon, B.R.; Bruno, S.; Manns, M.P.; Sulkowski, M.S.; Jacobson, I.M.; Reddy, K.R.; Goodman, Z.D.; Boparai, N.; et al. Boceprevir for untreated chronic HCV genotype 1 infection. N. Engl. J. Med. 2011, 364, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Katsume, A.; Tokunaga, Y.; Hirata, Y.; Munakata, T.; Saito, M.; Hayashi, H.; Okamoto, K.; Ohmori, Y.; Kusanagi, I.; Fujiwara, S.; et al. A serine palmitoyltransferase inhibitor blocks hepatitis C virus replication in human hepatocytes. Gastroenterology 2013, 145, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Ezzikouri, S.; Nishimura, T.; Kohara, M.; Benjelloun, S.; Kino, Y.; Inoue, K.; Matsumori, A.; Tsukiyama-Kohara, K. Inhibitory effects of Pycnogenol(R) on hepatitis C virus replication. Antivir. Res. 2015, 113, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Bartenschlager, R.; Lohmann, V.; Penin, F. The molecular and structural basis of advanced antiviral therapy for hepatitis C virus infection. Nat. Rev. Microbiol. 2013, 11, 482–496. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Treatment | Mutation | Virus Gene Region | Frequency (%) |
|---|---|---|---|
| IC50 × 6 | V36A | NS3 | 18.1 |
| 14 passages | T54V | NS3 | 26.9 |
| A156T | NS3 | 12.9 | |
| Q181H | NS5A | 25.2 | |
| P223S | NS5A | 23.3 | |
| 417P | NS5A | 15.8 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsukiyama-Kohara, K.; Kohara, M. Hepatitis C Virus: Viral Quasispecies and Genotypes. Int. J. Mol. Sci. 2018, 19, 23. https://doi.org/10.3390/ijms19010023
Tsukiyama-Kohara K, Kohara M. Hepatitis C Virus: Viral Quasispecies and Genotypes. International Journal of Molecular Sciences. 2018; 19(1):23. https://doi.org/10.3390/ijms19010023
Chicago/Turabian StyleTsukiyama-Kohara, Kyoko, and Michinori Kohara. 2018. "Hepatitis C Virus: Viral Quasispecies and Genotypes" International Journal of Molecular Sciences 19, no. 1: 23. https://doi.org/10.3390/ijms19010023
APA StyleTsukiyama-Kohara, K., & Kohara, M. (2018). Hepatitis C Virus: Viral Quasispecies and Genotypes. International Journal of Molecular Sciences, 19(1), 23. https://doi.org/10.3390/ijms19010023