The Herbal Bitter Drug Gentiana lutea Modulates Lipid Synthesis in Human Keratinocytes In Vitro and In Vivo

Abstract
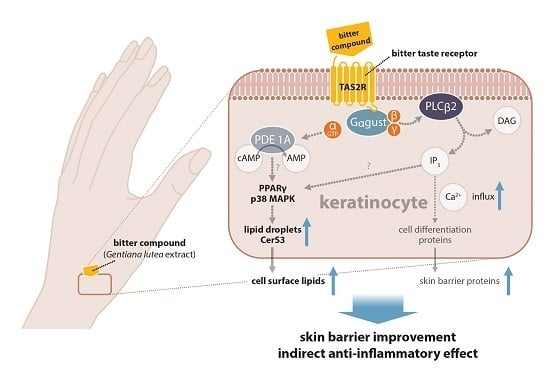
:
1. Introduction
2. Results
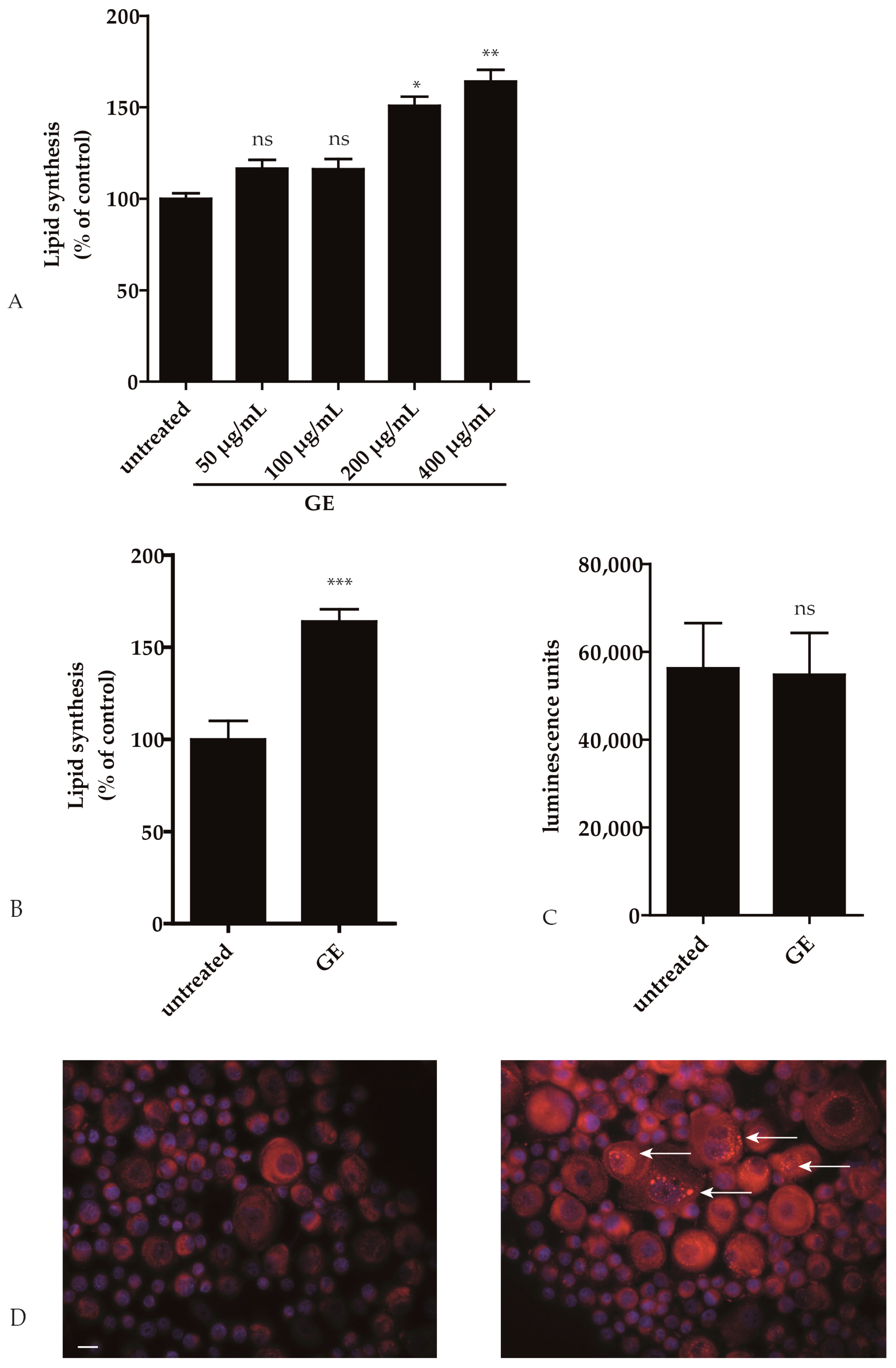
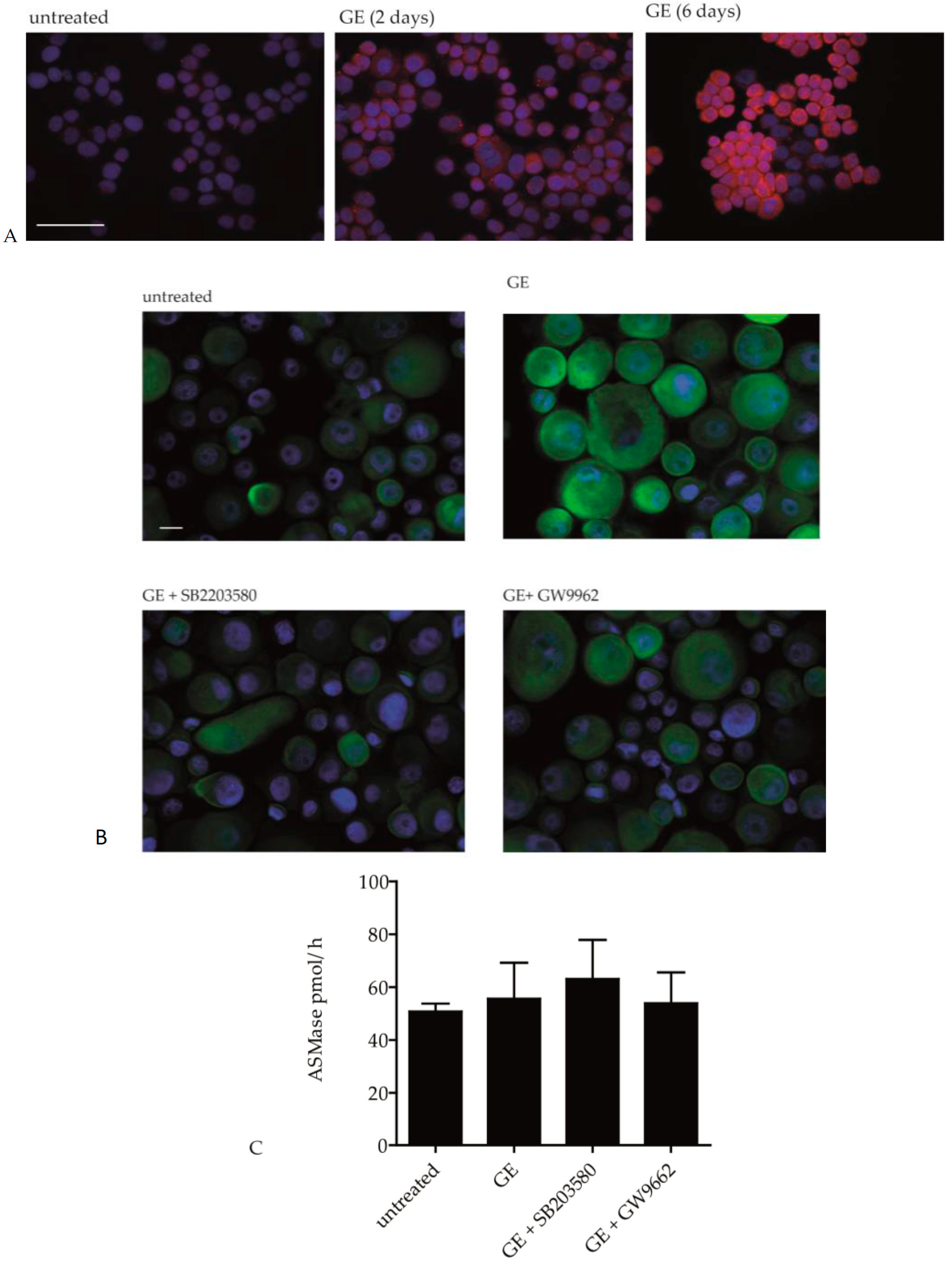
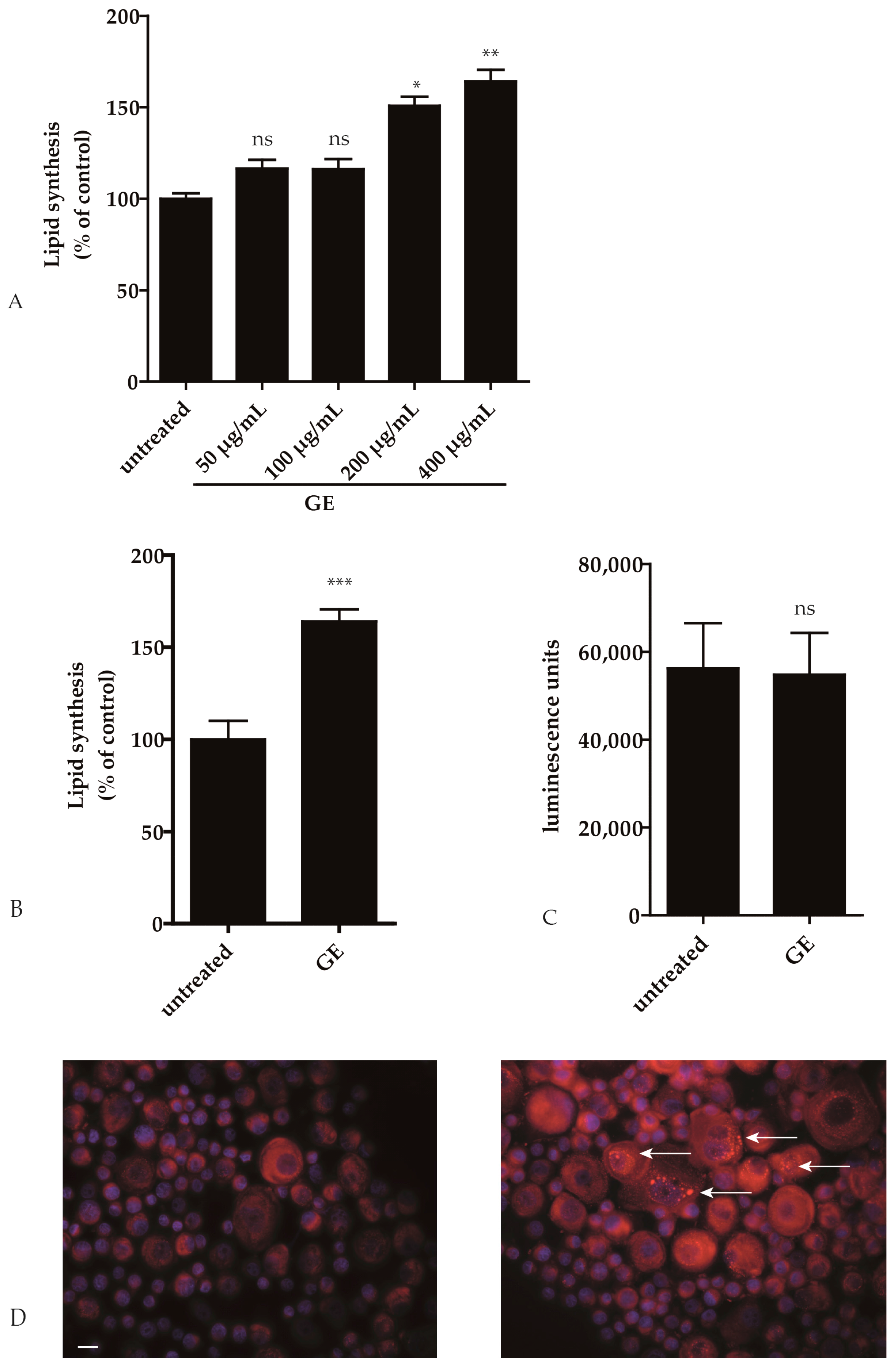
2.1. GE Enhances Lipid Synthesis in Keratinocytes
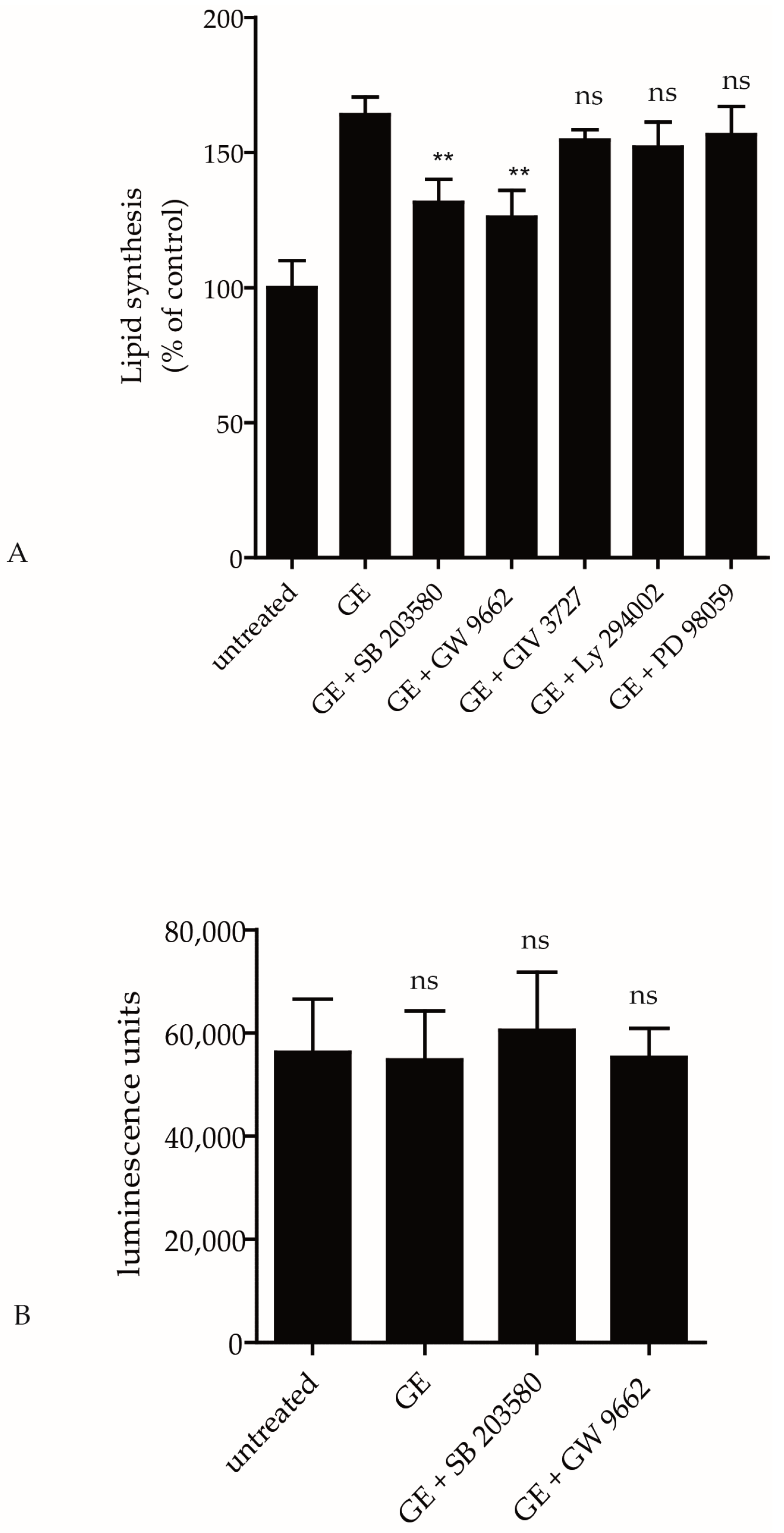
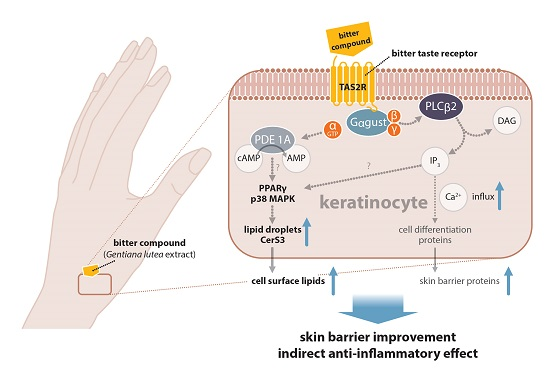
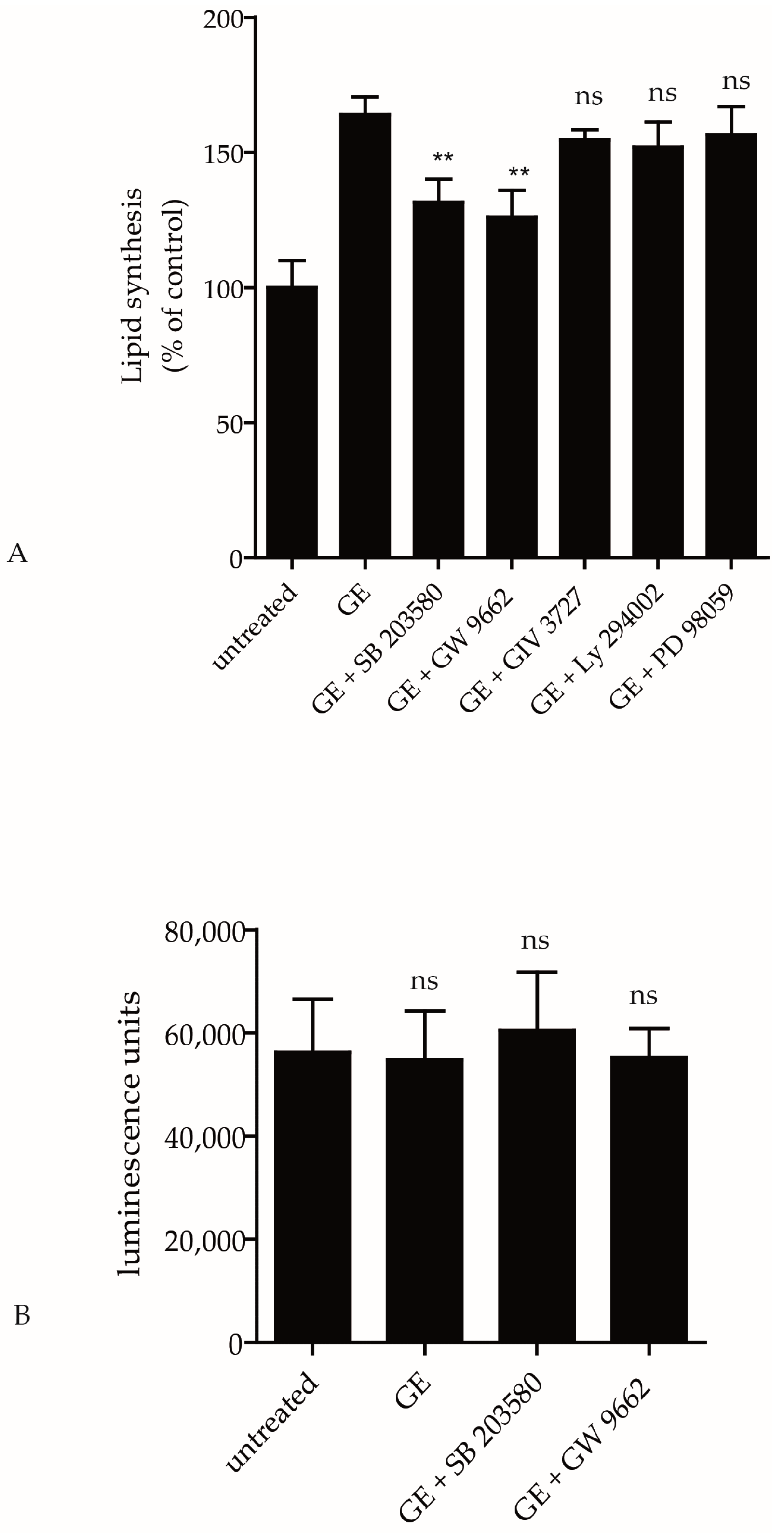
2.2. Pathways Involved in GE-Induced Lipid Production
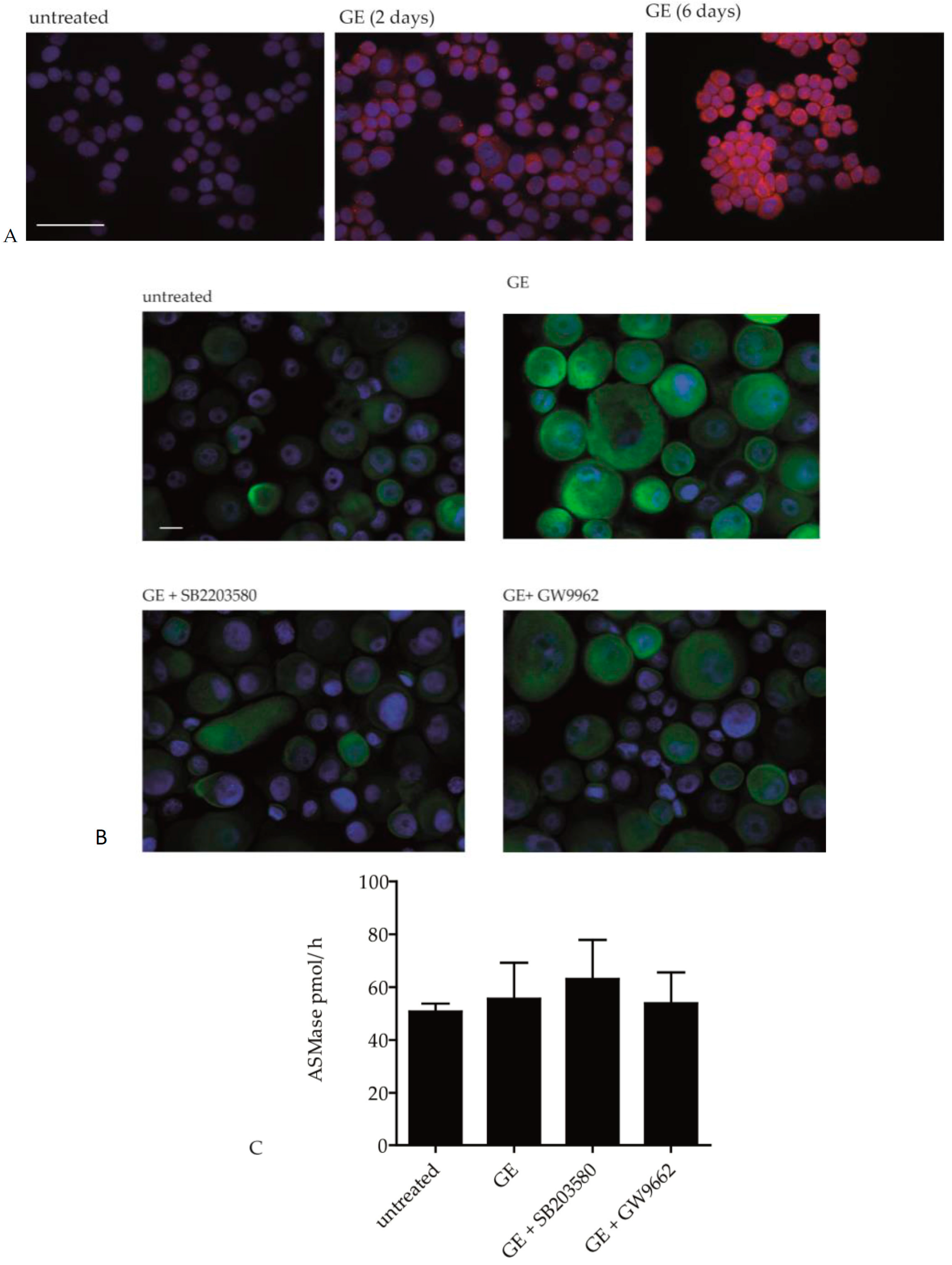
2.3. GE Modulates the Activity of Ceramide Metabolic Enzymes
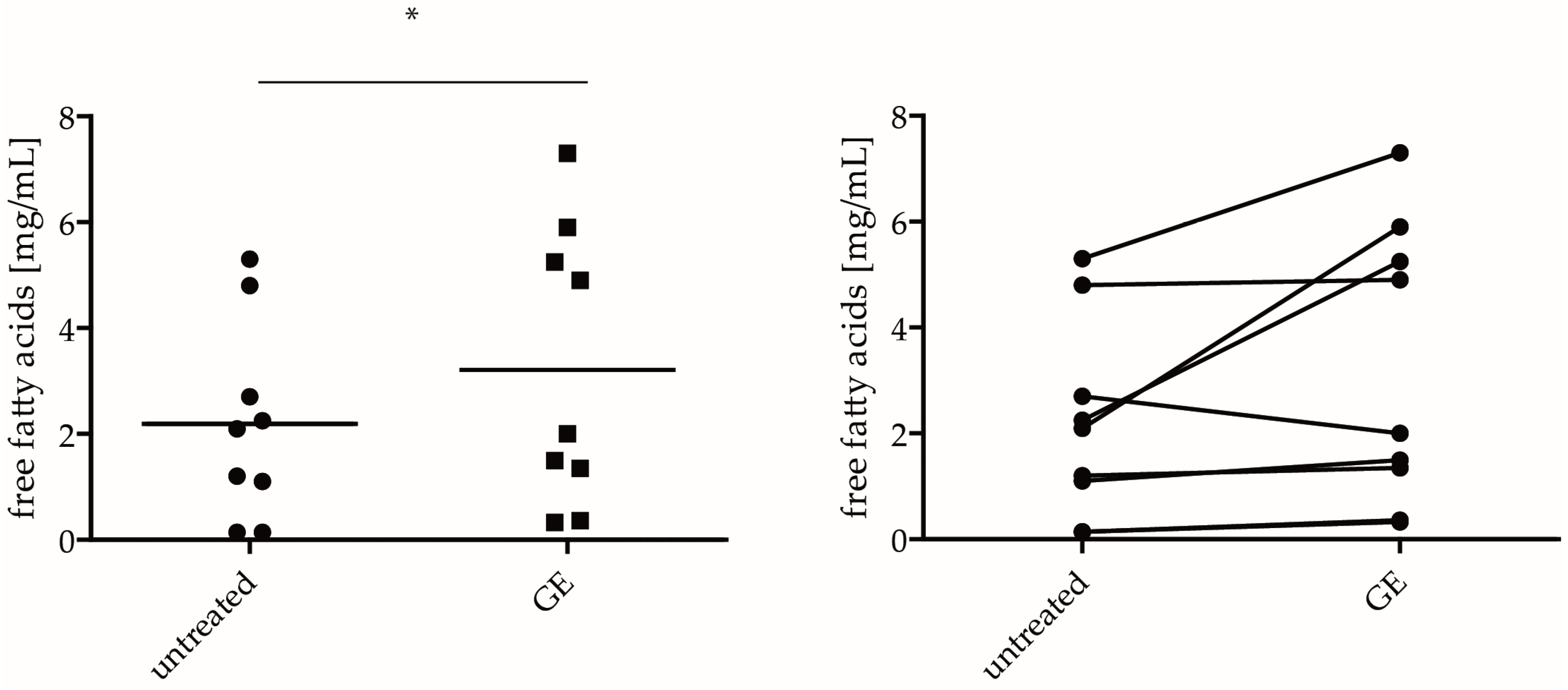
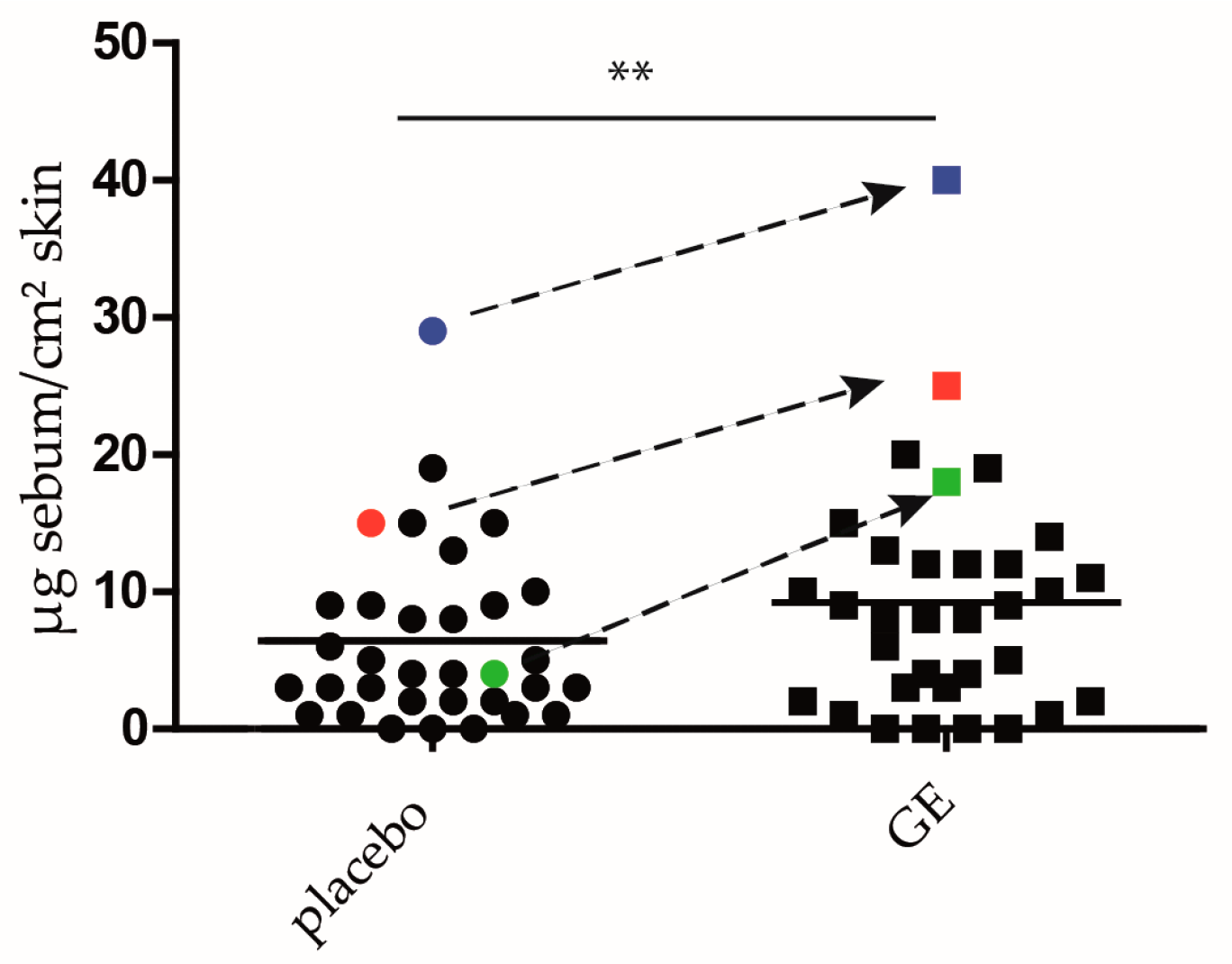
2.4. GE Enhances Lipid Production In Vivo
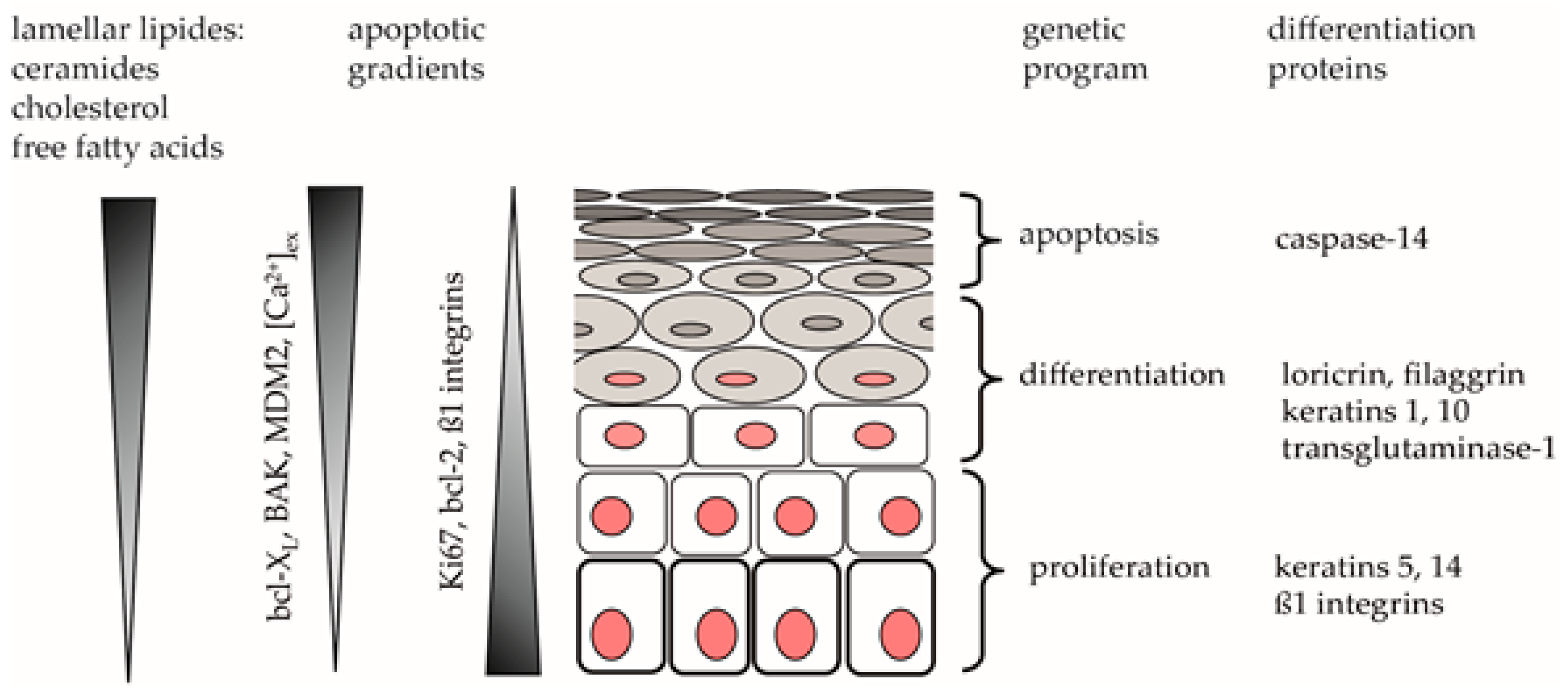
3. Discussion
4. Materials and Methods
4.1. Antibodies and Reagents
4.2. Preparation of Yellow Gentian Root High Pressure Ethanol (HPE) Extract
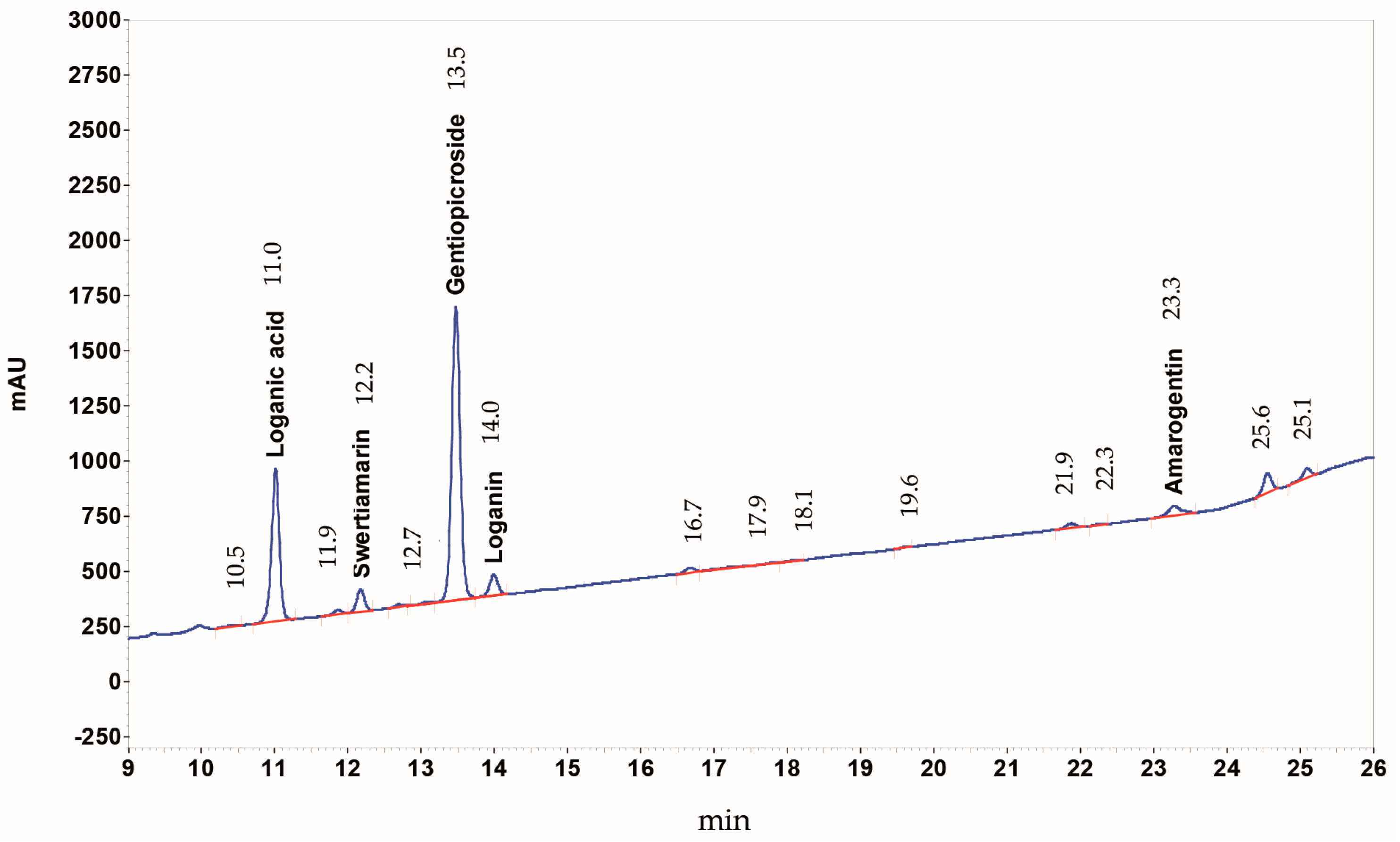
4.3. HPLC-DAD Analysis of Yellow Gentian Root HPE Extract
4.4. Cell Culture
4.5. Immunofluorescence
4.6. Cytotoxicity Assay
4.7. Enzyme Activity Assay for Sphingomyelinase
4.8. Determination of Intracellular Lipids in Keratinocytes
4.9. Isolation of Lipids from Keratinocytes
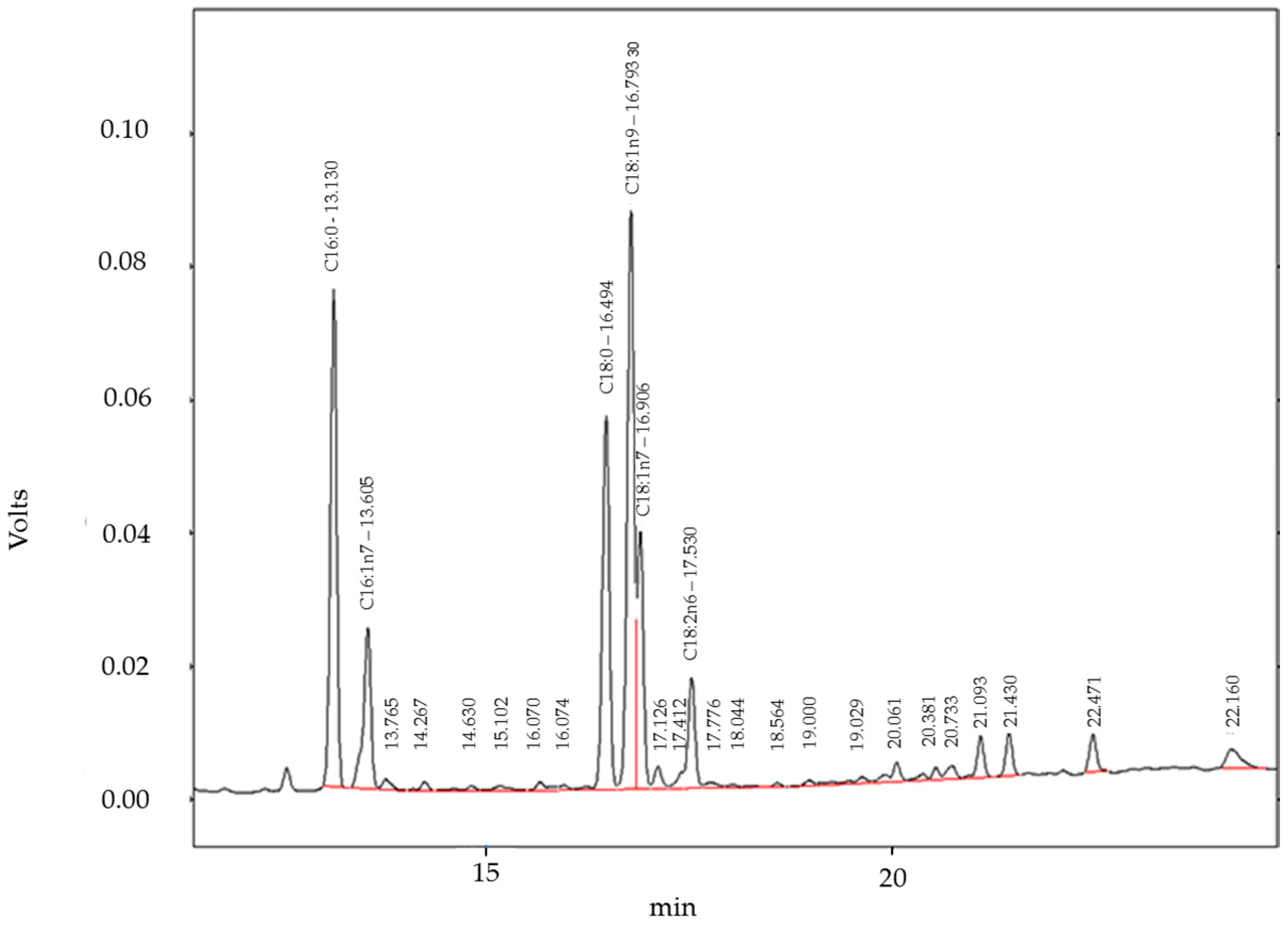
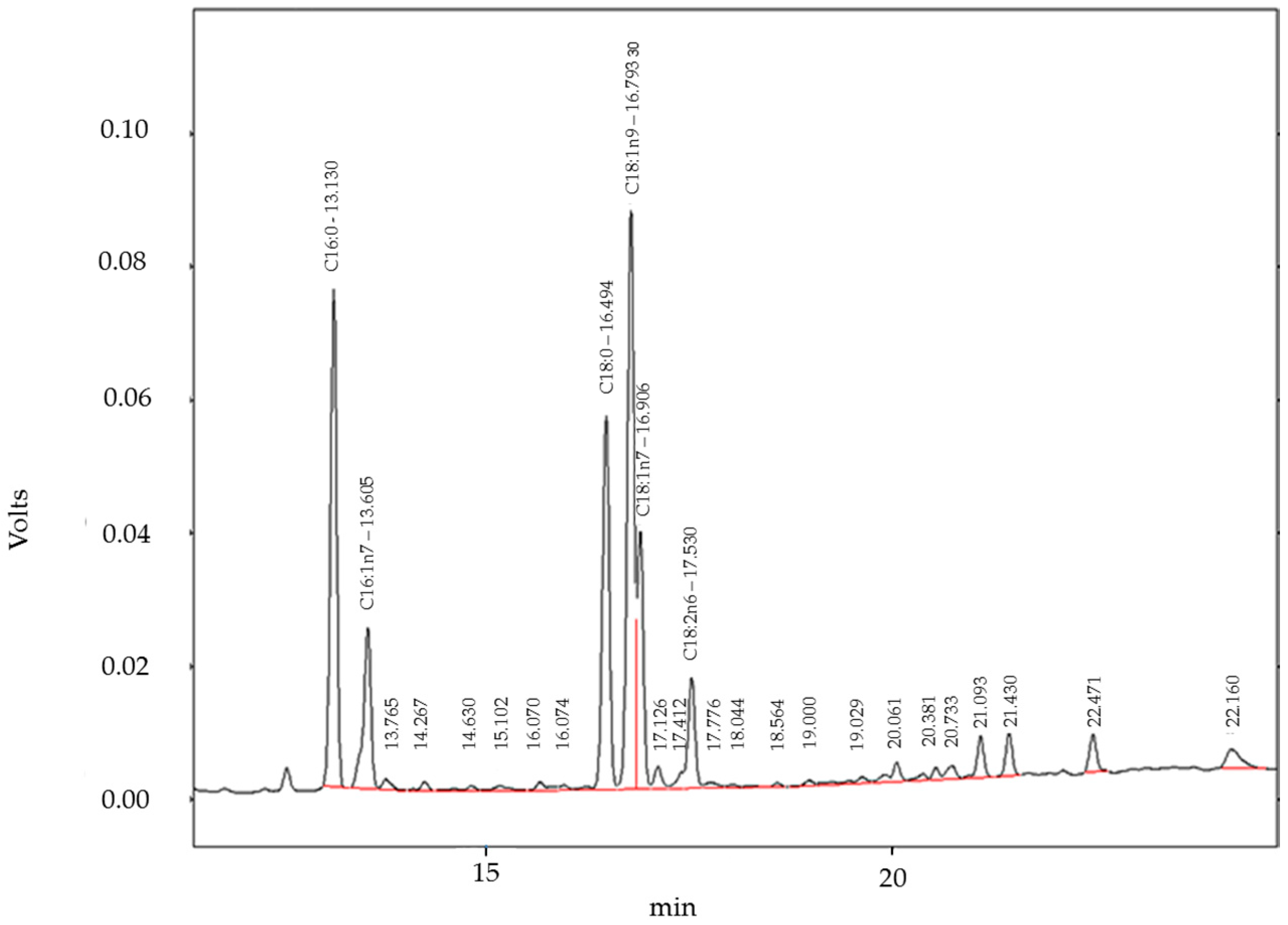
4.10. Gas Chromatography with Flame Ionization Detector (GC-FID) Analysis of Fatty Acids
4.11. Half Side Comparison
4.12. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ASMase | Shingomyelinase activity |
| BAK | Bcl-2 antagonist/killer1 |
| Ca2+ | Calcium |
| Bcl-2 | B-cell lymphoma 2 |
| CerS3 | Ceramid synthase 3 |
| GC-FID | Gaschromatography with flame ionization detector |
| INCI | International nomenclature of cosmetic ingredients |
| IL-6 | Interleukin-6 |
| HPE | High pressure ethanol |
| HPLC | High performance liquid chromatography |
| hPKs | Human primary keratinocytes |
| MDM2 | Mouse double minute 2 homolog |
| p38 MAPK | p38 mitogen-activated protein kinases |
| PGE2 | Prostaglandin E2 |
| PPARγ | Peroxisome proliferator activated receptor γ |
References
- Feingold, K.R.; Schmuth, M.; Elias, P.M. The regulation of permeability barrier homeostasis. J. Invest. Dermatol. 2007, 127, 1574–1576. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, P.; Mayer, E.P.; Fowler, S.D. Nile red: A selective fluorescent stain for intracellular lipid droplets. J. Cell Biol. 1985, 100, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R. The outer frontier: The importance of lipid metabolism in the skin. J. Lipid Res. 2009, 50, S417–S422. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y. Ceramide signaling in mammalian epidermis. Biochim. Biophys. Acta 2014, 1841, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.P.; Shin, K.-O.; Park, K.; Yun, H.J.; Mann, S.; Lee, Y.M.; Cho, Y. Vitamin C Stimulates Epidermal Ceramide Production by Regulating Its Metabolic Enzymes. Biomol. Ther. 2015, 23, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Motta, S.; Monti, M.; Sesana, S.; Mellesi, L.; Ghidoni, R.; Caputo, R. Abnormality of water barrier function in psoriasis. Role of ceramide fractions. Arch. Dermatol. 1994, 130, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G.; Abe, A.; Jin, K.; Higaki, Y.; Kawashima, M.; Hidano, A. Decreased level of ceramides in stratum corneum of atopic dermatitis: An etiologic factor in atopic dry skin? J. Investig. Dermatol. 1991, 96, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Mullen, T.D.; Hannun, Y.A.; Obeid, L.M. Ceramide synthases at the centre of sphingolipid metabolism and biology. Biochem. J. 2012, 441, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Goñi, F.M.; Alonso, A. Sphingomyelinases: Enzymology and membrane activity. FEBS Lett. 2002, 531, 38–46. [Google Scholar] [CrossRef]
- Yarosh, D.B.; Both, D.; Brown, D. Liposomal ursolic acid (merotaine) increases ceramides and collagen in human skin. Horm. Res. 2000, 54, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Kim, M.-K.; Jin, X.-J.; Oh, J.-H.; Kim, J.E.; Chung, J.H. Skin Aging and Photoaging Alter Fatty Acids Composition, Including 11,14,17-eicosatrienoic Acid, in the Epidermis of Human Skin. J. Korean Med. Sci. 2010, 25, 980–983. [Google Scholar] [CrossRef] [PubMed]
- Raj, D.; Brash, D.E.; Grossman, D. Keratinocyte apoptosis in epidermal development and disease. J. Investig. Dermatol. 2006, 126, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Wölfle, U.; Elsholz, F.A.; Kersten, A.; Haarhaus, B.; Müller, W.E.; Schempp, C.M. Expression and functional activity of the bitter taste receptors TAS2R1 and TAS2R38 in human keratinocytes. Skin Pharmacol. Physiol. 2015, 28, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem. Sens. 2010, 35, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, D.A.; Wang, W.C.; McIlmoyle, E.L.; Robinett, K.S.; Schillinger, R.M.; An, S.S.; Sham, J.S.; Liggett, S.B. Bitter taste receptors on airway smooth muscle bronchodilate by localized calcium signaling and reverse obstruction. Nat. Med. 2010, 16, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Xiong, G.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J. Clin. Investig. 2012, 122, 4145–4159. [Google Scholar] [CrossRef] [PubMed]
- Rozengurt, E.; Sternini, C. Taste receptor signaling in the mammalian gut. Curr. Opin. Pharmacol. 2007, 7, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Ponec, M.; Weerheim, A.; Kempenaar, J.; Mommaas, A.M.; Nugteren, D.H. Lipid composition of cultured human keratinocytes in relation to their differentiation. J. Lipid Res. 1988, 29, 949–961. [Google Scholar] [PubMed]
- Roudnitzky, N.; Behrens, M.; Engel, A.; Kohl, S.; Thalmann, S.; Hübner, S.; Lossow, K.; Wooding, S.P.; Meyerhof, W. Receptor Polymorphism and Genomic Structure Interact to Shape Bitter Taste Perception. PLoS Genet. 2015, 11, e1005530. [Google Scholar] [CrossRef] [PubMed]
- Rau, O.; Wurglics, M.; Dingermann, T.; Abdel-Tawab, M.; Schubert-Zsilavecz, M. Screening of herbal extracts for activation of the human peroxisome proliferator-activated receptor. Pharmazie 2006, 61, 952–956. [Google Scholar] [PubMed]
- Chon, S.-H.; Tannahill, R.; Yao, X.; Southall, M.D.; Pappas, A. Keratinocyte differentiation and upregulation of ceramide synthesis induced by an oat lipid extract via the activation of PPAR pathways. Exp. Dermatol. 2015, 24, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A.; Barak, Y.; Nagy, L.; Liao, D.; Tontonoz, P.; Evans, R.M. PPAR-gamma dependent and independent effects on macrophage-gene expression in lipid metabolism and inflammation. Nat. Med. 2001, 7, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Dobrosi, N.; Toth, B.I.; Nagy, G.; Dozsa, A.; Geczy, T.; Nagy, L.; Zouboulis, C.C.; Paus, R.; Kovacs, L.; Biro, T. Endocannabinoids enhance lipid synthesis and apoptosis of human sebocytes via cannabinoid receptor-2-mediated signaling. FASEB J. 2008, 22, 3685–3695. [Google Scholar] [CrossRef] [PubMed]
- Szatmari, I.; Töröcsik, D.; Agostini, M.; Nagy, T.; Gurnell, M.; Barta, E.; Chatterjee, K.; Nagy, L. PPARgamma regulates the function of human dendritic cells primarily by altering lipid metabolism. Blood 2007, 110, 3271–3280. [Google Scholar] [CrossRef] [PubMed]
- Gehart, H.; Kumpf, S.; Ittner, A.; Ricci, R. MAPK signalling in cellular metabolism: Stress or wellness? EMBO Rep. 2010, 11, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Holder, J.; Smith, S.; Kumar, S.; Eggo, M. Essential role of p38 mitogen-activated protein kinase in adipogenesis. In Proceedings of the 21st Joint Meeting of the British Endocrine Societies, Harrogate, UK, 8–11 April 2002. [Google Scholar]
- Cao, W.; Collins, Q.F.; Becker, T.C.; Robidoux, J.; Lupo, E.G.; Xiong, Y.; Daniel, K.W.; Floering, L.; Collins, S. p38 Mitogen-activated protein kinase plays a stimulatory role in hepatic gluconeogenesis. J. Biol. Chem. 2005, 280, 42731–42737. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A. Epidermal surface lipids. Dermatoendocrinology 2009, 1, 72–76. [Google Scholar] [CrossRef]
- Drake, D.R.; Brogden, K.A.; Dawson, D.V.; Wertz, P.W. Thematic review series: Skin lipids. Antimicrobial lipids at the skin surface. J. Lipid Res. 2008, 49, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Bonté, F.; Barré, P.; Pinguet, P.; Dusser, I.; Dumas, M.; Meybeck, A. Simarouba amara extract increases human skin keratinocyte differentiation. J. Ethnopharmacol. 1996, 53, 65–74. [Google Scholar] [CrossRef]
- Jennemann, R.; Rabionet, M.; Gorgas, K.; Epstein, S.; Dalpke, A.; Rothermel, U.; Bayerle, A.; van der Hoeven, F.; Imgrund, S.; Kirsch, J.; et al. Loss of ceramide synthase 3 causes lethal skin barrier disruption. Hum. Mol. Genet. 2012, 21, 586–608. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, Y.; Mitsutake, S.; Tsuji, K.; Kihara, A.; Igarashi, Y. Ceramide biosynthesis in keratinocyte and its role in skin function. Biochimie 2009, 91, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G. Lipid abnormalities in atopic dermatitis. J. Am. Acad. Dermatol. 2001, 45, S29–S32. [Google Scholar] [CrossRef] [PubMed]
- Betts, J.C.; Agranoff, A.B.; Nabel, G.J.; Shayman, J.A. Dissociation of endogenous cellular ceramide from NF-kappa B activation. J. Biol. Chem. 1994, 269, 8455–8458. [Google Scholar] [PubMed]
- Di Nardo, A.; Wertz, P.; Giannetti, A.; Seidenari, S. Ceramide and cholesterol composition of the skin of patients with atopic dermatitis. Acta Derm. Venereol. 1998, 78, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Geilen, C.C.; Bektas, M.; Wieder, T.; Orfanos, C.E. The vitamin D3 analogue, calcipotriol, induces sphingomyelin hydrolysis in human keratinocytes. FEBS Lett. 1996, 378, 88–92. [Google Scholar] [CrossRef]
- Lew, B.-L.; Cho, Y.; Kim, J.; Sim, W.-Y.; Kim, N.-I. Ceramides and cell signaling molecules in psoriatic epidermis: Reduced levels of ceramides, PKC-alpha, and JNK. J. Korean Med. Sci. 2006, 21, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Arikawa, J.; Ishibashi, M.; Kawashima, M.; Takagi, Y.; Ichikawa, Y.; Imokawa, G. Decreased levels of sphingosine, a natural antimicrobial agent, may be associated with vulnerability of the stratum corneum from patients with atopic dermatitis to colonization by Staphylococcus aureus. J. Investig. Dermatol. 2002, 119, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Fluhr, J.W.; Kao, J.; Jain, M.; Ahn, S.K.; Feingold, K.R.; Elias, P.M. Generation of free fatty acids from phospholipids regulates stratum corneum acidification and integrity. J. Investig. Dermatol. 2001, 117, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R. Thematic review series: Skin lipids. The role of epidermal lipids in cutaneous permeability barrier homeostasis. J. Lipid Res. 2007, 48, 2531–2546. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Mao-Qiang, M.; Taljebini, M.; Elias, P.M.; Feingold, K.R. Topical stratum corneum lipids accelerate barrier repair after tape stripping, solvent treatment and some but not all types of detergent treatment. Br. J. Dermatol. 1995, 133, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Geilen, C.C.; Wieder, T.; Orfanos, C.E. Ceramide signalling: Regulatory role in cell proliferation, differentiation and apoptosis in human epidermis. Arch. Dermatol. Res. 1997, 289, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Ballou, L.R.; Chao, C.P.; Holness, M.A.; Barker, S.C.; Raghow, R. Interleukin-1-mediated PGE2 production and sphingomyelin metabolism. Evidence for the regulation of cyclooxygenase gene expression by sphingosine and ceramide. J. Biol. Chem. 1992, 267, 20044–20050. [Google Scholar] [PubMed]
- Vieira, J.; Cardoso, C.S.; Pinto, J.; Patil, K.; Brazdil, P.; Cruz, E.; Mascarenhas, C.; Lacerda, R.; Gartner, A.; Almeida, S.; Alves, H.; Porto, G. A putative gene located at the MHC class I region around the D6S105 marker contributes to the setting of CD8+ T-lymphocyte numbers in humans. Int. J. Immunogenet. 2007, 34, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Danso, M.O.; van Drongelen, V.; Mulder, A.; van Esch, J.; Scott, H.; van Smeden, J.; El Ghalbzouri, A.; Bouwstra, J.A. TNF-α and Th2 Cytokines Induce Atopic Dermatitis–Like Features on Epidermal Differentiation Proteins and Stratum Corneum Lipids in Human Skin Equivalents. J. Investig. Dermatol. 2014, 134, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Rheinwald, J.G.; Green, H. Formation of a keratinizing epithelium in culture by a cloned cell line derived from a teratoma. Cell 1975, 6, 317–330. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | % Fatty Acids in hPKs (This Study) | % Fatty Acids in HaCaT Cells (This Study) | % Fatty Acids in Freshly Isolated Keratinocytes of Human Epidermis [18] | % Fatty Acids in Skin Models [18] |
|---|---|---|---|---|
| C16:0 | 15.5% | 10.8% | 15.8% | 22.2% |
| C16:1 w7 | 4.5% | 0.83% | 1.8% | 13.1% |
| C18:0 | 10% | 7.8% | 14.8% | 10.8% |
| C18:1 w9 | 11.8% | 10% | 16.6% (only marked as18:1) | 45.8% |
| C18:2 w6 | 11.8% | 17.5% | 23.3% (only marked as C18:2) | 1.1% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wölfle, U.; Haarhaus, B.; Seiwerth, J.; Cawelius, A.; Schwabe, K.; Quirin, K.-W.; Schempp, C.M. The Herbal Bitter Drug Gentiana lutea Modulates Lipid Synthesis in Human Keratinocytes In Vitro and In Vivo. Int. J. Mol. Sci. 2017, 18, 1814. https://doi.org/10.3390/ijms18081814
Wölfle U, Haarhaus B, Seiwerth J, Cawelius A, Schwabe K, Quirin K-W, Schempp CM. The Herbal Bitter Drug Gentiana lutea Modulates Lipid Synthesis in Human Keratinocytes In Vitro and In Vivo. International Journal of Molecular Sciences. 2017; 18(8):1814. https://doi.org/10.3390/ijms18081814
Chicago/Turabian StyleWölfle, Ute, Birgit Haarhaus, Jasmin Seiwerth, Anja Cawelius, Kay Schwabe, Karl-Werner Quirin, and Christoph M. Schempp. 2017. "The Herbal Bitter Drug Gentiana lutea Modulates Lipid Synthesis in Human Keratinocytes In Vitro and In Vivo" International Journal of Molecular Sciences 18, no. 8: 1814. https://doi.org/10.3390/ijms18081814
APA StyleWölfle, U., Haarhaus, B., Seiwerth, J., Cawelius, A., Schwabe, K., Quirin, K.-W., & Schempp, C. M. (2017). The Herbal Bitter Drug Gentiana lutea Modulates Lipid Synthesis in Human Keratinocytes In Vitro and In Vivo. International Journal of Molecular Sciences, 18(8), 1814. https://doi.org/10.3390/ijms18081814