The Effects of Artemisinin on the Cytolytic Activity of Natural Killer (NK) Cells

 ,
,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
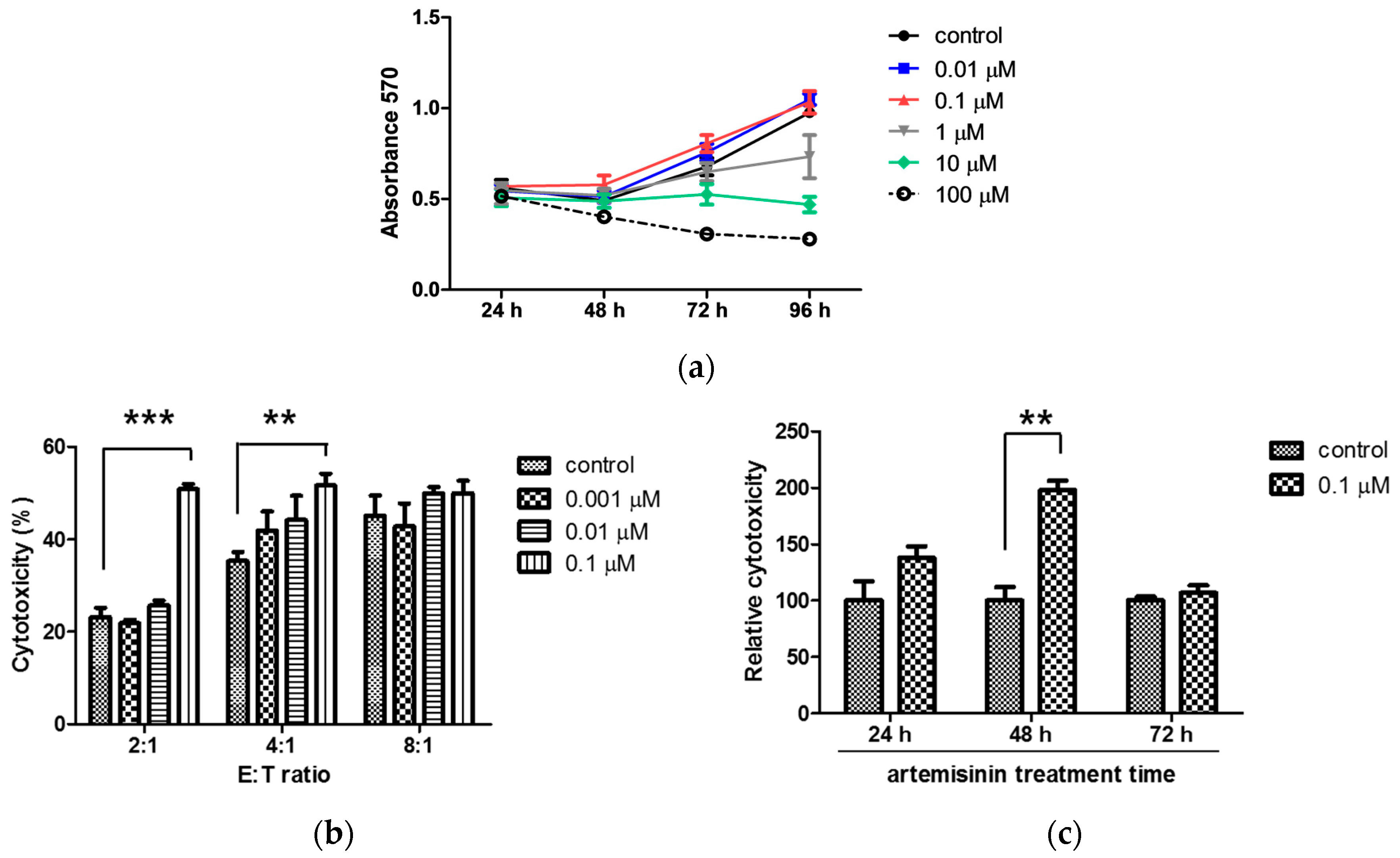
2.1. Artemisinin Enhances the Cytolytic Activity of NK-92MI Cells
2.2. Artemisinin Stimulates Granule Exocytosis of NK Cells
2.3. Artemisinin Stimulates ERK 1/2 Signaling Down-Stream of Activating Receptor
2.4. Artemisinin Increases Primary Human NK Cell Cytotoxicity
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Reagents
4.2. Cell Counting Kit (CCK)-8 Assay
4.3. NK Cytotoxicity Assay
4.4. CD107a Degranulation Assay
4.5. NKp30, NKp44, NKp46 and NKG2D Staining
4.6. Western Blotting
4.7. Isolation of Human PBLs
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Woodrow, C.J.; Haynes, R.K.; Krishna, S. Artemisinins. Postgrad. Med. J. 2005, 81, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.H.; Su, X. Artemisinin: Discovery from the Chinese herbal garden. Cell 2011, 146, 855–858. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.V.; Tekwani, B.L.; Singh, R.L.; Chauhan, V.S. Artemisinin, an endoperoxide antimalarial, disrupts the hemoglobin catabolism and heme detoxification systems in malarial parasite. J. Biol. Chem. 1999, 274, 19383–19388. [Google Scholar] [CrossRef] [PubMed]
- Dreher, D.; Junod, A.F. Role of oxygen free radicals in cancer development. Eur. J. Cancer 1996, 32, 30–38. [Google Scholar] [CrossRef]
- Tilaoui, M.; Mouse, H.A.; Jaafari, A.; Zyad, A. Differential effect of artemisinin against cancer cell lines. Nat. Prod. Bioprospect. 2014, 4, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.C.; Singh, N.P.; Sasaki, T. Development of artemisinin compounds for cancer treatment. Investig. New Drugs 2013, 31, 230–246. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Dunstan, H.; Sauerbrey, A.; Miyachi, H.; Chitambar, C.R. The anti-malarial artesunate is also active against cancer. Int. J. Oncol. 2001, 18, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Morvan, M.G.; Lanier, L.L. NK cells and cancer: You can teach innate cells new tricks. Nat. Rev. Cancer 2016, 16, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Pegram, H.J.; Andrews, D.M.; Smyth, M.J.; Darcy, P.K.; Kershaw, M.H. Activating and inhibitory receptors of natural killer cells. Immunol. Cell Biol. 2011, 89, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Smyth, M.J.; Hayakawa, Y.; Takeda, K.; Yagita, H. New aspects of natural-killer-cell surveillance and therapy of cancer. Nat. Rev. Cancer 2002, 2, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Stinchcombe, J.C.; Salio, M.; Cerundolo, V.; Pende, D.; Arico, M.; Griffiths, G.M. Centriole polarisation to the immunological synapse directs secretion from cytolytic cells of both the innate and adaptive immune systems. BMC Biol. 2011, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Stinchcombe, J.C.; Majorovits, E.; Bossi, G.; Fuller, S.; Griffiths, G.M. Centrosome polarization delivers secretory granules to the immunological synapse. Nature 2006, 443, 462–465. [Google Scholar] [CrossRef] [PubMed]
- Jahn, R.; Scheller, R.H. SNAREs—Engines for membrane fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Smyth, M.J.; Cretney, E.; Kelly, J.M.; Westwood, J.A.; Street, S.E.; Yagita, H.; Takeda, K.; van Dommelen, S.L.; Degli-Esposti, M.A.; Hayakawa, Y. Activation of NK cell cytotoxicity. Mol. Immunol. 2005, 42, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Krzewski, K.; Coligan, J.E. Human NK cell lytic granules and regulation of their exocytosis. Front. Immunol. 2012, 3, 335. [Google Scholar] [CrossRef] [PubMed]
- Alter, G.; Malenfant, J.M.; Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 2004, 294, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, T.; Shinohara, N.; Takayama, H.; Takaku, K.; Kondo, S.; Yonehara, S.; Nagai, K. Concanamycin A, a powerful tool for characterization and estimation of contribution of perforin- and Fas-based lytic pathways in cell-mediated cytotoxicity. J. Immunol. 1996, 156, 3678–3686. [Google Scholar] [PubMed]
- Graham, D.B.; Cella, M.; Giurisato, E.; Fujikawa, K.; Miletic, A.V.; Kloeppel, T.; Brim, K.; Takai, T.; Shaw, A.S.; Colonna, M.; et al. Vav1 controls DAP10-mediated natural cytotoxicity by regulating actin and microtubule dynamics. J. Immunol. 2006, 177, 2349–2355. [Google Scholar] [CrossRef] [PubMed]
- Zompi, S.; Hamerman, J.A.; Ogasawara, K.; Schweighoffer, E.; Tybulewicz, V.L.; Di Santo, J.P.; Lanier, L.L.; Colucci, F. NKG2D triggers cytotoxicity in mouse NK cells lacking DAP12 or Syk family kinases. Nat. Immunol. 2003, 4, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Ugolini, S.; Nunes, J.A. ADAPted secretion of cytokines in NK cells. Nat. Immunol. 2013, 14, 1108–1110. [Google Scholar] [CrossRef] [PubMed]
- Bhaw-Luximon, A.; Jhurry, D. Artemisinin and its derivatives in cancer therapy: Status of progress, mechanism of action, and future perspectives. Cancer Chemother. Pharmacol. 2017, 79, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, J.A.; Sundar, S.N.; Cheung, M.; Tin, A.S.; Modiano, J.; Firestone, G.L. Artemisinin blocks prostate cancer growth and cell cycle progression by disrupting Sp1 interactions with the cyclin-dependent kinase-4 (CDK4) promoter and inhibiting CDK4 gene expression. J. Biol. Chem. 2009, 284, 2203–2213. [Google Scholar] [CrossRef] [PubMed]
- Steinbruck, L.; Pereira, G.; Efferth, T. Effects of artesunate on cytokinesis and G2/M cell cycle progression of tumour cells and budding yeast. Cancer Genom. Proteom. 2010, 7, 337–346. [Google Scholar]
- Deng, Z.; Bald, I.; Illenberger, E.; Huels, M.A. Bond- and energy-selective carbon abstraction from D-ribose by hyperthermal nitrogen ions. Angew. Chem. Int. Ed. Engl. 2008, 47, 9509–9512. [Google Scholar] [CrossRef] [PubMed]
- Weifeng, T.; Feng, S.; Xiangji, L.; Changqing, S.; Zhiquan, Q.; Huazhong, Z.; Peining, Y.; Yong, Y.; Mengchao, W.; Xiaoqing, J.; et al. Artemisinin inhibits in vitro and in vivo invasion and metastasis of human hepatocellular carcinoma cells. Phytomedicine 2011, 18, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Wartenberg, M.; Wolf, S.; Budde, P.; Grunheck, F.; Acker, H.; Hescheler, J.; Wartenberg, G.; Sauer, H. The antimalaria agent artemisinin exerts antiangiogenic effects in mouse embryonic stem cell-derived embryoid bodies. Lab. Investig. 2003, 83, 1647–1655. [Google Scholar] [CrossRef] [PubMed]
- Pollard, J.W. Tumour-educated macrophages promote tumour progression and metastasis. Nat. Rev. Cancer 2004, 4, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Hu, K.; Li, S.; Zhu, J.; Gu, L.; Shen, H.; Hambly, B.D.; Bao, S.; Di, W. Dihydroartiminisin inhibits the growth and metastasis of epithelial ovarian cancer. Oncol. Rep. 2012, 27, 101–108. [Google Scholar] [PubMed]
- Guillerey, C.; Huntington, N.D.; Smyth, M.J. Targeting natural killer cells in cancer immunotherapy. Nat. Immunol. 2016, 17, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, H.; Kakuda, H.; Shimasaki, N.; Imai, C.; Ma, J.; Lockey, T.; Eldridge, P.; Leung, W.H.; Campana, D. Expansion of highly cytotoxic human natural killer cells for cancer cell therapy. Cancer Res. 2009, 69, 4010–4017. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, C.C.; Watzl, C.; Billadeau, D.D.; Leibson, P.J.; Burshtyn, D.N.; Long, E.O. Vav1 dephosphorylation by the tyrosine phosphatase SHP-1 as a mechanism for inhibition of cellular cytotoxicity. Mol. Cell Biol. 2003, 23, 6291–6299. [Google Scholar] [CrossRef] [PubMed]
- Colucci, F.; Rosmaraki, E.; Bregenholt, S.; Samson, S.I.; di Bartolo, V.; Turner, M.; Vanes, L.; Tybulewicz, V.; Di Santo, J.P. Functional dichotomy in natural killer cell signaling: Vav1-dependent and -independent mechanisms. J. Exp. Med. 2001, 193, 1413–1424. [Google Scholar] [CrossRef] [PubMed]
- Cote, M.; Menager, M.M.; Burgess, A.; Mahlaoui, N.; Picard, C.; Schaffner, C.; Al-Manjomi, F.; Al-Harbi, M.; Alangari, A.; Le Deist, F.; et al. Munc18-2 deficiency causes familial hemophagocytic lymphohistiocytosis type 5 and impairs cytotoxic granule exocytosis in patient NK cells. J. Clin. Investig. 2009, 119, 3765–3773. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.S.; Friedmann, K.S.; Knorck, A.; Hoxha, C.; Leidinger, P.; Backes, C.; Meese, E.; Keller, A.; Rettig, J.; Hoth, M.; et al. Syntaxin 8 is required for efficient lytic granule trafficking in cytotoxic T lymphocytes. Biochim. Biophys. Acta 2016, 1863, 1653–1664. [Google Scholar] [CrossRef] [PubMed]
- Dabrazhynetskaya, A.; Ma, J.; Guerreiro-Cacais, A.O.; Arany, Z.; Rudd, E.; Henter, J.I.; Karre, K.; Levitskaya, J.; Levitsky, V. Syntaxin 11 marks a distinct intracellular compartment recruited to the immunological synapse of NK cells to colocalize with cytotoxic granules. J. Cell Mol. Med. 2012, 16, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Hellewell, A.L.; Foresti, O.; Gover, N.; Porter, M.Y.; Hewitt, E.W. Analysis of familial hemophagocytic lymphohistiocytosis type 4 (FHL-4) mutant proteins reveals that S-acylation is required for the function of syntaxin 11 in natural killer cells. PLoS ONE 2014, 9, e98900. [Google Scholar] [CrossRef] [PubMed]
- Arneson, L.N.; Brickshawana, A.; Segovis, C.M.; Schoon, R.A.; Dick, C.J.; Leibson, P.J. Cutting edge: Syntaxin 11 regulates lymphocyte-mediated secretion and cytotoxicity. J. Immunol. 2007, 179, 3397–3401. [Google Scholar] [CrossRef] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Houh, Y.K.; Kim, K.E.; Park, S.; Hur, D.Y.; Kim, S.; Kim, D.; Bang, S.I.; Yang, Y.; Park, H.J.; Cho, D. The Effects of Artemisinin on the Cytolytic Activity of Natural Killer (NK) Cells. Int. J. Mol. Sci. 2017, 18, 1600. https://doi.org/10.3390/ijms18071600
Houh YK, Kim KE, Park S, Hur DY, Kim S, Kim D, Bang SI, Yang Y, Park HJ, Cho D. The Effects of Artemisinin on the Cytolytic Activity of Natural Killer (NK) Cells. International Journal of Molecular Sciences. 2017; 18(7):1600. https://doi.org/10.3390/ijms18071600
Chicago/Turabian StyleHouh, Youn Kyung, Kyung Eun Kim, Sunyoung Park, Dae Young Hur, Seonghan Kim, Daejin Kim, Sa Ik Bang, Yoolhee Yang, Hyun Jeong Park, and Daeho Cho. 2017. "The Effects of Artemisinin on the Cytolytic Activity of Natural Killer (NK) Cells" International Journal of Molecular Sciences 18, no. 7: 1600. https://doi.org/10.3390/ijms18071600
APA StyleHouh, Y. K., Kim, K. E., Park, S., Hur, D. Y., Kim, S., Kim, D., Bang, S. I., Yang, Y., Park, H. J., & Cho, D. (2017). The Effects of Artemisinin on the Cytolytic Activity of Natural Killer (NK) Cells. International Journal of Molecular Sciences, 18(7), 1600. https://doi.org/10.3390/ijms18071600