Enriching Diet with n-3 PUFAs to Help Prevent Cardiovascular Diseases in Healthy Adults: Results from Clinical Trials



Abstract
:
1. Introduction
2. Results
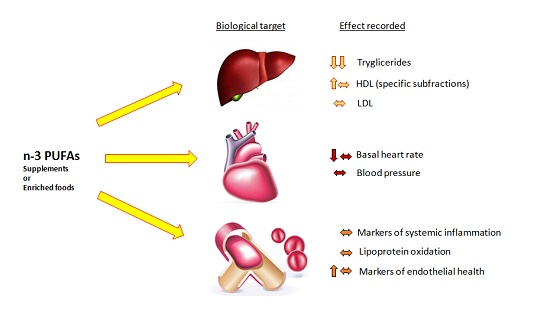
2.1. Effects on Lipid Profile
2.2. Effects on Cardiac Function and Blood Pressure
2.3. Effects on Thrombosis, Vascular Health and Inflammation
3. Discussion
4. Materials and Methods
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Global Health Risks. Avaliale online: http://www.who.int/healthinfo/global_burden_disease/GlobalHealthRisks_report_full.pdf (accessed on 31 May 2007).
- Galassi, A.; Reynolds, K.; He, J. Metabolic syndrome and risk of cardiovascular disease: A meta-analysis. Am. J. Med. 2006, 119, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Jump, D.B.; Depner, C.M.; Tripathy, S. n-3 fatty acid supplementation and cardiovascular disease. J. Lipid Res. 2012, 53, 2525–2545. [Google Scholar] [CrossRef] [PubMed]
- Reiner, Ž.; Catapano, A.L.; de Backer, G.; Graham, I.; Taskinen, M.R.; Wiklund, O.; Agewall, S.; Alegria, E.; Chapman, M.J.; Durrington, P.; et al. ESC/EAS Guidelines for the management of dyslipidaemias The Task Force for the management of dyslipidaemias of the European Society of Cardiology (ESC) and the European Atherosclerosis Society (EAS). Eur. Heart J. 2011, 32, 1769–1818. [Google Scholar]
- Watanabe, Y.; Tatsuno, I. n-3 polyunsaturated fatty acids for cardiovascular diseases: Present, past and future. Exp. Rev. Clin. Pharmacol. 2017, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. The role of marine n-3 fatty acids in inflammatory processes, atherosclerosis and plaque stability. Mol. Nutr. Food Res. 2012, 56, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Tetens, I. Opinion of the Scientific Panel on Dietetic products, nutrition and allergies [NDA] related to nutrition claims concerning n-3 fatty acids, monounsaturated fat, polyunsaturated fat and unsaturated fat. EFSA 2005. [Google Scholar] [CrossRef]
- DeFilippis, A.P.; Sperling, L.S. Understanding n-3’s. Am. Heart J. 2006, 151, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Dewailly, E.; Blanchet, C.; Lemieux, S.; Sauvé, L.; Gingras, S.; Ayotte, P.; Holub, B.J. n-3 Fatty acids and cardiovascular disease risk factors among the Inuit of Nunavik. Am. J. Clin. Nutr. 2001, 74, 464–473. [Google Scholar] [PubMed]
- Rizos, E.C.; Ntzani, E.E.; Bika, E.; Kostapanos, M.S.; Elisaf, M.S. Association between n-3 fatty acid supplementation and risk of major cardiovascular disease events: A systematic review and meta-analysis. JAMA 2012, 308, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Maehre, H.K.; Jensen, I.J.; Elvevoll, E.O.; Eilertsen, K.E. ω-3 Fatty Acids and Cardiovascular Diseases: Effects, Mechanisms and Dietary Relevance. Int. J. Mol. Sci. 2015, 16, 22636–22661. [Google Scholar] [CrossRef] [PubMed]
- Bowen, K.J.; Harris, W.S.; Kris-Etherton, P.M. n-3 Fatty Acids and Cardiovascular Disease: Are There Benefits? Curr. Treat. Options Cardiovasc. Med. 2016, 18, 69. [Google Scholar] [CrossRef] [PubMed]
- Walz, C.P.; Barry, A.R.; Koshman, S.L. n-3 polyunsaturated fatty acid supplementation in the prevention of cardiovascular disease. Can. Pharm. J. 2016, 149, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Maurizi, G.; Della Guardia, L.; Maurizi, A.; Poloni, A. Adipocytes properties and crosstalk with immune system in obesity-related inflammation. J. Cell Physiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Endo, J.; Arita, M. Cardioprotective mechanism of n-3 polyunsaturated fatty acids. J. Cardiol. 2016, 67, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Antollini, S.S.; Barrantes, F.J. Fatty Acid Regulation of Voltage- and Ligand-Gated Ion Channel Function. Front. Physiol. 2016, 7, 573. [Google Scholar] [CrossRef] [PubMed]
- Christou, G.A.; Christou, K.A.; Korantzopoulos, P.; Rizos, E.C.; Nikas, D.N.; Goudevenos, J.A. The Current Role of n-3 Fatty Acids in the Management of Atrial Fibrillation. Int. J. Mol. Sci. 2015, 16, 22870–22887. [Google Scholar] [CrossRef] [PubMed]
- Calò, L.; Martino, A.; Tota, C. The anti-arrhythmic effects of n-3 PUFAs. Int. J. Cardiol. 2013, 170, S21–S27. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, R.G.; James, M.J.; Gibson, R.A.; Edwards, J.R.; Stubberfield, J.; Stuklis, R.; Roberts-Thomson, K.; Young, G.D.; Cleland, L.G. Effects of fish-oil supplementation on myocardial fatty acids in humans. Am. J. Clin. Nutr. 2007, 85, 1222–1228. [Google Scholar] [PubMed]
- Liu, T.; Korantzopoulos, P.; Shehata, M.; Wang, X.; Kaul, S. Prevention of atrial fibrillation with n-3 fatty acids: A meta-analysis of randomised clinical trials. Heart 2011, 97, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Einvik, G.; Klemsdal, T.O.; Sandvik, L.; Hjerkinn, E.M. A randomized clinical trial on n-3 polyunsaturated fatty acids supplementation and all-cause mortality in elderly men at high cardiovascular risk. Eur. J. Cardiovasc. Prev. Rehabil. 2010, 17, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Albert, C.M.; Hennekens, C.H.; O’Donnell, C.J.; Ajani, U.A.; Carey, V.J.; Willett, W.C.; Ruskin, J.N.; Manson, J.E. Fish consumption and risk of sudden cardiac death. JAMA 1998, 279, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Oomen, C.M.; Feskens, E.J.; Räsänen, L.; Fidanza, F.; Nissinen, A.M.; Menotti, A.; Kok, F.J.; Kromhout, D. Fish consumption and coronary heart disease mortality in Finland, Italy, and The Netherlands. Am. J. Epidemiol. 2000, 151, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Lemaitre, R.N.; Kuller, L.H.; Burke, G.L.; Tracy, R.P.; Siscovick, D.S. Cardiac benefits of fish consumption may depend on the type of fish meal consumed: The Cardiovascular Health Study. Circulation 2003, 107, 1372–1377. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J. Fish consumption, fish oil, n-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, P.; Leach, C.; Ah-Sing, E.E.; Hussain, N.; Miller, G.J.; Millward, D.J.; Griffin, B.A. Influence of alpha-linolenic acid and fish-oil on markers of cardiovascular risk in subjects with an atherogenic lipoprotein phenotype. Atherosclerosis 2005, 181, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Tholstrup, T.; Hellgren, L.I.; Petersen, M.; Basu, S.; Straarup, E.M.; Schnohr, P.; Sandström, B. A solid dietary fat containing fish oil redistributes lipoprotein subclasses without increasing oxidative stress in men. J. Nutr. 2004, 134, 1051–1057. [Google Scholar] [PubMed]
- Hlais, S.; El-Bistami, D.; El Rahi, B.; Mattar, M.A.; Obeid, O.A. Combined fish oil and high oleic sunflower oil supplements neutralize their individual effects on the lipid profile of healthy men. Lipids 2013, 48, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Berge, R.K.; Ramsvik, M.S.; Bohov, P.; Svardal, A.; Nordrehaug, J.E.; Rostrup, E.; Bruheim, I.; Bjørndal, B. Krill oil reduces plasma triacylglycerol level and improves related lipoprotein particle concentration, fatty acid composition and redox status in healthy young adults—A pilot study. Lipids Health Dis. 2015, 14, 163. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, T.; Yoko Noda, Y.; Yuko Ohno, Y.; Sugiyama, H.; Oe, H.; Nakamura, K.; Kohno, K.; Ito, H. n-3 fatty acids improve postprandial lipemia and associated endothelial dysfunction in healthy individuals—A randomized cross-over trial. Biomed. Pharmacother. 2014, 68, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Abate, N. Role of subcutaneous adipose tissue in the pathogenesis of insulin resistance. J. Obes. 2013, 2013, 489187. [Google Scholar] [CrossRef] [PubMed]
- Buckley, J.D.; Burgess, S.; Murphy, K.J.; Howe, P.R. DHA-rich fish oil lowers heart rate during submaximal exercise in elite Australian Rules footballers. J. Sci. Med. Sport 2009, 12, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Singhal, A.; Lanigan, J.; Storry, C.; Low, S.; Birbara, T.; Lucas, A.; Deanfield, J. Docosahexaenoic acid supplementation, vascular function and risk factors for cardiovascular disease: A randomized controlled trial in young adults. J. Am. Heart Assoc. 2013, 2, e000283. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.D.; Holub, B.J. Differential eicosapentaenoic acid elevations and altered cardiovascular disease risk factor responses after supplementation with docosahexaenoic acid in postmenopausal women receiving and not receiving hormone replacement therapy. Am. J. Clin. Nutr. 2004, 79, 765–773. [Google Scholar] [PubMed]
- Cazzola, R.; Russo-Volpe, S.; Miles, E.A.; Rees, D.; Banerjee, T.; Roynette, C.E.; Wells, S.J.; Goua, M.; Wahle, K.W.; Calder, P.C.; et al. Age- and dose-dependent effects of an eicosapentaenoic acid-rich oil on cardiovascular risk factors in healthy male subjects. Atherosclerosis 2007, 193, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Superko, H.R.; Pendyala, L.; Williams, P.T.; Momary, K.M.; King, S.B.; Garrett, B.C. High-density lipoprotein subclasses and their relationship to cardiovascular disease. J. Clin. Lipidol. 2012, 6, 496–523. [Google Scholar] [CrossRef] [PubMed]
- Burillo, E.; Martín-Fuentes, P.; Mateo-Gallego, R.; Baila-Rueda, L.; Cenarro, A.; Ros, E.; Civeira, F. Omega-3 fatty acids and HDL. How do they work in the prevention of cardiovascular disease? Curr. Vasc. Pharmacol. 2012, 10, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Mensink, R.P.; Zock, P.L.; Kester, A.D.; Katan, M.B. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: A meta-analysis of 60 controlled trials. Am. J. Clin. Nutr. 2003, 77, 1146–1155. [Google Scholar] [PubMed]
- Wooten, J.S.; Biggerstaff, K.D.; Ben-Ezra, V. Responses of LDL and HDL particle size and distribution to omega-3 fatty acid supplementation and aerobic exercise. J. Appl. Physiol. 2009, 107, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.T.; Krauss, R.M.; Vranizan, K.M.; Stefanick, M.L.; Wood, P.D.; Lindgren, F.T. Associations of lipoproteins and apolipoproteins with gradient gel electrophoresis estimates of high density lipoprotein subfractions in men and women. Arterioscler. Thromb. 1992, 12, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Sioen, I.; Hacquebard, M.; Hick, G.; Maindiaux, V.; Larondelle, Y.; Carpentier, Y.A.; de Henauw, S. Effect of ALA-enriched food supply on cardiovascular risk factors in males. Lipids 2009, 44, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Nestel, P.J. Fish oil and cardiovascular disease: Lipids and arterial function. Am. J. Clin. Nutr. 2000, 71, 228S–231S. [Google Scholar] [PubMed]
- Mori, T.A.; Burke, V.; Puddey, I.B.; Watts, G.F.; O’Neal, D.N.; Best, J.D.; Beilin, L.J. Purified eicosapentaenoic and docosahexaenoic acids have differential effects on serum lipids and lipoproteins, LDL particle size, glucose, and insulin in mildly hyperlipidemic men. Am. J. Clin. Nutr. 2000, 71, 1085–1094. [Google Scholar] [PubMed]
- Suzukawa, M.; Abbey, M.; Howe, P.R.; Nestel, P.J. Effects of fish oil fatty acids on low density lipoprotein size, oxidizability, and uptake by macrophages. J. Lipid Res. 1995, 36, 473–484. [Google Scholar] [PubMed]
- Lamantia, V.; Sniderman, A.; Faraj, M. Nutritional management of hyperapoB. Nutr. Res. Rev. 2016, 29, 202–233. [Google Scholar] [CrossRef] [PubMed]
- Kaul, N.; Kreml, R.; Austria, J.A.; Richard, M.N.; Edel, A.L.; Dibrov, E.; Hirono, S.; Zettler, M.E.; Pierce, G.N. A comparison of fish oil, flaxseed oil and hempseed oil supplementation on selected parameters of cardiovascular health in healthy volunteers. J. Am. Coll. Nutr. 2008, 27, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Kirkhus, B.; Lamglait, A.; Eilertsen, K.E.; Falch, E.; Haider, T.; Vik, H.; Hoem, N.; Hagve, T.A.; Basu, S.; Olsen, E.; et al. Effects of similar intakes of marine n-3 fatty acids from enriched food products and fish oil on cardiovascular risk markers in healthy human subjects. Br. J. Nutr. 2012, 107, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Barceló-Coblijn, G.; Murphy, E.J.; Othman, R.; Moghadasian, M.H.; Kashour, T.; Friel, J.K. Flaxseed oil and fish-oil capsule consumption alters human red blood cell n-3 fatty acid composition: A multiple-dosing trial comparing 2 sources of n-3 fatty acid. Am. J. Clin. Nutr. 2008, 88, 801–809. [Google Scholar] [PubMed]
- Grimsgaard, S.; Bønaa, K.H.; Hansen, J.B.; Myhre, E.S. Effects of highly purified eicosapentaenoic acid and docosahexaenoic acid on hemodynamics in humans. Am. J. Clin. Nutr. 1998, 68, 52–59. [Google Scholar] [PubMed]
- Shah, A.P.; Ichiuji, A.M.; Han, J.K.; Traina, M.; El-Bialy, A.; Kamal Meymandi, S.; Yvonne Wachsner, R. Cardiovascular and endothelial effects of fish oil supplementation in healthy volunteers. J. Cardiovasc. Pharmacol. Ther. 2007, 12, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Delodder, F.; Tappy, L.; Liaudet, L.; Schneiter, P.; Perrudet, C.; Berger, M.M. Incorporation and washout of n-3 PUFA after high dose intravenous and oral supplementation in healthy volunteers. Clin. Nutr. 2015, 34, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Holguin, F.; Téllez-Rojo, M.M.; Lazo, M.; Mannino, D.; Schwartz, J.; Hernández, M.; Romieu, I. Cardiac autonomic changes associated with fish oil vs soy oil supplementation in the elderly. Chest 2005, 127, 1102–1107. [Google Scholar] [CrossRef]
- Geelen, A.; Zock, P.L.; Swenne, C.A.; Brouwer, I.A.; Schouten, E.G.; Katan, M.B. Effect of n-3 fatty acids on heart rate variability and baroreflex sensitivity in middle-aged subjects. Am. Heart J. 2003, 146, E4. [Google Scholar] [CrossRef]
- Nilsson, A.; Radeborg, K.; Salo, I.; Björck, I. Effects of supplementation with n-3 polyunsaturated fatty acids on cognitive performance and cardiometabolic risk markers in healthy 51 to 72 years old subjects: A randomized controlled cross-over study. Nutr. J. 2012, 11, 99. [Google Scholar] [CrossRef] [PubMed]
- Theobald, H.E.; Goodall, A.H.; Sattar, N.; Talbot, D.C.; Chowienczyk, P.J.; Sanders, T.A. Low-dose docosahexaenoic acid lowers diastolic blood pressure in middle-aged men and women. J. Nutr. 2007, 137, 973–978. [Google Scholar] [PubMed]
- Macartney, M.J.; Hingley, L.; Brown, M.A.; Peoples, G.E.; McLennan, P.L. Intrinsic heart rate recovery after dynamic exercise is improved with an increased n-3 index in healthy males. Br. J. Nutr. 2014, 112, 1984–1992. [Google Scholar] [CrossRef] [PubMed]
- Phang, M.; Lincz, L.F.; Garg, M.L. Eicosapentaenoic and docosahexaenoic acid supplementations reduce platelet aggregation and hemostatic markers differentially in men and women. J. Nutr. 2013, 143, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Conquer, J.A.; Cheryk, L.A.; Chan, E.; Gentry, P.A.; Holub, B.J. Effect of supplementation with dietary seal oil on selected cardiovascular risk factors and hemostatic variables in healthy male subjects. Thromb. Res. 1999, 96, 239–250. [Google Scholar] [CrossRef]
- Leigh-Firbank, E.C.; Minihane, A.M.; Leake, D.S.; Wright, J.W.; Murphy, M.C.; Griffin, B.A.; Williams, C.M. Eicosapentaenoic acid and docosahexaenoic acid from fish oils: Differential associations with lipid responses. Br. J. Nutr. 2002, 87, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Turini, M.E.; Crozier, G.L.; Donnet-Hughes, A.; Richelle, M.A. Short-term fish oil supplementation improved innate immunity, but increased ex vivo oxidation of LDL in man—A pilot study. Eur. J. Nutr. 2001, 40, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Mesa, M.D.; Buckley, R.; Minihane, A.M.; Yaqoob, P. Effects of oils rich in eicosapentaenoic and docosahexaenoic acids on the oxidizability and thrombogenicity of low-density lipoprotein. Atherosclerosis 2004, 175, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.A.; Thies, F.; Wallace, F.A.; Powell, J.R.; Hurst, T.L.; Newsholme, E.A.; Calder, P.C. Influence of age and dietary fish oil on plasma soluble adhesion molecule concentrations. Clin. Sci. 2001, 100, 91–100. [Google Scholar] [CrossRef] [PubMed]
- De Caterina, R.; Basta, G.; Lazzerini, G.; Dell’Omo, G.; Petrucci, R.; Morale, M.; Carmassi, F.; Pedrinelli, R. Soluble vascular cell adhesion molecule-1 as a biohumoral correlate of atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 2646–2654. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lu, N.; Chen, D.; Meng, L.; Zheng, Y.; Hui, R. Effects of n-3 PUFA supplementation on plasma soluble adhesion molecules: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2012, 95, 972–980. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.J.; Morel-Kopp, M.C.; Tofler, G.H.; Ward, C.M. The effect of n-3 polyunsaturated fatty acids on fibrin and thrombin generation in healthy subjects and subjects with cardiovascular disease. Semin. Thromb. Hemost. 2015, 41, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Guillot, N.; Caillet, E.; Laville, M.; Calzada, C.; Lagarde, M.; Véricel, E. Increasing intakes of the long-chain omega-3 docosahexaenoic acid: Effects on platelet functions and redox status in healthy men. FASEB J. 2009, 23, 2909–2916. [Google Scholar] [CrossRef] [PubMed]
- Colussi, G.; Catena, C.; Novello, M.; Bertin, N.; Sechi, L.A. Impact of n-3 polyunsaturated fatty acids on vascular function and blood pressure: Relevance for cardiovascular outcomes. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Elherik, K.; Bolton-Smith, C.; Barr, R.; Hill, A.; Murrie, I.; Belch, J.J. The effects of dietary fatty acid supplementation on endothelial function and vascular tone in healthy subjects. Cardiovasc. Res. 2003, 59, 955–962. [Google Scholar] [CrossRef]
- Fahs, C.A.; Yan, H.; Ranadive, S.; Rossow, L.M.; Agiovlasitis, S.; Wilund, K.R.; Fernhall, B. The effect of acute fish-oil supplementation on endothelial function and arterial stiffness following a high-fat meal. Appl. Physiol. Nutr. Metab. 2010, 35, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Fisher, E.; Lake, E.; McLeod, R.S. Apolipoprotein B100 quality control and the regulation of hepatic very low density lipoprotein secretion. J. Biomed. Res. 2014, 28, 178–193. [Google Scholar] [PubMed]
- Flock, M.R.; Skulas-Ray, A.C.; Harris, W.S.; Gaugler, T.L.; Fleming, J.A.; Kris-Etherton, P.M. Effects of supplemental long-chain n-3 fatty acids and erythrocyte membrane fatty acid content on circulating inflammatory markers in a randomized controlled trial of healthy adults. Prostaglandins Leukot. Essent. Fatty Acids 2014, 91, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Falsey, A.R.; Walsh, E.E.; Francis, C.W.; Looney, R.J.; Kolassa, J.E.; Hall, W.J.; Abraham, G.N. Response of C-reactive protein and serum amyloid A to influenza A infection in older adults. J. Infect. Dis. 2001, 183, 995–999. [Google Scholar] [CrossRef] [PubMed]
- Kasapis, C.; Thompson, P.D. The effects of physical activity on serum C-reactive protein and inflammatory markers: A systematic review. J. Am. Coll. Cardiol. 2005, 45, 1563–1569. [Google Scholar] [CrossRef] [PubMed]
- Steptoe, A.; Hamer, M.; Chida, Y. The effects of acute psychological stress on circulating inflammatory factors in humans: A review and meta-analysis. Brain Behav. Immun. 2007, 21, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Omega-3 Supplements: In Depth. Available online: https://nccih.nih.gov/health/omega3/introduction.htm (accessed on 31 May 2007).
- Decsi, T.; Kennedy, K. Sex-specific differences in essential fatty acid metabolism. Am. J. Clin. Nutr. 2011, 94, 1914S–1919S. [Google Scholar] [CrossRef] [PubMed]
- Flynn, M.A. Empowering people to be healthier: Public health nutrition through the Ottawa Charter. Proc. Nutr. Soc. 2015, 74, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Craig, W.J.; Mangels, A.R.; American Dietetic Association. Position of the American Dietetic Association: Vegetarian diets. J. Am. Diet. Assoc. 2009, 109, 1266–1282. [Google Scholar] [PubMed]

{kind=link}
| Effects on Lipid Profile | ||||||
|---|---|---|---|---|---|---|
| Study | Type | Subjects | Age | Duration | Supplementation | Effect |
| Hlais et al. (2013) [28] | RCT | 98 | 18–35 | 12 weeks | Fish oil (2 g/day) vs. n-9 rich oil (8 g/day) | ↓ TCG |
| Berge et al. (2015) [29] | RCT | 17 | 18–36 | 28 days | 832.5 mg/day DHA + EPA | ↓ TCG, VLDL, chylomicrons ↑ LDL ↑ large LDL ⇔ HDL |
| Miyoshi et al. (2014) [30] | RCrT | 10/5 | 20–85 | 4 weeks | 1.9g DHA + 1.5g EPA/day | ↓ Postprandial TGC elevation, VLDL, TGC in chylomicrons ⇔ LDL |
| Buckley et al. (2009) [32] | RCT | 25 | >18 | 5 weeks | DHA rich fish oil | ↓ TCG |
| Singhal et al. (2013) [33] | RCT | 328 | 18–37 | 16 weeks | 1.6 g DHA/day vs. placebo | ↓ TCG, ↓ VLDL |
| Stark et al. (2004) [34] | RCT | 14 | 45–70 | 28 days | 2.8 g DHA vs. placebo | ↓ TGC, ↑ HDL, ↓ TGC/HDL |
| Cazzola et al. (2007) [35] | RCT | 93 62 | 18–42 53–70 | 12 weeks | 1.35/2.7/4 g/day EPA | ↓ TCG ⇔ LDL ⇔ HDL |
| Tholstrup et al. (2004) [27] | RCT | 16 | 35–75 | 3 weeks | Dietary replacement 4 g/day PUFA-rich oil | ↓ TCG, VLDL, IDL ↑ LDL2b APOB ⇔ HDL ↑ HDL2b |
| Wooten et al. (2009) [39] | IT | 11 | 19–47 | 42 days | 2.45 g/day EPA 1.61 g/day DHA | ⇔ HDL and LDL Shifting from HDL subclass type 3 to type 2 |
| Sioen et al. (2009) [41] | IT | 59 | 22–65 | 12 weeks | ALA 5g + 1.5 of other PUFAs/day | ↑ HDL |
| Kaul et al. (2008) [46] | RCT | 86 | >18 | 12 weeks | 2 g/day fish oil vs. placebo/flaxseed/hempseed oil | ⇔ Total cholesterol col ⇔ LDL ⇔ HDL ⇔ TCG |
| Kirkhus et al. (2012) [47] | RCT | 159 | 18–70 | 7 weeks | 1 g EPA and DHA/day vs. placebo | ⇔ Total cholesterol ⇔ LDL ⇔ HDL ⇔ TCG |
| Nilsson et al. (2012) [54] | RCT | 40 | Middle-aged to elderly | 5 weeks | 3 g/day fish oil vs. placebo | ↓TGC |
| Barceló-Coblijn et al. (2008) [48] | RCT | 62 | >40 | 12 weeks | 1.2 g/ 2.4 g/ 3.6 g flaxseed oil/day vs. 0.6 g or 1.2 g fish oil/day vs. placebo | ⇔ Total cholesterol ⇔ HDL ⇔ TCG |
| Cardiac Function and Blood Pressure | ||||||
| Grimsgaard et al. (1998) [49] | RCT | 224 | 36–56 | 4 g/day DHA + EPA vs. 4 g/day corn oil | ↓ HR ↑ ventricular diastolic capacity, ⇔ systolic BP ⇔ diastolic BP | |
| Shah et al. (2007) [50] | RCT | 26 | 26–36 | 14 days | 1 g/day fish oil vs. 1 g/day corn oil | ↓ resting HR ⇔ systolic BP ⇔ diastolic BP |
| Stark et al. (2004) [34] | RCT | 14 | 45–70 | 28 days | 2.8 g DHA vs. placebo | ↓ resting HR |
| Delodder et al. (2015) [51] | IT | 8 | 23.3 (mean) | 1 intravenous infusion + 3 oral somministrations | 0.6 g/kg body weight of n-3 PUFAs + 0.6 g/kg/die | ↓ maximal HR |
| Holguin et al. (2005) [52] | RCT | 58 | >65 | 6 months | 2 g/day fish oil vs. 2 g/day soy oil | ↑ HR variability with fish oil ↑ HR variability with soy oil |
| Geelen et al. (2003) [53] | RCT | 84 | 50–70 | 12 weeks | 3.5 g/day fish oil vs. placebo | ⇔ HR variability and baroreflex sensitivity |
| Nilsson et al. (2012) [54] | RCT | 40 | Middle-aged to elderly | 5 weeks | 3 g/day fish oil vs. placebo | ↓ systolic BP |
| Sioen et al. (2009) [41] | IT | 59 | 22–65 | 12 weeks | ALA 5g + 1.5 of other PUFAs/day | ↓ diastolic BP |
| Theobald et al. (2007) [55] | RCT | 38 | 45–65 y | 3 months | 0.7 g DHA/day vs. placebo | ↓ diastolic BP |
| Macartney et al. (2014) [56] | RCT | 39 | 18–40 | 8 weeks | DHA 560 mg/day + EPA 140 mg/day vs. placebo | ↑ HR decrease in the recovery period ↓ HR during submaximal exercise |
| Effects on Thrombosis, Vascular Health and Inflammation | ||||||
| Phang et al. (2013) [57] | RCT | 94 | 39 (mean) | 4 weeks | 1000 mg EPA + 200 mg DHA/day vs. 200 mg EPA + 1000 mg DHA/day | ↓ platelet aggregation |
| Conquer et al. (1999) [58] | RCT | 19 | 29.5 (mean) | 42 days | 20 g seal oil vs. placebo | ↑ protein C, ↓ plasma fibrinogen |
| Mesa et al. (2004) [61] | RCT | 42 | 23–65 | 4 weeks | EPA-rich oil 9 g/day or a DHA-rich oil 9 g/day vs. placebo | ⇔ lag time for oxidation, ⇔ oxidation rate, ⇔ thrombotic tendency of oxidized LDL |
| Tholstrup et al. (2004) [27] | RCT | 16 | 35–75 | 3 weeks | Dietary replacement 4 g/day PUFA-rich oil | ⇔8-iso-PGF2α ⇔ Vitamin E |
| Kirkhus et al. (2012) [47] | RCT | 159 | 18–70 | 7 weeks | 1 g EPA and DHA/day vs. placebo | ⇔ inflammatory markers ⇔ markers for oxidative stress |
| Miles et al. (2001) [62] | RCT | 16 12 | <40 >55 | 12 weeks | 1.2 g/day of EPA + DHA | ↓ soluble E-selectin and VCAM-1 in the elderlies |
| Cazzola et al. (2007) [35] | RCT | 93 62 | 18–42 53–70 | 12 weeks | 1.35/2.7/4 g/day EPA | ↓ VCAM-1 |
| Barceló-Coblijn et al. (2008) [48] | RCT | 62 | >40 | 12 weeks | 1.2 g/2.4 g/3.6 g flaxseed oil/day vs. 0.6 or 1.2 g fish oil/day vs. placebo | ⇔ soluble VCAM-1 ⇔ C-reactive protein ⇔ Tumor Necrosis Factor α |
| Phang et al. (2013) [57] | RCT | 94 | 39.6 (mean) | 4 weeks | 200 mg EPA + 1000 mg DHA vs.1000 mg EPA + 200 mg DHA vs. placebo | ⇔ soluble CD36 |
| McEwen et al. (2015) [65] | IT | 40 | 21–64 | 4 weeks | 120 mg/day EPA + 520/day DHA | ↓ fibrin generation |
| Guillot et al. (2009) [66] | IT | 12 | 53–65 | 2 weeks each dose | 200, 400, 800, and 1600 mg/day DHA | ↓ platelet reactivity |
| Kaul et al. (2008) [46] | RCT | 86 | >18 | 12 weeks | 2 g/day fish oil vs. placebo/flaxseed/hempseed oil | ⇔ collagen- stimulated platelet aggregation ⇔ thrombin-stimulated platelet aggregation ⇔ inflammatory markers |
| Khan et al. (2003) [68] | RCT | 173 | 40–65 | 8 months | Placebo/oleic acid rich sunflower oil/evening primrose oil/soya bean oil/tuna fish oil/tuna-evening primrose oil mix | ↑ peak-response to acetylcholine |
| Shah et al. (2007) [50] | RCT | 26 | 26–36 | 14 days | 1 g/day fish oil vs. 1 g/day corn oil | ↑ endothelium-dependent brachial artery flow-mediated vasodilation, ↑ endothelium-independent nitroglycerin-mediated vasodilation |
| Singhal et al. (2013) [33] | RCT | 328 | 18–37 | 16 weeks | 1.6 g DHA/day vs. placebo | ⇔ endothelium-dependent brachial artery flow-mediated vasodilation |
| Miyoshi et al. (2014) [30] | RCT | 10/5 | 20–85 | 4 weeks | 1.9g DHA + 1.5g EPA/day | ↓ postprandial endothelial dysfunction |
| Fahs et al. (2010) [69] | RCT | 20 | 25 (mean) | 1 meal | 540 mg EPA + 360 mg DHA + 3 IU Vitamin E | No brachial artery flow-mediated dilatation impairment after a high fat meal |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manuelli, M.; Della Guardia, L.; Cena, H. Enriching Diet with n-3 PUFAs to Help Prevent Cardiovascular Diseases in Healthy Adults: Results from Clinical Trials. Int. J. Mol. Sci. 2017, 18, 1552. https://doi.org/10.3390/ijms18071552
Manuelli M, Della Guardia L, Cena H. Enriching Diet with n-3 PUFAs to Help Prevent Cardiovascular Diseases in Healthy Adults: Results from Clinical Trials. International Journal of Molecular Sciences. 2017; 18(7):1552. https://doi.org/10.3390/ijms18071552
Chicago/Turabian StyleManuelli, Matteo, Lucio Della Guardia, and Hellas Cena. 2017. "Enriching Diet with n-3 PUFAs to Help Prevent Cardiovascular Diseases in Healthy Adults: Results from Clinical Trials" International Journal of Molecular Sciences 18, no. 7: 1552. https://doi.org/10.3390/ijms18071552
APA StyleManuelli, M., Della Guardia, L., & Cena, H. (2017). Enriching Diet with n-3 PUFAs to Help Prevent Cardiovascular Diseases in Healthy Adults: Results from Clinical Trials. International Journal of Molecular Sciences, 18(7), 1552. https://doi.org/10.3390/ijms18071552