Moringa Leaves Prevent Hepatic Lipid Accumulation and Inflammation in Guinea Pigs by Reducing the Expression of Genes Involved in Lipid Metabolism


 and
and 
Abstract
:1. Introduction
2. Results
2.1. Food Intake and Body and Liver Weight
2.2. Plasma Lipids, Glucose and Liver Enzymes
2.3. Lipoprotein Size and Subfractions
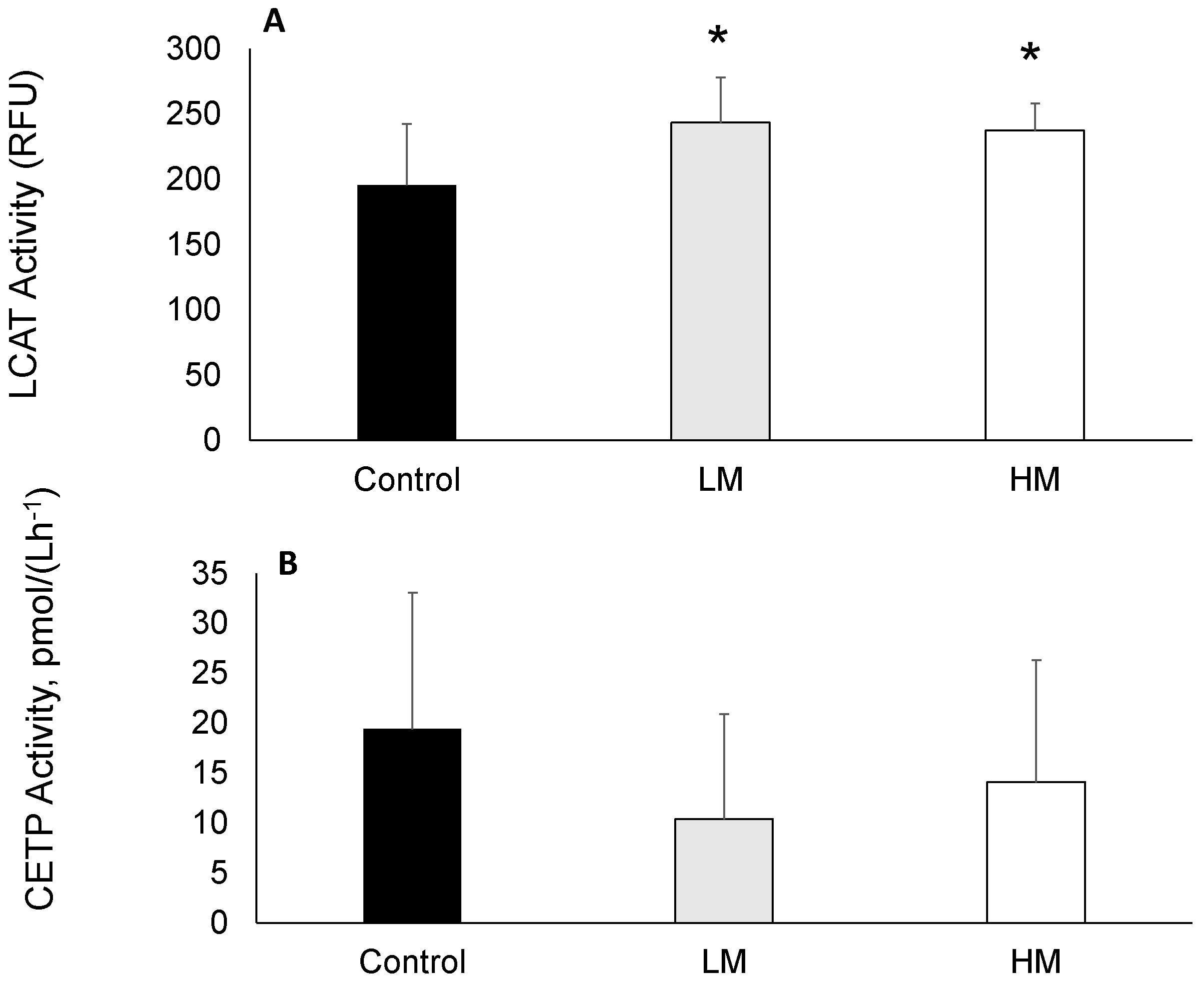
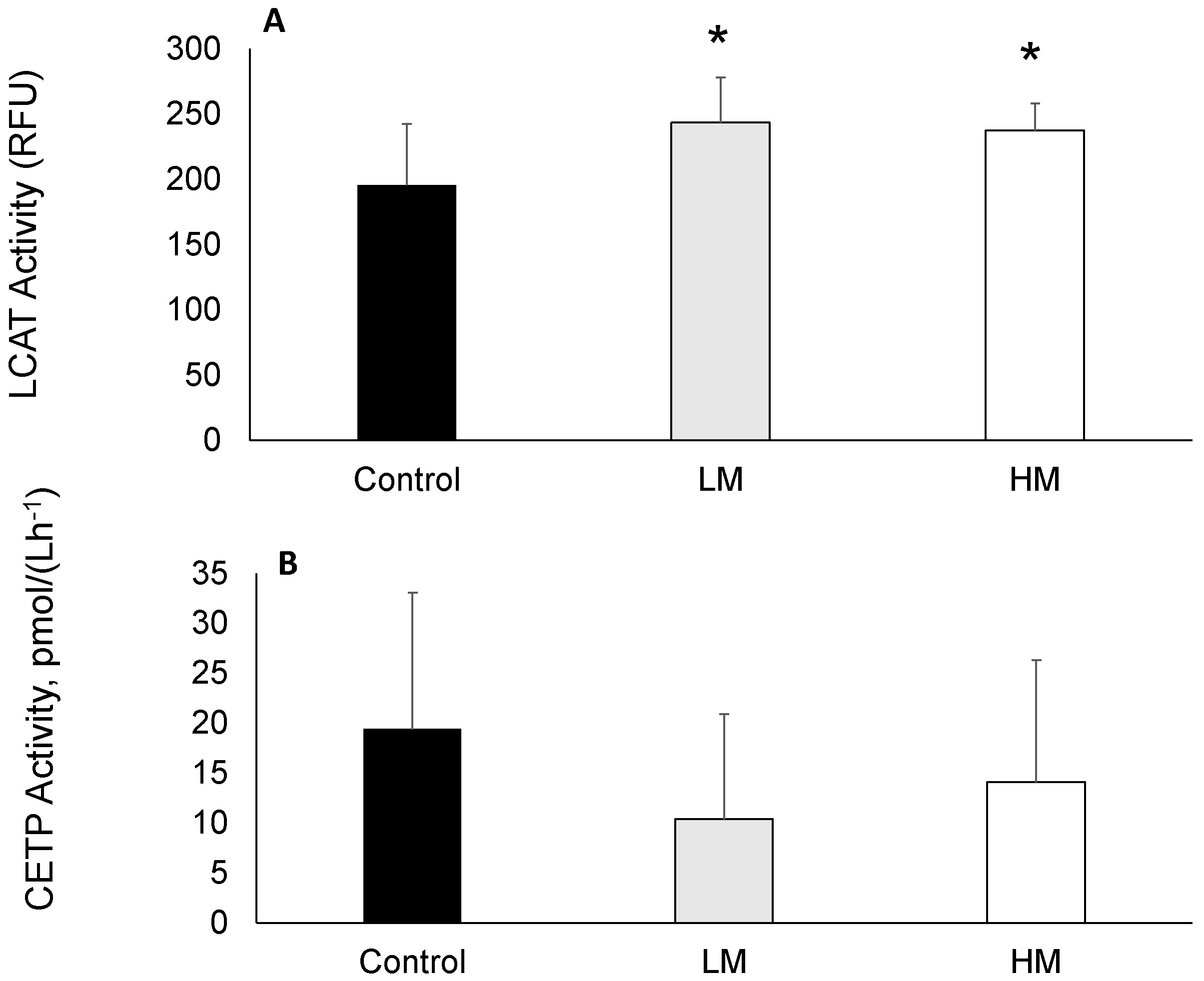
2.4. LCAT and CETP Activity
2.5. Hepatic Lipids
2.6. Liver Inflammation
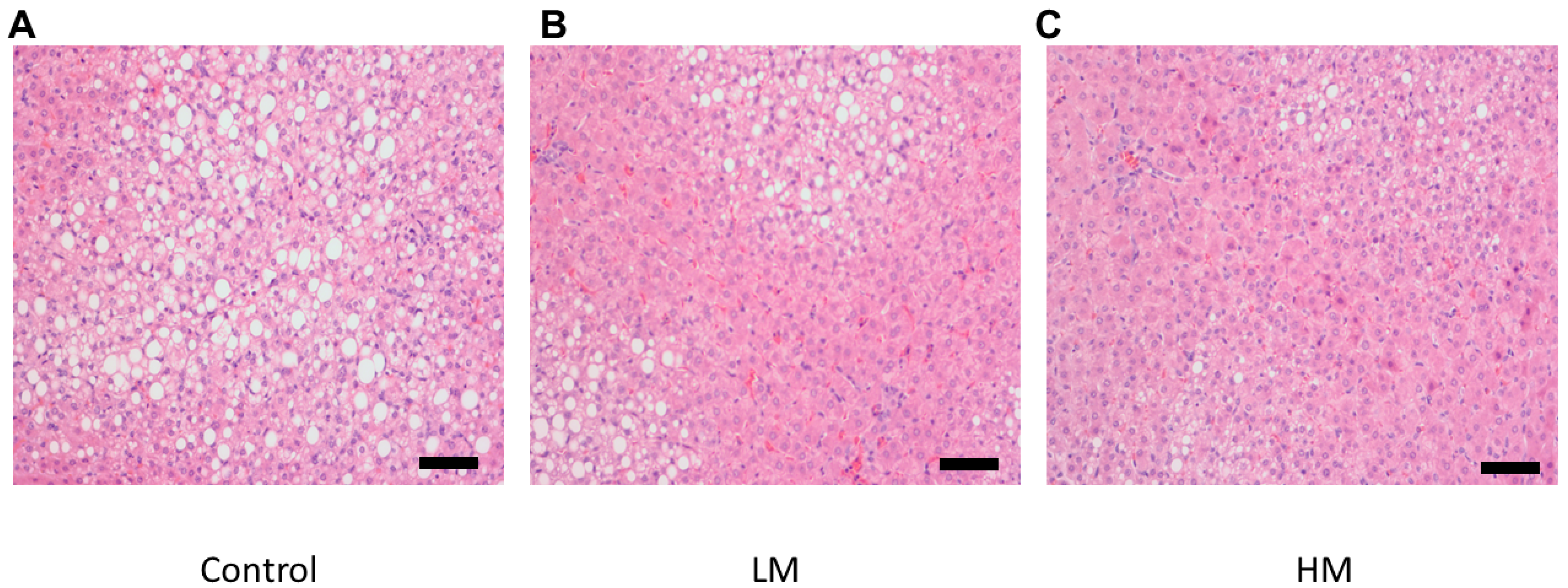
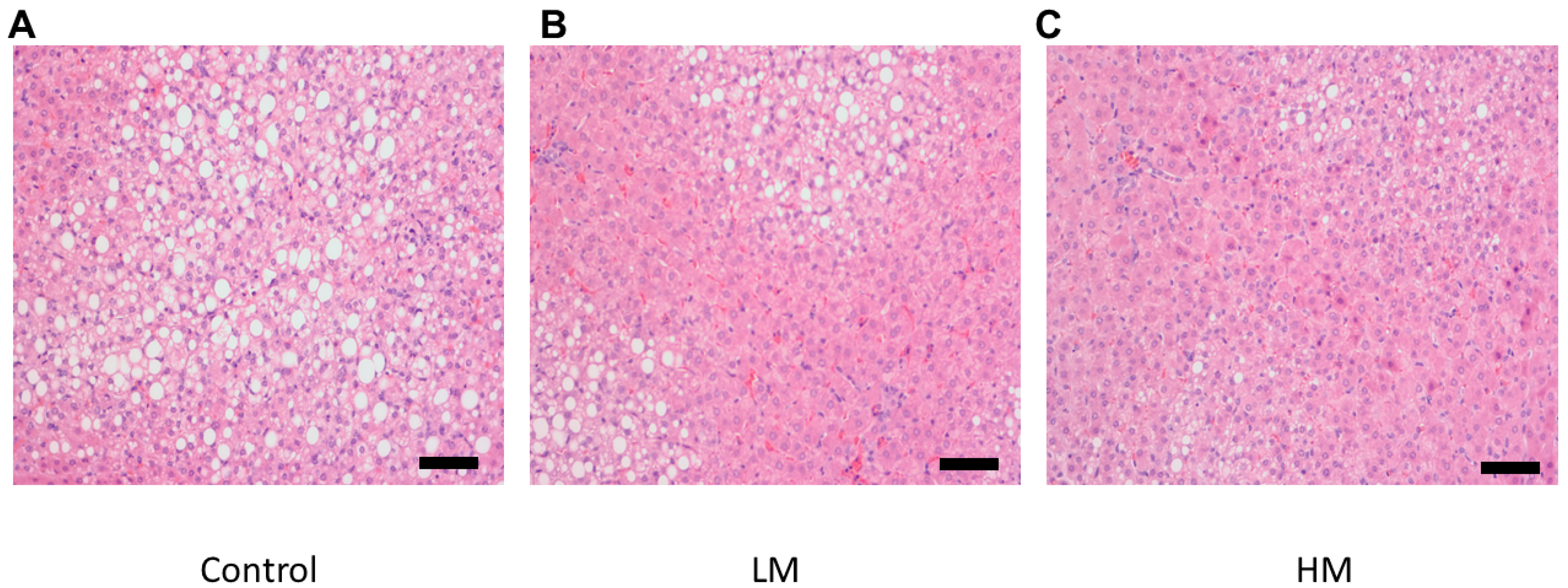
2.7. Histology
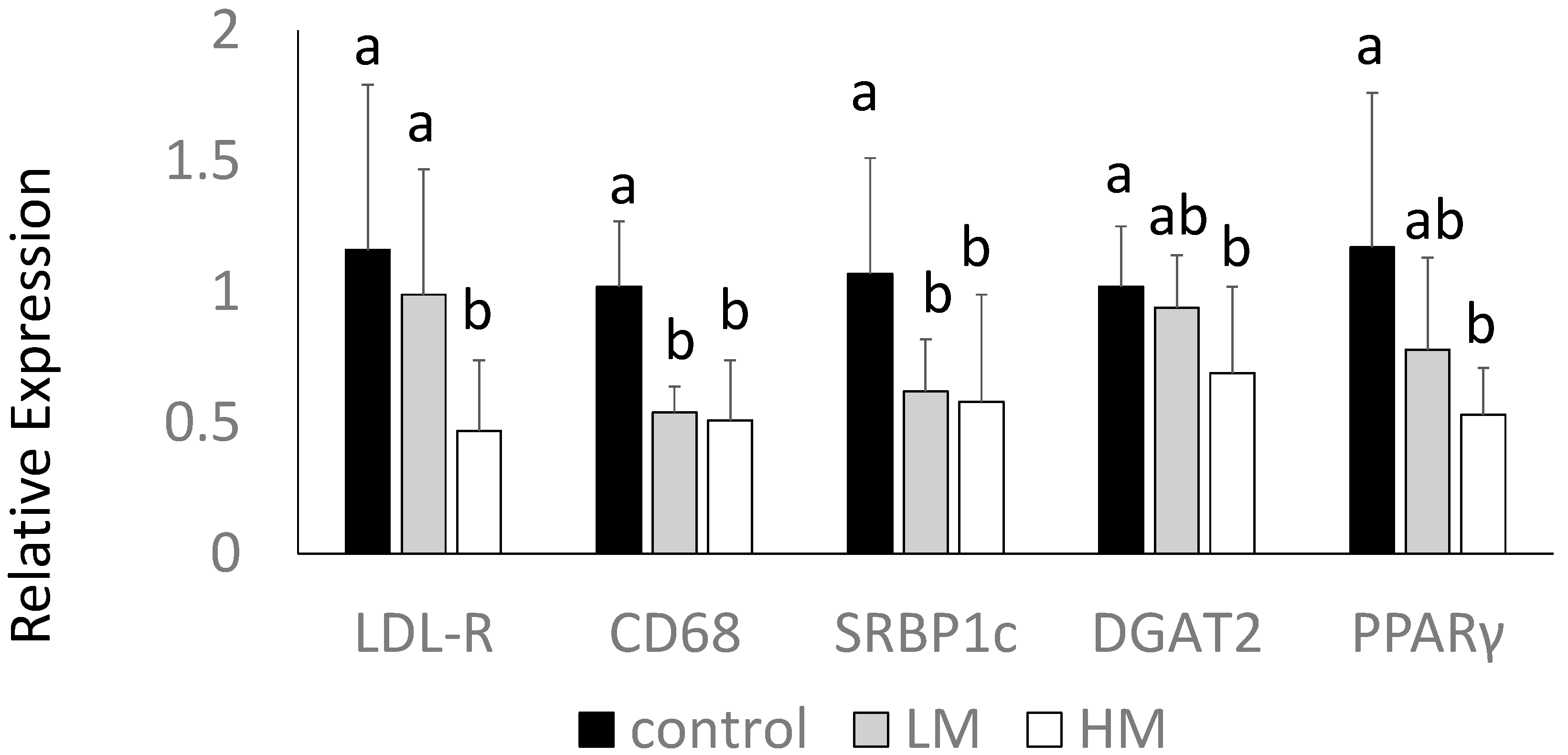
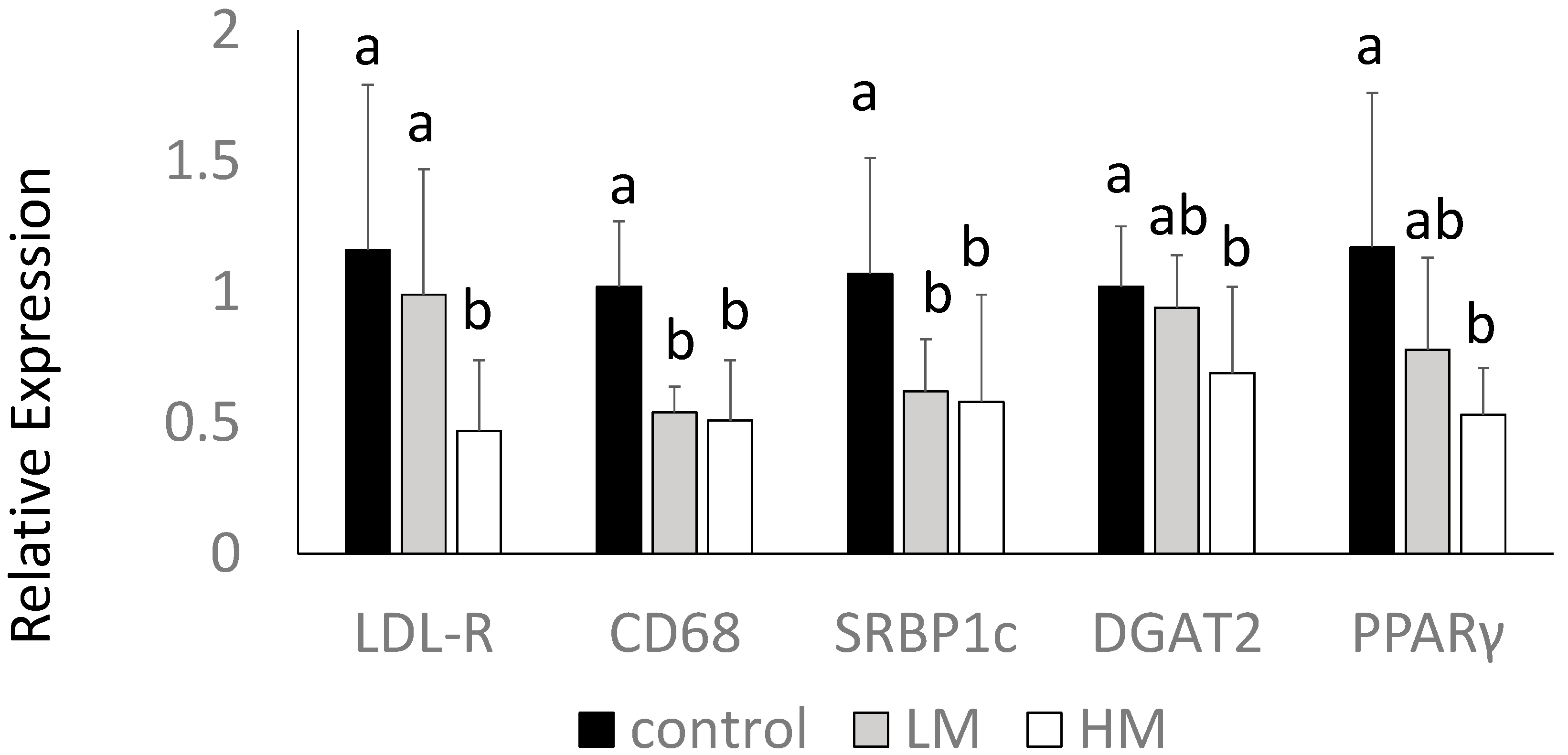
2.8. Hepatic Gene Expression
3. Discussion
3.1. Moringa Leaves and Plasma Lipids and Liver Enzymes
3.2. Moringa Leaves and Hepatic Lipids, Inflammation, and Gene Expression
4. Materials and Methods
4.1. Experimental Design
4.2. Plasma Lipids, Glucose and Liver Enzymes
4.3. Lipoprotein Subfraction and Size
4.4. Lecithin Cholesterol Acyl Transferase (LCAT) and Choleterol Ester Protein (CETP) Acitivities
4.5. Hepatic Lipids
4.6. Histologic Evaluation
4.7. Inflammatory Cytokine Concentration in Liver
4.8. Gene Expression
4.9. RNA Isolation, cDNA Synthesis and Real-Time qRT-PCR
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Angulo, P. Nonalcoholic fatty liver disease. N. Engl. J. Med. 2014, 346, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Tolman, K.G.; Dalpiaz, A.S. Treatment of non-alcoholic fatty liver disease. Ther. Clin. Risk Manag. 2007, 3, 1153–1163. [Google Scholar] [PubMed]
- Krawczyk, M.; Bonfrate, L.; Portincasa, P. Nonalcoholic fatty liver disease. Best Pract. Res. Clin. Gastroenterol. 2010, 24, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Milic, N.; Peta, V.; Alfieri, F.; de Lorenzo, A.; Bellentani, S. Alimentary regimen in non-alcoholic fatty liver disease: Mediterranean diet. World J. Gastroenterol. 2014, 20, 16831–16840. [Google Scholar] [CrossRef] [PubMed]
- Masarone, M.; Federico, A.; Abenavoli, L.; Loguercio, C.; Persico, M. Non alcoholic fatty liver: Epidemiology and natural history. Rev. Recent Clin. Trials 2014, 9, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Murillo, A.; DiMarco, D.; Fernandez, M. The potential of non-provitamin A carotenoids for the prevention and treatment of non-alcoholic fatty liver disease. Biology 2016, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tan, H.-Y.; Wang, N.; Zhang, Z.-J.; Lao, L.; Wong, C.-W.; Feng, Y. The role of oxidative stress and antioxidants in liver diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed]
- Leone, A.; Spada, A.; Battezzati, A.; Schiraldi, A.; Aristil, J.; Bertoli, S. Moringa oleifera seeds and oil: Characteristics and uses for human health. Int. J. Mol. Sci. 2016, 17, 2141. [Google Scholar] [CrossRef] [PubMed]
- Leone, A.; Spada, A.; Battezzati, A.; Schiraldi, A.; Aristil, J.; Bertoli, S. Cultivation, genetic, ethnopharmacology, phytochemistry and pharmacology of Moringa oleifera leaves: An overview. Int. J. Mol. Sci. 2015, 16, 12791–12835. [Google Scholar] [CrossRef] [PubMed]
- Adedapo, A.A.; Falayi, O.O.; Oyagbemi, A.A. Evaluation of the analgesic, anti-inflammatory, anti-oxidant, phytochemical and toxicological properties of the methanolic leaf extract of commercially processed Moringa oleifera in some laboratory animals. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmari, A.K.; Al-Elaiwi, A.M.; Athar, M.T.; Tariq, M.; Al Eid, A.; Al-Asmary, S.M. A review of hepatoprotective plants used in Saudi traditional medicine. Evid. Based Complement. Altern. Med. 2014, 2014, 890842. [Google Scholar] [CrossRef] [PubMed]
- Ghasi, S.; Nwobodo, E.; Ofili, J.O. Hypocholesterolemic effects of crude extract of leaf of Moringa oleifera Lam in high-fat diet fed wistar rats. J. Ethnopharmacol. 2000, 69, 21–25. [Google Scholar] [CrossRef]
- Fakurazi, S.; Hairuszah, I.; Nanthini, U. Moringa oleifera Lam prevents acetaminophen induced liver injury through restoration of glutathione level. Food Chem. Toxicol. 2008, 46, 2611–2615. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, D.; Rai, P.K.; Kumar, A.; Mehta, S.; Watal, G. Effect of Moringa oleifera Lam leaves aqueous extract therapy on hyperglycemic rats. J. Ethnopharmacol. 2009, 123, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, D.; Rai, P.K.; Mehta, S.; Chatterji, S.; Shukla, S.; Rai, D.K.; Sharma, G.; Sharma, B.; Khair, S.; Watal, G. Role of Moringa oleifera in regulation of diabetes-induced oxidative stress. Asian Pac. J. Trop. Med. 2013, 6, 426–432. [Google Scholar] [CrossRef]
- Nandave, M.; Ojha, S.K.; Joshi, S.; Kumari, S.; Arya, D.S. Moringa oleifera leaf extract prevents isoproterenol-induced myocardial damage in rats: Evidence for an antioxidant, antiperoxidative, and cardioprotective intervention. J. Med. Food 2009, 12, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Mehta, L.K.; Balaraman, R.; Amin, A.H.; Bafna, P.A.; Gulati, O.D. Effect of fruits of Moringa oleifera on the lipid profile of normal and hypercholesterolaemic rabbits. J. Ethnopharmacol. 2003, 86, 191–195. [Google Scholar] [CrossRef]
- Chávez-Santoscoy, R.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Effect of flavonoids and saponins extracted from black bean (Phaseolus vulgaris L.) seed coats as cholesterol micelle disruptors. Plant Foods Hum. Nutr. 2013, 68, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Minaiyan, M.; Asghari, G.; Taheri, D.; Saeidi, M.; Nasr-Esfahani, S. Anti-inflammatory effect of Moringa oleifera Lam seeds on acetic acid-induced acute colitiis in rats. Avicenna J. Phytomed. 2014, 4, 127–136. [Google Scholar] [PubMed]
- Singh, B.N.; Singh, B.R.; Singh, R.L.; Prakash, D.; Dhakarey, R.; Upadhyay, G.; Singh, H.B. Oxidative DNA damage protective activity, antioxidant and anti-quorum sensing potentials of Moringa oleifera. Food Chem. Toxicol. 2009, 47, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Labbe, D.; Provencal, M.; Lamy, S.; Boivin, D.; Gingras, D.; Beliveau, R.; Be, R.; Provenc, M. The flavonols quercetin, kaempferol, and myricetin inhibit hepatocyte growth factor-induced medulloblastoma cell migration. J. Nutr. 2009, 139, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Pari, L.; Kumar, N.A. Hepatoprotective activity of Moringa oleifera on antitubercular drug-induced liver damage in rats. J. Med. Food 2002, 5, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Sharifudin, S.A.; Fakurazi, S.; Hidayat, M.T.; Hairuszah, I.; Aris Mohd Moklas, M.; Arulselvan, P. Therapeutic potential of Moringa oleifera extracts against acetaminophen-induced hepatotoxicity in rats. Pharm. Biol. 2013, 51, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Hamza, A.A. Ameliorative effects of Moringa oleifera Lam seed extract on liver fibrosis in rats. Food Chem. Toxicol. 2010, 48, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Sinha, M.; Das, D.K.; Bhattacharjee, S.; Majumdar, S.; Dey, S. Leaf extract of Moringa oleifera prevents ionizing radiation-induced oxidative stress in mice. J. Med. Food 2011, 14, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Hartman, M.J. Review of the safety and efficacy of Moringa oleifera. Phytother. Res. 2015, 29, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Al-Sarraj, T.; Saadi, H.; Volek, J.S.; Fernandez, M.L. Carbohydrate restriction favorably alters lipoprotein metabolism in Emirati subjects classified with the metabolic syndrome. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Carr, T.P.; Andresen, C.J.; Rudel, L.L. Enzymatic determination of triglyceride, free cholesterol, and total cholesterol in tissue lipid extracts. Clin. Biochem. 1993, 26, 39–42. [Google Scholar] [CrossRef]
- Kim, J.E.; Leite, J.O.; Smyth, J.A.; Clark, R.M.; Fernandez, M.L. A lutein-enriched diet prevents cholesterol accumulation and decreases oxidized LDL and inflammatory cytokines in the aorta of guinea pigs. J. Nutr. 2011, 141, 1458–1463. [Google Scholar] [CrossRef] [PubMed]
- Murillo, A.G.; Aguilar, D.; Norris, G.H.; Dimarco, D.M.; Missimer, A.; Hu, S.; Smyth, J.A.; Gannon, S.; Blesso, C.N.; Luo, Y.; et al. Compared with powdered lutein, a lutein nanoemulsion increases plasma and liver lutein, protects against hepatic steatosis, and affects lipoprotein metabolism in guinea pigs. J. Nutr. 2016, 146, 1961–1969. [Google Scholar] [CrossRef] [PubMed]
- Farrell, N.; Norris, G.; Lee, S.G.; Chun, O.K.; Blesso, C.N. Anthocyanin-rich black elderberry extract improves markers of HDL function and reduces aortic cholesterol in hyperlipidemic mice. Food Funct. 2015, 6, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Mu, Y.M.; Li, T.T.; Yang, Y.L.; Zhang, M.T.; Li, Y.S.; Zhang, W.K.; Tang, H.B.; Shang, H.C. Gynura procumbens reverses acute and chronic ethanol-induced liver steatosis through MAPK/SREBP-1c-dependent and -independent pathways. J. Agric. Food Chem. 2015, 63, 8460–8471. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Cao, J.; Feng, Q.; Peng, J.; Hu, Y. Roles of chlorogenic acid on regulating glucose and lipids metabolism: A review. Evid. Based Complement. Altern. Med. 2013, 2013, 801457. [Google Scholar] [CrossRef] [PubMed]
- Gebregiorgis, F.; Negesse, T.; Nurfeta, A. Feed intake and utilization in sheep fed graded levels of dried Moringa (Moringa stenopetala) leaf as a supplement to Rhodes grass hay. Trop. Anim. Health Prod. 2012, 44, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Giannini, E.G.; Testa, R.; Savarino, V. Liver enzyme alteration: A guide for clinicians. Can. Med. Assoc. J. 2005, 172, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Ouédraogo, M.; Lamien-Sanou, A.; Ramdé, N.; Ouédraogo, A.S.; Ouédraogo, M.; Zongo, S.P.; Goumbri, O.; Duez, P.; Guissou, P.I. Protective effect of Moringa oleifera leaves against gentamicin-induced nephrotoxicity in rabbits. Exp. Toxicol. Pathol. 2013, 65, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.L.; Vergara-Jimenez, M.; Conde, K.; Behr, T.; Abdel-Fattah, G. Regulation of apolipoprotein B-containing lipoproteins by dietary soluble fiber in guinea pigs. Am. J. Clin. Nutr. 1997, 65, 814–822. [Google Scholar] [PubMed]
- Osono, Y.; Woollett, L.A.; Herz, J.; Dietschy, J.M. Role of the low density lipoprotein receptor in the flux of cholesterol through the plasma and across the tissues of the mouse. J. Clin. Invest. 1995, 95, 1124–1132. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.L.; Ruiz, L.R.; Conde, A.K.; Sun, D.M.; Erickson, S.K.; McNamara, D.J. Psyllium reduces plasma LDL in guinea pigs by altering hepatic cholesterol homeostasis. J. Lipid Res. 1995, 36, 1128–1138. [Google Scholar] [PubMed]
- Fernandez, M.L.; Vergara-Jimenez, M.; Conde, K.; Abdel-Fattah, G. Dietary carbohydrate type and fat amount alter VLDL and LDL metabolism in guinea pigs. J. Nutr. 1996, 126, 2494–2504. [Google Scholar] [PubMed]
- Duprez, D.A.; Otvos, J.; Tracy, R.P.; Feingold, K.R.; Gross, M.D.; Lima, J.A.C.; Mackey, R.H.; Neaton, J.D.; Sanchez, O.A.; Jacobs, D.R. High-density lipoprotein subclasses and noncancer chronic inflammatory-related events versus cardiovascular events: The multi-ethnic study of atherosclerosis. J. Am. Heart Assoc. 2015, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Mutungi, G.; Waters, D.; Ratliff, J.; Puglisi, M.; Clark, R.M.; Volek, J.S.; Luz, M. Eggs distinctly modulate plasma carotenoid and lipoprotein subclasses in adult men following a carbohydrate-restricted diet. J. Nutr. Biochem. 2009, 21, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Ossoli, A.; Pavanello, C.; Calabresi, L. High-density lipoprotein, lecithin: Cholesterol acyltransferase, and atherosclerosis. Endocrinol. Metab. 2016, 31, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Barter, P.J.; Rye, K.-A. Cholesteryl ester transfer protein inhibition as a strategy to reduce cardiovascular risk. J. Lipid Res. 2012, 53, 1755–1766. [Google Scholar] [CrossRef] [PubMed]
- De Ogburn, R.; Leite, J.O.; Ratliff, J.; Volek, J.S.; McGrane, M.M.; Fernandez, M.L. Effects of increased dietary cholesterol with carbohydrate restriction on hepatic lipid metabolism in guinea pigs. Comp. Med. 2012, 62, 109–115. [Google Scholar]
- Atawodi, S.E.; Atawodi, J.C.; Idakwo, G.A.; Pfundstein, B.; Haubner, R.; Wuntele, G.; Bartsch, H.; Owen, R.W. Evaluation of the polypehnol content and antioxidant properties of methanol extracts of the leaves, stem and root barks of Moringa oleifera Lam. J. Med. Food 2010, 13, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Dong, L.; Dang, X.; Liu, Y.; Jiang, J.; Wang, Y.; Lu, X.; Guo, X. Effect of chlorogenic acid on LPS-induced proinflammatory signaling in hepatic stellate cells. Inflamm. Res. 2013, 62, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.S.; Jeon, S.M.; Kim, M.J.; Yeo, J.; Seo, K.I.; Choi, M.S.; Lee, M.K. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obese mice. Food Chem. Toxicol. 2010, 48, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D. Sterol regulatory element-binding proteins: Transcriptional activators of lipid synthesis. Biochem. Soc. Trans. 2002, 30, 1091–1095. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, G.N. The role of cholesterol in the pathogenesis of NASH. Trends Endocrinol. Metab. 2016, 27, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Waterman, C.; Cheng, D.M.; Rojas-Silva, P.; Poulev, A.; Dreifus, J.; Lila, M.A.; Raskin, I. Stable, water extractable isothiocyanates from Moringa oleifera leaves attenuate inflammation in vitro. Phytochemistry 2014, 103, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Shi, L.; Sheng, Y.; Zhang, J.; Lu, B.; Ji, L. Chlorogenic acid suppresses monocrotaline-induced sinusoidal obstruction syndrome: The potential contribution of NFκB, Egr1, Nrf2, MAPKs and PI3K signals. Environ. Toxicol. Pharmacol. 2016, 46, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Tobe, K.; Aoyama, M.; Sakamoto, K.; Ohsugi, M.; Kamei, N.; Nemoto, S.; Inoue, A.; Ito, Y.; Uchida, S.; et al. Expression of DGAT2 in white adipose tissue is regulated by central leptin action. J. Biol. Chem. 2005, 280, 3331–3337. [Google Scholar] [CrossRef] [PubMed]
- Casaschi, A.; Wang, Q.; Dang, K.O.; Richards, A.; Theriault, A. Intestinal Apolipoprotein B secretion is inhibited by the flavonoid quercitin: Potential role of microsomal triglyceride transfer protein and dicaylglycerol acyltranferase. Lipids 2002, 37, 547–652. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | Low Moringa | High Moringa |
|---|---|---|---|
| Total cholesterol (mg/dL) | 294.6 ± 167.0 a | 192.9 ± 84.7 a | 251.6 ± 70.3 a |
| VLDL cholesterol (mg/dL) | 11.0 ± 2.9 a | 10.6 ± 5.0 a | 11.9 ± 4.0 a |
| LDL cholesterol (mg/dL) | 244.5 ± 166.4 a | 155.0 ± 73.9 a | 187.7 ± 73.4 a |
| HDL cholesterol (mg/dL) | 39.1 ± 28.3 a | 27.4 ± 24.2 a | 52.0 ± 30.3 a |
| Triglycerides (mg/dL) | 55.9 ± 13.9 a | 53.6 ± 24.7 a | 59.5 ± 18.9 a |
| Glucose (mg/dL) | 158.4 ± 16.5 a | 154.4 ± 23.4 a | 164.6 ± 18.9 a |
| ALT (IU/L) | 239.3 ± 144.6 a | 109.4 ± 43.7 b | 141.9 ± 73.6 b |
| AST (IU/L) | 552.9 ± 479.4 a | 276.9 ± 146.5 a | 299.4 ± 195.1 a |
| Parameter | Control | Low Moringa | High Moringa |
|---|---|---|---|
| Total VLDL (nmol/L) | 55.2 ±1 28.1 a | 45.4 ± 21.4 a | 61.5 ± 1.7 a |
| Large VLDL (60–100 nm) | 1.7 ± 1.3 a | 1.1 ± 0.9 a | 1.1 ± 0.9 a |
| Medium VLDL (40–60 nm) | 14.3 ± 7.0 a | 13.7 ± 7.6 a | 16.7 ± 7.2 a |
| Small VLDL (30–40 nm) | 39.1 ± 21.2 a | 30.6 ± 14.0 a | 16.7 ± 7.2 a |
| Total LDL (nmol/L) | 531 ± 219 a | 563 ± 173 a | 709 ± 284 a |
| IDL (nmol/L) | 63 ± 31 a | 80 ± 33 a | 107 ± 52 a |
| Large LDL (23–30 nm) | 148 ± 61 a | 102 ± 46 a | 104 ± 30 a |
| Small LDL (18–23 mn) | 320 ± 160 a | 382 ± 123 a | 498 ± 252 a |
| Total HDL (µmol/L) | 1.5±0.6 a | 0.9 ± 0.7 a | 1.5 ± 0.5 |
| Large HDL (10–13 nm) | 0.19±0.13 a | 0.14±0.09 a | 0.21±0.08 a |
| Medium HDL (8.2–10) | 0.56± 0.55 a | 0.11±0.21 b | 0.21±0.13 b |
| Small HDL (7.3–8.2) | 0.8±0.6 a | 0.6±0.7 a | 1.0±0.5 a |
| VLDL size (nm) | 51.0 ± 5.7 a | 46.4 ± 5.5 a | 47.4 ± 5.5 |
| LDL size (nm) | 20.8 ± 0.6 a | 20.3 ± 0.5 a | 20.3 ± 0.7 a |
| HDL size (nm) | 9.5 ± 1.0 a | 10.1 ± 0.7 a | 9.7 ± 0.5 a |
| Parameter | Control | Low Moringa | High Moringa |
|---|---|---|---|
| Total Cholesterol (mmol/g) | 28.1 ± 7.2 a | 23.3 ± 4.1 a,b | 16.0 ± 7.8 b |
| Free Cholesterol (mmol/g) | 11.6 ± 2.3 a | 8.3 ± 2.6 b | 6.5 ± 4.2 b |
| Esterified cholesterol (mmol/g) | 16.3 ± 5.4 a | 15,0 ± 2.8 a,b | 9.8 ± 4.7 b |
| Triglycerides (mmol/g) | 57.7 ± 11.5 a | 48.8 ± 9.9 a,b | 34.5 ± 13.0 b |
| Parameter | Control | Low Moringa | High Moringa |
|---|---|---|---|
| IL-1β (ng/g) | 196.4 ± 49.2 a | 180.3 ± 40.9 a,b | 143.1 ± 31.5 b |
| IL-6 (ng/g) | 30.1 ± 3.7 a | 27.5 ± 4.9 a | 37.3 ± 16.4 a |
| IL-10 (ng/g) | 233 ± 54 a | 208 ± 42 a,b | 160 ± 38 b |
| IFNγ (ng/g) | 194.4 ± 20.5 a | 173.4 ± 39.2 a,b | 153.4 ± 42.3 b |
| MCP-1 (ng/g) | 11.4 ± 1.1 a | 10.3 ± 1.2 a | 21.9 ± 21.7 a |
| TNFα (ng/g) | 31.1 ± 4.9 a | 27.4 ± 4.4 a | 35.6 ± 19.3 a |
| Component | Amount (g/100 g) |
|---|---|
| Carbohydrate | 30 |
| Fiber | 44 |
| Protein | 5 |
| Fat | 3 |
| Moisture | 10 |
| Minerals | 8 |
| Component | Control | Low Moringa (LM) | High Moringa (HM) | |||
|---|---|---|---|---|---|---|
| g/100 g | % Energy | g/100 g | % Energy | g/100 g | % Energy | |
| Protein 1 | 22 | 23 | 20 | 23 | 17 | 23 |
| Carbohydrate | 41 | 41.9 | 39 | 41.9 | 38 | 41.9 |
| Fat 2 | 15.1 | 35.1 | 14.8 | 35.1 | 14.7 | 35.1 |
| Vitamins 3 | 1.1 | - | 1.1 | - | 1.0 | - |
| Minerals 3 | 8.1 | - | 8.1 | - | 7.8 | - |
| Cellulose | 10 | - | 7 | - | 5 | - |
| Guar Gum | 2.5 | - | 2.5 | - | 2.5 | - |
| Cholesterol | 0.25 | - | 0.25 | - | 0.25 | - |
| Moringa 4 | 0 | - | 10 | - | 15 | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almatrafi, M.M.; Vergara-Jimenez, M.; Murillo, A.G.; Norris, G.H.; Blesso, C.N.; Fernandez, M.L. Moringa Leaves Prevent Hepatic Lipid Accumulation and Inflammation in Guinea Pigs by Reducing the Expression of Genes Involved in Lipid Metabolism. Int. J. Mol. Sci. 2017, 18, 1330. https://doi.org/10.3390/ijms18071330
Almatrafi MM, Vergara-Jimenez M, Murillo AG, Norris GH, Blesso CN, Fernandez ML. Moringa Leaves Prevent Hepatic Lipid Accumulation and Inflammation in Guinea Pigs by Reducing the Expression of Genes Involved in Lipid Metabolism. International Journal of Molecular Sciences. 2017; 18(7):1330. https://doi.org/10.3390/ijms18071330
Chicago/Turabian StyleAlmatrafi, Manal Mused, Marcela Vergara-Jimenez, Ana Gabriela Murillo, Gregory H. Norris, Christopher N. Blesso, and Maria Luz Fernandez. 2017. "Moringa Leaves Prevent Hepatic Lipid Accumulation and Inflammation in Guinea Pigs by Reducing the Expression of Genes Involved in Lipid Metabolism" International Journal of Molecular Sciences 18, no. 7: 1330. https://doi.org/10.3390/ijms18071330
APA StyleAlmatrafi, M. M., Vergara-Jimenez, M., Murillo, A. G., Norris, G. H., Blesso, C. N., & Fernandez, M. L. (2017). Moringa Leaves Prevent Hepatic Lipid Accumulation and Inflammation in Guinea Pigs by Reducing the Expression of Genes Involved in Lipid Metabolism. International Journal of Molecular Sciences, 18(7), 1330. https://doi.org/10.3390/ijms18071330