
Sulfiredoxin May Promote Cervical Cancer Metastasis via Wnt/β-Catenin Signaling Pathway

Abstract
:1. Introduction
2. Results
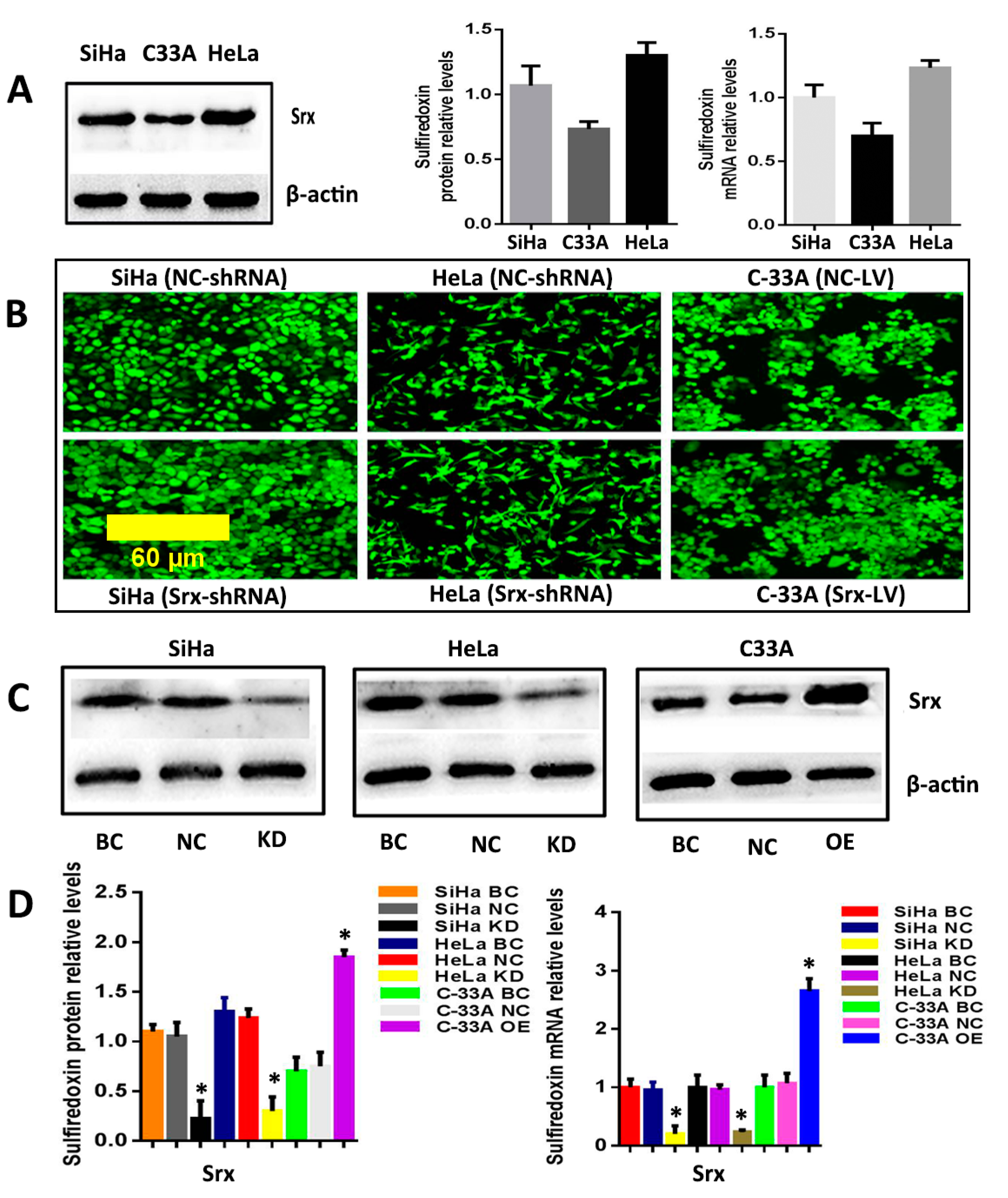
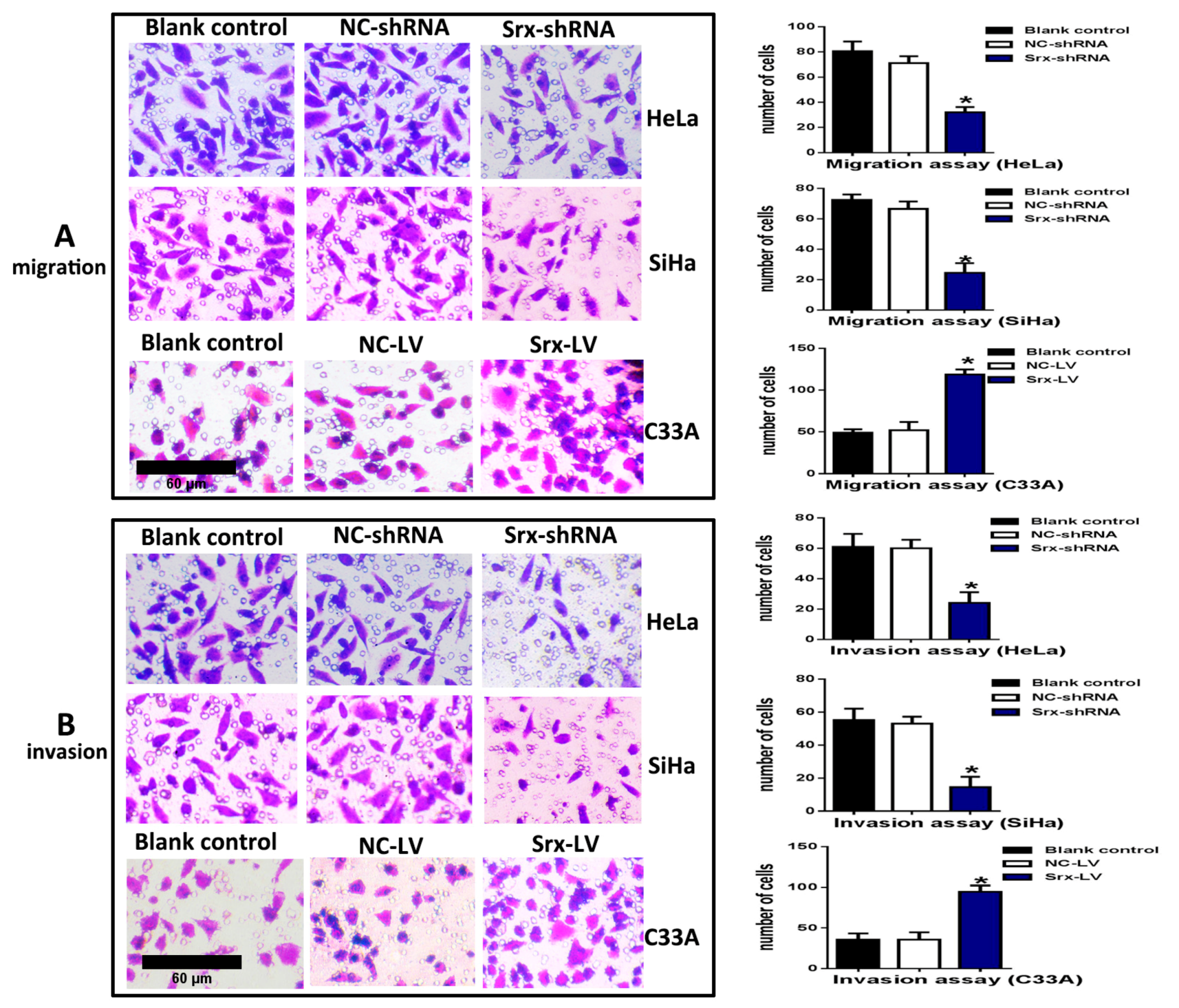
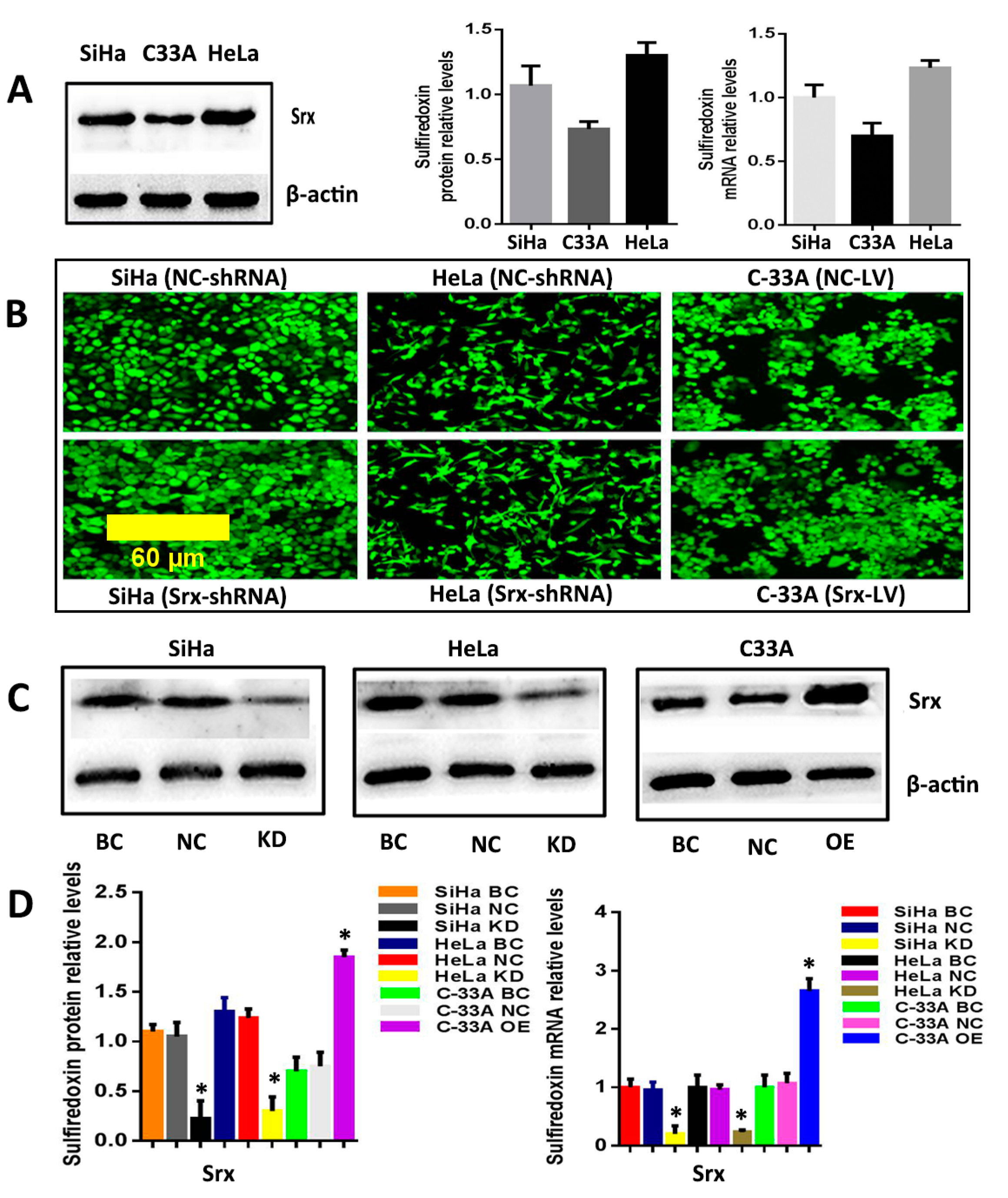
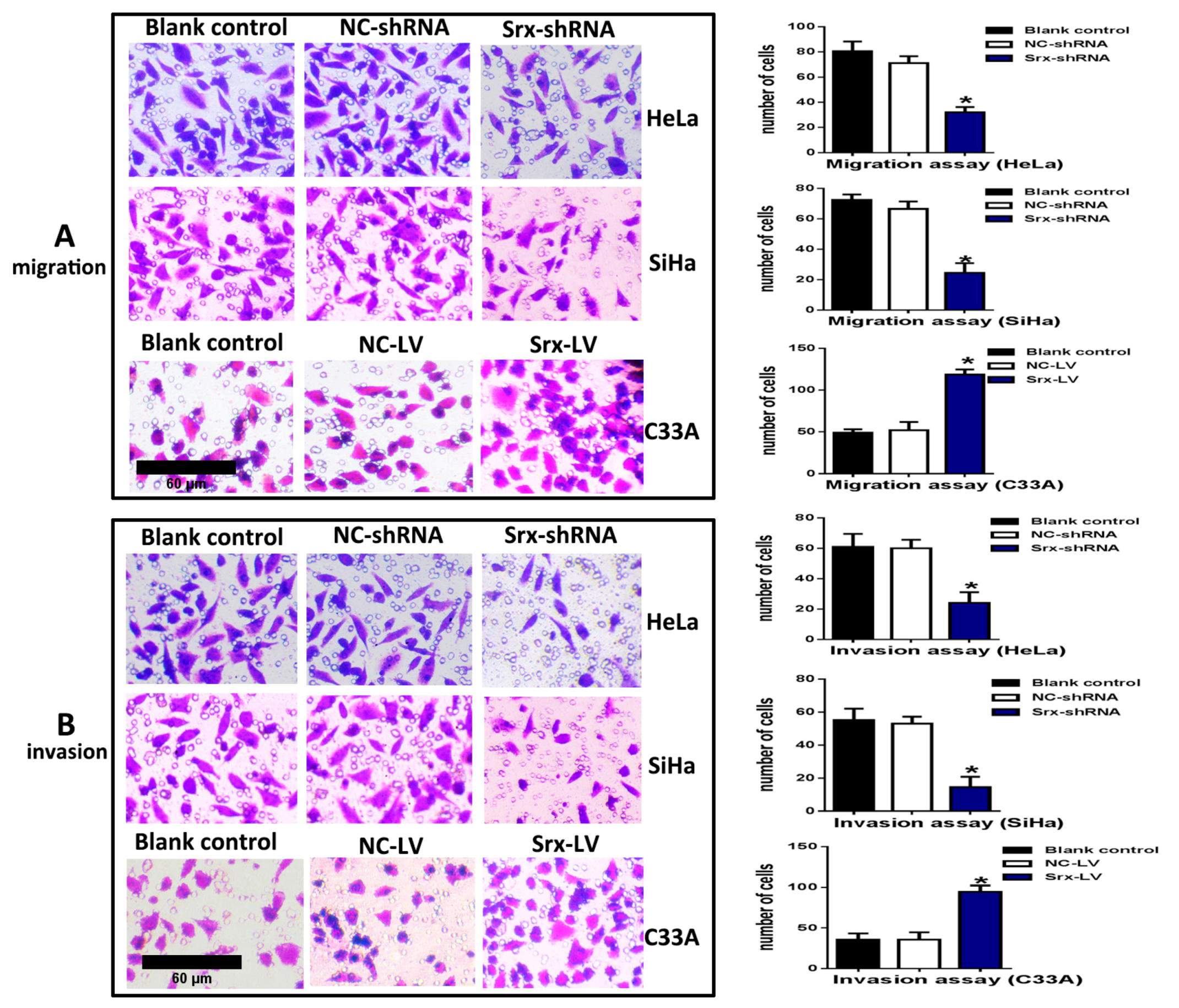
2.1. Srx Promotes the Migration and Invasion of Cervical Cancer Cells
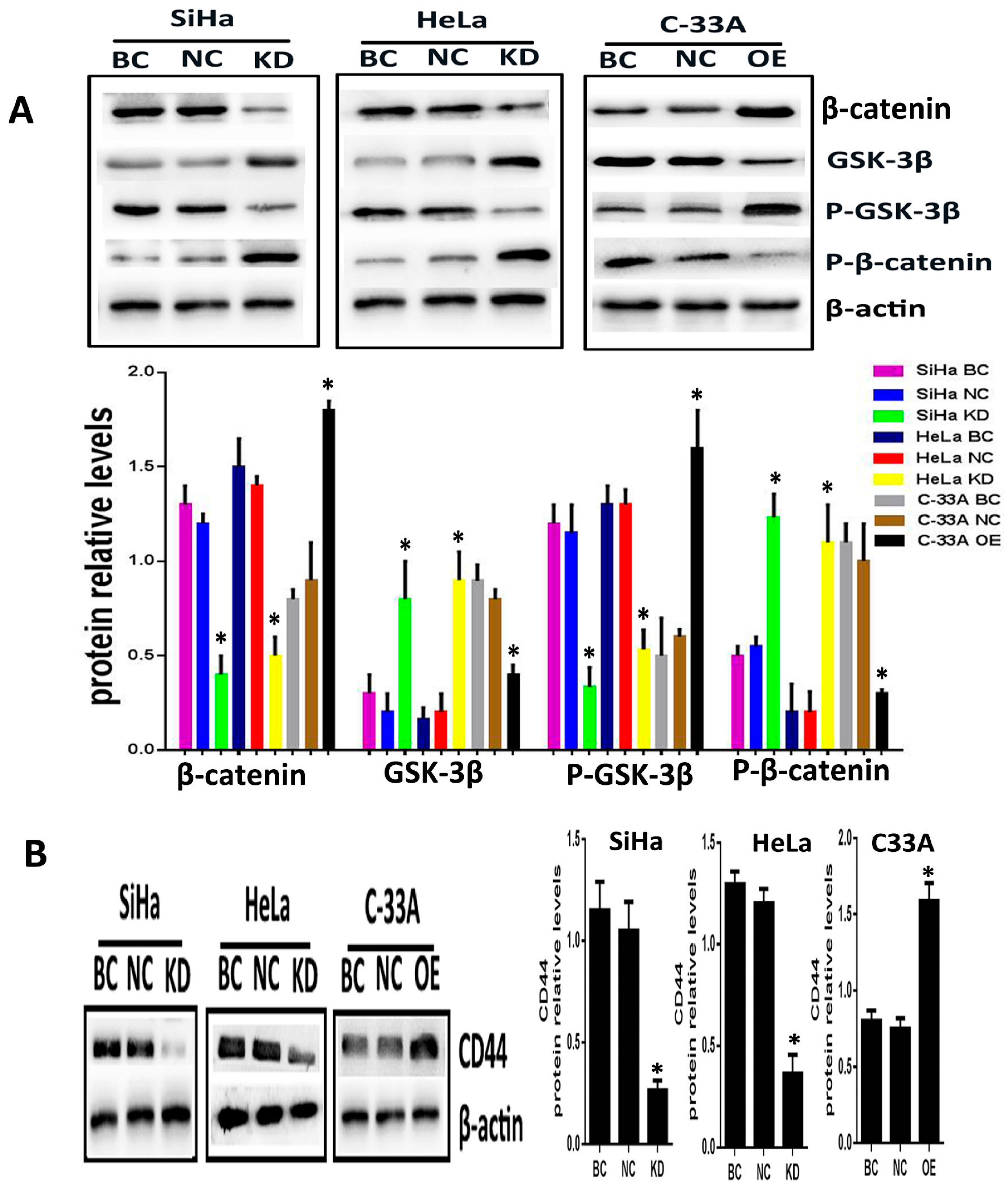
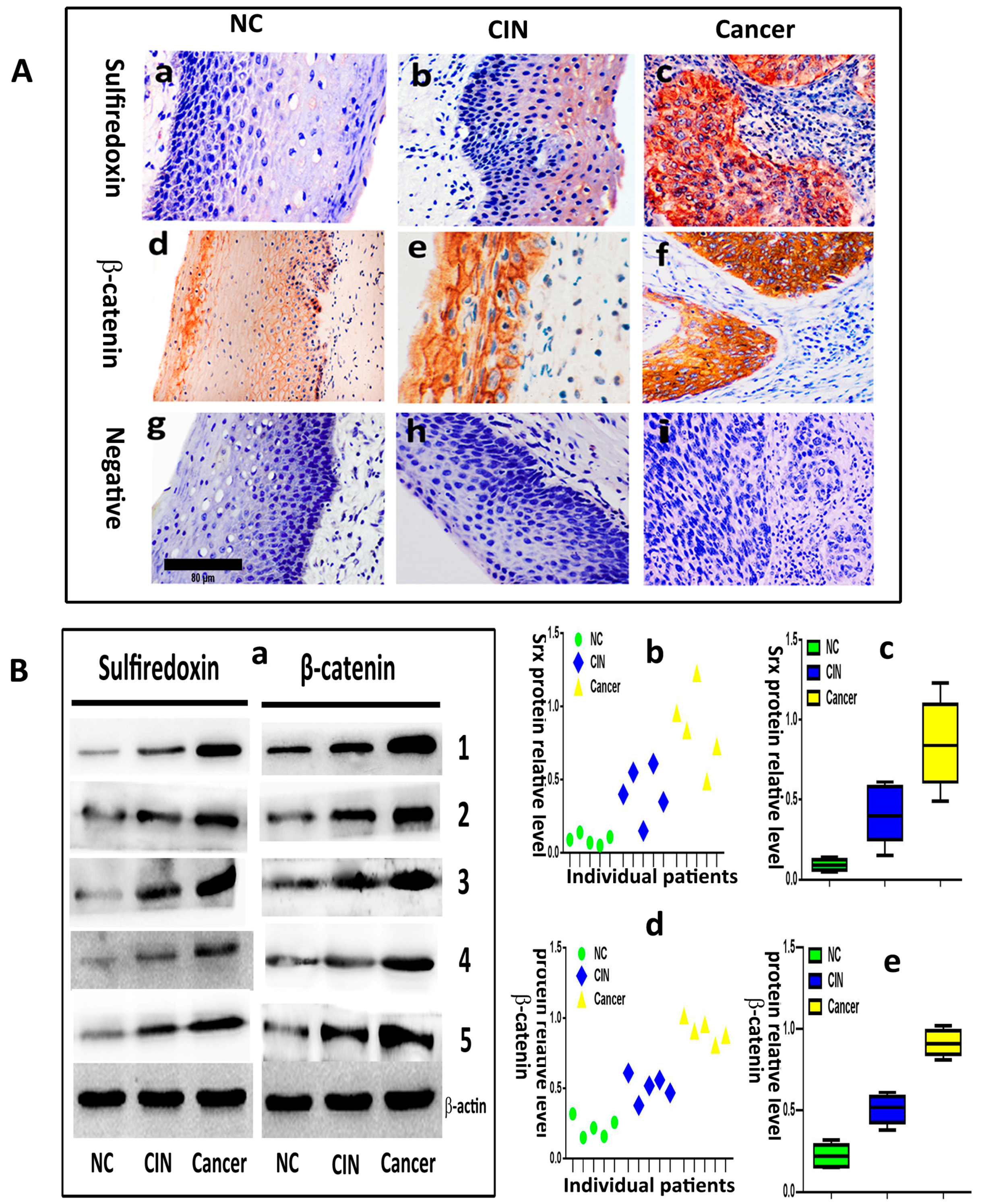
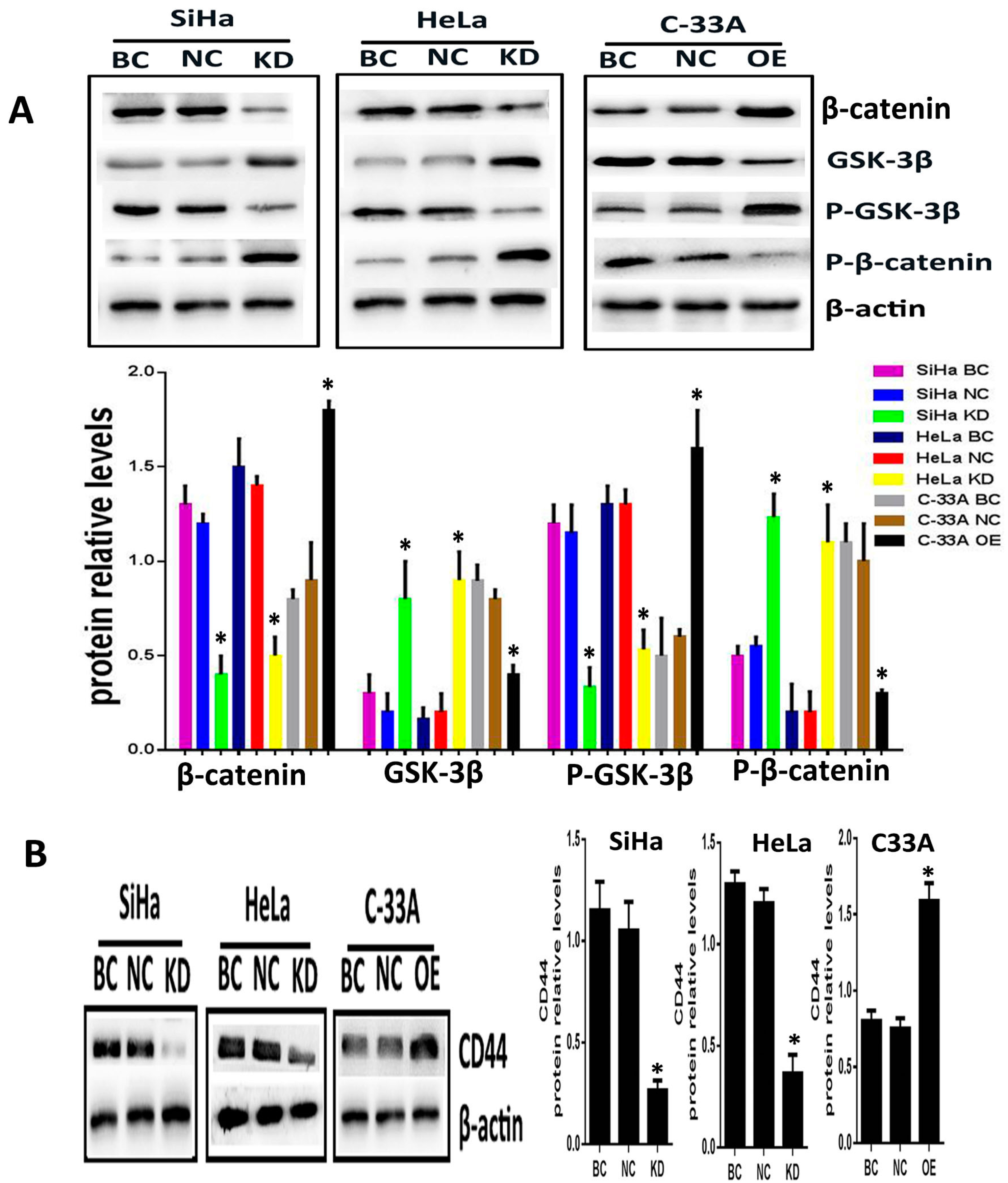
2.2. Silencing or Overexpression of Srx Resulted in Alteration of Proteins Levels Associated with Wnt/β-Catenin Signaling Pathway
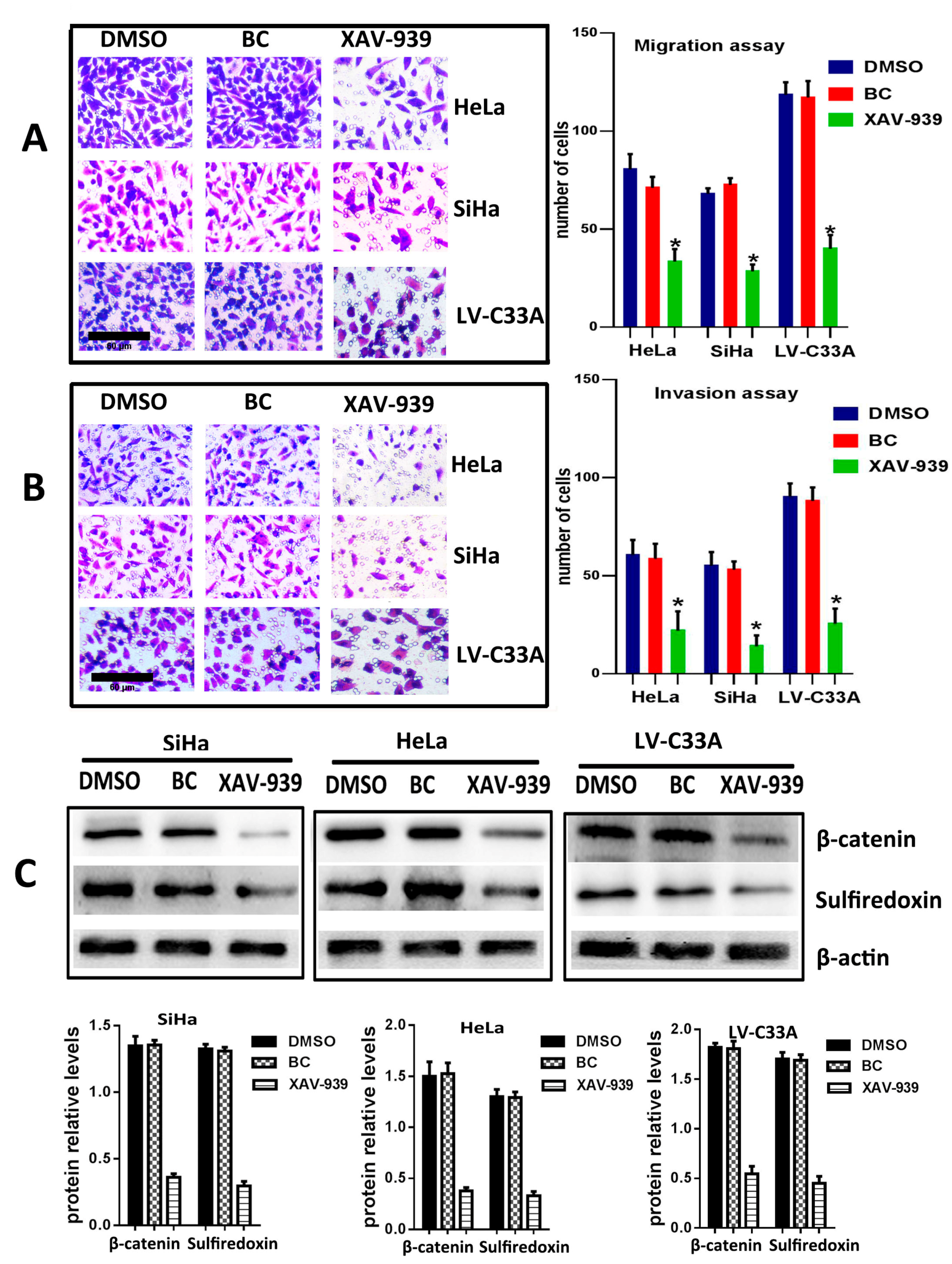
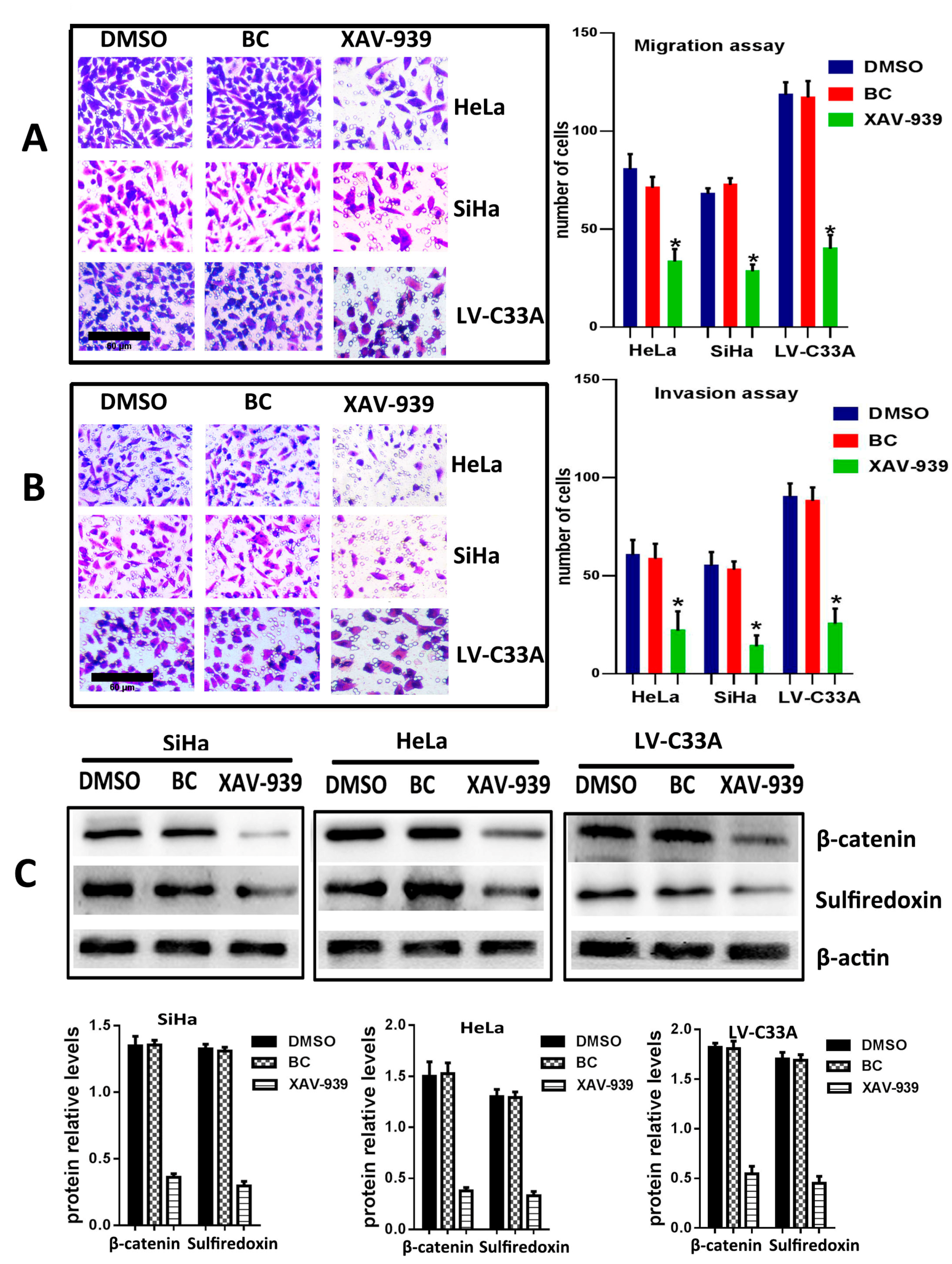
2.3. The Suppression of Wnt/β-Catenin Signaling Pathway by XAV-939 Inhibits Migration, Invasion and Srx Expression in Cervical Cancer Cell Lines
3. Discussion
4. Methods and Materials
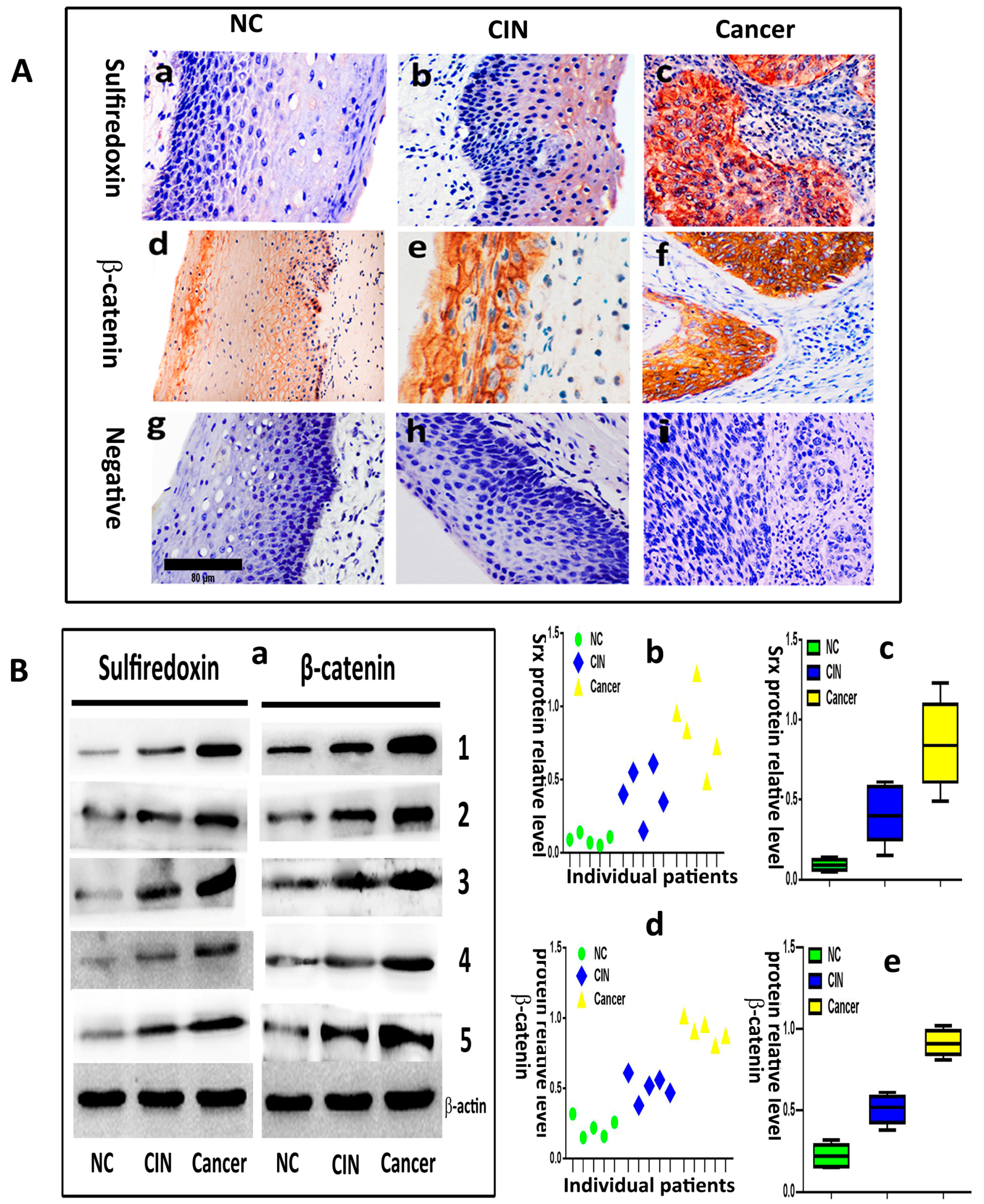
4.1. Clinical Patient Specimens
4.2. Cell Lines and Reagents
4.3. Immunohistochemistry (IHC)
4.4. Lentiviral Transduction and Establishment of Stable Cell Lines
4.5. RT-qPCR
4.6. Western Blot Analysis (WB)
4.7. Cell Migration and Invasion Assay
4.8. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fei, L.; Wang, T.; Tang, S. SOX14 promotes proliferation and invasion of cervical cancer cells through Wnt/β-catenin pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 1698–1704. [Google Scholar]
- Qian, C.; Zheng, P.S.; Yang, W.T. EZH2-mediated repression of GSK-3β and TP53 promotes Wnt/β-catenin signaling-dependent cell expansion in cervical carcinoma. Oncotarget 2016, 7, 36115–36129. [Google Scholar]
- Cheng, Y.; Guo, Y.; Zhang, Y.; You, K.; Li, Z.; Geng, L. MicroRNA-106b is involved in transforming growth factor β1-induced cell migration by targeting disabled homolog 2 in cervical carcinoma. J. Exp. Clin. Cancer Res. 2016, 35, 36116–36129. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.Y.; Han, S.H.; Sung, S.H.; Lee, H.E.; Kim, Y.M.; Noh, Y.H.; Bae, S.H.; Rhee, S.G.; Chang, T.S. Sulfiredoxin protein is critical for redox balance and survival of cells exposed to low steady-state levels of H2O2. J. Biol. Chem. 2012, 287, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Findlay, V.J.; Tapiero, H.; Townsend, D.M. Sulfiredoxin: A potential therapeutic agent? Biomed. Pharmacother. 2005, 59, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Bae, S.H.; Toledano, M.B.; Rhee, S.G. Role of sulfiredoxin as a regulator of peroxiredoxin function and regulation of its expression. Free Radic. Biol. Med. 2012, 53, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Park, S.J.; Chang, T.S.; Lee, D.Y.; Rhee, S.G. Molecular mechanism of the reduction of cysteine sulfinic acid of peroxiredoxin to cysteine by mammalian sulfiredoxin. J. Biol. Chem. 2006, 281, 14400–14407. [Google Scholar] [CrossRef] [PubMed]
- Biteau, B.; Labarre, J.; Toledano, M.B. ATP-dependent reduction of cysteine-sulphinic acid by S. cerevisiae sulphiredoxin. Nature 2003, 425, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yu, S.; Wu, J.; Zou, Y.Y.; Zhao, Y. Sulfiredoxin-1 protects PC12 cells against oxidative stress induced by hydrogen peroxide. J. Neurosci. Res. 2013, 91, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M.; Jiang, H.; Wua, L.; Chawsheena, H.A.; Wei, Q. The sulfiredoxin–peroxiredoxin (Srx–Prx) axis in cell signal transduction and cancer development. Cancer Lett. 2015, 366, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, G.R.; Kim, J.; Baek, J.Y.; Jo, Y.J.; Hong, S.E.; Kim, S.H.; Lee, J.; Lee, H.I.; Park, S.K.; et al. Sulfiredoxin inhibitor induces preferential death of cancer cells through reactive oxygen species-mediated mitochondrial damage. Free Radic. Biol. Med. 2016, 91, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wu, L.; Chen, J.; Mishra, M.; Chwsheen, H.A.; Zhu, H.; Wei, Q. Sulfiredoxin promotes colorectal cancer cell invasion and metastasis through a novel mechanism of enhancing EGFR signaling. Mol. Cancer Res. 2015, 13, 1554–1566. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Jiang, H.; Chwsheen, H.A.; Mishra, M.; Young, M.R.; Gerard, M.; Toledano, M.B.; Colburn, N.H.; Wei, Q. Tumor promoter-induced sulfiredoxin is required for mouse skin tumorigenesis. Carcinogenesis 2014, 35, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Lee, H.L.; Lee, K.B.; Park, J.H.; Chung, W.Y.; Lee, K.S.; Sheen, S.S.; Park, K.J.; Hwang, S.C. Nuclear factor E2-related factor 2 dependent overexpression of sulfiredoxin and peroxiredoxin III in human lung cancer. Korean J. Intern. Med. 2011, 26, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.X.; Qin, L.; Lan, K.Y.; Wang, S.; Wang, H. The expression and clinical significance of Srx- and E-cadherin in cervical squamous cell carcinoma tissue. Immunol. J. 2015, 31, 1067–1071. [Google Scholar]
- Bowers, R.R.; Manevich, Y.; Townsend, D.M.; Tew, K.D. Sulfiredoxin redox-sensitive interaction with S100A4 and non-muscle myosin IIA regulates cancer cell motility. Biochemistry 2012, 51, 7740–7754. [Google Scholar] [CrossRef] [PubMed]
- Rath, G.; Jawanjal, P.; Salhan, S.; Nalliah, M.; Dhawan, I. Clinical significance of inactivated glycogen synthase kinase 3β in HPV-associated cervical cancer: Relationship with Wnt/β-catenin pathway activation. Am. J. Reprod. Immunol. 2015, 73, 460–478. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, N.; Zhang, Y.; Wang, S.; Pang, X.; Zhang, S. Wnt-11 overexpression promoting the invasion of cervical cancer cells. Tumour Biol. 2016, 37, 11789–11798. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Huang, Y.; Cao, X.; Xu, J.; Zhang, L.; Wang, J.; Huang, L.; Huang, S.; Yuan, L.; Jia, W.; et al. Wnt2 promotes cervical carcinoma metastasis and induction of epithelial-mesenchymal transition. PLoS ONE 2016, 11, e0160414. [Google Scholar] [CrossRef] [PubMed]
- Puerto-Galan, L.; Perez-Ruiz, J.M.; Guinea, M.; Cejudo, F.J. The contribution of NADPH thioredoxin reductase C (NTRC) and sulfiredoxin to 2-Cys peroxiredoxin overoxidation in arabidopsis thaliana chloroplasts. J. Exp. Bot. 2015, 66, 2957–2966. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Fu, Z.; Wang, H.; Feng, J.; Wei, J.; Guo, J. Peroxiredoxin 2 knockdown by RNA interference inhibits the growth of colorectal cancer cells by downregulating Wnt/β-catenin signaling. Cancer Lett. 2014, 343, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Jiang, H.; Baker, A.; Dodge, L.K.; Gerard, M.; Young, M.R.; Toledano, M.B.; Colburn, N.H. Loss of sulfiredoxin renders mice resistant to azoxymethane/dextran sulfate sodium-induced colon carcinogenesis. Carcinogenesis 2013, 34, 1403–1410. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Jiang, H.; Matthews, C.P.; Colburn, N.H. Sulfiredoxin is an AP-1 target gene that is required for transformation and shows elevated expression in human skin malignancies. Proc. Natl. Acad. Sci. USA 2008, 105, 19738–19743. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Jiang, H.; Xiao, Z.; Baker, A.; Young, M.R.; Veenstra, T.D.; Colburn, N.H. Sulfiredoxin–peroxiredoxin IV axis promotes human lung cancer progression through modulation of specific phosphokinase signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 7004–7009. [Google Scholar] [CrossRef] [PubMed]
- Maryam, K.; Mohammed, C.S.; Wang, J.; Wei, Q.; Wang, X.; Collier, Z.; Tang, S.; Liu, H.; Zhang, F.; Huang, J.; et al. Wnt/β-catenin signaling plays an ever-expanding role in stem cell self-renewal, tumorigenesis and cancer chemoresistance. Genes Dis. 2016, 3, 11–40. [Google Scholar]
- Aminuddin, A.; Ng, P.Y. Promising druggable target in head and neck squamous cell carcinoma: Wnt signaling. Front. Pharmacol. 2016, 7, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Zhang, S.; Wang, P.; Yang, C.; Shang, C.; Yang, J.; Wang, J. LncRNA, TUG1 regulates the oral squamous cell carcinoma progression possibly via interacting with Wnt/β-catenin signaling. Gene 2017, 608, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Liang, L.; Ouyang, K.; Li, Z.; Yi, X. MALAT1 induces tongue cancer cells’ EMT and inhibits apoptosis through Wnt/β-catenin signaling pathway. J. Oral Pathol. Med. 2017, 46, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Gang, W.H.; Fu, Y.S.; Zhou, K.M.; Guo, Y.; Yuan, M.; Dong, H.J. The expression of β-catenin and galectin-3 in cervical carcinoma and its clinical pathological significances. West. China Med. J. 2015, 30, 1452–1456. [Google Scholar]
- Moon, J.C.; Kim, G.M.; Kim, E.K.; Lee, H.N.; Ha, B.; Lee, S.Y.; Jiang, H.H. Reversal of 2-Cys peroxiredoxin oligomerization by sulfiredoxin. Biochem. Biophys. Res. Commun. 2013, 432, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Noh, Y.H.; Baek, J.Y.; Jeong, W.; Rhee, S.G.; Chang, T.S. Sulfiredoxin translocation into mitochondria plays a crucial role in reducing hyperoxidized peroxiredoxin III. J. Biol. Chem. 2009, 284, 8470–8477. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Case | Expression of Srx | Expression of β-Catenin | ||||||
|---|---|---|---|---|---|---|---|---|---|
| + (%) | − (%) | χ2 | p | + (%) | − (%) | χ2 | p | ||
| NC | 20 | 3(15.0) | 17(85.0) | 4(20) | 16(80.0) | ||||
| CIN | 30 | 14(46.7) | 16(53.3) | 16(53.3) | 14(46.7) | ||||
| CINI | 5 | 1(20.0) | 4(80.0) | 1(20.0) | 3(80.0) | ||||
| CINII | 12 | 5(41.7) | 7(58.3) | 6(50.0) | 6(50.0) | ||||
| CINIII | 13 | 8(61.5) | 5(38.5) | 9(69.2) | 4(30.8) | ||||
| CSCC | 90 | 66(73.3) | 22(26.7) | 71(78.9) | 19(21.1) | ||||
| NC vs. CIN | 3.980 | 0.021 | 4.253 | 0.018 | |||||
| CINvs. Cancer | 7.200 | 0.013 | 7.370 | 0.010 | |||||
| NC vs. Cancer | 21.386 | 0.000 | 23.514 | 0.000 | |||||
| NC vs. CIN vs. Cancer | 26.476 | 0.000 | 27.037 | 0.000 | |||||
| Variable | N | Expression of Srx | Expression of β-Catenin | ||||
|---|---|---|---|---|---|---|---|
| + (%) | − (%) | p | + (%) | − (%) | p | ||
| Age (year) | |||||||
| ≥45 | 42 | 33(78.6) | 9(21.4) | 0.209 | 36(85.7) | 6(14.3) | 0.110 |
| <45 | 48 | 33(68.7) | 15(31.3) | 35(72.9) | 13(27.1) | ||
| FIGO stage | |||||||
| I | 70 | 49(70.0) | 21(30.0) | 0.146 | 52(74.3) | 18(25.7) | 0.037 |
| II | 20 | 17(85.0) | 3(15.0) | 19(95.0) | 1(5.0) | ||
| Tumor size (cm) | |||||||
| ≥4 | 10 | 6(60.0) | 4(40.0) | 0.255 | 6(60.0) | 4(40.0) | 0.129 |
| <4 | 80 | 60(75.0) | 20(25.0) | 65(81.2) | 15(18.8) | ||
| Histologic differentiation | |||||||
| Well and moderate | 63 | 44(69.8) | 19(30.2) | 0.190 | 46(73.0) | 17(27.0) | 0.030 |
| Poor | 27 | 22(81.5) | 5(18.5) | 25(92.6) | 2(7.4) | ||
| Lymph node metastasis | |||||||
| + | 27 | 24(88.9) | 3(11.1) | 0.023 | 25(92.6) | 2(7.4) | 0.030 |
| − | 63 | 42(66.7) | 21(33.3) | 46(73.0) | 17(27.0) | ||
| Infiltration of haemal tube | |||||||
| + | 21 | 19(90.5) | 2(9.5) | 0.034 | 15(71.4) | 6(28.6) | 0.252 |
| − | 69 | 47(68.1) | 22(31.9) | 56(81.2) | 13(18.8) | ||
| Depth of cancer invasion | |||||||
| Light layer | 54 | 36(66.7) | 18(33.3) | 0.064 | 41(75.9) | 13(24.1) | 0.284 |
| Deep layer | 36 | 30(83.3) | 6(6.7) | 30(83.3) | 16(16.7) | ||
| Expression of β-Catenin | Expression of Srx | |
|---|---|---|
| Positive (n = 66) | Negative (n = 24) | |
| Positive (n = 71) | 58 | 13 |
| Negative (n = 19) | 8 | 11 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, K.; Zhao, Y.; Fan, Y.; Ma, B.; Yang, S.; Liu, Q.; Linghu, H.; Wang, H. Sulfiredoxin May Promote Cervical Cancer Metastasis via Wnt/β-Catenin Signaling Pathway. Int. J. Mol. Sci. 2017, 18, 917. https://doi.org/10.3390/ijms18050917
Lan K, Zhao Y, Fan Y, Ma B, Yang S, Liu Q, Linghu H, Wang H. Sulfiredoxin May Promote Cervical Cancer Metastasis via Wnt/β-Catenin Signaling Pathway. International Journal of Molecular Sciences. 2017; 18(5):917. https://doi.org/10.3390/ijms18050917
Chicago/Turabian StyleLan, Kangyun, Yuni Zhao, Yue Fan, Binbin Ma, Shanshan Yang, Qin Liu, Hua Linghu, and Hui Wang. 2017. "Sulfiredoxin May Promote Cervical Cancer Metastasis via Wnt/β-Catenin Signaling Pathway" International Journal of Molecular Sciences 18, no. 5: 917. https://doi.org/10.3390/ijms18050917
APA StyleLan, K., Zhao, Y., Fan, Y., Ma, B., Yang, S., Liu, Q., Linghu, H., & Wang, H. (2017). Sulfiredoxin May Promote Cervical Cancer Metastasis via Wnt/β-Catenin Signaling Pathway. International Journal of Molecular Sciences, 18(5), 917. https://doi.org/10.3390/ijms18050917