
Health Effects of Psidium guajava L. Leaves: An Overview of the Last Decade


 ,
, 

Abstract
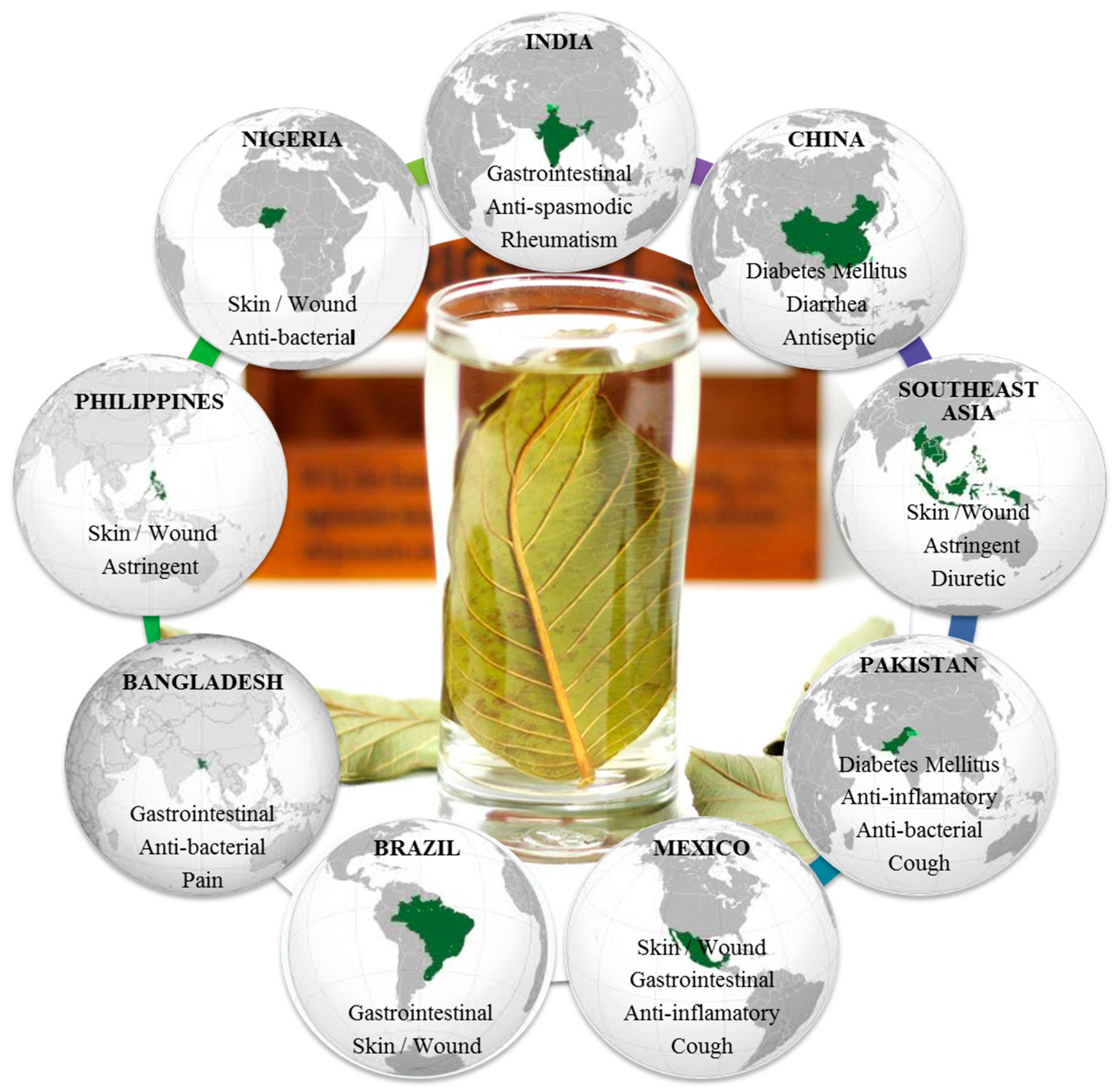
1. Introduction
2. Pharmacological Properties
2.1. In Vitro Studies
2.1.1. Infectious and Parasitic Diseases
2.1.2. Neoplasms
2.1.3. Diseases of the Blood and Immune System
2.1.4. Endocrine and Metabolic Diseases
2.1.5. Diseases of the Circulatory System
2.1.6. Diseases of the Digestive System
2.1.7. Diseases of the Skin and Subcutaneous Tissue
2.1.8. Other Activities Related to Several Diseases
2.2. In Vivo Studies
2.2.1. Infectious and Parasitic Diseases
2.2.2. Neoplasms
2.2.3. Diseases of the Blood and Immune System
2.2.4. Endocrine and Metabolic Diseases
2.2.5. Diseases of the Circulatory System
2.2.6. Diseases of the Digestive System
2.2.7. Diseases of the Skin and Subcutaneous Tissue
2.2.8. Other Activities Related to Several Diseases
2.3. Clinical Trials
3. Other Applications
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Anyinam, C. Ecology and ethnomedicine: Exploring links between current environmental crisis and indigenous medical practices. Soc. Sci. Med. 1995, 40, 321–329. [Google Scholar] [CrossRef]
- Patwardhan, B.; Warude, D.; Pushpangadan, P.; Bhatt, N. Ayurveda and traditional Chinese medicine: A comparative overview. Evid. Based Complement. Altern. Med. 2005, 2, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Fabricant, D.S.; Farnsworth, N.R. The value of plants used in traditional medicine for drug discovery. Environ. Health Perspect. 2001, 109, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.F. Fruits of Warm Climates; Creative Resource Systems, Inc.: Winterville, NC, USA, 1987. [Google Scholar]
- Salazar, D.M.; Melgarejo, P.; Martínez, R.; Martínez, J.J.; Hernández, F.; Burguera, M. Phenological stages of the guava tree (Psidium guajava L.). Sci. Hortic. 2006, 108, 157–161. [Google Scholar] [CrossRef]
- Gutiérrez, R.M.P.; Mitchell, S.; Solis, R.V. Psidium guajava: A review of its traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2008, 117, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Shruthi, S.D.; Roshan, A.; Sharma, S.; Sunita, S. A review on the medicinal plant Psidium guajava Linn. (Myrtaceae). J. Drug Deliv. Ther. 2013, 3, 162–168. [Google Scholar]
- Morais-Braga, M.F.B.; Carneiro, J.N.P.; Machado, A.J.T.; dos Santos, A.T.L.; Sales, D.L.; Lima, L.F.; Figueredo, F.G.; Coutinho, H.D.M. Psidium guajava L., from ethnobiology to scientific evaluation: Elucidating bioactivity against pathogenic microorganisms. J. Ethnopharmacol. 2016, 194, 1140–1152. [Google Scholar] [CrossRef] [PubMed]
- Sanda, K.A.; Grema, H.A.; Geidam, Y.A.; Bukar-Kolo, Y.M. Pharmacological aspects of P. guajava: An update. Int. J. Pharmacol. 2011, 7, 316–324. [Google Scholar]
- Bernal, J.; Mendiola, J.A.; Ibáñez, E.; Cifuentes, A. Advanced analysis of nutraceuticals. J. Pharm. Biomed. Anal. 2011, 55, 758–774. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Nishimura, Y.; Kurata, N.; Iwase, M.; Yasuhara, H. Effects of continuous ingestion of herbal teas on intestinal CYP3A in the rat. J. Pharmacol. Sci. 2007, 103, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Yasuoka, A.; Abe, K. Functional food science and food for specified health use policy in Japan: State of the art. Curr. Opin. Lipidol. 2008, 19, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Milyani, R. Inhibitory effect of some plant extracts on clinical isolates of Staphylococcus aureus. Afr. J. Microbiol. Res. 2012, 6, 6517–6524. [Google Scholar] [CrossRef]
- Anas, K.; Jayasree, P.R.; Vijayakumar, T.; Manish Kumar, P.R. In vitro antibacterial activity of Psidium guajava Linn. leaf extract on clinical isolates of multidrug resistant Staphylococcus aureus. Indian J. Exp. Biol. 2008, 46, 41–46. [Google Scholar] [PubMed]
- Chah, K.F.; Eze, C.A.; Emuelosi, C.E.; Esimone, C.O. Antibacterial and wound healing properties of methanolic extracts of some Nigerian medicinal plants. J. Ethnopharmacol. 2006, 104, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.; Chanda, S. In Vitro antimicrobial activity of Psidium guajava L. leaf extracts against clinically important pathogenic microbial strains. Braz. J. Microbiol. 2007, 38, 452–458. [Google Scholar] [CrossRef]
- Dhiman, A.; Nanda, A.; Narasimhan, B. In vitro antimicrobial activity of methanolic leaf extract of Psidium guajava L. J. Pharm. Bioallied Sci. 2011, 3, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.R.V.; Dias, A.L.T.; Carvalho, R.R.; Souza, C.R.F.; Oliveira, W.P. Antioxidant and antimicrobial activities of Psidium guajava L. spray dried extracts. Ind. Crops Prod. 2014, 60, 39–44. [Google Scholar] [CrossRef]
- De Araújo, A.A.; Soares, L.A.L.; Assunção Ferreira, M.R.; de Souza Neto, M.A.; da Silva, G.R.; de Araújo, R.F.; Guerra, G.C.B.; de Melo, M.C.N. Quantification of polyphenols and evaluation of antimicrobial, analgesic and anti-inflammatory activities of aqueous and acetone-water extracts of Libidibia ferrea, Parapiptadenia rigida and Psidium guajava. J. Ethnopharmacol. 2014, 156, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Nisha, K.; Darshana, M.; Madhu, G.; Bhupendra, M.K. GC-MS Analysis and anti-microbial activity of Psidium guajava (leaves) grown in Malva region of India. Int. J. Drug Dev. Res. 2011, 3, 237–245. [Google Scholar]
- Bezerra Morais-Braga, M.F.; Lima Sales, D.; dos Santos Silva, F.; Pereira Chaves, T.; de Carvalho Nilo Bitu, V.; Torres Avilez, W.M.; Ribeiro-Filho, J.; Douglas Melo Coutinho, H. Psidium guajava L. and Psidium brownianum Mart ex DC. potentiate the effect of antibiotics against Gram-positive and Gram-negative bacteria. Eur. J. Integr. Med. 2016, 8, 683–687. [Google Scholar] [CrossRef]
- Betoni, J.E.C.; Passarelli Mantovani, R.; Nunes Barbosa, L.; Di Stasi, L.C.; Fernandes Junior, A. Synergism between plant extract and antimicrobial drugs used on Staphylococcus aureus diseases. Mem. Inst. Oswaldo Cruz 2006, 101, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Metwally, A.M.; Omar, A.A.; Harraz, F.M.; El Sohafy, S.M. Phytochemical investigation and antimicrobial activity of Psidium guajava L. leaves. Pharmacogn. Mag. 2010, 6, 212–218. [Google Scholar] [PubMed]
- Mailoa, M.N.; Mahendradatta, M.; Laga, A.; Djide, N. Antimicrobial activities of tannins extract from guava leaves (Psidium guajava L.) on pathogens microbial. Int. J. Sci. Technol. Res. 2014, 3, 236–241. [Google Scholar]
- Ghosh, P.; Mandal, A.; Chakraborty, P.; Rasul, M.G.; Chakraborty, M.; Saha, A. Triterpenoids from Psidium guajava with biocidal activity. Indian J. Pharm. Sci. 2010, 72, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Zahir, A.A.; Rahuman, A.A.; Bagavan, A.; Santhoshkumar, T.; Mohamed, R.R.; Kamaraj, C.; Rajakumar, G.; Elango, G.; Jayaseelan, C.; Marimuthu, S. Evaluation of botanical extracts against Haemaphysalis bispinosa Neumann and Hippobosca maculata Leach. Parasitol. Res. 2010, 107, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.S.; Sykes, M.L.; Akanji, M.A.; Avery, V.M. Anti-trypanosomal and cytotoxic activity of ethanolic extracts of Psidium guajava leaves in Alamar Blue based assays. Vet. Arh. 2011, 81, 623–633. [Google Scholar]
- Kaushik, N.K.; Bagavan, A.; Rahuman, A.A.; Zahir, A.A.; Kamaraj, C.; Elango, G.; Jayaseelan, C.; Kirthi, A.V.; Santhoshkumar, T.; Marimuthu, S.; et al. Evaluation of antiplasmodial activity of medicinal plants from North Indian Buchpora and South Indian Eastern Ghats. Malar. J. 2015, 14, 65. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.C.; Mahmud, R.; Noordin, R.; Pillai Piaru, S.; Perumal, S.; Ismail, S. Free radicals scavenging activity, cytotoxicity and anti-parasitic activity of essential oil of Psidium guajava L. leaves against Toxoplasma gondii. J. Essent. Oil Bear. Plants 2013, 16, 32–38. [Google Scholar] [CrossRef]
- Chanu, T.R.; Pai, V.; Chakraborty, R.; Raju, B.; Lobo, R.; Ballal, M. Screening for antidiarrheal activity of Psidium guajava: A possible alternative in the treatment against diarrhea causing enteric pathogens. J. Chem. Pharm. Res. 2011, 3, 961–967. [Google Scholar]
- Thiyagarajan, S.; Jamal, A. Evaluation of Lethal Activity of Psidium guajava Linn. extracts on bacterial pathogens causing diarrheal infections. Int. J. Res. Ayurveda Pharm. 2015, 6, 111–117. [Google Scholar] [CrossRef]
- Rahim, N.; Gomes, D.J.; Watanabe, H.; Rahman, S.R.; Chomvarin, C.; Endtz, H.P.; Alam, M. Antibacterial activity of Psidium guajava leaf and bark against multidrug-resistant Vibrio cholerae: Implication for cholera control. Jpn. J. Infect. Dis. 2010, 63, 271–274. [Google Scholar] [PubMed]
- Birdi, T.; Daswani, P.; Brijesh, S.; Tetali, P.; Natu, A.; Antia, N. Newer insights into the mechanism of action of Psidium guajava L. leaves in infectious diarrhoea. BMC Complement. Altern. Med. 2010, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.A.; Andrade Neto, M.; Bezerra, J.N.S.; Macrae, A.; De Sousa, O.V.; Fonteles-Filho, A.A.; Vieira, R.H.S.D.F. Antibacterial activity of guava, Psidium guajava Linnaeus, leaf extracts on diarrhea-causing enteric bacteria isolated from seabob shrimp, Xiphopenaeus kroyeri (Heller). Rev. Inst. Med. Trop. Sao Paulo 2008, 50, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Nwinyi, O.; Chinedu, S.N.; Ajani, O.O. Evaluation of antibacterial activity of Pisidium guajava and Gongronema latifolium. J. Med. Plants Res. 2008, 2, 189–192. [Google Scholar]
- Sriwilaijaroen, N.; Fukumoto, S.; Kumagai, K.; Hiramatsu, H.; Odagiri, T.; Tashiro, M.; Suzuki, Y. Antiviral effects of Psidium guajava Linn. (guava) tea on the growth of clinical isolated H1N1 viruses: Its role in viral hemagglutination and neuraminidase inhibition. Antivir. Res. 2012, 94, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Nakamura, T.; Hosokawa, T.; Suzuki-Yamamoto, T.; Yamashita, H.; Kimoto, M.; Tsuji, H.; Yoshida, H.; Hada, T.; Takahashi, Y. Antiproliferative activity of guava leaf extract via inhibition of prostaglandin endoperoxide H synthase isoforms. Prostaglandins Leukot. Essent. Fatty Acids 2009, 80, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Sul´ain, M.D.; Zazali, K.E.; Ahmad, N. Screening on anti-proliferative activity of Psidium guajava Leaves extract towards selected cancer cell lines. J. US China Med. Sci. 2012, 9, 30–37. [Google Scholar]
- Vieira Braga, T.; Gonçalves Rodrigues das Dores, R.; Soncin Ramos, C.; Gontijo Evangelista, F.C.; Márcia da Silva Tinoco, L.; de Pilla Varotti, F.; Carvalho, M.D.G.; de Paula Sabino, A. Antioxidant, antibacterial and antitumor activity of ethanolic extract of the Psidium guajava leaves. Am. J. Plant Sci. 2014, 5, 3492–3500. [Google Scholar] [CrossRef]
- Kaileh, M.; Vanden Berghe, W.; Boone, E.; Essawi, T.; Haegeman, G. Screening of indigenous Palestinian medicinal plants for potential anti-inflammatory and cytotoxic activity. J. Ethnopharmacol. 2007, 113, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.C.; Peng, C.H.; Chen, K.C.; Hsieh, C.L.; Peng, R.Y. The aqueous soluble polyphenolic fraction of Psidium guajava leaves exhibits potent anti-angiogenesis and anti-migration actions on DU145 cells. Evid. Based Complement. Altern. Med. 2011, 2011, 219069. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.C.; Hsieh, C.L.; Huang, K.D.; Ker, Y.B.; Chyau, C.C.; Peng, R.Y. Anticancer activity of rhamnoallosan against DU-145 cells is kinetically complementary to coexisting polyphenolics in Psidium guajava budding leaves. J. Agric. Food Chem. 2009, 57, 6114–6122. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L.Y.; Longato, G.B.; Ruiz, A.L.T.G.; Tinti, S.V.; Possenti, A.; Vendramini-costa, D.B. In vitro, in vivo and in silico analysis of the anticancer and estrogen-like activity of guava leaf extracts. Curr. Med. Chem. 2014, 21, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Park, K.R.; Nam, D.; Yun, H.M.; Lee, S.G.; Jang, H.J.; Sethi, G.; Cho, S.K.; Ahn, K.S. β-Caryophyllene oxide inhibits growth and induces apoptosis through the suppression of PI3K/AKT/mTOR/S6K1 pathways and ROS-mediated MAPKs activation. Cancer Lett. 2011, 312, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Manosroi, J.; Dhumtanom, P.; Manosroi, A. Anti-proliferative activity of essential oil extracted from Thai medicinal plants on KB and P388 cell lines. Cancer Lett. 2006, 235, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.S.; Carley, S. Cytotoxic activity of hexane extracts of Psidium guajava L. (myrtaceae) and Cassia alata L. (caesalpineaceae) in kasumi-1 and OV2008 cancer cell lines. Trop. J. Pharm. Res. 2012, 11, 201–207. [Google Scholar] [CrossRef]
- Ryu, N.H.; Park, K.R.; Kim, S.M.; Yun, H.M.; Nam, D.; Lee, S.G.; Jang, H.J.; Ahn, K.S.; Kim, S.-H.; Shim, B.S.; et al. A hexane fraction of guava leaves (Psidium guajava L.) induces anticancer activity by suppressing AKT/mammalian target of rapamycin/ribosomal p70 S6 kinase in human prostate cancer cells. J. Med. Food 2011, 15, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Hwang, J.H.; Park, S.Y.; Jin, Y.J.; Ko, H.C.; Moon, S.W.; Kim, S.J. Fermented guava leaf extract inhibits LPS-induced COX-2 and iNOS expression in Mouse macrophage cells by inhibition of transcription factor NF-kappaB. Phyther. Res. 2008, 22, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Jeong, S.W.; Cho, S.K.; Ahn, K.S.; Kim, B.K.; Kim, J.C. Anti-inflammatory effects of 4 medicinal plant extracts in lipopolysaccharide-induced RAW 264.7 cells. Food Sci. Biotechnol. 2013, 22, 213–220. [Google Scholar] [CrossRef]
- Jang, M.; Jeong, S.-W.; Cho, S.K.; Ahn, K.S.; Lee, J.H.; Yang, D.C.; Kim, J.-C. Anti-Inflammatory Effects of an Ethanolic Extract of Guava (Psidium guajava L.) Leaves In Vitro and In Vivo. J. Med. Food 2014, 17, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.S.; Sukumaran, V.; Giri, S.S.; Park, S.C. Flavonoid fraction of guava leaf extract attenuates lipopolysaccharide-induced inflammatory response via blocking of NF-κB signalling pathway in Labeo rohita macrophages. Fish Shellfish Immunol. 2015, 47, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Jeong, S.-W.; Cho, S.K.; Yang, H.J.; Yoon, D.-S.; Kim, J.-C.; Park, K.-H. Improvement in the anti-inflammatory activity of guava (Psidium guajava L.) leaf extracts through optimization of extraction conditions. J. Funct. Foods 2014, 10, 161–168. [Google Scholar] [CrossRef]
- Madduluri, S.; Sitaram, B. In vitro evaluation of anti inflammatory activity of methanolic and ethanolic leaf extracts of five indigenous plants in South India. Int. J. PharmTech Res. 2014, 6, 569–574. [Google Scholar]
- Laily, N.; Kusumaningtyas, R.W.; Sukarti, I.; Rini, M.R.D.K. The potency of guava Psidium guajava (L.) leaves as a Functional immunostimulatory ingredient. Procedia Chem. 2015, 14, 301–307. [Google Scholar] [CrossRef]
- Soman, S.; Rauf, A.A.; Indira, M.; Rajamanickam, C. Antioxidant and antiglycative potential of ethyl acetate fraction of Psidium guajava leaf extract in streptozotocin-induced diabetic rats. Plant Foods Hum. Nutr. 2010, 65, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-W.; Hsieh, C.-L.; Wang, H.-Y.; Chen, H.-Y. Inhibitory effects of guava (Psidium guajava L.) leaf extracts and its active compounds on the glycation process of protein. Food Chem. 2009, 113, 78–84. [Google Scholar] [CrossRef]
- Wang, H.; Du, Y.-J.; Song, H.-C. α-Glucosidase and α-amylase inhibitory activities of guava leaves. Food Chem. 2010, 123, 6–13. [Google Scholar] [CrossRef]
- Deguchi, Y.; Miyazaki, K. Anti-hyperglycemic and anti-hyperlipidemic effects of guava leaf extract. Nutr. Metab. Lond. 2010, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Kong, F.; Ni, H.; Mo, Z.; Wan, J.B.; Hua, D.; Yan, C. Structural characterization, α-glucosidase inhibitory and DPPH scavenging activities of polysaccharides from guava. Carbohydr. Polym. 2016, 144, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Eidenberger, T.; Selg, M.; Krennhuber, K. Inhibition of dipeptidyl peptidase activity by flavonol glycosides of guava (Psidium guajava L.): A key to the beneficial effects of guava in type II diabetes mellitus. Fitoterapia 2013, 89, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lim, Y.; Kwon, O. Selected phytochemicals and culinary plant extracts inhibit fructose uptake in caco-2 cells. Molecules 2015, 20, 17393–17404. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.-C.; Shen, S.-C.; Wu, J.S.-B. Effect of guava (Psidium guajava L.) leaf extract on glucose uptake in rat hepatocytes. J. Food Sci. 2009, 74, H132–H138. [Google Scholar] [CrossRef] [PubMed]
- Basha, S.K.; Kumari, V.S. In vitro antidiabetic activity of Psidium guajava leaves extracts. Asian Pac. J. Trop. Dis. 2012, 2 (Suppl. S1), 98–100. [Google Scholar] [CrossRef]
- Basha, S.K.; Govindaraju, K.; Manikandan, R.; Ahn, J.S.; Bae, E.Y.; Singaravelu, G. Phytochemical mediated gold nanoparticles and their PTP 1B inhibitory activity. Colloids Surf. B Biointerfaces 2010, 75, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, H.; Qin, L.; Liu, T.; Gao, M. Guava leaf extracts inhibit 3T3-L1 adipocyte differentiation via activating AMPK. J. Nutr. Ther. 2012, 1, 107–113. [Google Scholar]
- Liu, C.-W.; Wang, Y.-C.; Hsieh, C.-C.; Lu, H.-C.; Chiang, W.-D. Guava (Psidium guajava Linn.) leaf extract promotes glucose uptake and glycogen accumulation by modulating the insulin signaling pathway in high-glucose-induced insulin-resistant mouse FL83B cells. Process Biochem. 2015, 50, 1128–1135. [Google Scholar] [CrossRef]
- Chang, W.-C.; Shen, S.-C. Effect of water extracts from edible myrtaceae plants on uptake of 2-(n-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino)-2-deoxyglucose in TNF-α-treated FL83B mouse hepatocytes. Phytother. Res. 2013, 27, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Arasakumari, M.; Prabu, P.; Amalraj, A.J. Anti-diabetic and aldose reductase inhibitory potential of Psidium guajava by in vitro analysis. Int. J. Pharm. Pharm. Sci. 2016, 8, 271–276. [Google Scholar] [CrossRef]
- Owen, P.L.; Matainaho, T.; Sirois, M.; Johns, T. Endothelial cytoprotection from oxidized LDL by some crude melanesian plant extracts is not related to their antioxidant capacity. J. Biochem. Mol. Toxicol. 2007, 21, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.L.; Huang, C.N.; Lin, Y.C.; Peng, R.Y. Molecular action mechanism against apoptosis by aqueous extract from guava budding leaves elucidated with human umbilical vein endothelial cell (HUVEC) model. J. Agric. Food Chem. 2007, 55, 8523–8533. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-L.; Lin, Y.-C.; Yen, G.-C.; Chen, H.-Y. Preventive effects of guava (Psidium guajava L.) leaves and its active compounds against α-dicarbonyl compounds-induced blood coagulation. Food Chem. 2007, 103, 528–535. [Google Scholar] [CrossRef]
- Anyachukwu Irondi, E.; Olalekan Agboola, S.; Oboh, G.; Augusti Boligon, A.; Linde Athayde, M.; Shode, F.O. Guava leaves polyphenolics-rich extract inhibits vital enzymes implicated in gout and hypertension in vitro. J. Intercult. Ethnopharmacol. 2016, 5, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Prabu, G.R.; Gnanamani, A.; Sadulla, S. Guaijaverin—A plant flavonoid as potential antiplaque agent against Streptococcus mutans. J. Appl. Microbiol. 2006, 101, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Shafiei, Z.; Haji Abdul Rahim, Z.; Philip, K.; Thurairajah, N. Antibacterial and anti-adherence effects of a plant extract mixture (PEM) and its individual constituent extracts (Psidium sp., Mangifera sp., and Mentha sp.) on single- and dual-species biofilms. PeerJ 2016, 4, e2519. [Google Scholar] [CrossRef] [PubMed]
- Razak, F.A.; Othman, R.Y.; Rahim, Z.H.A. The effect of Piper betle and Psidium guajava extracts on the cell-surface hydrophobicity of selected early settlers of dental plaque. J. Oral Sci. 2006, 48, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Fathilah, A.R.; Rahim, Z.H.A.; Othman, Y.; Yusoff, M. Bacteriostatic effect of Piper betle and Psidium guajava extracts on dental plaque bacteria. Pak. J. Biol. Sci. 2009, 12, 518–521. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chandrashekar, B.R.; Nagarajappa, R.; Singh, R.; Thakur, R. An in vitro study on the anti-microbial effi cacy of ten herbal extracts on primary plaque colonizers. J. Young Pharm. 2014, 6, 33–39. [Google Scholar] [CrossRef]
- John, N.R.; Gala, V.C.; Sawant, C.S. Inhibitory effects of plant extracts on multi-species dental biofilm formation in vitro. Int. J. Pharm. Bio Sci. 2013, 4, 487–495. [Google Scholar]
- John, N.R.; Gala, V.C.; Bhagwat, A.M.; Sawant, C.S. HPTLC analysis of eclipta prostrata and Psidium guajava extracts and their effect on cell-surface hydrophobicity of a consortium of dental plaque isolates. Int. J. Pharm. Pharm. Sci. 2013, 5, 1–6. [Google Scholar]
- Shekar, C.; Nagarajappa, R.; Singh, R.; Thakur, R. Antimicrobial efficacy of Acacia nilotica, Murraya koenigii L. Sprengel, Eucalyptus hybrid, and Psidium guajava on primary plaque colonizers: An in vitro comparison between hot and cold extraction process. J. Indian Soc. Periodontol. 2015, 19, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Kwamin, F.; Gref, R.; Haubek, D.; Johansson, A. Interactions of extracts from selected chewing stick sources with Aggregatibacter actinomycetemcomitans. BMC Res. Notes 2012, 5, 203. [Google Scholar] [CrossRef] [PubMed]
- Thalikunnil, S.T.; Ashok, A.; Sukesh, K. Screening of psidium gaujava for effective phytomedicines and study on its antibacterial effect against dental caries bacteria. Int. J. Pharm. Pharm. Sci. 2012, 4, 400–401. [Google Scholar]
- Jayakumari, S.; Anbu, J.; Ravichandiran, V.; Nithya, S.; Anjana, A.; Sudharani, D. Evaluation of toothache activity of methanolic extract and its various fractions from the leaves Psidium guajava Linn. Int. J. Pharma. Bio Sci. 2012, 3, 238–249. [Google Scholar]
- Ravi, K.; Divyashree, P. Psidium guajava: A review on its potential as an adjunct in treating periodontal disease. Pharmacogn. Rev. 2014, 8, 96–100. [Google Scholar] [PubMed]
- Chen, H.-H. Hepatoprotective effect of guava (Psidium guajava L.) Leaf extracts on ethanol-induced injury on clone 9 rat liver cells. Food Nutr. Sci. 2011, 2, 983–988. [Google Scholar] [CrossRef][Green Version]
- Qa’dan, F.; Thewaini, A.-J.; Ali, D.A.; Afifi, R.; Elkhawad, A.; Matalka, K.Z. The antimicrobial activities of Psidium guajava and Juglans regia leaf extracts to acne-developing organisms. Am. J. Chin. Med. 2005, 33, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, E.M. The use of Psidium guajava Linn. in treating wound, skin and soft tissue infections. Sci. Res. Essay 2009, 4, 605–611. [Google Scholar]
- Padrón-Márquez, B.; Viveros-Valdez, E.; Oranday-Cárdenas, A.; Carranza-Rosales, P. Antifungal activity of Psidium guajava organic extracts against dermatophytic fungi. J. Med. Plants Res. 2012, 6, 5435–5438. [Google Scholar] [CrossRef]
- Suwanmanee, S.; Kitisin, T.; Luplertlop, N. In vitro screening of 10 edible thai plants for potential antifungal properties. Evid. Based Complement. Altern. Med. 2014, 2014, 138587. [Google Scholar] [CrossRef] [PubMed]
- You, D.-H.; Park, J.-W.; Yuk, H.-G.; Lee, S.-C. Antioxidant and tyrosinase inhibitory activities of different parts of guava (Psidium guajava L.). Food Sci. Biotechnol. 2011, 20, 1095–1100. [Google Scholar] [CrossRef]
- Han, E.H.; Hwang, Y.P.; Choi, J.H.; Yang, J.H.; Seo, J.K.; Chung, Y.C.; Jeong, H.G. Psidium guajava extract inhibits thymus and activation-regulated chemokine (TARC/CCL17) production in human keratinocytes by inducing heme oxygenase-1 and blocking NF-κB and STAT1 activation. Environ. Toxicol. Pharmacol. 2011, 32, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-U.; Yeon Weon, K.; Nam, D.-Y.; Hyun Nam, J.; Kyung Kim, W. Skin protective effect of guava leaves against UV-induced melanogenesis via inhibition of ORAI1 channel and tyrosinase activity. Exp. Dermatol. 2016, 25, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Abreu, P.R.C.; Almeida, M.C.; Bernardo, R.M.; Bernardo, L.C.; Brito, L.C.; Garcia, E.A.C.; Fonseca, A.S.; Bernardo-Filho, M. Guava extract (Psidium guajava) alters the labelling of blood constituents with technetium-99m. J. Zhejiang Univ. Sci. B 2006, 7, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Han, E.H.; Hwang, Y.P.; Kim, H.G.; Park, J.H.; Choi, J.H.; Im, J.H.; Khanal, T.; Park, B.H.; Yang, J.H.; Choi, J.M.; et al. Ethyl acetate extract of Psidium guajava inhibits IgE-mediated allergic responses by blocking FcεRI signaling. Food Chem. Toxicol. 2011, 49, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Pachanawan, A.; Phumkhachorn, P.; Rattanachaikunsopon, P. Potential of Psidium guajava supplemented fish diets in controlling Aeromonas hydrophila infection in tilapia (Oreochromis niloticus). J. Biosci. Bioeng. 2008, 106, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.L.; Li, Z.J.; Yang, K.; Lin, H.Z.; Guo, Z.X. Effect of guava leaves on growth and the non-specific immune response of Penaeus monodon. Fish Shellfish Immunol. 2014, 40, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, S.O.; Akanji, M.A.; Oguntoye, S.A. Ethanolic leaf extract of Psidium guajava: Phyto-chemical and trypanocidal activity in rats infected with Trypanosoma brucei brucei. J. Med. Plants Res. 2009, 3, 420–423. [Google Scholar]
- Akanji, M.A.; Adeyemi, O.S.; Oguntoye, S.O.; Sulyman, F. Psidium guajava extract reduces trypanosomosis associated lipid peroxidation and raises glutathione concentrations in infected animals. EXCLI J. 2009, 8, 148–154. [Google Scholar]
- Rajendran, C.; Begam, M.; Kumar, D.; Baruah, I.; Gogoi, H.K.; Srivastava, R.B.; Veer, V. Antiplasmodial activity of certain medicinal plants against chloroquine resistant Plasmodium berghei infected white albino BALB/c mice. J. Parasit. Dis. 2014, 38, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Birdi, T. Psidium guajava leaf extract prevents intestinal colonization of Citrobacter rodentium in the mouse model. J. Ayurveda Integr. Med. 2015, 6, 50. [Google Scholar] [PubMed]
- Geidam, Y.A.; Ambali, A.G.; Onyeyili, P.A.; Tijjani, M.B.; Gambo, H.I.; Gulani, I.A. Antibacterial efficacy of ethyl acetate fraction of Psidium guajava leaf aqueous extract on experimental Escherichia coli (O78) infection in chickens. Vet. World 2015, 8, 358–362. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shittu, O.B.; Ajayi, O.L.; Bankole, S.O.; Popoola, T.O.S. Intestinal ameliorative effects of traditional Ogi-tutu, Vernonia amygdalina and Psidium guajava in mice infected with Vibrio cholera. Afr. Health Sci. 2016, 16, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Jan, O.Q.; Kamili, N.; Ashraf, A.; Iqbal, A.; Sharma, R.K.; Rastogi, A. Haematobiochemical parameters of goats fed tannin rich Psidium guajava and Carissa spinarum against Haemonchus contortus infection in India. J. Parasit. Dis. 2013, 39, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Seo, N.; Ito, T.; Wang, N.; Yao, X.; Tokura, Y.; Furukawa, F.; Takigawa, M.; Kitanaka, S. Anti-allergic Psidium guajava extracts exert an antitumor effect by inhibition of T regulatory cells and resultant augmentation of Th1 cells. Anticancer Res. 2005, 25, 3763–3770. [Google Scholar] [PubMed]
- Adeyemi, O.S.; Akanji, M.A.; Ekanem, J.T. Anti-anaemic properties of the ethanolic extracts of Psidium guajava in Trypanosoma brucei brucei Infected Rats. Res. J. Pharmacol. 2010, 4, 74–77. [Google Scholar] [CrossRef]
- Udem, S.C.; Anyanwu, M.U.; Obidike, R.I.; Udem, N.D. The effects of Psidium guajava Linn. (Myrtaceae) leaf chloroform extract on some hematological and biochemical parameters in mice. Comp. Clin. Pathol. 2011, 20, 47–51. [Google Scholar] [CrossRef]
- Raj, V.B.; Rao, R.M.; Kumar, R.K.; Srinivas, K. Analgesic effect of ethanol extracted leaves of Psidium guajava leaves in animal models. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 1796–1801. [Google Scholar]
- Stalin D., J. A study on the analgesic property of methanolic and aqueous extracts of dried leaf of Psidium guajava Linn. Int. J. Adv. Res. Pharm. Bio Sci. 2013, 3, 1–6. [Google Scholar]
- Akinloye, O.; Akinmoladun, A.C.; Farombi, E.O. Modulatory effect of Psidium guajava linn and ocimum gratissimum Linn on lipid profile and selected biochemical indices in rabbits fed high cholesterol diet. J. Complement. Integr. Med. 2010, 7. [Google Scholar] [CrossRef]
- Mesquita Freire, J.; Patto de Abreu, C.M.; da Silveira Duarte, S.M.; Borges Araújo de Paula, F.; Ribeiro Lima, A. Evaluation of the protective effect of guava fruits and leaves on oxidative stress. Acta Sci. Biol. Sci. 2014, 36, 35–40. [Google Scholar]
- Oh, W.K.; Lee, C.H.; Lee, M.S.; Bae, E.Y.; Sohn, C.B.; Oh, H.; Kim, B.Y.; Ahn, J.S. Antidiabetic effects of extracts from. Psidium Guajava 2005, 96, 411–415. [Google Scholar]
- Mansoori Bahrani, A.H.; Zaheri, H.; Soltani, N.; Kharazmi, F. Effect of the administration of Psidium guava leaves on blood glucose, lipid profiles and sensitivity of the vascular mesenteric bed to Phenylephrine in streptozotocin-induced diabetic rats. J. Diabetes Mellit. 2012, 2, 138–145. [Google Scholar] [CrossRef]
- Shen, S.-C.; Cheng, F.-C.; Wu, N.-J. Effect of guava (Psidium guajava Linn.) leaf soluble solids on glucose metabolism in type 2 diabetic rats. Phytother. Res. 2008, 22, 1458–1464. [Google Scholar] [CrossRef] [PubMed]
- Ogueri, C.C.; Elekwa, I.; Ude, V.C.; Ugbogu, A.E. Effect of aqueous extract of guava (Psidium guajava) leaf on blood glucose and liver enzymes in alloxan induced diabetic rats. Br. J. Pharm. Res. 2014, 4, 1079–1087. [Google Scholar] [CrossRef]
- Shakeera Banu, M.; Sujatha, K.; Sridharan, G.; Manikandan, R. Antihyperglycemic and antihyperlipidemic potentials of Psidium guajava in alloxan-induced diabetic rats. Asian J. Pharm. Clin. Res. 2013, 6, 88–89. [Google Scholar]
- Adeyemi, O.S.; Akanji, M.A. Biochemical changes in the kidney and liver of rats following administration of ethanolic extract of Psidium guajava leaves. Hum. Exp. Toxicol. 2011, 30, 1266–1274. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.S.; Akanji, M.A. Psidium guajava leaf extract: Effects on rat serum homeostasis and tissue morphology. Comp. Clin. Pathol. 2012, 21, 401–407. [Google Scholar] [CrossRef]
- Ademiluyi, A.O.; Oboh, G.; Ogunsuyi, O.B.; Oloruntoba, F.M. A comparative study on antihypertensive and antioxidant properties of phenolic extracts from fruit and leaf of some guava (Psidium guajava L.) varieties. Comp. Clin. Pathol. 2015, 25, 363–374. [Google Scholar] [CrossRef]
- Chiwororo, W.D.H.; Ojewole, J.A.O. Biphasic effect of Psidium guajava Linn. (Myrtaceae) leaf aqueous extract on rat isolated vascular smooth muscles. J. Smooth Muscle Res. 2008, 44, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Olatunji-Bello, I.I.; Odusanya, A.J.; Raji, I.; Ladipo, C.O. Contractile effect of the aqueous extract of Psidium guajava leaves on aortic rings in rat. Fitoterapia 2007, 78, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Hosokawa, T.; Morinaka, T.; Irino, S.; Hirano, S.; Kobayashi, H.; Yoshioka, A.; Suzuki-Yamamoto, T.; Yokoro, M.; Kimoto, M.; et al. Antiatherogenic effect of guava leaf extracts inhibiting leucocyte-type 12-lipoxygenase activity. Food Chem. 2012, 131, 1069–1075. [Google Scholar] [CrossRef]
- Takahashi, Y.; Otsuki, A.; Mori, Y.; Kawakami, Y.; Ito, H. Inhibition of leukocyte-type 12-lipoxygenase by guava tea leaves prevents development of atherosclerosis. Food Chem. 2015, 186, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Soman, S.; Rajamanickam, C.; Rauf, A.A.; Indira, M. Beneficial effects of Psidium guajava leaf extract on diabetic myocardium. Exp. Toxicol. Pathol. 2013, 65, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Livingston Raja, N.R.; Sundar, K. Psidium guajava Linn confers gastro protective effects on rats. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 151–156. [Google Scholar] [PubMed]
- Tende, J.A.; Eze, E.D.; Tende, Y.A.; Onaadepo, O.; Shaibu, A. Anti-ulcerogenic activity of guava (Psidium guajava) leaves extract in rats. Ann. Exp. Biol. 2013, 1, 6–9. [Google Scholar]
- Umana Uduak, E.; Timbuak, J.A.; Musa, S.A.; Ikyembe, D.T.; Abdurrashid, S.; Hamman, W.O. Ulceroprotective effect of methanol extract of Psidium guajava leaves on ethanol induced gastric ulcer in adult wistar rats. Asian J. Med. Sci. 2012, 4, 75–78. [Google Scholar]
- Jayakumari, S.; Anbu, J.; Ravichandiran, V.; Anjana, A.; Siva Kumar, G.M.; Maharaj, S. Antiulcerogenic and free radical scavenging activity of flavonoid fraction of Psidium guajava Linn leaves. Int. J. Pharm. Pharm. Sci. 2012, 4, 170–174. [Google Scholar]
- Ojewole, J.A.O.; Awe, E.O.; Chiwororo, W.D.H. Antidiarrhoeal activity of Psidium guajava Linn. (Myrtaceae) leaf aqueous extract in rodents. J. Smooth Muscle Res. 2008, 44, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Ezekwesili, J.O.; Nkemdilim, U.U.; Okeke, C.U. Mechanism of antidiarrhoeal effect of ethanolic extract of Psidium guajava leaves. Biokemistri 2010, 22, 85–90. [Google Scholar]
- Shah, A.J.; Begum, S.; Hassan, S.I.; Ali, S.N.; Siddiqui, B.S.; Gilani, A.-H. Pharmacological basis for the medicinal use of Psidium guajava leave in hyperactive gut disorders. Bangladesh J. Pharmacol. 2011, 6, 100–105. [Google Scholar] [CrossRef][Green Version]
- Roy, C.K.; Kamath, J.V.; Asad, M. Hepatoprotective activity of Psidium guajava Linn. leaf extract. Indian J. Exp. Biol. 2006, 44, 305–311. [Google Scholar] [PubMed]
- D’Mello, P.; Rana, M. Hepatoprotective activity of Psidium guajava extract and its phospholipid complex in paracetamol induced hepatic damage in rats. Int. J. Phytomed. 2010, 2, 85–93. [Google Scholar]
- Taju, G.; Jayanthi, M.; Majeed, S.A. Evaluation of hepatoprotective and antioxidant activity of Psidium guajava leaf extract against acetaminophen induced liver injury in rats. Int. J. Toxicol. Appl. Pharmacol. 2011, 1, 13–20. [Google Scholar]
- Osaman, M.; Ahamad, M.; Mahfouz, S.; Elaby, S. Biochemical studies on the hepatoprotective effects of pomegranate and guava ethanol extracts. N. Y. Sci. J. 2011, 4, 1–15. [Google Scholar]
- Mohamed, E.A.K. Hepatoprotective effect of aqueous leaves extract of Psidium guajava and Zizyphus spina-christi against paracetamol induced hepatotoxicity in rats. J. Appl. Sci. Res. 2012, 8, 2800–2806. [Google Scholar]
- Choi, J.H.; Park, B.H.; Kim, H.G.; Hwang, Y.P.; Han, E.H.; Jin, S.W.; Seo, J.K.; Chung, Y.C.; Jeong, H.G. Inhibitory effect of Psidium guajava water extract in the development of 2,4-dinitrochlorobenzene-induced atopic dermatitis in NC/Nga mice. Food Chem. Toxicol. 2012, 50, 2923–2929. [Google Scholar] [CrossRef] [PubMed]
- Porta Santos Fernandes, K.; Kalil Bussadori, S.; Martins Marques, M.; Sumie Wadt Yamashita, N.; Bach, E.; Domingues Martins, M. Healing and cytotoxic effects of Psidium guajava (Myrtaceae) leaf extracts. Braz. J. Oral Sci. 2010, 9, 9–14. [Google Scholar]
- Baroroh, H.N.; Utami, E.D. Harwoko inhibitory effect of ethanolic extract of Psidium guajava leaves in rat active cutaneus anaphylaxis reaction. Int. J. Pharm. Clin. Res. 2016, 8, 1–5. [Google Scholar]
- Wang, X.; Ye, K.; Lv, Y.; Wei, S.; Li, X.; Ma, J.; Zhang, X.; Ye, C. Ameliorative effect and underlying mechanisms of total triterpenoids from Psidium guajava Linn (myrtaceae) leaf on high-fat streptozotocin-induced diabetic peripheral neuropathy in rats. Trop. J. Pharm. Res. 2016, 15, 327–333. [Google Scholar] [CrossRef]
- Kumar, A.; Kumarchandra, R.; Rai, R.; Rao, B. Radiomodulatory role of Psidium guajava leaf extracts against X-ray induced genotoxicity, oxidative stress and apoptosis in albino wistar rat model. J. Appl. Pharm. Sci. 2016, 6, 58–65. [Google Scholar] [CrossRef]
- Ekaluo, U.B.; Ikpeme, E.V.; Uno, U.U.; Umeh, S.O.; Erem, F.A. Protective role of aqueous guava leaf extract against caffeine induced spermatotoxicity in albino rats. Res. J. Med. Plant 2016, 10, 98–105. [Google Scholar] [CrossRef][Green Version]
- Kaneko, K.; Suzuki, K.; Iwadate-Iwata, E.; Kato, I.; Uchida, K.; Onoue, M. Evaluation of food-drug interaction of guava leaf tea. Phyther. Res. 2013, 27, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Vladislavovna Doubova, S.; Reyes Morales, H.; Flores Hernández, S.; Martínez-García, M.D.C.; González de Cossío Ortiz, M.; Chávez Soto, M.A.; Rivera Arce, E.; Lozoya, X. Effect of a Psidii guajavae folium extract in the treatment of primary dysmenorrhea: A randomized clinical trial. J. Ethnopharmacol. 2007, 110, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, D.; Nayana, O.V.; Thomas, L. Antimicrobial plant extract incorporated gelatin beads for potential application in pharmaceutical industry. Biotechnol. Res. 2016, 2, 11–14. [Google Scholar]
- Giri, S.S.; Sen, S.S.; Chi, C.; Kim, H.J.; Yun, S.; Park, S.C.; Sukumaran, V. Effect of guava leaves on the growth performance and cytokine gene expression of Labeo rohita and its susceptibility to Aeromonas hydrophila infection. Fish Shellfish Immunol. 2015, 46, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Fawole, F.J.; Sahu, N.P.; Pal, A.K.; Ravindran, A. Haemato-immunological response of Labeo rohita (Hamilton) fingerlings fed leaf extracts and challenged by Aeromonas hydrophila. Aquac. Res. 2016, 47, 3788–3799. [Google Scholar] [CrossRef]
- Gobi, N.; Ramya, C.; Vaseeharan, B.; Malaikozhundan, B.; Vijayakumar, S.; Kadarkarai, M.; Benelli, G. Oreochromis mossambicus diet supplementation with Psidium guajava leaf extracts enhance growth, immune, antioxidant response and resistance to Aeromonas hydrophila. Fish Shellfish Immunol. 2016, 58, 572–583. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Origin | Extraction Method | Major Constituent | Microorganisms | Assay | Main Results | Ref. |
|---|---|---|---|---|---|---|
| Saudi Arabia | Decoction (30 min) | - | Staphylococcus aureus strains | Agar well diffusion assay | At 200 µL: iz ≤ 30 mm. | [13] |
| India | Soxhlet with MeOH (12 h), maceration in H2O (4 h) | - | S. aureus strains | Agar well diffusion assay, time-kill of bacterial cell, SDS-PAGE analysis, and cellular toxicity to human erythrocytes assays | At 20 mg/L: iz ≤ 20 mm, MIC: 25 µg/mL (MeOH) and 7.5 mg/mL (H2O). MBC: 1.25 and 12.5 mg/mL, respectively, 10 h to kill bacteria, ↑ degradation of protein, no hemolysis. | [14] |
| Nigeria | Maceration in MeOH (48 h) | - | S. aureus, Escherichia coli, Pseudomonas aeruginosa, Proteus spp., and Shigella spp. | Agar well diffusion assay | At 20 mg/mL: iz ≤ 18 mm; 81.8% prevention growth. | [15] |
| India | Maceration with agitation in MeOH, Ac, and DMF (12 h) | - | G-p and G-n bacteria and fungi (91 clinically important strains) | Disc diffusion assay | At 25 mg/mL: against g-p 70% MeOH > 80% Ac > 50% DE, ↓ 76.36% g-n bacteria. Fungi 56% Ac > 38% ME > 31% DMF. No activity against Citrobacter spp., Alcaligenes fecalis, and Aspergillus spp. | [16] |
| India | Soxhlet with MeOH (4 h) | Phytochemical screening: mainly flavonoid-glycosides and tannins | Bacteria (Bacillus subtilis, S. aureus and E. coli), and fungi (Candida albicans and Aspergillus niger) | Paper disc diffusion assay | At 50 μg/mL: iz ≤ 12.6 mm and 10 mm for bacterial and fungi strains, respectively. E. coli: MIC 0.78 μg/mL, MBC 50 μg/mL, and MFC 12.5 μg/mL. | [17] |
| Brazil | Maceration with stirring in EtOH:H2O 70% (v/v) (50 °C, 1 h) | TPC: 25.93 (% m/m, dry base), TFC: 23.48 (mg/g, dry base) | Fungi (C. albicans, Candida krusei, and Candida glabrata), G-p (S. aureus) and G-n (E. coli and P. aeruginosa) | Microdilution assay | MIC = 80–100 µg/mL (C. krusei, C. glabrata and S. aureus) and MBC, MFC ≤ 250–1000 µg/mL (the others). | [18] |
| Brazil | Turbo-extraction with water or Ac:H2O 70% (v/v) (20 min) | Gallic acid: 0.065 µg/g, Catechin: 1.04 µg/g | G-p strains (S. aureus, Staphylococcus epidermidis, and Enterococcus faecalis) G-n (E. coli, Salmonella enteritidis, Shigella flexneri, and Klebsiella pneumoniae) | Agar-diffusion and microdilution assays | At 5 mg/mL: iz ≤ 20 mm, MIC = 39 μg/mL (S. epidermis), MIC < 600 μg/mL (the others). | [19] |
| India | Soxhlet with n-hexane | Methyl 2,6,10-trimethyltridecanoate (28.86%) and Methyl octadecanoate (22.18%) | G-p: S. aureus, Streptoccocus faecalis, Bacillus subtillis, Lactobacillus spp., Enterococcus aerogenes, Acinetobacter spp. G-n: E. coli, Proteus vulgari, Enterobacter aerogenes, Salmonella typhimurium, P. aeruginosa, and K. pneumoniae | Agar well diffusion assay | At 80 µL: iz ≤ 27 mm, MIC = 3–10 µL. | [20] |
| Brazil | Maceration in EtOH:H2O 70% (v/v) (72 h) | - | E. coli, P. aeruginosa, and S. Aureus | Microdilution assay | Only S. aureus (MIC = 256 mg/mL). Synergic effect with ciprofloxacin and gentamicin at 1024 mg/mL. | [21] |
| Brazil | Maceration in MeOH:H2O 70% (v/v) (48h) | - | S. aureus strains | Disc diffusion assay | MIC 90% = 0.52 mg/mL, at 131.75 mg/mL synergic effect with tetracycline, chloramphenicol, erythromycin, vancomycin, oxacillin, cephalothin, ampicillin, cefoxitin, cotrimoxazole. | [22] |
| Egypt | Maceration in EtOH:H2O 50% (v/v) | Quercetin, avicularin, guajaverin, isoquercitrin, hyperin | S. aureus, E. coli, P. aeruginosa, and C. albicans | Agar well diffusion assay | S. aureus: ↑ iz quercetin (28 mm), MIC (μg/mL) guajaverin (0.09–0.19) < avicularin (0.09–0.38) < quercetin (1.25) for all the microorganism tested. | [23] |
| Indonesia | Maceration in EtOH:H2O 30% (v/v) (3 days) | Tannins (2.35 mg/g) | E. coli, P. aureginosa, S. aureus, A. niger and C. Albicans | Paper disc diffusion method | iz ≤ 15 mm. | [24] |
| India | Soxhlet with toluene (72 h) | Betulinic acid and lupeol | Fungi: Calletotricheme camellie, Fussarium equisitae, Alterneria alternate, Curvularia eragrostidies, and Colletrichum Gleosproides. Bacteria: E. Coli, B. Subtillis, S. aureus, and Enterobactor | Slide germination method | Bacteria: MIC < 100–200 μg/mL, fungi: MIC < 2.5–10 μg/mL. | [25] |
| India | Soxhlet with Ac (8 h) | - | H. bispinosa Neumann (Acarina: Ixodidae) and H. maculata Leach (Diptera: Hippoboscidae) | Antiparasitic activity method of FAO (2004) | At 3 mg/mL: mortality 100% H. maculate, 78% H. bispinosa, parasite dead H. maculata (LC50 = 0.646 mg/mL). | [26] |
| Nigeria | Maceration with agitation in EtOH:H2O 20% and 80% (v/v) (24 h) | - | Trypanosoma brucei brucei and HEK293 | Alamar Blue assays | At 238.10 μg/mL: IC50 (T. b. brucei) = 6.3 μg/mL and 48.9 μg/mL for 80% and 20% extracts, respectively, IC50 (HEK293) 30.1 and 24.16%, respectively. | [27] |
| India | Soxhlet with ethyl acetate and MeOH (8 h) | - | Plasmodium falciparum strains | SYBR green assay | IC50 9–18 μg/mL, resistance indices = 0.6 and 1.4 in MeOH and ethyl acetate, respectively. | [28] |
| Malaysia | Hydrodistillation (3 h) | - | Toxoplasma gondii | MTT assay with Vero cells | At 200 µg/mL: No cytotoxic effect (EC50 = 37.54 µg/mL), anti-parasitic activity (EC50 of 3.94 µg/mL). | [29] |
| India | Soxhlet with EtOH, and maceration in H2O (6 days) | - | E. coli, Shigella spp., Salmonella spp., Aeromonas spp., S. aureus, and Candida spp. | Agar well diffusion assay | H2O: iz ≤ 30 mm (max C. albicans). EtOH: iz ≤ 31 mm (max Aeromonas hydrophila). | [30] |
| India | Soxhlet with EtOH:H2O 70% (v/v), MeOH, ethyl acetate, and H2O | Phytochemical analysis: tannins, saponins, flavonoids, terpenoids, sugars | E. coli, Salmonella spp., and Vibrio cholerae | Agar well diffusion assay | At 1000 µg/mL: iz ≤ 30 mm. MeOH: MIC (100%) > 250 µg/mL. EtOH:H2O:MICs (38–65%) > 500 µg/mL and > 750 µg/mL. Ethyl acetate and H2O: MICs > 750 µg/mL. | [31] |
| Bangladesh | Maceration in H2O and MeOH:H2O 75% (v/v) (48 h) | - | V. cholera | Agar well diffusion assay | MICs = 1250 µg/mL (H2O), 850 µg/mL for (MeOH:H2O). Antibacterial resistance to trimethoprim/sulfomethoxazole, furazolidone, tetracycline, and erythromycin. | [32] |
| India | Decoction | Major component: quercetin (2 mg/g) | E. coli (heat labile (HLT) and cholera toxin (CT)), V. cholerae, Shigella flexneri | Microtitre plate based assay. Assays for bacterial colonization (adherence and invasion) and enterotoxins | At 2.7 mg/mL: (EC50 = 0.98 (S. flexneri) and 2.88% (V. cholerae). ↓ adherence and invasion to epithelial cells (EC50 = 0.37–1.25% and 0.04–0.25%, respectively). The effect on adherence is not due to quercetin and the invasion is lower than with the extract. ↓ Production of HLT and CT (EC50 = 1.03 and 2.69%) and binding to glioside monosialic acid enzyme (EC50 = 0.06 and 2.51%). | [33] |
| Brazil | Soxhlet with n-hexane, ethyl acetate, MeOH, H2O (24 h) | - | S. aureus, Salmonella spp., and E. coli | Disc diffusion method | At 1938 µg/disc: iz = 7.00–11.25 mm (Soxhlet), and 11–18 mm (H2O). No inhibition to E. coli (H2O) and Salmonella spp. (hexane and ethyl acetate). | [34] |
| Nigeria | Soxhlet with EtOH:H2O 60% (v/v) (5 h), and H2O (3 h) | - | E. coli and S. aureus | Agar well diffusion assay | At 10 mg/mL: H2O: iz = 9–16 mm and 8–11 mm, MICs = 5 and 2.5 mg/mL (E. coli and S. aureus, respectively). EtOH:H2O: iz 12–21 and 11–14 mm, MICs = 1.25 and 0.625 mg/mL, respectively. | [35] |
| Japan | Infusion (8 min) | Tannin content: 1.11 mg/mL | H1N1 virus strains | 19-h Influenza growth inhibition assay | At 0.4 mg/mL: inhibition growth (IC50 = 0.05–0.42%). | [36] |
| Origin | Extraction Method | Major Constituent | Cell Line | Assay | Main Results | Ref. |
|---|---|---|---|---|---|---|
| Japan | Maceration in EtOH:H2O 50% (v/v) | TPC: 71 g/100 g | Human colon adenocarcinoma (COLO320DMA) | Cyclooxygenase and cell proliferation assays | At 1 mg/mL: ↓ human cyclooxygenase activity (IC50 55 and 560 µg/mL PGHS-1 and 2, respectively), ↓ IC50 5.1 µg/mL (PGSH) and 4.5 µg/mL (cyclooxygenase). | [37] |
| At 100 µg/mL: Quercetin ↓ IC50 = 5.3 (PGSH-1) and 250 µg/mL (PGSH-2), ↓ DNA synthesis rate. | ||||||
| Malaysia | Soxhlet with ether, MeOH, and H2O | - | Cervical cancer (HeLa), breast cancer (MDA-MB-231) and osteosarcoma (MG-63). Control: non-malignant Madin-Darby canine kidney (MDCK) | Methylene blue assay | At 10 mg/mL: HeLa: No anti-proliferative activity. | [38] |
| MDA-MB-231: IC50 ether extract (4.2 µg/mL) > MeOH (18.6 µg/mL) > H2O (55.7 µg/mL). | ||||||
| MG-63: same order (IC50 of 5.42, 23.25, and 61.88 µg/mL, respectively). | ||||||
| MDCK: cytotoxic effect of ether and MeOH extract (IC50 = 5.03 and 11.55 µg/mL, respectively). | ||||||
| Brazil | Maceration in EtOH | TPC: 766.08 mg/g, TFC: 118.90 mg/g | HeLa, colorectal carcinoma (RKO-AS45-1), and lung fibroblast (Wi-26VA4) | MTT assay | At 1 mg/mL: IC50 = 15.6 µg/mL (HeLa), 21.2 (RKO) µg/mL, and 68.9 µg/mL (Wi-26VA4). | [39] |
| Palestine | Maceration in DCM:MeOH 50% (v/v) (24 h) | - | Murine fibrosarcoma (L929sA), and human breast cancer (MDA-MB-231 and MCF-7) | MTT assay | IC50 = 55 µg/mL (L929sA), 820 µg/mL (MCF7 cells), no cytotoxic effect on MDA-MB-231 cells. | [40] |
| Taiwan | Decoction (30 min) | - | Human prostate carcinoma (DU-145) | MTT, ELISA, gelatinolytic zymography, wound scratch, and chicken chorioallantoic membrane assays | At 0.25 mg/mL: cell suppression (IC50 0.57 mg/mL). ↓ Expressions of VEGF (76.9%), IL-6 (98.8%) and IL-8 (98%), and MMP-2 (100%) and MMP-9 (100%). Suppressed the cell migration (30.9%) and the angiogenesis. | [41] |
| Taiwan | Decoction (1 h) | TPC: 470.0 mg/g Individual compounds: gallic acid (348), catechin (102), epicatechin (60), rutin (100), quercetin (102), and rutin (100) in mg/g | Human prostate epithelial (PZ-HPV-7) and DU-145 | MTT assay | At 1 mg/mL: 100% suppression DU-145 cells. PZ-HPV-7 cells followed an auto-decaying process. Cell-killing rate coefficient (kapp) = 0.03 × 103 phenolic compounds cells/mg h. | [42] |
| Brazil | Soxhlet with DCM. Maceration with EtOH | Guajadial, psidial A, and psiguajadial A and B | Leukemia (K-562), MCF-7, ovarian cancer (NCI/ADR-RES), lung (NCI-H460), melanoma (UACC-62), prostate (PC-3), colon (HT-29), ovarian (OVCAR-3), and kidney (786-0) | Protocol established by NCI (ELISA test) | At 250 µg/mL: Anti-proliferative activity DCM > EtOH, inhibition growths: 26 (OVCAR-3)-65 (UACC-62) µg/mL due to the major compounds. | [43] |
| Japan | Maceration with sonication in MeOH:H2O 80% (v/v) (3 h) and isolation | CPO | PC-3 and MCF-7 | MTT, annexin V antibody, TUNEL, and western blot assays | At 50 µg/mL: ↓ cell proliferation, ↑ early and late apoptotic effect, down-regulation of PI3K/AKT/mTOR/S6K1 pathway, up-regulation of MAPKs, JNK, ERKs, and p38 MAPK. | [44] |
| Thailand | Hydrodistillation | Human mouth epidermal carcinoma (KB) and murine leukemia (P388) | MTT assay | At 0.15 mg/mL: KB: 75% cytotoxic effect, IC50 = 0.04 mg/mL; At 0.08 mg/mL: P388: 80% cytotoxic effect, IC50 = 0.05 mg/mL. | [45] | |
| Jamaica | Maceration in hexane (4 days) | - | Leukemia (Kasumi-1) | MTT assay | IC50 = 200 µg/mL. | [46] |
| Japan | Maceration with sonication in MeOH:H2O 80% (v/v) (3 h). Fractionation with hexane | 60 compounds (in hexane fraction): β-eudesmol (11.98%), α-copaene (7.97%), phytol (7.95%), α-patchoulene (3.76%), and CPO (3.63%) | Human prostate cancer (PC-3 and LNCaP) | MTT, annexin V antibody, TUNEL, and western blot assays | At 150 µg/mL: PC-3: ↑ apoptotic effect of the hexane fraction (15%), ↓ effect on early apoptotic cells, ↑ effect for late apoptosis, via the suppression of PI3K/AKT/mTOR/S6K1 and MAPK signalling cascades in both cell lines. | [47] |
| Origin | Extraction Method | Major Constituent | Cells | Assay | Main Results | Ref. |
|---|---|---|---|---|---|---|
| Korea | Maceration in MeOH:H2O 70% (v/v) (5 days) | - | LPS-stimulated RAW 264.7 (Mouse macrophage) | Griess, MTT, ELISA kit, western blot, transient transfection, and luciferase assays | At 125 µg/mL: no cytotoxic effect, ↑ 44–62% inhibition rates. ↓ LPS-induced NO and PEG2 ↓ iNOS and COX-2 (↓ I-κBα degradation, ↓ activation NF-κB). | [48] |
| Palestine | Maceration in DCM:MeOH 50% (v/v) (24 h) | - | L929sA fibroblast | Transfection and luciferase assays | At 62.5 µg/mL: ↓ expression of IL-6 and NF-κB luciferase reporter gene construct via the NF-κB transactivation level, since no ↓ inhibition of NF-κB/DNA binding. | [40] |
| Korea | Extraction in MeOH:H2O 70% (v/v) (6 h) | TPC: 426.84 mg (GAE)/g | LPS-stimulated RAW 264.7 | MTT, Griess, and ELISA test assays | At 30 µg/mL: no cytotoxic effect. ↓ LPS-induced NO (52.58%) and the production of PGE2 (43.45). | [49] |
| Korea | Extraction in EtOH:H2O 55% (v/v) (4.9 h, 47 °C) | Gallic acid (0.2) and catechin (4.4) in mg/g | LPS-stimulated RAW 264.7 | MTT, Griess, ELISA test, RT-PCR, and total western blot assays | At 50 µg/mL: no cytotoxic effect. ↓ LPS-induced NO (>65%) by ↓ iNOS, ↓ PGE2 (to basal level) via ↓ COX-2 mRNA. ↓ IL-6. ↓ iNOS and COX-2 due to the down-regulation of ERK1/2 pathway, because no effect was found to other proteins at the dose tested. | [50] |
| India | Maceration in MeOH:H2O 90% (v/v) (x3) | - | LPS-stimulated in Labeo rohita head-kidney macrophages | MTT, Greiss, ELISA, RT-PCR, and western blot assays | At 200 µg/mL, ↓ LPS-induced NO (75%) by ↓ iNOS-mRNA, ↓ PGE2 (45%) via ↓ production COX-2-mRNA, TNF-α, IL-1β, IL-10, and mRNA expression. Suppressed phosphorylation of MAPK (↓ I-κBα degradation ↓ activation NF-κB). | [51] |
| Korea | Soxhlet with EtOH:H2O 55% (v/v) (4.9 h, 47 °C) | Gallic acid (0.09) and catechin (0.72) in mg/g | LPS-stimulated RAW 264.7 | MTT, Greiss and ELISA test assays | At 30 µg/mL: no cytotoxic effect. ↓ LPS-induced NO (47.5%) and PGE2 (45.8). | [52] |
| India | Maceration with agitation in MeOH and EtOH (24 h) | - | Human blood | HRBC membrane stabilization method | At 200 µg/mL: ↑ 13.8–14.4% prevention of lysis of the membrane. | [53] |
| Indonesia | Maceration with agitation in EtOH:H2O 96% (v/v) (6 h) | TPC: 101.93 mg GAE/g | Human lymphocyte | MTT assay | 0.5 μg/mL: Stimulation index 1.54%. | [54] |
| Origin | Compound | Assay | Main Results | Ref. |
|---|---|---|---|---|
| India | Ethyl acetate fraction | In vitro glycation of BSA-fluorescence measurement | In vitro AGEs formation with IC50 of 38.95 ± 3.08 μg/mL. | [55] |
| Taiwan | Gallic acid, catechin and quercetin | In vitro glycation of BSA-fluorescence measurement; Fructosamine assay and Girard-T assay | At 100 μg/mL: 80% inhibitory effects on the formation of α-dicarbonyl compounds at a concentration of 50 µg/mL, inhibitory effects on AGEs formation in BSA glycation systems. | [56] |
| China | Quercetin, kaempferol, myricetin | Rat intestinal sucrase and maltase inhibitory activities; Porcine pancreatic α-amylase inhibitory activity | At 1.5 mg/mL: inhibitory activities with IC50 values of 3.5, 5.2 and 3.0 mM against sucrase, with IC50 values of 4.8, 5.6 and 4.1 mM against maltase and with IC50 values of 4.8, 5.3 and 4.3 mM against α-amylase, respectively. Synergistic effect against α-glucosidase. | [57] |
| China | Water-soluble polysaccharides, including GP90 and P90 | α-Glucosidase inhibition assay | α-Glucosidase inhibition activity with an EC50 of 2.27 µg/mL and 0.18 mg/mL. | [59] |
| - | Peltatoside, hyperoside, isoquercitrin, guaijaverin and flavonol-glycosides | Spectrophotometric assay; absorption assay into CaCo-2 cells | Concentration of the compounds (0.01 to 0.06 µmol/mL). Individual flavonol-glycosides inhibited DP-IV dose-dependently. The ethanolic guava leaves extract (380 µg/mL) showed a dose-dependent inhibition of DP-IV, with an IC50 of 380 μg/mL test assay solution; the highest uptake was from Guaijaverin. | [60] |
| Korea | Quercetin and catechin | Fructose transport in CaCo-2 cell systems | At 1 mg/mL: inhibition of fructose uptake (55%). At 30 µg/mL: quercetin contributed to both, GLUT2 and 5 transporters, and catechin to GLUT5-mediated fructose uptake inhibition. | [61] |
| India | Guavanoic acid | Spectrophotometric assay | At 27 μg/mL: remarkable PTP1B inhibitory activity (90%) and in vitro stability in various physiological medium including saline, histidine, cysteine, BSA, HSA and buffers (pH 5, 7 and 9). IC50 = 1.14 μg/mL. | [64] |
| India | n-Hexane, methanol, ethanol and aqueous leaf extracts | Inhibitory glucose diffusion | At 50 g/L: the methanol extract was the most potent with the lowest mean glucose concentration of 201 ± 1.69 mg/dL at the end of 27 h (↓ 93% uptake). | [63] |
| Japan | 70% Ethanol extract | Oil Red O Assay; Real-Time RT | At 100 μg/mL: inhibition of 3T3-L1 differentiation via down-regulation of adipogenic transcription factors and markers (mRNA levels of PPAR-γ, C/EBP-α, and aP2), and suppression of mitotic clonal expansion (at day 4 and 8). | [65] |
| Taiwan | Aqueous extract | Glucose uptake test; bicinchonic acid method; Western-blot analysis | At 400 µg/mL: ↑ IR (25.1%), p-IR (46.2%), p-IRS (51.2%), PI3K (32.2%), Akt (46.1%), p-Akt (36.3%), GLUT-2 (46.8%), and total glycogen synthase (45.5%). | [66] |
| Taiwan | Vescalagin | Glucose-uptake test | At 100 µg/mL: Enhancement of glucose uptake in TNF-α-induced insulin-resistant. | [67] |
| Origin | Extraction Method | Microorganism(s)/Cells | Assay | Main Results | Ref. |
|---|---|---|---|---|---|
| India | Soxhlet with MeOH (4.5 h) | S. mutans strains | Agar well diffusion assay, effect on acid production, on sucrose-dependent adherence to smooth glass surfaces, and on sucrose-induced cellular aggregation, and MATH assays | MIC > 5 mg/mL (MeOH). MIC = 2–4 mg/mL (guaijaverin) At sub-MIC (0.125–2 mg/mL): ↑ pH (5 to 6–7), hydrophobicity indexes (3.2–72%), ↓ sucrose-dependent adherence (34–84%) and aggregation. | [73] |
| Malaysia | Decoction | S. sanguinis and S. mutans | NAM model system | At 60.95 mg/mL: MIC = 7.62 (S. sanguinis) and 3.81(S. mutans.) mg/mL. MBC values = 15.24 and 30.48 mg/mL, respectively. At 0.5 mg/mL: ↓ adherence 57 and 60% (single-species) and 88–89% (dual-species). | [74] |
| Malaysia | Sonication with H2O (10 min) | S. sanguinis, S. mitis, and Actinomyces spp. | MATH assay | At 1 mg/mL: ↓ 54.1%, 49.9% and 40.6%, respectively, cell-surface hydrophobicity. At 20 mg/mL: was 64.7, 60.5, and 55.5%, respectively. | [75] |
| Malaysia | Decoction | S. sanguinis, S. mitis, and Actinomyces spp. | Bacterial growth and generation time rates determinations | At 4 mg/mL: Time growth = 1.22 (S. sanguinis, Actinomyces spp) and 2.06 h (S. mitis) ↓ growth 42.6%, 51.2% and 55%. | [76] |
| India | Maceration with stirring in EtOH (2 days) | S. mutans, S. sanguinis, and S. salivarius | Agar well diffusion assay | At 10 mg/mL: inhibition zones of 21.17, 18.58, and 23.00 mm, respectively. | [77] |
| India | Maceration (2 days) and Soxhlet (6 h) with EtOH, H2O, and EtOH:H2O 50% (v/v) | S. mutans and S. mitis | Agar well diffusion assay, sucrose-dependent adherence and cellular co-aggregation activities, and biofilm formation sterile acrylic tooth determinations | At 15 mg/mL: inhibition zone for H2O (11.8 mm) to EtOH:H2O (25 mm), both by Soxhlet. MIC = 1 mg/mL. EtOH:H2O extract: at >0.05 mg/mL: ↓ adherence and co-aggregation, at MIC, ↓ the viable count of dental biofilm (3.50 log10 CFU/mL). | [78] |
| India | Soxhlet with EtOH:H2O 50% (v/v) (6 h) | S. mutans and S. mitis | MATH assay | At >1 mg/mL ↓ hydrophobicity (index < 40%). | [79] |
| India | Maceration with stirring (2 days) and Soxhlet with EtOH | S. mutans, S. sanguinis, and S. salivarius | Agar well diffusion assay | At 10 mg/mL: ↑ inhibition zones for maceration extracts (19–23 mm). | [80] |
| Ghana | Maceration with agitation in EtOH:H2O 70% (v/v) (24 h) | Aggregatibacter actinomycetemcomitans strains | Agar well diffusion assay, release of the cytosol enzyme lactate dehydrogenase, fluorescence assisted cell sorter, and ELISA assays | No growth inhibitory effect, although neutralized the cell death and pro-inflammatory response, and restored the morphological alterations induced by the leukotoxin. These effects were due to the direct binding of guava compounds and the leukotoxin. | [81] |
| India | Maceration in Ac, EtOH, chloroform, MeOH and H2O (15 days at 22 °C) | Neisseria catarrhalis, S. mutans, S. salivarius, Streptococcus viridans, Bacillus megaterium, and P. aeruginosa | Agar well diffusion assay | ↑ Inhibition zones in Ac (15–29 mm), except for N. catarrhalis (20 mm in MeOH). | [82] |
| India | Maceration in MeOH (72 h). Fractionation with ethyl acetate | S. aureus and S. mutans | HRBC membrane stabilization method, disc and agar well diffusion assays | MeOH and ethyl acetate fraction ↑ protection (84–99%) to the inflammatory response. Inhibition zones (25–100 µg/mL) = 10.5 to 22 mm by both methods. MICs = 0.48 (ethyl acetate) and 0.62 (MeOH) mg/mL. | [83] |
| Taiwan | Maceration EtOH, Ac, H2O (room temperature and 60 °C) (24 h) | Clone 9 rat liver cells | WST-1 and ALT assays | At >500 μg/mL cytotoxic effect of EtOH and Ac and 600 μg/mL for H2O. At <200 μg/mL normal values were observed for H2O and Ac, and EtOH (<500 μg/mL). At <100 μg/mL: Hepato-protective effect in EtOH and H2O (full range). | [85] |
| Origin | Extraction Method | Subject | Treatment | Main Results | Ref. |
|---|---|---|---|---|---|
| Thailand | Maceration in H2O, EtOH, and ether (24 h) | Oreochromis niloticus | Aeromonas hydrophila | LD50 = 3.44 × 106 CFU/mL. ↓ Mortality of the subjects. | [95] |
| China | - | Penaeus monodon | Yellow-head virus, white spot syndrome virus, and Vibrio harveyi | Survival rate = 80–95% (↑ Weight (2 to 6 g)). In serum (↑ feed): ↓ PO (7.50 U/mL) and SOD (178.33 U/mL), ↑ NOS (64.80 U/mL). In hepato-pancreas: ↑ SOD (57.32 U/mg), ACP (23.28 U/mg), AKP (19.35 U/mg), and LSZ (3459.946 U/mg). | [96] |
| Nigeria | Maceration with agitation in EtOH:H2O 80% (v/v) (24 h) | Albino rats | T. b. brucei | At 300 mg/kg: ↓ parasitemia; ↑ survival in 24 days. | [97] |
| Nigeria | Maceration with agitation in EtOH:H2O 80% (v/v) (24 h) | Albino rats | T. b. brucei | Administration 1–7 days. ↑ GSH: liver (5.4 to 8.1), kidney (3.3 to 6.0), and serum (0.8 to 2.4), restored in kidney and serum. In the brain, no effect was found. ↓ MDA: serum (13.9 to 5.9), brain (42.8 to 18.1), kidney (27.3 to 17.6), and liver (38.2 to 19.2). | [98] |
| India | Decoction of the leaves (10 min) | BALB/c mice | Plasmodium berghei | At 350 and 1000 mg/kg ↓ parasitemia (73.7% and 85.8%); ↑ survival 15 and 18 days. | [99] |
| India | Extraction in EtOH:H2O 50% (v/v) | Swiss mice | Citrobacter rodentium | At 300 mg/kg: ↓ infection (day 4) of the treatment, and no infection at day 19 (control group at day 24). | [100] |
| Nigeria | Hidrodistillation and fractionation with ethyl acetate | ISA brown male chicks | E. coli | At 100 mg/kg: In 10 days ↓ signs of villous collapse (stunting, matting and fusion of villi), number of wet droppings (12-6); ↑ activity, weight gaining, and feed intake (from 27 to 45 g) in contrast to the infected ones (from 30 to 18 g); ↓ bacterial shedding load (from 60 to 45 CFU/mL). | [101] |
| Nigeria | Decoction of the leaves | Adult mice | V. cholera | At 250 mg/kg: Histopathological observations: mild degenerative, secretory, and inflammatory changes with goblet cells and with most of the exudate (neutrophils and lymphocytes). | [102] |
| India | - | Adult male goat | Haemonchus contortus | 90 Days feeding: ↑ Hb (7.2 to 8.6 g/dL), PCV (20.2 to 29.3%), total protein (4.8 to 6.3 g/dL), GLO (2.3 to 3.8 g/dL) (↑ control (2.8)), glucose (43.9 to 52.6 g/dL), and calcium (8.7 to 9.6 mg/dL); ↓ blood urea (47.9 to 29.8 mg/dL) (↓ control (41)). Phosphorus balance, serum albumin levels and serum enzyme activity did not show variation. | [103] |
| Origin | Extraction Method | Subject | Treatment | Main Results | Ref. |
|---|---|---|---|---|---|
| Nigeria | Maceration with agitation in EtOH:H2O 80% (v/v) (24 h) | Wistar rats | T. b. brucei/no infected | Treatment (1–7 days) at 150 mg/kg: ↑ Hb (6.5 to 10.7 g/dL), PCV (28.6 to 34.4%), RBCC (4.1 to 5.0 × 1012/L), MCV (53.6 to 64.3 fL), and MCHC (21.4 to 31.4 g/dL); ↓ WBC (23.2 to 19.4 × 109/dL) and neutrophil levels (28.9 to 27.3 × 103/mL).Compared to no infected subjects: similar values that obtained in treated-infected animals but with opposite conclusions. | [105] |
| Nigeria | Extraction in chloroform (24 h) | Mice | No infected | Treatment (28 days) at 45.9 mg/mL: no differences in Hb (12 to 11 g/dL), PCV (37 to 35%), RBCC (6.1 to 5.1 × 106/L), and MCHC (33 to 32 g/dL), and neutrophil levels (13 to 12%); ↑ lymphocyte levels (85 to 92%) and MCV (61 to 69 fL). | [106] |
| Korea | Extraction in EtOH:H2O 55% (v/v) (4.9 h, 47 °C) | Sprague-Dawley rats and mice | Freund’s complete adjuvant-induced hyperalgesia/LPS-induced endotoxic shock | At 400 mg/kg: PWL restored; ↑ 67% survival rate (72 h) by ↓ TNF-α (500 to 325 pg/mL) and IL-6 (80 to 58 ng/mL). | [50] |
| Brazil | Turbo-extraction in water and acetone: H2O 70% (v/v) (20 min) | Swiss mice | Carrageenan-induced peritonitis, acetic acid-induced abdominal writhing and hot plate test | At 50mg/kg: number of leukocyte migration into the peritoneal cavity H2O < H2O -acetone extract. No central analgesic activity. Peripheral analgesic activity: ↓ number writhing response (from 50 to 15 count). | [19] |
| India | Maceration in EtOH (7 days) | Wistar rats | Acetic acid-induced writhing | At 2 mg/kg ↓ 66% number writhing response (from 67 to 54 count). Comparable to diclofenac sodium (75%). | [107] |
| India | Distillation with MeOH and H2O | Wistar rats | Acetic acid-induced writhing and hot plate test | At 10 and 30 mg/kg ↓ responses time (at 9.4 and 10.6 s) compared to the analgesic drug Pentazocine (14 s). | [108] |
| Origin | Subject | Treatment | Main Results | Ref. |
|---|---|---|---|---|
| Nigeria | Rabbits | High-cholesterol diet | At 250 mg/kg: ↓ TC (15%); ↑ HDL (69%); ↓ LDL (74%); ↓ hyperglycemia 43%. | [109] |
| Brazil | Wistar rats | High-cholesterol diet | At 369.89 mg phenolic compound in the extract/g: ↓ TC (29–35%), TG (59–73%); ↑ HDL (46%); ↓ VLDL+LDL; ↓ enzyme activity (SOD (6.2 to 5.7 U/mg protein), GP (4.6 to 2.3 µmol/g protein). | [110] |
| Korea | Leprdb/Leprdb juvenile and adult mice | Diabetes spontaneous mutation | At 10 mg/kg: 87% inhibition PTP1B; ↓ glucose levels 31% and 42% respectively. | [111] |
| Iran | Wistar rat | Streptozotocin-induced diabetes | At 1mg/L: ↓ Ca/Mg ratio (18 to 12), glucose level, TG (100 to 65 mg/dL), TC (68 to 48 mg/dL), ↑ HDL (18 to 40 mg/dL), ↓ LDL, and VLDL to normal levels; ↓ alteration in vascular reactivity (110 to 50 mmHg). | [112] |
| Taiwan | Sprague-Dawley rats | Low-dose streptozotocin and nicotinamide-induced diabetes | At 400 mg/kg: ↓ blood glucose level (230 to 140 mg/dL); ↑ plasma insulin level and glucose utilization (normal levels); ↑ enzyme activity (hepatic hexokinase (8 to 11 U/mg protein), phosphofructokinase (18 to 25 U/mg protein) and glucose-6-phosphate dehydrogenase (11 to 25 U/mg protein). | [113] |
| India | Sprague-Dawley rats | Streptozotocin-induced diabetes | At 100 mg/kg: ↓ blood glucose level (4 to 1 mg/mL) and lipid peroxidation (2 to 1 mmol/100 g tissue); ↑ enzyme activity (CAT (6 to 10 × 103 U/mg protein), SOD (6 to 10 U/mg protein), GPx (0.4 to 0.6 U/mg protein), GRd (0.1 to 0.3 U/mg protein). | [55] |
| Nigeria | Albino rats | Alloxan-induced diabetes | At 200 mg/kg: ↑ average weight (99 to 209g); ↓ blood glucose level (15 to 8 mmol/L); ↓ alanine aminotransferase activity (32 to 24 U/L). | [114] |
| India | Albino rats | Alloxan-induced diabetes | At 500 mg/kg: ↓ blood glucose level, TC (231 to 163 mg/dL), TG (133 to 69 mg/dL), LDL (186 to 126 mg/dL), VLDL (26 to 13 mg/dL); ↑ HDL (18 to 23 mg/dL). | [115] |
| Nigeria | Wister rats | - | At 150 mg/kg: ↑ ALP (300, 175and 650 IU), AST (500, 400, 450 IU), ALT (1200, 1200, 1800 IU), ACP (750, 650, 900 IU) activity in the kidney, liver, and serum, respectively. | [116] |
| Nigeria | Mice | - | At 49.3 mg/mL: ↑ AST (93 to 126 iµ/L), ALT (30 to 35 iµ/L), ALP (57 to 66 iµ/L), conjugate bilirubin (0.2 to 0.3 mg/dL) and creatinine (0.9 to 1.2 mg/dL). | [106] |
| Nigeria | Albino rats | - | At 150 mg/kg: ↑ serum urea (2.9 to 6 mmol/L) and creatinine (2.7 to 4 mmol/L); ↓ concentration of serum Na+ (122 to 99 mmol/L). | [117] |
| Origin | Extraction Method | Subject | Treatment | Main Results | Ref. |
|---|---|---|---|---|---|
| India | Extraction with MeOH | Wistar rats | ASP, PL, and EtOH-induced ulcers | At 200 mg/kg: PL-induced ulcers: ↓ 64% ulcer formation (ui = 2.1), ↓ GV (5 to 2 mL), acid secretion (88 to 64 mEq/L/100 g), ↑ pH (2 to 5).Comparable to omeprazole; ASP (↓ 70.5%, ui = 2.5) and EtOH (↓ 70.4%, ui = 8.7)-induced systems. | [124] |
| Nigeria | Maceration in H2O (24 h) | Albino rats | EtOH-induced ulcers | At 1000 mg/kg: ↓ MNL (9.4 to 2) ui (4.7 to 1). | [125] |
| Nigeria | Maceration with agitation in MeOH (24 h) | Wistar rats | EtOH-induced ulcers | At 1000 mg/kg: ↓ ui (17.7 to 6.3), ↑ protection (64.4%). | [126] |
| India | Maceration in EtOH:H2O 90% (v/v) (72 h). | Wistar rats | PL and EtOH-induced ulcers | At 200 mg/kg: PL-induced: ↓ ulcer formation (77 to 84%), ui (5 to 1.3), GV (1.4 to 0.5 mL/100g), and acid secretion (28 to 23 mEq/L); ↑ pH (2.0 to 3.4). EtOH-induced: ↓ (63% to 79%, ui = 1.6 to 5.6), and gastric lesions (5.6–1.9). | [127] |
| South Africa | Maceration in H2O (48 h) | Wistar rats and BALB/c mice | Castor oil-induced diarrhea and castor oil-induced enteropooling | At 400 mg/kg: ↑ 83.3% rat protection, ↓ 75% fluid accumulation in rats; ↓ 87.73% transit in rats and 77.2% in mice; ↓ 64.35% of contractions in mice. | [128] |
| Nigeria | Soxhlet with EtOH:H2O 70% (v/v) | Wistar rats | Castor oil-induced diarrhea | At 80 mg/kg: ↓ 53.03% transit in rats and ↓ 67.70% intestinal contractions. | [129] |
| Pakistan | Maceration with EtOH | BALB/c mice, rabbit jejunum | Castor oil-induced diarrhea, K+-induced motility | At 1 g/kg: ↑ 81.1% mice protection; Spasmolytic effect (0.3–1 mg/mL) ↓ spontaneous contractions EC50 = 0.66 mg/mL in rabbits. | [130] |
| India | Decoction (1 h) | Wistar rats | CCl4, PCM, and TAA-induced liver injury | At 500 mg/kg: CCl4: ↓ ALT (384 to 17 U/L), AST (642 to 152 U/L), ALP (750 to 489 U/L), and bilirubin (1.6 to 0.3 mg/dL), ↓ control levels; PCM: ↓ ALT (384 to 87 U/L), AST (642 to 179 U/L), ALP (750 to 338 U/L), and bilirubin (1.6 to 0.6 mg/dL); TAA: ↓ ALT (337 to 32 U/L), AST (438 to 237 U/L), and ALP (770 to 479 U/L). | [131] |
| India | Soxhlet with EtOH | Wistar rats | PCM-induced liver injury | At 400 mg/kg: ↓ SGOT (475 to 370), SGPT (158 to 128), ALP (814 to 729), and bilirubin (0.7 to 0.6); ↑ total protein (5.15 to 5.6), albumin (2.6 to 3.1), and GLO (2.1 to 2.4). Histopathological observations: less diffuse granular degeneration and mild periportal lymphocytic infiltration. | [132] |
| India | Decoction (1 h) | Wistar rats | Acetaminophen-induced liver injury | At 500 mg/kg: ↓ AST (121 to 77 IU/L), ALT (80 to 57 IU/L), ALP (115 to 67 IU/L), and total bilirubin (4 to 2 mg/dL). Restored: total protein (5 to 7 g/dL), LPO (7 to 2 nmol/mg protein), GPx (13 to 19 µmol/mg protein), GSH (15 to 23 µmol/mg protein), CAT (14 to 24 µmol/mg protein), and SOD (48 to 63 µmol/mg protein). Histopathological observations: normal lobular structure. | [133] |
| Egypt | Maceration with agitation in EtOH:H2O 70% (v/v) (24 h) | Albino rats | CCl4-induced liver injury | At 500 mg/kg: ↓ ALT (94 to 55 U/mL), AST (199 to 82 U/mL), GGT (71 to 23 U/mL), lysosomal enzymes (50%), and LPO (7 to 3 nmol/mg protein); ↑ SOD (15 to 39 U/mg protein), CAT (5 to 15 µg/mg protein), GSH (6 to 8 µg/mg protein), GST (13 to 25 mM/min/mg protein), total protein (48 to 58 g/L), albumin (29 to 38 g/L), GLO (19 to 21 g/L). | [134] |
| Egypt | Decoction (1 h) | Wistar rats | PCM-induced liver injury | ↓ AST (342 to 156 U/L), ALT (359 to 80 U/L), ALP (288 to 263 U/L), LDH (207 to 143 U/L), GGT (11 to 7 U/L), and total bilirubin (0.3 to 0.2 mg/dL). Restored SOD (13 to 24 U/g) and CAT (5 to 17 U/g). | [135] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-de-Cerio, E.; Verardo, V.; Gómez-Caravaca, A.M.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Health Effects of Psidium guajava L. Leaves: An Overview of the Last Decade. Int. J. Mol. Sci. 2017, 18, 897. https://doi.org/10.3390/ijms18040897
Díaz-de-Cerio E, Verardo V, Gómez-Caravaca AM, Fernández-Gutiérrez A, Segura-Carretero A. Health Effects of Psidium guajava L. Leaves: An Overview of the Last Decade. International Journal of Molecular Sciences. 2017; 18(4):897. https://doi.org/10.3390/ijms18040897
Chicago/Turabian StyleDíaz-de-Cerio, Elixabet, Vito Verardo, Ana María Gómez-Caravaca, Alberto Fernández-Gutiérrez, and Antonio Segura-Carretero. 2017. "Health Effects of Psidium guajava L. Leaves: An Overview of the Last Decade" International Journal of Molecular Sciences 18, no. 4: 897. https://doi.org/10.3390/ijms18040897
APA StyleDíaz-de-Cerio, E., Verardo, V., Gómez-Caravaca, A. M., Fernández-Gutiérrez, A., & Segura-Carretero, A. (2017). Health Effects of Psidium guajava L. Leaves: An Overview of the Last Decade. International Journal of Molecular Sciences, 18(4), 897. https://doi.org/10.3390/ijms18040897