Abstract
Today, there is increasing interest in discovering new bioactive compounds derived from ethnomedicine. Preparations of guava (Psidium guajava L.) leaves have traditionally been used to manage several diseases. The pharmacological research in vitro as well as in vivo has been widely used to demonstrate the potential of the extracts from the leaves for the co-treatment of different ailments with high prevalence worldwide, upholding the traditional medicine in cases such as diabetes mellitus, cardiovascular diseases, cancer, and parasitic infections. Moreover, the biological activity has been attributed to the bioactive composition of the leaves, to some specific phytochemical subclasses, or even to individual compounds. Phenolic compounds in guava leaves have been credited with regulating blood-glucose levels. Thus, the aim of the present review was to compile results from in vitro and in vivo studies carried out with guava leaves over the last decade, relating the effects to their clinical applications in order to focus further research for finding individual bioactive compounds. Some food applications (guava tea and supplementary feed for aquaculture) and some clinical, in vitro, and in vivo outcomes are also included.
1. Introduction
Ethnomedicine, which refers to the study of traditional medical practice, is an integral part of the culture and the interpretation of health by indigenous populations in many parts of the world [1]. For example, Indian Ayurveda and traditional Chinese medicine are among the most enduring folk medicines still practiced. These systems try to promote health and improve the quality of life, with therapies based on the use of indigenous drugs of natural origin [2]. Given that plants have been widely used as herbal medicines, several approaches are now being carried out to discover new bioactive compounds [3].
Psidium guajava L., popularly known as guava, is a small tree belonging to the myrtle family (Myrtaceae) [4]. Native to tropical areas from southern Mexico to northern South America, guava trees have been grown by many other countries having tropical and subtropical climates, thus allowing production around the world [5]. Traditionally, preparations of the leaves have been used in folk medicine in several countries, mainly as anti-diarrheal remedy [6]. Moreover, other several uses have been described elsewhere on all continents, with the exception of Europe [6,7,8]. Figure 1 summarizes the main traditional uses of guava leaves in the main producer countries. Depending upon the illness, the application of the remedy is either oral or topical. The consumption of decoction, infusion, and boiled preparations is the most common way to overcome several disorders, such as rheumatism, diarrhea, diabetes mellitus, and cough, in India, China, Pakistan, and Bangladesh [6,7,8,9], while in Southeast Asia the decoction is used as gargle for mouth ulcers [6,8,9] and as anti-bactericidal in Nigeria [8,9]. For skin and wound applications, poultice is externally used in Mexico, Brazil, Philippines, and Nigeria [6,7,8,9]. In addition, chewing stick is used for oral care in Nigeria [9].
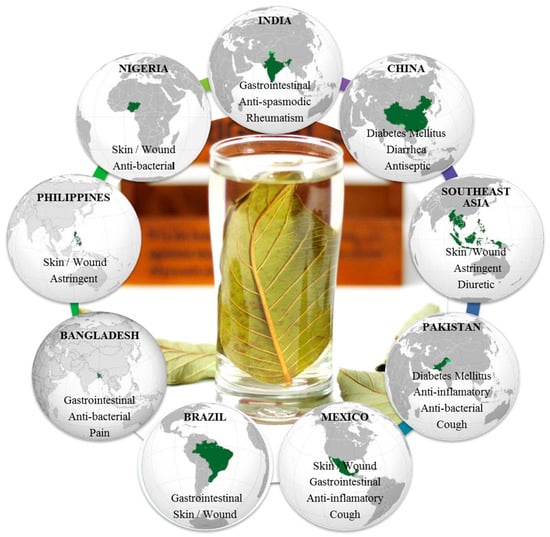
Figure 1.
Main traditional uses of guava leaves in the principal producer countries.
Currently, there is increasing interest in studying of plants regarding their chemical components of bioactive compounds, their effects on several diseases, and their use for human health as functional foods and/or nutraceuticals [10]. In recent years, guava leaves tea and some complementary guava products are available in several shops in Japan as well as on the Internet [11], because guava leaf phenolic compounds have been claimed to be food for specified health use (FOSHU), since they have beneficial health effects related to the modulation of blood–sugar level [12]. Thus, the aim of this review was to summarize the biological activities, in vitro and in vivo, studied in the last decade on P. guajava L. leaves, relating them to the international classification of diseases provided by the World Health Organization. In addition, the beneficial effects of some applications of guava leaves are also been examined. For this purpose, a comprehensive review of the literature from 2004 to 2016 was done, although more recent studies have also been included. Reviewed journals, websites, books, and several databases as “Scopus”, “Google Scholar”, “PubMed”, and “ScienceDirect”, were used to compile them. To ensure that relevant works are included, terms such as “Psidium guajava”, “guava”, “leaves”, “in vitro”, “in vivo”, “clinical”, “trial”, “food application”, and those related with the diseases such as “bacteria”, “cancer”, “blood”, “glycaemia”, and “oral”, among others were matched in the search. Only complete available works published in English, Spanish, and Portuguese have been included.
2. Pharmacological Properties
2.1. In Vitro Studies
2.1.1. Infectious and Parasitic Diseases
Aqueous and organic extracts of guava leaves have been demonstrated to have antibacterial activity due to an inhibitory effect against antibiotics-resistant clinical isolates of Staphylococcus aureus strains [13,14]. Despite using the same diffusion method, differences are noticed in their inhibition zones, as shown in Table 1, probably due to extraction method or the dose assayed. A methanol extract exerted antibacterial effects, preventing the growth of different strains from several bacteria such as Staphylococcus aureus, Escherichia coli, Pseudomonas aeruginosa, Proteus spp., and Shigella spp. [15]. Furthermore, different extracts of the leaves such as aqueous, acetone–water, methanolic, spray-dried extracts, and the essential oil, showed potential inhibitory activity against Gram-positive and Gram-negative bacteria and fungi [16,17,18,19,20]. In these works, it is noticeable that Gram-positive bacteria exhibited greater inhibition zones and minimum inhibitory concentrations (MICs) than Gram-negative. Concerning the anti-fungal activity, less inhibition than bacteria is reported [16,17], except for Candida krusei and Candida glabrata which provided higher inhibition [18], and for Aspergillus spp. for which no activity was found [16] (Table 1). Moreover, Bezerra et al. [21] evaluated the effect of guava leaves on different bacterial strains, concluding that the synergistic action between the leaves and various antibiotics boosted its anti-bacterial activity. This effect was also observed by Betoni et al. [22] with target drugs for the protein synthesis, cell-wall synthesis, and folic acid. However, the latter did not find synergic effect with gentamicin, perhaps because the time of maceration was lower than the time used by Bezerra et al. [21], and also the solvent was different (Table 1).

Table 1.
In vitro assays against infectious and parasitic diseases.
Metwally et al. [23] associated the antimicrobial activity against some bacteria and fungi with five flavonoids isolated from the leaves. This effect was also related to the concentration of tannins in the leaves [24] and to the content of gallic acid and catechin [19]. Additionally, the activity against bacterial and fungal pathogens was traced to betulinic acid and lupeol [25]. In fact, these works are focused on the activity of these compounds, rather than on the effect of the whole extract of the leaves.
In addition, leaf acetone extract of P. guajava has also exhibited moderate acaricidal and insecticidal activities causing the dead of Hippobosca maculata adult fly [26].
Furthermore, Adeyemi et al. [27] suggested that an ethanol extract from the leaves function as a trypanocide agent, since its inhibition of Trypanosoma brucei brucei growth proved similar to that of the reference drugs. Kaushik et al. [28] proposed the leaves as an anti-malaria agent, due to their inhibitory activity and the resistance indices. Furthermore, the effect of guava leaf essential oil against toxoplasmosis caused by the growth of Toxoplasma gondii were reported [29]. Additionally, guava leaves were proposed for the treatment of diarrhea caused by enteric pathogens, since it showed significant inhibitory activity against Vibrio cholerae and V. parahemolyticus, Aeromonas hydrophila, Escherichia coli, Shigella spp. and Salmonella spp. [30,31,32]. It is suppose that the same plant origin and similar extraction procedure makes that these works show comparable inhibition zones for the bacteria tested [30,31], in contrast to the leaves of India and Bangladesh, where MIC values did not show any concordance [31,32] (Table 1). In addition, a reduction was described for S. flexneri and V. cholera invasion and for their adherence to the human laryngeal epithelial cells, and for the production of E. coli heat labile toxin and cholera toxin, as well as their binding to ganglioside monosialic acid enzyme [33]. Moreover, other studies also demonstrated the antimicrobial effect of some bacteria that cause gastrointestinal disorders by different methods [34,35]. In contrast to previous results [20,31], no inhibition of the hydrodistillation and n-hexane extract was found against E. coli Salmonella spp. [31] (Table 1).
Furthermore, guava leaf tea helped control of the growth of influenza viruses, including oseltamivir-resistant strains, via the prevention of viral entry into host cells, probably due to the presence of flavonols [36].
2.1.2. Neoplasms
All the results published regarding anti-cancer properties have been summarized in Table 2.
Table 2.
In vitro studies against neoplasm.
Kawakami et al. [37] evaluated the anti-proliferative activity of guava leaf extract in human-colon adenocarcinoma cell line (COLO320DMA). These authors found that the extract depressed the proliferation rate due to the presence of quercetin and quercetin glycosides. Moreover, different extracts were tested on three cancer cell lines (cervical cancer (HeLa), breast cancer (MDA-MB-231), and osteosarcoma (MG-63)). The extracts showed no anti-proliferative activity towards HeLa cells, although they displayed activity against MDA-MB-231 and MG-63, the ether extract being the most effective, followed by methanol and water extracts. However, ether and methanol extracts presented a cytotoxic effect on non-malignant cell Madine Darby canine kidney (MDCK) [38]. In contrast, an ethanol extract from the stem and leaves reported significant anti-tumor activity on HeLa and colorectal carcinoma (RKO-AS45-1), whereas its effect was less significant for a lung fibroblast cell line (Wi-26VA4) [39]. This difference could be due to the origin of the leaves, compounds in the steam, or even to the extraction method selected. In this context, an organic guava leaf extract provided molecular evidence of cytotoxic or anti-tumor activity in human breast carcinoma benign cells (MCF-7) and also in murine fibrosarcoma (L929sA) [40]. A fact worthy to comment is that the difference noticed in the cytotoxic effect on MDA-MB-231 cell line might be because the extraction differs [38,40]. Furthermore, the aqueous extract of budding guava leaves displayed an anti-tumor effect against human prostate epithelial (PZ-HPV-7) and carcinoma (DU-145) cells in view of the cell-killing-rate coefficients, as well as anti-angiogenesis and anti-migration activities, respectively [41,42].
Regarding the bioactivity of terpenes from guava, an enriched mixture of guajadial, psidial A, and psiguadial A and B proved anti-proliferative effect for nine human cancer lines: leukemia (K-562), breast (MCF-7), resistant ovarian cancer (NCI/ADR-RES), lung (NCI-H460), melanoma (UACC-62), prostate (PC-3), colon (HT-29), ovarian (OVCAR-3), and kidney (786-0) [43]. The apoptotic effect of β-caryophyllene oxide (CPO) on MCF-7 and PC-3 cell lines was also demonstrated because of its ability to interfere with multiple signaling cascades involved in tumor genesis [44]. Moreover, the essential oil from guava leaves exerted an anti-proliferative effect on human-mouth epidermal carcinoma (KB) and murine leukemia (P388) cell lines [45], while a hexane fraction of the leaves showed a cytotoxic effect against leukemia (Kasumi-1) cancer-cell line at higher half maximal inhibitory concentration (IC50), probably due to a less concentration of the bioactive compounds of the leaves [46]. Finally, cytotoxic and apoptotic effect in PC-3 cells and apoptotic effect in LNCaP cells was reported. The lack of cytotoxic effect in LNCaP might be because the cell growth is androgen-dependent, while in PC-3 is androgen-independent. [47]. Comparing these data with those reported by Park et al. [44], high concentration is needed for causing cell death, and a weak effect is found on early apoptotic cell. The main difference between these works is the composition of the extract, so it could be concluded that an antagonist effect is produced amongst the isolated compounds by Ryu et al. [47].
2.1.3. Diseases of the Blood and Immune System
A fermented guava leaf extract was tested in mouse macrophage (RAW 264.7) cells. The results confirmed its potential to decrease the expression of lipopolysaccharide-inducible nitric oxide synthase and cyclooxygenase-2 proteins level, two pro-inflammatory mediators, through the down-regulation of nuclear factor-κB transcriptional activity (NF-κB) [48]. This biological activity was also reported in other works [40,49,50]. Briefly, Jang et al. [49] evaluating the prostaglandin E2 production found that the inhibitory effect was highly correlated to the total phenolic content. Kaileh et al. [40] suggested that the suppression of the nuclear factor-κB could be at the transcriptional level because of the lack of binding between nuclear factor-κB and DNA in murine fibrosarcoma (L929sA) and two breast-cancer cell lines (MDA-MB231 and MCF7). At the same time, Jang et al. [50] found that the lipopolysaccharide-induced production of nitric oxide and prostaglandin E2 was due to the ability of guava leaf extract to suppress phosphorylation in protein expression. Moreover, Sen et al. [51] verified the inhibition of nuclear factor-κB activation in Labeo rohita head-kidney macrophages by the flavonoid fraction of guava leaf extract and Jang et al. [52] improved the inhibition of lipopolysaccharide-induced prostaglandin E2 and nitric oxide production by optimizing of the extraction conditions. Furthermore, methanol and ethanol leaf extracts also showed the inhibition of hypotonicity-induced lysis of erythrocyte membrane [53]. Meanwhile, Laily et al. [54] suggested the use of guava leaves as immune-stimulant agent because they modulated the lymphocyte proliferation response.
The results for this activity, confirm the potential of guava leaves as an anti-inflammatory treatment and as immune-system stimulatory agent. As is shown in Table 3, a general trend is reported in every work, although the differences noticed in the data are probably due to the different extraction method and to the doses assayed, or even to the harvesting time of the leaves. However, the mechanism should be further studied since two different pathways are suggested for the down-regulation of NF-κB.
Table 3.
In vitro assays against diseases of the blood and immune system.
2.1.4. Endocrine and Metabolic Diseases
Several works have focused on elucidating the anti-diabetic compounds present in guava leaves (Table 4). Although the origin of the leaves remains different, the presence of these compounds has demonstrated the hypoglycemic effect of the leaves via different assays. However, the main mode of action seems to be due to an inhibition of the enzymes related to this activity.
Table 4.
Compounds in guava leaves with anti-diabetic properties in in vitro assays.
The anti-glycative potential of the guava leaves was evaluated, with the conclusion that the extract inhibited, in vitro, the formation of advanced glycation end-products formation [55]. Moreover, the aqueous guava leaf extract, in an albumin/glucose model system, also exerted the same effect and indeed inhibited Amadori products. Gallic acid, catechin and quercetin exhibited over 80% inhibitory effects whereas ferulic acid showed no activity [56]. In another study, seven pure flavonoid compounds (quercetin, kaempferol, guaijaverin, avicularin, myricetin, hyperin, and apigenin) showed strong inhibitory activities against sucrase, maltase, and α-amylase, and a clear synergistic effect against α-glucosidase [57]. Moreover, Deguchi and Miyazaki [58] suggested that the component that inhibited the in vitro activities of α-glucosidase enzymes in guava extract was a polymerized polyphenol. In addition, polysaccharides from guava leaves also exhibited α-glucosidase inhibition [59].
Eidenberger et al. [60] demonstrated the dose-dependent inhibition of guava leaf ethanol extracts on dipeptidyl-peptidase-IV due to the individual flavonol-glycosides: peltatoside, hyperoside, methylquercetin hexoside, isoquercitrin, quercetin/morin pentoside, guaijaverin, and quercetin/morin pentoside. Additionally, the individual flavonol-glycosides found in the guava extract reported no significant differences compared with the uptake of the whole guava extract into epithelial cells (CaCo-2) [60]. In the same cell line, the inhibition of fructose uptake was also tested by Lee et al. [61], who confirmed that catechin and quercetin contributed to the inhibition of glucose transporters. In addition, the enhancement of aqueous guava leaf extract was investigated with regard to glucose uptake in rat clone 9 hepatocytes. Moreover, quercetin was proposed as the active compound responsible for promoting glucose uptake in liver cells and contributing to the alleviation of hypoglycemia in diabetes [62]. Furthermore, Basha and Kumari [63] also estimated the glucose uptake of different extracts. The methanol extract of guava leaves was found to be the most efficient in lowering glucose levels. Basha et al. [64] demonstrated the ability of guavanoic-acid-mediated gold nanoparticles to inhibit the protein tyrosine phosphatase 1B activity.
Indeed, a guava leaf ethanol extract was tested in pre-adipocyte cell line (3T3-L1), which showed its ability to inhibit adipocyte differentiation via down-regulation of adipogenic transcription factors and markers, and hence may prevent obesity in vivo [65]. To evaluate the potential of the leaves on glucose uptake and glycogen synthesis, an aqueous extract was used in insulin-resistant mouse (FL83B) cells. The results confirmed the improved expression and phosphorylation of insulin signaling-related proteins, promoting glycogen synthesis and glycolysis pathways. In fact, this work provides new insights into the mechanisms through which the guava extract improves insulin resistance in the hepatocytes [66]. In the same cell line, vescalagin was postulated as the active component that may alleviate the insulin resistance in mouse hepatocytes [67].
In this sense, the latest study made in L6 myoblasts and myotubes cells confirmed that the glucose uptake recruitment followed a wortmannin-dependent pathway. In addition, guava leaves also inhibited aldose reductase activity, up-regulated gene- and protein-level expression of several insulin receptors and also improved cellular-level glucose uptake [68].
2.1.5. Diseases of the Circulatory System
Cardiovascular disorders have been related to the endothelial cell damage that causes atherosclerosis. In this sense, extracts from budding guava leaves demonstrated a protective, in vitro, effect in bovine aortal endothelial cells, delaying low-density lipoprotein oxidation and preventing oxidized low-density lipoprotein cytotoxicity [69]. A similar effect was also noted in human umbilical-vein endothelial cell due to the ability of saving cell-viability reduction, suppressing reactive oxygen species production and nitric oxide release, as well as inhibiting the expression of NF-κB [70]. Moreover, budding guava leaves also showed their ability as an anticoagulant in plasma, since they reduced thrombin clotting time and inhibited the activity of antithrombin III. Thus, they could help to reduce the development of cardiovascular complications [71].
In addition, flavonoids and phenolic acids in the leaves could contribute to the prevention and amelioration of gout and hypertension, since, in rat-tissues homogenates, they inhibit the activity of two enzymes related to the development of both diseases (xanthine oxidase and angiotensin 1-converting enzymes) [72].
2.1.6. Diseases of the Digestive System
Guaijaverin, isolated from guava leaves, displayed high inhibitory activity against Streptococcus mutans. In fact, guaijaverin exhibited its ability as an anti-plaque agent, becoming an alternative for oral care [73]. Furthermore, guava leaves showed greater bactericidal effect on early (Streptococcus sanguinis) and late (S. mutans) colonizers compared to Mangifera indica L. and Mentha piperita L. leaves, whereas, when they are compared with the plant extract mixture, the effect is slightly lower. By contrast, guava leaves showed similar and higher anti-adherence effect than the plant mixture [74]. In another study, the whole extract was tested on the cell-surface hydrophobicity of selected early settlers and primary colonizers of dental plaque, showing its ability to alter and disturb the surface characteristics of the agents, making them less adherent [75,76,77], and also delayed in the generation of dental biofilm by targeting growth, adherence, and co-aggregation [78]. This property could be due to the presence of flavonoids and tannins detected in P. guajava [79]. Shekar et al. [80] also confirmed the use of the leaves as anti-plaque agents against Streptococcus mutans, S. sanguinis, and S. salivarius. Kwamin et al. [81] discovered the effectiveness of guava leaf extract in the leukotoxin neutralization of Aggregatibacter actinomycetemcomitans, leading it to be considered as a possible agent for the treatment of aggressive forms of periodontitis. In addition, extracts rich in guava flavonoids have demonstrated their potential for preventing dental caries due to the growth inhibition of the oral flora [82]. Moreover, its soothing of toothache has been verified based on the analgesic, anti-inflammatory, and anti-microbial activity properties [83] and it has been reviewed positively as an adjutant for treating periodontal disease [84].
Concerning the liver disorders, the cytotoxic and hepato-protective effects of guava leaves were reported. Studies carried out in clone 9 cells treated with different extracts of the leaves showed that only ethanol and acetone extracts tend to have cytotoxicity effect at high concentrations. Moreover, the ethanol extract showed hepato-protective activity, although the hot-water extract reported greater effect and lower cytotoxicity [85].
Table 5 compiles the methodology followed and the results reported in the present works. It is important to keep in mind that the origin, the selection of the extraction method or solvent, and the concentration of the extract tested generally provide different data. For example, comparing data for inhibition zones, best results are noticed at long maceration time in acetone, which seems to be a better extracting solvent than ethanol [77,78,80,82]. Hydrophobicity depends on the origin of the leaves, the extraction method, and the concentration of the extract tested, and it also depends on the lipophilic (index > 70%) or hydrophilic nature of the strain [73,75,79]. Finally, minimum inhibitory concentration relies on all factors.
Table 5.
In vitro assays against diseases related to the digestive system.
2.1.7. Diseases of the Skin and Subcutaneous Tissue
Qa’dan et al. [86] described the antimicrobial effect of a leaf extract against the main developer of acne lesions, Propionibacterium acnes, and other organisms isolated from acne lesions. The antimicrobial activity was also displayed against pathogenic bacteria associated with wound, skin, and soft-tissue infections [87]. Furthermore, antifungal properties have also been studied by Padrón-Márquez et al. [88]. The acetone and methanol extracts displayed relevant activity against dermatophytic fungi, and thus could be considered as new agents against skin disease. Furthermore, phenols from the leaves were tested on human-skin fibroblast cells and showed antifungal properties [89].
In addition, the tyrosinase inhibitory activities of 4 different parts (branch, fruit, leaf, and seed) of guava, extracted with acetone, ethanol, methanol, and water were tested by You et al. [90] who reported that the ethanol extract from the leaves reached the highest activity. Therefore, the leaves might be appropriate for both boosting the whitening of skin and inhibiting browning. In addition, in a human keratinocyte cell line, an ethyl acetate extract showed a positive effect on atopic dermatitis via the inhibition of cytokine-induced Th2 chemokine expression [91].
Lee et al. [92] carried out the first electrophysiological study based on ultraviolet (UV)-induced melanogenesis with guava leaves. The authors suggested the use of guava leaves for both direct and indirect prevention of skin melanogenesis caused by UV radiation. In fact they demonstrate that methanolic guava leaves extract inhibits tyrosinase, that is the key enzyme in melanin synthesis, and ORAI1 channel that has shown to be associated with UV-induced melanogenesis.
2.1.8. Other Activities Related to Several Diseases
An aqueous guava extract showed its ability to decrease the radiolabeling of blood constituent due to an antioxidant action and/or because it alters the membrane structures involved in ion transport into cells [93]. Guava leaves also have been demonstrated to possess anti-allergic effects in rat mast (RBL-2H3) cell line by the inhibition of degranulation and cytokine production, as well as blocking high-affinity immunoglobulin E-receptor signaling [94].
2.2. In Vivo Studies
2.2.1. Infectious and Parasitic Diseases
After checking the effect of guava leaf extract, in vitro, against Aeromonas hydrophila, in vivo experiments were carried out in tilapia (Oreochromis niloticus), indicating the potential use of P. guajava as environmentally friendly antibiotic [95]. The leaves also had anti-viral and anti-bacterial activity towards shrimp (Penaeus monodon) pathogens such as yellow-head virus, white spot syndrome virus, and Vibrio harvey. In addition, guava leaf extract improved the activities of prophenoloxidase and nitric oxide synthase in serum, and of superoxide dismutase, acid phosphatase, alkaline phosphatase, and lysozyme in serum and hepatopancreas [96].
Furthermore, guava leaves have been suggested for managing sleeping sickness, since they exhibited trypanocidal effect in albino rats [97]; the extract ameliorate the tissue-lipid peroxidation associated to trypanosomosis, as well as raising the level of the glutathione concentration [98]. The leaves also showed anti-malarial effect in BALB/c mice infected with Plasmodium berghei via parasitemia suppression [99]. Moreover, guava leaves are also recommended for treating infectious diarrhea since they prevented intestinal colonization of Citrobacter rodentium in Swiss albino mice [100]. In chicks, guava leaf extract enabled the control of diarrhea produced by E. coli and reduced the severity of its symptomatology [101]. In mice, the improvement of cholera symptoms caused by V. cholerae, a human pathogen, was also confirmed by Shittu et al. [102].
In addition, anti-helminthic properties towards gastro-intestinal nematodes have been found, as a result of the presence of condensed tannins in the guava plant, which raised the levels of hemoglobin, packed cell volume, total protein, globulin, glucose, and calcium, and lowered the levels of blood urea [103].
All the results published regarding in vivo anti-bacterial properties have been summarized in Table 6.
Table 6.
In vivo anti-bacterial effect.
2.2.2. Neoplasms
Only one study is available on the anti-tumor effect that could be related to the phenolic composition of guava leaves. An ethanol extract of the leaves was administrated to B6 mice after inoculation of melanoma cells. The results suggested that the extract had a vaccine effect, but not a therapeutic effect, against tumors through by depressing T regulatory cells [104].
Moreover, the meroterpene-enriched fraction of guava leaves, containing guajadial, psidial A, and psiguadial A and B, was evaluated in vivo in a solid Ehrlich murine breast-adenocarcinoma model. The results suggested that these compounds may act as phytoestrogens, presenting tissue-specific antagonistic and agonistic activity on estrogen receptors [43]. These data partially confirmed the results in vitro obtained by Ryu et al. [47].
2.2.3. Diseases of the Blood and Immune System
Among blood diseases, anemia indicates a failure in the immune system. In this sense, guava extract presented an anti-anemic effect in trypanosomosis-infected Wistar rats, improving the values of hemoglobin, packed cell volume, red-blood cell counts, mean corpuscular volume, and mean concentration hemoglobin count while decreasing white-blood cell and neutrophil levels [105]. Moreover, the same trend in the hematological analyses was also recorded in mice. After the administration of guava leaf extract, no alterations on the erythron were detected [106]. Nevertheless, results differ because subjects under study are different, also the duration of the treatment, the extraction method and the dose assayed (Table 7).
Table 7.
In vivo studies against diseases of the blood and immune system.
The anti-inflammatory response of the leaves was dose-dependent in induced hyperalgesia in Sprague-Dawley rats, decreasing in paw-withdrawal latency, and significantly improving the survival rate of mice with lethal endotoxemia [50]. Moreover, the anti-inflammatory activity of aqueous and acetone–water extracts of the leaves was also confirmed in Swiss mice by reducing the amount of leukocyte migration. The acetone–water extract also exhibited peripheral analgesic activity, probably by blocking the effect or the release of endogenous substances that excite pain-nerve endings [19]. The analgesic effect in albino rats was also reported. The ethanol extract reduced the writhing response [107], and a jumping response was found after the administration of a distilled extract (combination of methanol and aqueous extracts) [108]. In this case, the writhing response for both Swiss mice and Wistar rats seems to be comparable, although the dose assayed is completely different (Table 7).
2.2.4. Endocrine and Metabolic Diseases
Guava leaves have shown their potential against one of the diseases with the highest incidence level worldwide, diabetes mellitus, and also towards biochemical changes caused by the disease. In spite of being leaves from different countries, treatments in different subjects or even different data, the same trend is followed in these works (Table 8).
Table 8.
Endocrine and metabolic in vivo assays with guava leaves.
The effect of aqueous guava leaf extract was investigated in rabbits, fed a high-cholesterol diet. Treatment with guava leaves reduced the plasma-cholesterol level, caused a remarkable spike in high-density lipoprotein, a dip in low-density lipoprotein levels, and significantly reduced the associated hyperglycemia. In addition, the extract showed hypolipidemic and hypoglycemic potentials in hypercholesterolemic rabbits [109]. Furthermore, guava leaves reduced oxidative stress induced by hypercholesterolemia in rats [110].
In addition, the anti-diabetic effect was also evaluated in Leprdb/Leprdb mice and significant blood-glucose-lowering effects were observed. In addition, histological analysis revealed a significant reduction in the number of lipid droplets, which, furthermore, at least in part, could be mediated via the inhibition of protein tyrosine phosphatase 1B [111].
In streptozotocin-induced diabetic rats, the administration of oral doses of aqueous and ethanol extracts from guava leaves could alter the Ca:Mg ratio [112]; however, in low-dose streptozotocin and nicotinamide-induced Sprague-Dawley diabetic rats, long-term administration of guava leaf extracts raised the plasma-insulin level, the glucose utilization, and the activity of hepatic enzymes [113]. Moreover, the leaves also lowered blood glucose levels and decreased protein glycation [55].
In agreement with the above, a lower blood-glucose level was also reported in alloxan-induced diabetic rats. Additionally, no side effects were observed in certain liver enzymes (alkaline phosphatase and aspartate aminotransferase) whereas alanine aminotransferase activity declined [114]. In alloxan-induced diabetic rats, a decrease was also found in blood glucose, total cholesterol, triglycerides, low-density lipoprotein cholesterol, very low-density lipoprotein cholesterol, and a significant increase in high-density lipoprotein cholesterol after 21 days of treatment with guava leaf ethanolic extract [115].
Among the works that evaluated only biochemical parameters, guava leaf extract promoted changes due to an alteration on the activity of alkaline phosphatase, aspartate aminotransferase, alanine aminotransferase, and acid phosphatase in the kidney, liver, and serum [106,116]. In addition, Adeyemi and Akanji [117] evaluated the effect of daily administration of guava leaves, demonstrating the alteration of the serum homeostasis and the pathological variations in rat tissues.
2.2.5. Diseases of the Circulatory System
Ademiluyi et al. [118] assessed the lipid peroxidation in rats after checking the antihypertensive effect, in vitro, of red and white guava leaves. The work concluded that the activity may be related to rosmarinic acid, eugenol, carvacrol, catechin, and caffeic acid since they were the major constituents of their extracts. In addition, this activity was supported by the biphasic and contractile effect on rat vascular smooth muscles [119,120].
In addition, atherosclerosis development was reduced in apoE-knockout mice by guava leaf extracts. In fact, the effect was connected to the presence of ethyl gallate and quercetin [121,122]. In streptozotocin-induced diabetic rats, vascular reactivity to vasoconstrictor agents was reduced, as was vessel atherosclerosis [112]. Furthermore, Soman et al. [123] found that an ethyl acetate fraction of guava leaves reduced cardiac hypertrophy in streptozotocin-induced diabetic rats due to an anti-glycative effect.
2.2.6. Diseases of the Digestive System
In the digestive system, formed by the gastrointestinal tract plus the group organs necessary for the digestion, guava leaves have demonstrated activity towards different parts.
On the one hand, the leaves have shown the ability to protect the stomach against ulceration by inhibiting gastric lesions, reducing gastric secretory volume, and acid secretion, and raising the gastric pH [124,125,126]. This anti-ulcer activity, resulting from the protection of the mucosa, was related to the flavonoids in the leaves [127]. Despite of the subject employed for the assay, similar data are reported in these works (Table 9). The anti-diarrheal activity of guava leaf aqueous extract was evaluated on experimentally induced diarrhea in rodents. The extract performed in the same way as the control drugs, offering protection, inhibiting intestinal transit, and delaying gastric emptying [128]. Another study attributed this activity to a dual action between the antimicrobial effect and the reduction in gastrointestinal motility ability of the extract [129]. In rabbits, the anti-spasmodic effects were connected to a calcium channel blocking activity, which explains the inhibitory effect on gut motility. The anti-diarrheal protection was also tested in mice [130]. As is shown in Table 9, the anti-diarrheal activity is dose-dependent, although the protection varied depending on the subjects.
Table 9.
In vivo assays for digestive system related diseases.
On the other hand, guava leaves exhibited hepato-protective effect due to the reduction of serum parameters of hepatic enzymes markers and histopathological alterations in the acute liver damage induced in rats [131,132,133,134,135]. Here, a dose-dependent effect is also found. However, decoction of the leaves seems to be the best option for the extraction of the compounds that exhibited this activity (Table 9).
2.2.7. Diseases of the Skin and Subcutaneous Tissue
Guava leaves have been suggested as a therapeutic agent to control pruritus in atopic dermatitis. The improvement of the skin lesions was due to a reduction in serum immunoglobulin E level and in the eczematous symptoms [136]. Moreover, the epithelium was repaired with connective tissue and absence or moderate presence of inflammatory cells by the leaves. As a result, the leaves exhibited wound healing properties [137]. Furthermore, guava leaf extract was tested on rat skin, and exhibited inhibitory activity towards an active cutaneous anaphylaxis reaction [138].
2.2.8. Other Activities Related to Several Diseases
Triterpenoids from guava leaves were suggested as a potential therapeutic approach for treating diabetic peripheral neuropathy, as they enhanced physical functions and offered neuronal protection towards the suppression of the expression of pro-inflammatory cytokines [139]. In addition, the leaves can act as radio modulators for cancer patients because by preventing DNA damage and apoptosis. [140], as well as protective agents by restoring the normal values of sperm viability, sperm count, sperm motility, and sperm-head abnormality caused by caffeine-induced spermatotoxicity [141].
Moreover, the consumption of guava leaf tea was evaluated, in vivo, in the inhibition of cytochrome P450 (CYP) 3A-mediated drug metabolism by the interaction between guava tea and several drugs [11,142]. Matsuda et al. [11] investigated the consequence of the ingestion of guava tea for two weeks in rats, and the effect with an anxiolytic drug. The short-term consumption of the tea had little effect on the assays performed. This weak influence was due to the absence of interaction between the tea and midazolam in the metabolism studied. In addition, two in vivo studies were made in rats, to evaluate the interaction of guava leaf tea with an anti-coagulant drug (warfarin) [142]. Kaneko et al. [141] suggested that because the tea contained compounds that block the affinity between the enzyme and phenolic compounds of the tea, long-term administration showed a low probability of causing drug-metabolizing enzymes. Moreover, short-term administration revealed that the tea did not interfere with coagulation, meaning that the tea consumption did not alter the pharmacological effect and displayed no side effects.
2.3. Clinical Trials
To test the effect of guava leaf extract, several randomized clinical trials have been conducted during the last two decades, although only two studies are available in the last decade. One of the studies consisted of evaluating the effect of guava leaf extract pills on primary dysmenorrhea disorder. For this, 197 women were divided into four groups, and each received a different dosage: 3 and 6 mg extract/day, 300 mg placebo/day and 1200 mg ibuprofen/day. The administration took place over five days during three consecutive cycles. The results demonstrated that 6 mg extract/day alleviated menstrual pain and could replace the use of medicaments like ibuprofen. In fact, guava leaves could be used as a broad-spectrum phyto-drug and not only as an anti-spasmodic agent [143]. Furthermore, Deguchi and Miyazaki [58] reviewed several works regarding the effect of the intake of a commercial guava leaf tea (Bansoureicha®, Yakult Honsha, Tokyo, Japan) on different pathologies of diabetes mellitus illness such as the influence on postprandial blood glucose, on insulin resistance and on hypertriglyceridemia and hypercholesterolemia. The authors concluded that the ingestion of guava leaf tea can ameliorate the symptoms of diabetes mellitus and that it could be used as an alimentotherapy.
3. Other Applications
Further applications found with guava leaves are listed below: firstly, to prepare gelatin beads with marine-fish gelatin for various applications such as medicine, and the food and pharmaceutical industries [144]. Secondly, Giri et al. [145] suggested guava leaves as supplementary feed for the fish species Labeo rohita, due to the immune-stimulatory effect. The same conclusion was reached by Fawole et al. [146] in L. rohita. Thirdly, Gobi et al. [147] reported that guava leaf powder, mixed with a commercial diet, strengthened the immunological response of Oreochromis mossambicus, and recommended the leaves as feed complement in aquaculture.
4. Conclusions
Traditional claims generally require experimental research to establish their effectiveness. In this regard, ethnomedicine applications of Psidium guajava L. leaves have been verified by several researches over the last decade against many disorders, demonstrating its potential in the treatment of the most common worldwide diseases. In addition, the effects of the leaves have been related to individual compounds such as quercetin, catechin, vescalagin, gallic acid, peltatoside, hyperoside, isoquercitrin, and guaijaverin.
Future prospects should be aimed at investigating the biodiversity of guava and/or the purification of the different compounds present in guava leaves in order to obtain functional ingredients for further uses as alternative agents in natural therapeutic approaches.
Acknowledgments
The author Vito Verardo thanks the Spanish Ministry of Economy and Competitiveness (MINECO) for “Ramony Cajal” post-doctoral contract.
Author Contributions
Elixabet Díaz-de-Cerio contributed to the literature review and manuscript redaction; Vito Verardo and Ana María Gómez-Caravaca contributed to the conception of the idea and framework writing; and Alberto Fernández-Gutiérrez and Antonio Segura-Carretero supervised the progress of work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Anyinam, C. Ecology and ethnomedicine: Exploring links between current environmental crisis and indigenous medical practices. Soc. Sci. Med. 1995, 40, 321–329. [Google Scholar] [CrossRef]
- Patwardhan, B.; Warude, D.; Pushpangadan, P.; Bhatt, N. Ayurveda and traditional Chinese medicine: A comparative overview. Evid. Based Complement. Altern. Med. 2005, 2, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Fabricant, D.S.; Farnsworth, N.R. The value of plants used in traditional medicine for drug discovery. Environ. Health Perspect. 2001, 109, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.F. Fruits of Warm Climates; Creative Resource Systems, Inc.: Winterville, NC, USA, 1987. [Google Scholar]
- Salazar, D.M.; Melgarejo, P.; Martínez, R.; Martínez, J.J.; Hernández, F.; Burguera, M. Phenological stages of the guava tree (Psidium guajava L.). Sci. Hortic. 2006, 108, 157–161. [Google Scholar] [CrossRef]
- Gutiérrez, R.M.P.; Mitchell, S.; Solis, R.V. Psidium guajava: A review of its traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2008, 117, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Shruthi, S.D.; Roshan, A.; Sharma, S.; Sunita, S. A review on the medicinal plant Psidium guajava Linn. (Myrtaceae). J. Drug Deliv. Ther. 2013, 3, 162–168. [Google Scholar]
- Morais-Braga, M.F.B.; Carneiro, J.N.P.; Machado, A.J.T.; dos Santos, A.T.L.; Sales, D.L.; Lima, L.F.; Figueredo, F.G.; Coutinho, H.D.M. Psidium guajava L., from ethnobiology to scientific evaluation: Elucidating bioactivity against pathogenic microorganisms. J. Ethnopharmacol. 2016, 194, 1140–1152. [Google Scholar] [CrossRef] [PubMed]
- Sanda, K.A.; Grema, H.A.; Geidam, Y.A.; Bukar-Kolo, Y.M. Pharmacological aspects of P. guajava: An update. Int. J. Pharmacol. 2011, 7, 316–324. [Google Scholar]
- Bernal, J.; Mendiola, J.A.; Ibáñez, E.; Cifuentes, A. Advanced analysis of nutraceuticals. J. Pharm. Biomed. Anal. 2011, 55, 758–774. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Nishimura, Y.; Kurata, N.; Iwase, M.; Yasuhara, H. Effects of continuous ingestion of herbal teas on intestinal CYP3A in the rat. J. Pharmacol. Sci. 2007, 103, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Yasuoka, A.; Abe, K. Functional food science and food for specified health use policy in Japan: State of the art. Curr. Opin. Lipidol. 2008, 19, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Milyani, R. Inhibitory effect of some plant extracts on clinical isolates of Staphylococcus aureus. Afr. J. Microbiol. Res. 2012, 6, 6517–6524. [Google Scholar] [CrossRef]
- Anas, K.; Jayasree, P.R.; Vijayakumar, T.; Manish Kumar, P.R. In vitro antibacterial activity of Psidium guajava Linn. leaf extract on clinical isolates of multidrug resistant Staphylococcus aureus. Indian J. Exp. Biol. 2008, 46, 41–46. [Google Scholar] [PubMed]
- Chah, K.F.; Eze, C.A.; Emuelosi, C.E.; Esimone, C.O. Antibacterial and wound healing properties of methanolic extracts of some Nigerian medicinal plants. J. Ethnopharmacol. 2006, 104, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.; Chanda, S. In Vitro antimicrobial activity of Psidium guajava L. leaf extracts against clinically important pathogenic microbial strains. Braz. J. Microbiol. 2007, 38, 452–458. [Google Scholar] [CrossRef]
- Dhiman, A.; Nanda, A.; Narasimhan, B. In vitro antimicrobial activity of methanolic leaf extract of Psidium guajava L. J. Pharm. Bioallied Sci. 2011, 3, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.R.V.; Dias, A.L.T.; Carvalho, R.R.; Souza, C.R.F.; Oliveira, W.P. Antioxidant and antimicrobial activities of Psidium guajava L. spray dried extracts. Ind. Crops Prod. 2014, 60, 39–44. [Google Scholar] [CrossRef]
- De Araújo, A.A.; Soares, L.A.L.; Assunção Ferreira, M.R.; de Souza Neto, M.A.; da Silva, G.R.; de Araújo, R.F.; Guerra, G.C.B.; de Melo, M.C.N. Quantification of polyphenols and evaluation of antimicrobial, analgesic and anti-inflammatory activities of aqueous and acetone-water extracts of Libidibia ferrea, Parapiptadenia rigida and Psidium guajava. J. Ethnopharmacol. 2014, 156, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Nisha, K.; Darshana, M.; Madhu, G.; Bhupendra, M.K. GC-MS Analysis and anti-microbial activity of Psidium guajava (leaves) grown in Malva region of India. Int. J. Drug Dev. Res. 2011, 3, 237–245. [Google Scholar]
- Bezerra Morais-Braga, M.F.; Lima Sales, D.; dos Santos Silva, F.; Pereira Chaves, T.; de Carvalho Nilo Bitu, V.; Torres Avilez, W.M.; Ribeiro-Filho, J.; Douglas Melo Coutinho, H. Psidium guajava L. and Psidium brownianum Mart ex DC. potentiate the effect of antibiotics against Gram-positive and Gram-negative bacteria. Eur. J. Integr. Med. 2016, 8, 683–687. [Google Scholar] [CrossRef]
- Betoni, J.E.C.; Passarelli Mantovani, R.; Nunes Barbosa, L.; Di Stasi, L.C.; Fernandes Junior, A. Synergism between plant extract and antimicrobial drugs used on Staphylococcus aureus diseases. Mem. Inst. Oswaldo Cruz 2006, 101, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Metwally, A.M.; Omar, A.A.; Harraz, F.M.; El Sohafy, S.M. Phytochemical investigation and antimicrobial activity of Psidium guajava L. leaves. Pharmacogn. Mag. 2010, 6, 212–218. [Google Scholar] [PubMed]
- Mailoa, M.N.; Mahendradatta, M.; Laga, A.; Djide, N. Antimicrobial activities of tannins extract from guava leaves (Psidium guajava L.) on pathogens microbial. Int. J. Sci. Technol. Res. 2014, 3, 236–241. [Google Scholar]
- Ghosh, P.; Mandal, A.; Chakraborty, P.; Rasul, M.G.; Chakraborty, M.; Saha, A. Triterpenoids from Psidium guajava with biocidal activity. Indian J. Pharm. Sci. 2010, 72, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Zahir, A.A.; Rahuman, A.A.; Bagavan, A.; Santhoshkumar, T.; Mohamed, R.R.; Kamaraj, C.; Rajakumar, G.; Elango, G.; Jayaseelan, C.; Marimuthu, S. Evaluation of botanical extracts against Haemaphysalis bispinosa Neumann and Hippobosca maculata Leach. Parasitol. Res. 2010, 107, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.S.; Sykes, M.L.; Akanji, M.A.; Avery, V.M. Anti-trypanosomal and cytotoxic activity of ethanolic extracts of Psidium guajava leaves in Alamar Blue based assays. Vet. Arh. 2011, 81, 623–633. [Google Scholar]
- Kaushik, N.K.; Bagavan, A.; Rahuman, A.A.; Zahir, A.A.; Kamaraj, C.; Elango, G.; Jayaseelan, C.; Kirthi, A.V.; Santhoshkumar, T.; Marimuthu, S.; et al. Evaluation of antiplasmodial activity of medicinal plants from North Indian Buchpora and South Indian Eastern Ghats. Malar. J. 2015, 14, 65. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.C.; Mahmud, R.; Noordin, R.; Pillai Piaru, S.; Perumal, S.; Ismail, S. Free radicals scavenging activity, cytotoxicity and anti-parasitic activity of essential oil of Psidium guajava L. leaves against Toxoplasma gondii. J. Essent. Oil Bear. Plants 2013, 16, 32–38. [Google Scholar] [CrossRef]
- Chanu, T.R.; Pai, V.; Chakraborty, R.; Raju, B.; Lobo, R.; Ballal, M. Screening for antidiarrheal activity of Psidium guajava: A possible alternative in the treatment against diarrhea causing enteric pathogens. J. Chem. Pharm. Res. 2011, 3, 961–967. [Google Scholar]
- Thiyagarajan, S.; Jamal, A. Evaluation of Lethal Activity of Psidium guajava Linn. extracts on bacterial pathogens causing diarrheal infections. Int. J. Res. Ayurveda Pharm. 2015, 6, 111–117. [Google Scholar] [CrossRef]
- Rahim, N.; Gomes, D.J.; Watanabe, H.; Rahman, S.R.; Chomvarin, C.; Endtz, H.P.; Alam, M. Antibacterial activity of Psidium guajava leaf and bark against multidrug-resistant Vibrio cholerae: Implication for cholera control. Jpn. J. Infect. Dis. 2010, 63, 271–274. [Google Scholar] [PubMed]
- Birdi, T.; Daswani, P.; Brijesh, S.; Tetali, P.; Natu, A.; Antia, N. Newer insights into the mechanism of action of Psidium guajava L. leaves in infectious diarrhoea. BMC Complement. Altern. Med. 2010, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.A.; Andrade Neto, M.; Bezerra, J.N.S.; Macrae, A.; De Sousa, O.V.; Fonteles-Filho, A.A.; Vieira, R.H.S.D.F. Antibacterial activity of guava, Psidium guajava Linnaeus, leaf extracts on diarrhea-causing enteric bacteria isolated from seabob shrimp, Xiphopenaeus kroyeri (Heller). Rev. Inst. Med. Trop. Sao Paulo 2008, 50, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Nwinyi, O.; Chinedu, S.N.; Ajani, O.O. Evaluation of antibacterial activity of Pisidium guajava and Gongronema latifolium. J. Med. Plants Res. 2008, 2, 189–192. [Google Scholar]
- Sriwilaijaroen, N.; Fukumoto, S.; Kumagai, K.; Hiramatsu, H.; Odagiri, T.; Tashiro, M.; Suzuki, Y. Antiviral effects of Psidium guajava Linn. (guava) tea on the growth of clinical isolated H1N1 viruses: Its role in viral hemagglutination and neuraminidase inhibition. Antivir. Res. 2012, 94, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Nakamura, T.; Hosokawa, T.; Suzuki-Yamamoto, T.; Yamashita, H.; Kimoto, M.; Tsuji, H.; Yoshida, H.; Hada, T.; Takahashi, Y. Antiproliferative activity of guava leaf extract via inhibition of prostaglandin endoperoxide H synthase isoforms. Prostaglandins Leukot. Essent. Fatty Acids 2009, 80, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Sul´ain, M.D.; Zazali, K.E.; Ahmad, N. Screening on anti-proliferative activity of Psidium guajava Leaves extract towards selected cancer cell lines. J. US China Med. Sci. 2012, 9, 30–37. [Google Scholar]
- Vieira Braga, T.; Gonçalves Rodrigues das Dores, R.; Soncin Ramos, C.; Gontijo Evangelista, F.C.; Márcia da Silva Tinoco, L.; de Pilla Varotti, F.; Carvalho, M.D.G.; de Paula Sabino, A. Antioxidant, antibacterial and antitumor activity of ethanolic extract of the Psidium guajava leaves. Am. J. Plant Sci. 2014, 5, 3492–3500. [Google Scholar] [CrossRef]
- Kaileh, M.; Vanden Berghe, W.; Boone, E.; Essawi, T.; Haegeman, G. Screening of indigenous Palestinian medicinal plants for potential anti-inflammatory and cytotoxic activity. J. Ethnopharmacol. 2007, 113, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.C.; Peng, C.H.; Chen, K.C.; Hsieh, C.L.; Peng, R.Y. The aqueous soluble polyphenolic fraction of Psidium guajava leaves exhibits potent anti-angiogenesis and anti-migration actions on DU145 cells. Evid. Based Complement. Altern. Med. 2011, 2011, 219069. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.C.; Hsieh, C.L.; Huang, K.D.; Ker, Y.B.; Chyau, C.C.; Peng, R.Y. Anticancer activity of rhamnoallosan against DU-145 cells is kinetically complementary to coexisting polyphenolics in Psidium guajava budding leaves. J. Agric. Food Chem. 2009, 57, 6114–6122. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L.Y.; Longato, G.B.; Ruiz, A.L.T.G.; Tinti, S.V.; Possenti, A.; Vendramini-costa, D.B. In vitro, in vivo and in silico analysis of the anticancer and estrogen-like activity of guava leaf extracts. Curr. Med. Chem. 2014, 21, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Park, K.R.; Nam, D.; Yun, H.M.; Lee, S.G.; Jang, H.J.; Sethi, G.; Cho, S.K.; Ahn, K.S. β-Caryophyllene oxide inhibits growth and induces apoptosis through the suppression of PI3K/AKT/mTOR/S6K1 pathways and ROS-mediated MAPKs activation. Cancer Lett. 2011, 312, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Manosroi, J.; Dhumtanom, P.; Manosroi, A. Anti-proliferative activity of essential oil extracted from Thai medicinal plants on KB and P388 cell lines. Cancer Lett. 2006, 235, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.S.; Carley, S. Cytotoxic activity of hexane extracts of Psidium guajava L. (myrtaceae) and Cassia alata L. (caesalpineaceae) in kasumi-1 and OV2008 cancer cell lines. Trop. J. Pharm. Res. 2012, 11, 201–207. [Google Scholar] [CrossRef]
- Ryu, N.H.; Park, K.R.; Kim, S.M.; Yun, H.M.; Nam, D.; Lee, S.G.; Jang, H.J.; Ahn, K.S.; Kim, S.-H.; Shim, B.S.; et al. A hexane fraction of guava leaves (Psidium guajava L.) induces anticancer activity by suppressing AKT/mammalian target of rapamycin/ribosomal p70 S6 kinase in human prostate cancer cells. J. Med. Food 2011, 15, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Hwang, J.H.; Park, S.Y.; Jin, Y.J.; Ko, H.C.; Moon, S.W.; Kim, S.J. Fermented guava leaf extract inhibits LPS-induced COX-2 and iNOS expression in Mouse macrophage cells by inhibition of transcription factor NF-kappaB. Phyther. Res. 2008, 22, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Jeong, S.W.; Cho, S.K.; Ahn, K.S.; Kim, B.K.; Kim, J.C. Anti-inflammatory effects of 4 medicinal plant extracts in lipopolysaccharide-induced RAW 264.7 cells. Food Sci. Biotechnol. 2013, 22, 213–220. [Google Scholar] [CrossRef]
- Jang, M.; Jeong, S.-W.; Cho, S.K.; Ahn, K.S.; Lee, J.H.; Yang, D.C.; Kim, J.-C. Anti-Inflammatory Effects of an Ethanolic Extract of Guava (Psidium guajava L.) Leaves In Vitro and In Vivo. J. Med. Food 2014, 17, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.S.; Sukumaran, V.; Giri, S.S.; Park, S.C. Flavonoid fraction of guava leaf extract attenuates lipopolysaccharide-induced inflammatory response via blocking of NF-κB signalling pathway in Labeo rohita macrophages. Fish Shellfish Immunol. 2015, 47, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Jeong, S.-W.; Cho, S.K.; Yang, H.J.; Yoon, D.-S.; Kim, J.-C.; Park, K.-H. Improvement in the anti-inflammatory activity of guava (Psidium guajava L.) leaf extracts through optimization of extraction conditions. J. Funct. Foods 2014, 10, 161–168. [Google Scholar] [CrossRef]
- Madduluri, S.; Sitaram, B. In vitro evaluation of anti inflammatory activity of methanolic and ethanolic leaf extracts of five indigenous plants in South India. Int. J. PharmTech Res. 2014, 6, 569–574. [Google Scholar]
- Laily, N.; Kusumaningtyas, R.W.; Sukarti, I.; Rini, M.R.D.K. The potency of guava Psidium guajava (L.) leaves as a Functional immunostimulatory ingredient. Procedia Chem. 2015, 14, 301–307. [Google Scholar] [CrossRef]
- Soman, S.; Rauf, A.A.; Indira, M.; Rajamanickam, C. Antioxidant and antiglycative potential of ethyl acetate fraction of Psidium guajava leaf extract in streptozotocin-induced diabetic rats. Plant Foods Hum. Nutr. 2010, 65, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-W.; Hsieh, C.-L.; Wang, H.-Y.; Chen, H.-Y. Inhibitory effects of guava (Psidium guajava L.) leaf extracts and its active compounds on the glycation process of protein. Food Chem. 2009, 113, 78–84. [Google Scholar] [CrossRef]
- Wang, H.; Du, Y.-J.; Song, H.-C. α-Glucosidase and α-amylase inhibitory activities of guava leaves. Food Chem. 2010, 123, 6–13. [Google Scholar] [CrossRef]
- Deguchi, Y.; Miyazaki, K. Anti-hyperglycemic and anti-hyperlipidemic effects of guava leaf extract. Nutr. Metab. Lond. 2010, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Kong, F.; Ni, H.; Mo, Z.; Wan, J.B.; Hua, D.; Yan, C. Structural characterization, α-glucosidase inhibitory and DPPH scavenging activities of polysaccharides from guava. Carbohydr. Polym. 2016, 144, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Eidenberger, T.; Selg, M.; Krennhuber, K. Inhibition of dipeptidyl peptidase activity by flavonol glycosides of guava (Psidium guajava L.): A key to the beneficial effects of guava in type II diabetes mellitus. Fitoterapia 2013, 89, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lim, Y.; Kwon, O. Selected phytochemicals and culinary plant extracts inhibit fructose uptake in caco-2 cells. Molecules 2015, 20, 17393–17404. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.-C.; Shen, S.-C.; Wu, J.S.-B. Effect of guava (Psidium guajava L.) leaf extract on glucose uptake in rat hepatocytes. J. Food Sci. 2009, 74, H132–H138. [Google Scholar] [CrossRef] [PubMed]
- Basha, S.K.; Kumari, V.S. In vitro antidiabetic activity of Psidium guajava leaves extracts. Asian Pac. J. Trop. Dis. 2012, 2 (Suppl. S1), 98–100. [Google Scholar] [CrossRef]
- Basha, S.K.; Govindaraju, K.; Manikandan, R.; Ahn, J.S.; Bae, E.Y.; Singaravelu, G. Phytochemical mediated gold nanoparticles and their PTP 1B inhibitory activity. Colloids Surf. B Biointerfaces 2010, 75, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Yoshitomi, H.; Qin, L.; Liu, T.; Gao, M. Guava leaf extracts inhibit 3T3-L1 adipocyte differentiation via activating AMPK. J. Nutr. Ther. 2012, 1, 107–113. [Google Scholar]
- Liu, C.-W.; Wang, Y.-C.; Hsieh, C.-C.; Lu, H.-C.; Chiang, W.-D. Guava (Psidium guajava Linn.) leaf extract promotes glucose uptake and glycogen accumulation by modulating the insulin signaling pathway in high-glucose-induced insulin-resistant mouse FL83B cells. Process Biochem. 2015, 50, 1128–1135. [Google Scholar] [CrossRef]
- Chang, W.-C.; Shen, S.-C. Effect of water extracts from edible myrtaceae plants on uptake of 2-(n-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino)-2-deoxyglucose in TNF-α-treated FL83B mouse hepatocytes. Phytother. Res. 2013, 27, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Arasakumari, M.; Prabu, P.; Amalraj, A.J. Anti-diabetic and aldose reductase inhibitory potential of Psidium guajava by in vitro analysis. Int. J. Pharm. Pharm. Sci. 2016, 8, 271–276. [Google Scholar] [CrossRef]
- Owen, P.L.; Matainaho, T.; Sirois, M.; Johns, T. Endothelial cytoprotection from oxidized LDL by some crude melanesian plant extracts is not related to their antioxidant capacity. J. Biochem. Mol. Toxicol. 2007, 21, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.L.; Huang, C.N.; Lin, Y.C.; Peng, R.Y. Molecular action mechanism against apoptosis by aqueous extract from guava budding leaves elucidated with human umbilical vein endothelial cell (HUVEC) model. J. Agric. Food Chem. 2007, 55, 8523–8533. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-L.; Lin, Y.-C.; Yen, G.-C.; Chen, H.-Y. Preventive effects of guava (Psidium guajava L.) leaves and its active compounds against α-dicarbonyl compounds-induced blood coagulation. Food Chem. 2007, 103, 528–535. [Google Scholar] [CrossRef]
- Anyachukwu Irondi, E.; Olalekan Agboola, S.; Oboh, G.; Augusti Boligon, A.; Linde Athayde, M.; Shode, F.O. Guava leaves polyphenolics-rich extract inhibits vital enzymes implicated in gout and hypertension in vitro. J. Intercult. Ethnopharmacol. 2016, 5, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Prabu, G.R.; Gnanamani, A.; Sadulla, S. Guaijaverin—A plant flavonoid as potential antiplaque agent against Streptococcus mutans. J. Appl. Microbiol. 2006, 101, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Shafiei, Z.; Haji Abdul Rahim, Z.; Philip, K.; Thurairajah, N. Antibacterial and anti-adherence effects of a plant extract mixture (PEM) and its individual constituent extracts (Psidium sp., Mangifera sp., and Mentha sp.) on single- and dual-species biofilms. PeerJ 2016, 4, e2519. [Google Scholar] [CrossRef] [PubMed]
- Razak, F.A.; Othman, R.Y.; Rahim, Z.H.A. The effect of Piper betle and Psidium guajava extracts on the cell-surface hydrophobicity of selected early settlers of dental plaque. J. Oral Sci. 2006, 48, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Fathilah, A.R.; Rahim, Z.H.A.; Othman, Y.; Yusoff, M. Bacteriostatic effect of Piper betle and Psidium guajava extracts on dental plaque bacteria. Pak. J. Biol. Sci. 2009, 12, 518–521. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chandrashekar, B.R.; Nagarajappa, R.; Singh, R.; Thakur, R. An in vitro study on the anti-microbial effi cacy of ten herbal extracts on primary plaque colonizers. J. Young Pharm. 2014, 6, 33–39. [Google Scholar] [CrossRef]
- John, N.R.; Gala, V.C.; Sawant, C.S. Inhibitory effects of plant extracts on multi-species dental biofilm formation in vitro. Int. J. Pharm. Bio Sci. 2013, 4, 487–495. [Google Scholar]
- John, N.R.; Gala, V.C.; Bhagwat, A.M.; Sawant, C.S. HPTLC analysis of eclipta prostrata and Psidium guajava extracts and their effect on cell-surface hydrophobicity of a consortium of dental plaque isolates. Int. J. Pharm. Pharm. Sci. 2013, 5, 1–6. [Google Scholar]
- Shekar, C.; Nagarajappa, R.; Singh, R.; Thakur, R. Antimicrobial efficacy of Acacia nilotica, Murraya koenigii L. Sprengel, Eucalyptus hybrid, and Psidium guajava on primary plaque colonizers: An in vitro comparison between hot and cold extraction process. J. Indian Soc. Periodontol. 2015, 19, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Kwamin, F.; Gref, R.; Haubek, D.; Johansson, A. Interactions of extracts from selected chewing stick sources with Aggregatibacter actinomycetemcomitans. BMC Res. Notes 2012, 5, 203. [Google Scholar] [CrossRef] [PubMed]
- Thalikunnil, S.T.; Ashok, A.; Sukesh, K. Screening of psidium gaujava for effective phytomedicines and study on its antibacterial effect against dental caries bacteria. Int. J. Pharm. Pharm. Sci. 2012, 4, 400–401. [Google Scholar]
- Jayakumari, S.; Anbu, J.; Ravichandiran, V.; Nithya, S.; Anjana, A.; Sudharani, D. Evaluation of toothache activity of methanolic extract and its various fractions from the leaves Psidium guajava Linn. Int. J. Pharma. Bio Sci. 2012, 3, 238–249. [Google Scholar]
- Ravi, K.; Divyashree, P. Psidium guajava: A review on its potential as an adjunct in treating periodontal disease. Pharmacogn. Rev. 2014, 8, 96–100. [Google Scholar] [PubMed]
- Chen, H.-H. Hepatoprotective effect of guava (Psidium guajava L.) Leaf extracts on ethanol-induced injury on clone 9 rat liver cells. Food Nutr. Sci. 2011, 2, 983–988. [Google Scholar] [CrossRef]
- Qa’dan, F.; Thewaini, A.-J.; Ali, D.A.; Afifi, R.; Elkhawad, A.; Matalka, K.Z. The antimicrobial activities of Psidium guajava and Juglans regia leaf extracts to acne-developing organisms. Am. J. Chin. Med. 2005, 33, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, E.M. The use of Psidium guajava Linn. in treating wound, skin and soft tissue infections. Sci. Res. Essay 2009, 4, 605–611. [Google Scholar]
- Padrón-Márquez, B.; Viveros-Valdez, E.; Oranday-Cárdenas, A.; Carranza-Rosales, P. Antifungal activity of Psidium guajava organic extracts against dermatophytic fungi. J. Med. Plants Res. 2012, 6, 5435–5438. [Google Scholar] [CrossRef]
- Suwanmanee, S.; Kitisin, T.; Luplertlop, N. In vitro screening of 10 edible thai plants for potential antifungal properties. Evid. Based Complement. Altern. Med. 2014, 2014, 138587. [Google Scholar] [CrossRef] [PubMed]
- You, D.-H.; Park, J.-W.; Yuk, H.-G.; Lee, S.-C. Antioxidant and tyrosinase inhibitory activities of different parts of guava (Psidium guajava L.). Food Sci. Biotechnol. 2011, 20, 1095–1100. [Google Scholar] [CrossRef]
- Han, E.H.; Hwang, Y.P.; Choi, J.H.; Yang, J.H.; Seo, J.K.; Chung, Y.C.; Jeong, H.G. Psidium guajava extract inhibits thymus and activation-regulated chemokine (TARC/CCL17) production in human keratinocytes by inducing heme oxygenase-1 and blocking NF-κB and STAT1 activation. Environ. Toxicol. Pharmacol. 2011, 32, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-U.; Yeon Weon, K.; Nam, D.-Y.; Hyun Nam, J.; Kyung Kim, W. Skin protective effect of guava leaves against UV-induced melanogenesis via inhibition of ORAI1 channel and tyrosinase activity. Exp. Dermatol. 2016, 25, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Abreu, P.R.C.; Almeida, M.C.; Bernardo, R.M.; Bernardo, L.C.; Brito, L.C.; Garcia, E.A.C.; Fonseca, A.S.; Bernardo-Filho, M. Guava extract (Psidium guajava) alters the labelling of blood constituents with technetium-99m. J. Zhejiang Univ. Sci. B 2006, 7, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Han, E.H.; Hwang, Y.P.; Kim, H.G.; Park, J.H.; Choi, J.H.; Im, J.H.; Khanal, T.; Park, B.H.; Yang, J.H.; Choi, J.M.; et al. Ethyl acetate extract of Psidium guajava inhibits IgE-mediated allergic responses by blocking FcεRI signaling. Food Chem. Toxicol. 2011, 49, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Pachanawan, A.; Phumkhachorn, P.; Rattanachaikunsopon, P. Potential of Psidium guajava supplemented fish diets in controlling Aeromonas hydrophila infection in tilapia (Oreochromis niloticus). J. Biosci. Bioeng. 2008, 106, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.L.; Li, Z.J.; Yang, K.; Lin, H.Z.; Guo, Z.X. Effect of guava leaves on growth and the non-specific immune response of Penaeus monodon. Fish Shellfish Immunol. 2014, 40, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, S.O.; Akanji, M.A.; Oguntoye, S.A. Ethanolic leaf extract of Psidium guajava: Phyto-chemical and trypanocidal activity in rats infected with Trypanosoma brucei brucei. J. Med. Plants Res. 2009, 3, 420–423. [Google Scholar]
- Akanji, M.A.; Adeyemi, O.S.; Oguntoye, S.O.; Sulyman, F. Psidium guajava extract reduces trypanosomosis associated lipid peroxidation and raises glutathione concentrations in infected animals. EXCLI J. 2009, 8, 148–154. [Google Scholar]
- Rajendran, C.; Begam, M.; Kumar, D.; Baruah, I.; Gogoi, H.K.; Srivastava, R.B.; Veer, V. Antiplasmodial activity of certain medicinal plants against chloroquine resistant Plasmodium berghei infected white albino BALB/c mice. J. Parasit. Dis. 2014, 38, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Birdi, T. Psidium guajava leaf extract prevents intestinal colonization of Citrobacter rodentium in the mouse model. J. Ayurveda Integr. Med. 2015, 6, 50. [Google Scholar] [PubMed]
- Geidam, Y.A.; Ambali, A.G.; Onyeyili, P.A.; Tijjani, M.B.; Gambo, H.I.; Gulani, I.A. Antibacterial efficacy of ethyl acetate fraction of Psidium guajava leaf aqueous extract on experimental Escherichia coli (O78) infection in chickens. Vet. World 2015, 8, 358–362. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shittu, O.B.; Ajayi, O.L.; Bankole, S.O.; Popoola, T.O.S. Intestinal ameliorative effects of traditional Ogi-tutu, Vernonia amygdalina and Psidium guajava in mice infected with Vibrio cholera. Afr. Health Sci. 2016, 16, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Jan, O.Q.; Kamili, N.; Ashraf, A.; Iqbal, A.; Sharma, R.K.; Rastogi, A. Haematobiochemical parameters of goats fed tannin rich Psidium guajava and Carissa spinarum against Haemonchus contortus infection in India. J. Parasit. Dis. 2013, 39, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Seo, N.; Ito, T.; Wang, N.; Yao, X.; Tokura, Y.; Furukawa, F.; Takigawa, M.; Kitanaka, S. Anti-allergic Psidium guajava extracts exert an antitumor effect by inhibition of T regulatory cells and resultant augmentation of Th1 cells. Anticancer Res. 2005, 25, 3763–3770. [Google Scholar] [PubMed]
- Adeyemi, O.S.; Akanji, M.A.; Ekanem, J.T. Anti-anaemic properties of the ethanolic extracts of Psidium guajava in Trypanosoma brucei brucei Infected Rats. Res. J. Pharmacol. 2010, 4, 74–77. [Google Scholar] [CrossRef]
- Udem, S.C.; Anyanwu, M.U.; Obidike, R.I.; Udem, N.D. The effects of Psidium guajava Linn. (Myrtaceae) leaf chloroform extract on some hematological and biochemical parameters in mice. Comp. Clin. Pathol. 2011, 20, 47–51. [Google Scholar] [CrossRef]
- Raj, V.B.; Rao, R.M.; Kumar, R.K.; Srinivas, K. Analgesic effect of ethanol extracted leaves of Psidium guajava leaves in animal models. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 1796–1801. [Google Scholar]
- Stalin D., J. A study on the analgesic property of methanolic and aqueous extracts of dried leaf of Psidium guajava Linn. Int. J. Adv. Res. Pharm. Bio Sci. 2013, 3, 1–6. [Google Scholar]
- Akinloye, O.; Akinmoladun, A.C.; Farombi, E.O. Modulatory effect of Psidium guajava linn and ocimum gratissimum Linn on lipid profile and selected biochemical indices in rabbits fed high cholesterol diet. J. Complement. Integr. Med. 2010, 7. [Google Scholar] [CrossRef]
- Mesquita Freire, J.; Patto de Abreu, C.M.; da Silveira Duarte, S.M.; Borges Araújo de Paula, F.; Ribeiro Lima, A. Evaluation of the protective effect of guava fruits and leaves on oxidative stress. Acta Sci. Biol. Sci. 2014, 36, 35–40. [Google Scholar]
- Oh, W.K.; Lee, C.H.; Lee, M.S.; Bae, E.Y.; Sohn, C.B.; Oh, H.; Kim, B.Y.; Ahn, J.S. Antidiabetic effects of extracts from. Psidium Guajava 2005, 96, 411–415. [Google Scholar]
- Mansoori Bahrani, A.H.; Zaheri, H.; Soltani, N.; Kharazmi, F. Effect of the administration of Psidium guava leaves on blood glucose, lipid profiles and sensitivity of the vascular mesenteric bed to Phenylephrine in streptozotocin-induced diabetic rats. J. Diabetes Mellit. 2012, 2, 138–145. [Google Scholar] [CrossRef]
- Shen, S.-C.; Cheng, F.-C.; Wu, N.-J. Effect of guava (Psidium guajava Linn.) leaf soluble solids on glucose metabolism in type 2 diabetic rats. Phytother. Res. 2008, 22, 1458–1464. [Google Scholar] [CrossRef] [PubMed]
- Ogueri, C.C.; Elekwa, I.; Ude, V.C.; Ugbogu, A.E. Effect of aqueous extract of guava (Psidium guajava) leaf on blood glucose and liver enzymes in alloxan induced diabetic rats. Br. J. Pharm. Res. 2014, 4, 1079–1087. [Google Scholar] [CrossRef]
- Shakeera Banu, M.; Sujatha, K.; Sridharan, G.; Manikandan, R. Antihyperglycemic and antihyperlipidemic potentials of Psidium guajava in alloxan-induced diabetic rats. Asian J. Pharm. Clin. Res. 2013, 6, 88–89. [Google Scholar]
- Adeyemi, O.S.; Akanji, M.A. Biochemical changes in the kidney and liver of rats following administration of ethanolic extract of Psidium guajava leaves. Hum. Exp. Toxicol. 2011, 30, 1266–1274. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.S.; Akanji, M.A. Psidium guajava leaf extract: Effects on rat serum homeostasis and tissue morphology. Comp. Clin. Pathol. 2012, 21, 401–407. [Google Scholar] [CrossRef]
- Ademiluyi, A.O.; Oboh, G.; Ogunsuyi, O.B.; Oloruntoba, F.M. A comparative study on antihypertensive and antioxidant properties of phenolic extracts from fruit and leaf of some guava (Psidium guajava L.) varieties. Comp. Clin. Pathol. 2015, 25, 363–374. [Google Scholar] [CrossRef]
- Chiwororo, W.D.H.; Ojewole, J.A.O. Biphasic effect of Psidium guajava Linn. (Myrtaceae) leaf aqueous extract on rat isolated vascular smooth muscles. J. Smooth Muscle Res. 2008, 44, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Olatunji-Bello, I.I.; Odusanya, A.J.; Raji, I.; Ladipo, C.O. Contractile effect of the aqueous extract of Psidium guajava leaves on aortic rings in rat. Fitoterapia 2007, 78, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Hosokawa, T.; Morinaka, T.; Irino, S.; Hirano, S.; Kobayashi, H.; Yoshioka, A.; Suzuki-Yamamoto, T.; Yokoro, M.; Kimoto, M.; et al. Antiatherogenic effect of guava leaf extracts inhibiting leucocyte-type 12-lipoxygenase activity. Food Chem. 2012, 131, 1069–1075. [Google Scholar] [CrossRef]
- Takahashi, Y.; Otsuki, A.; Mori, Y.; Kawakami, Y.; Ito, H. Inhibition of leukocyte-type 12-lipoxygenase by guava tea leaves prevents development of atherosclerosis. Food Chem. 2015, 186, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Soman, S.; Rajamanickam, C.; Rauf, A.A.; Indira, M. Beneficial effects of Psidium guajava leaf extract on diabetic myocardium. Exp. Toxicol. Pathol. 2013, 65, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Livingston Raja, N.R.; Sundar, K. Psidium guajava Linn confers gastro protective effects on rats. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 151–156. [Google Scholar] [PubMed]
- Tende, J.A.; Eze, E.D.; Tende, Y.A.; Onaadepo, O.; Shaibu, A. Anti-ulcerogenic activity of guava (Psidium guajava) leaves extract in rats. Ann. Exp. Biol. 2013, 1, 6–9. [Google Scholar]
- Umana Uduak, E.; Timbuak, J.A.; Musa, S.A.; Ikyembe, D.T.; Abdurrashid, S.; Hamman, W.O. Ulceroprotective effect of methanol extract of Psidium guajava leaves on ethanol induced gastric ulcer in adult wistar rats. Asian J. Med. Sci. 2012, 4, 75–78. [Google Scholar]
- Jayakumari, S.; Anbu, J.; Ravichandiran, V.; Anjana, A.; Siva Kumar, G.M.; Maharaj, S. Antiulcerogenic and free radical scavenging activity of flavonoid fraction of Psidium guajava Linn leaves. Int. J. Pharm. Pharm. Sci. 2012, 4, 170–174. [Google Scholar]
- Ojewole, J.A.O.; Awe, E.O.; Chiwororo, W.D.H. Antidiarrhoeal activity of Psidium guajava Linn. (Myrtaceae) leaf aqueous extract in rodents. J. Smooth Muscle Res. 2008, 44, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Ezekwesili, J.O.; Nkemdilim, U.U.; Okeke, C.U. Mechanism of antidiarrhoeal effect of ethanolic extract of Psidium guajava leaves. Biokemistri 2010, 22, 85–90. [Google Scholar]
- Shah, A.J.; Begum, S.; Hassan, S.I.; Ali, S.N.; Siddiqui, B.S.; Gilani, A.-H. Pharmacological basis for the medicinal use of Psidium guajava leave in hyperactive gut disorders. Bangladesh J. Pharmacol. 2011, 6, 100–105. [Google Scholar] [CrossRef][Green Version]
- Roy, C.K.; Kamath, J.V.; Asad, M. Hepatoprotective activity of Psidium guajava Linn. leaf extract. Indian J. Exp. Biol. 2006, 44, 305–311. [Google Scholar] [PubMed]
- D’Mello, P.; Rana, M. Hepatoprotective activity of Psidium guajava extract and its phospholipid complex in paracetamol induced hepatic damage in rats. Int. J. Phytomed. 2010, 2, 85–93. [Google Scholar]
- Taju, G.; Jayanthi, M.; Majeed, S.A. Evaluation of hepatoprotective and antioxidant activity of Psidium guajava leaf extract against acetaminophen induced liver injury in rats. Int. J. Toxicol. Appl. Pharmacol. 2011, 1, 13–20. [Google Scholar]
- Osaman, M.; Ahamad, M.; Mahfouz, S.; Elaby, S. Biochemical studies on the hepatoprotective effects of pomegranate and guava ethanol extracts. N. Y. Sci. J. 2011, 4, 1–15. [Google Scholar]
- Mohamed, E.A.K. Hepatoprotective effect of aqueous leaves extract of Psidium guajava and Zizyphus spina-christi against paracetamol induced hepatotoxicity in rats. J. Appl. Sci. Res. 2012, 8, 2800–2806. [Google Scholar]
- Choi, J.H.; Park, B.H.; Kim, H.G.; Hwang, Y.P.; Han, E.H.; Jin, S.W.; Seo, J.K.; Chung, Y.C.; Jeong, H.G. Inhibitory effect of Psidium guajava water extract in the development of 2,4-dinitrochlorobenzene-induced atopic dermatitis in NC/Nga mice. Food Chem. Toxicol. 2012, 50, 2923–2929. [Google Scholar] [CrossRef] [PubMed]
- Porta Santos Fernandes, K.; Kalil Bussadori, S.; Martins Marques, M.; Sumie Wadt Yamashita, N.; Bach, E.; Domingues Martins, M. Healing and cytotoxic effects of Psidium guajava (Myrtaceae) leaf extracts. Braz. J. Oral Sci. 2010, 9, 9–14. [Google Scholar]
- Baroroh, H.N.; Utami, E.D. Harwoko inhibitory effect of ethanolic extract of Psidium guajava leaves in rat active cutaneus anaphylaxis reaction. Int. J. Pharm. Clin. Res. 2016, 8, 1–5. [Google Scholar]
- Wang, X.; Ye, K.; Lv, Y.; Wei, S.; Li, X.; Ma, J.; Zhang, X.; Ye, C. Ameliorative effect and underlying mechanisms of total triterpenoids from Psidium guajava Linn (myrtaceae) leaf on high-fat streptozotocin-induced diabetic peripheral neuropathy in rats. Trop. J. Pharm. Res. 2016, 15, 327–333. [Google Scholar] [CrossRef]
- Kumar, A.; Kumarchandra, R.; Rai, R.; Rao, B. Radiomodulatory role of Psidium guajava leaf extracts against X-ray induced genotoxicity, oxidative stress and apoptosis in albino wistar rat model. J. Appl. Pharm. Sci. 2016, 6, 58–65. [Google Scholar] [CrossRef]
- Ekaluo, U.B.; Ikpeme, E.V.; Uno, U.U.; Umeh, S.O.; Erem, F.A. Protective role of aqueous guava leaf extract against caffeine induced spermatotoxicity in albino rats. Res. J. Med. Plant 2016, 10, 98–105. [Google Scholar] [CrossRef][Green Version]
- Kaneko, K.; Suzuki, K.; Iwadate-Iwata, E.; Kato, I.; Uchida, K.; Onoue, M. Evaluation of food-drug interaction of guava leaf tea. Phyther. Res. 2013, 27, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Vladislavovna Doubova, S.; Reyes Morales, H.; Flores Hernández, S.; Martínez-García, M.D.C.; González de Cossío Ortiz, M.; Chávez Soto, M.A.; Rivera Arce, E.; Lozoya, X. Effect of a Psidii guajavae folium extract in the treatment of primary dysmenorrhea: A randomized clinical trial. J. Ethnopharmacol. 2007, 110, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, D.; Nayana, O.V.; Thomas, L. Antimicrobial plant extract incorporated gelatin beads for potential application in pharmaceutical industry. Biotechnol. Res. 2016, 2, 11–14. [Google Scholar]
- Giri, S.S.; Sen, S.S.; Chi, C.; Kim, H.J.; Yun, S.; Park, S.C.; Sukumaran, V. Effect of guava leaves on the growth performance and cytokine gene expression of Labeo rohita and its susceptibility to Aeromonas hydrophila infection. Fish Shellfish Immunol. 2015, 46, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Fawole, F.J.; Sahu, N.P.; Pal, A.K.; Ravindran, A. Haemato-immunological response of Labeo rohita (Hamilton) fingerlings fed leaf extracts and challenged by Aeromonas hydrophila. Aquac. Res. 2016, 47, 3788–3799. [Google Scholar] [CrossRef]
- Gobi, N.; Ramya, C.; Vaseeharan, B.; Malaikozhundan, B.; Vijayakumar, S.; Kadarkarai, M.; Benelli, G. Oreochromis mossambicus diet supplementation with Psidium guajava leaf extracts enhance growth, immune, antioxidant response and resistance to Aeromonas hydrophila. Fish Shellfish Immunol. 2016, 58, 572–583. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).