
The Peculiar Glycolytic Pathway in Hyperthermophylic Archaea: Understanding Its Whims by Experimentation In Silico


Abstract
:
1. Introduction
2. Results
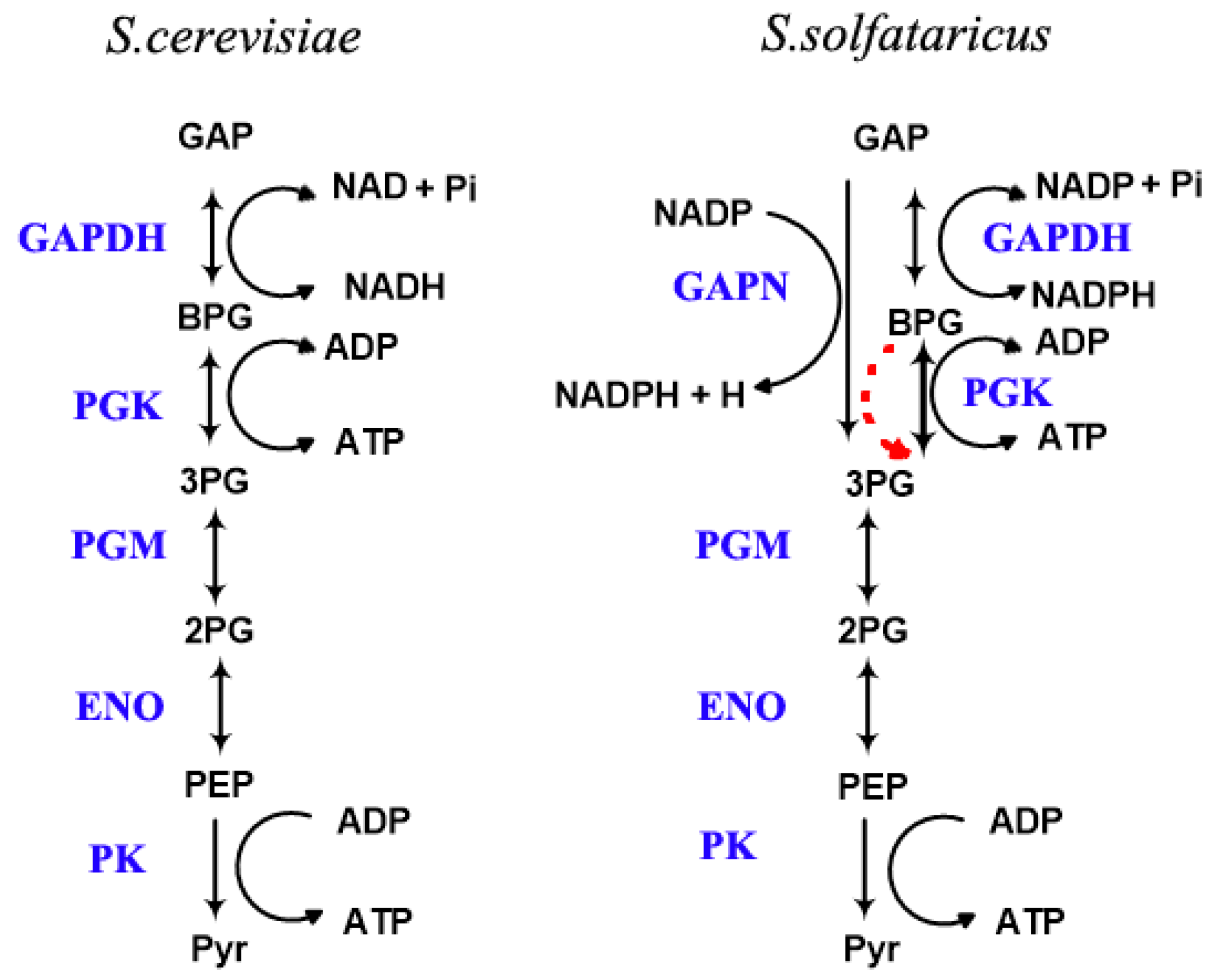
2.1. The Scheme of the Model
2.2. The Three Proposed Hypotheses
2.3. In Silico Prevalidation of the Hypotheses
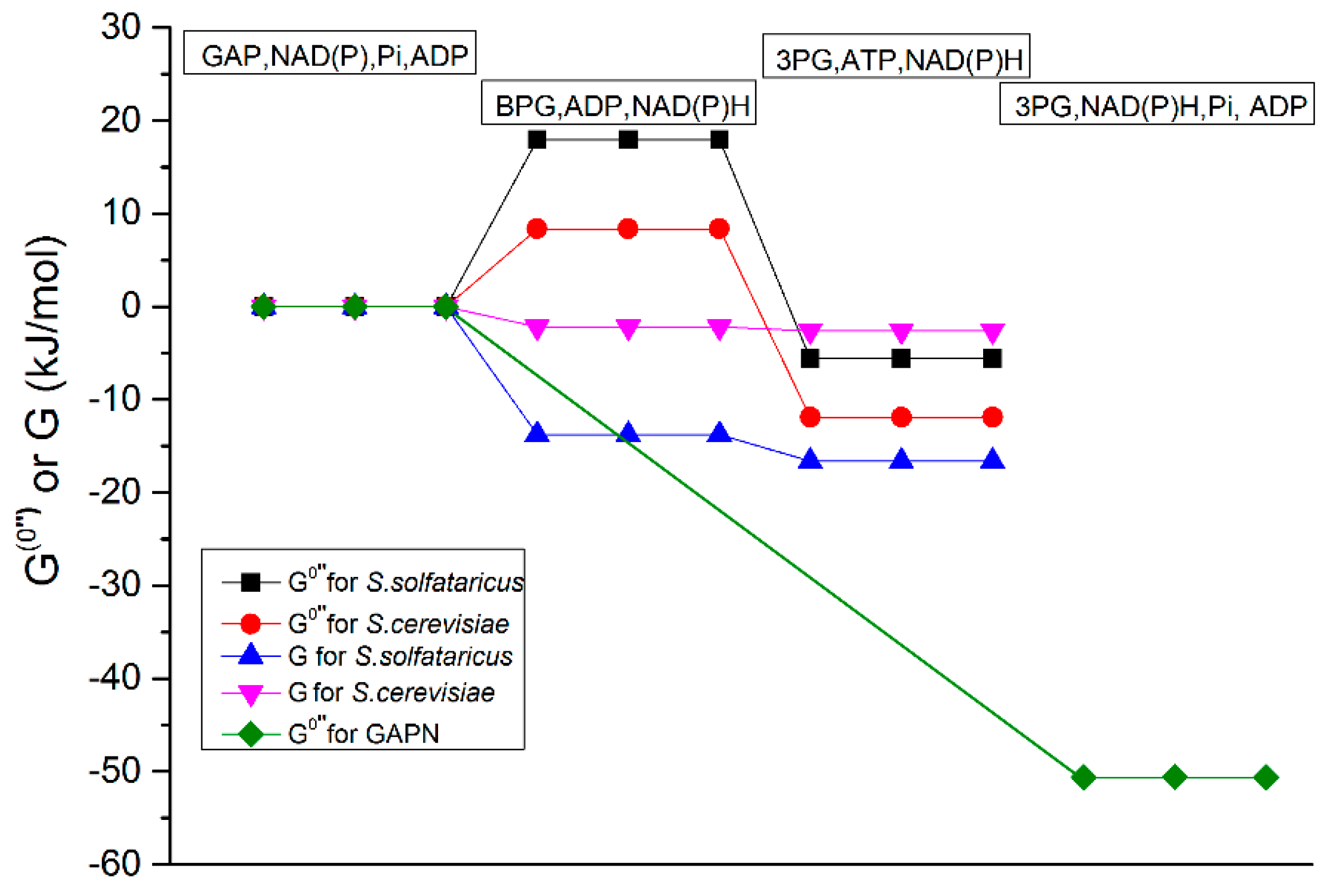
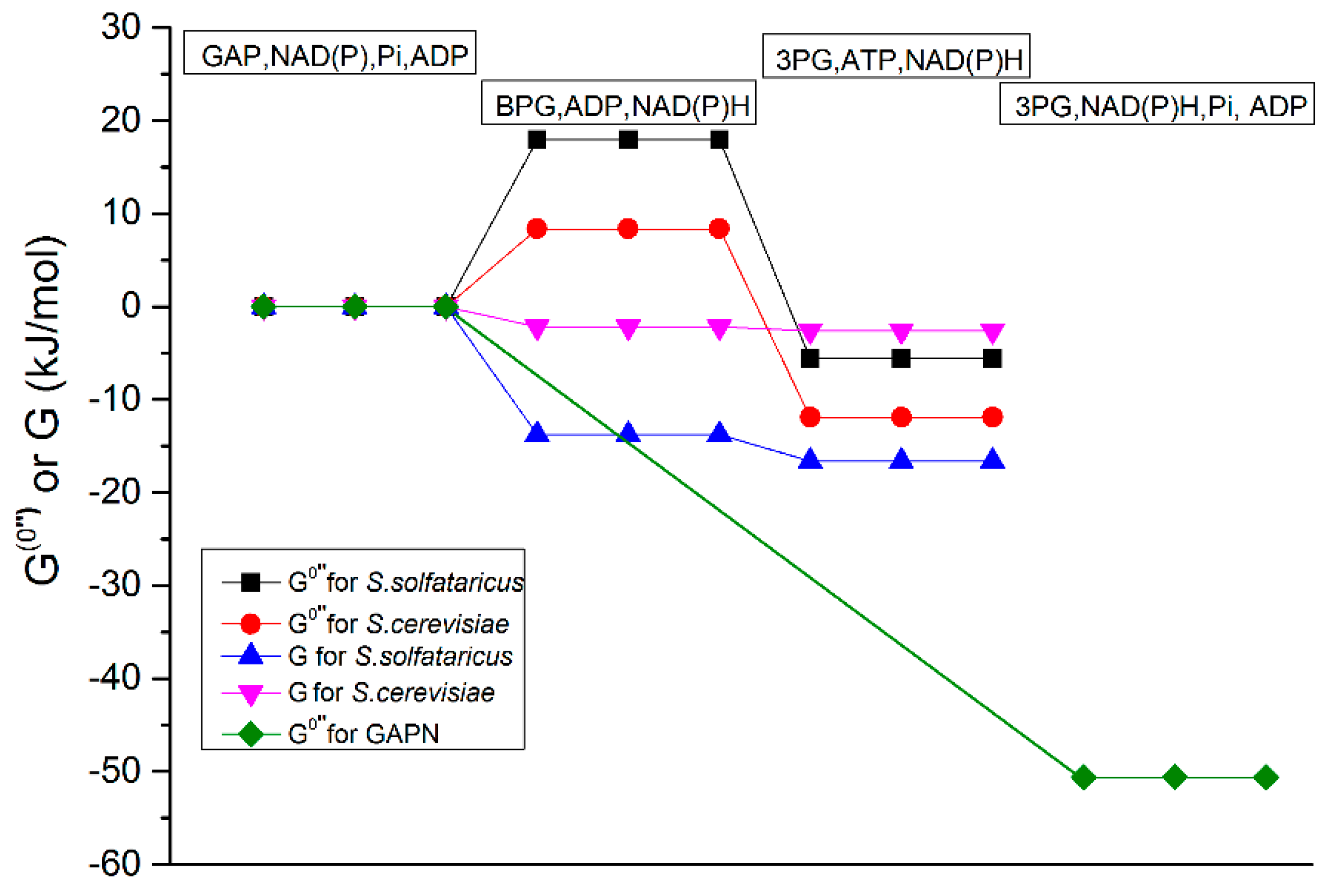
2.3.1. Test of the First Hypothesis: Is the GAPDH Reaction Thermodynamically too Unfavorable?
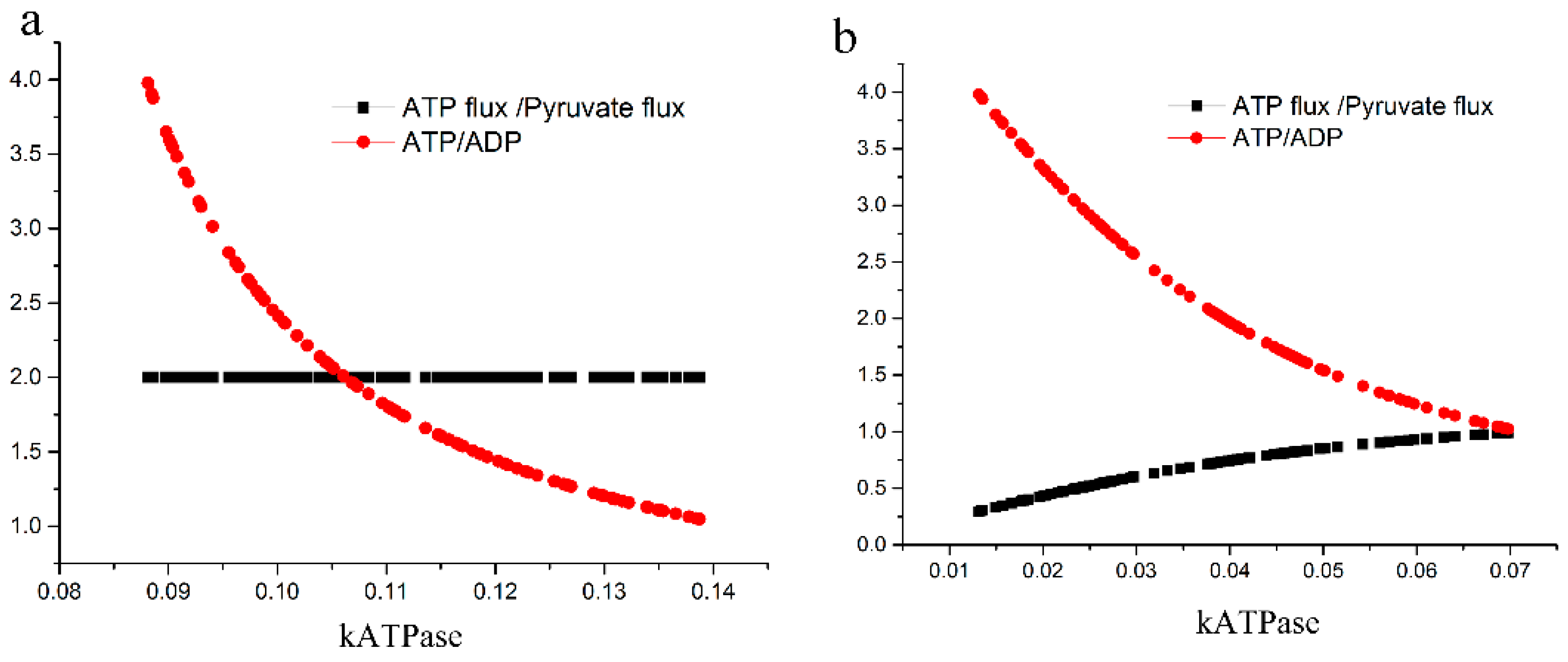
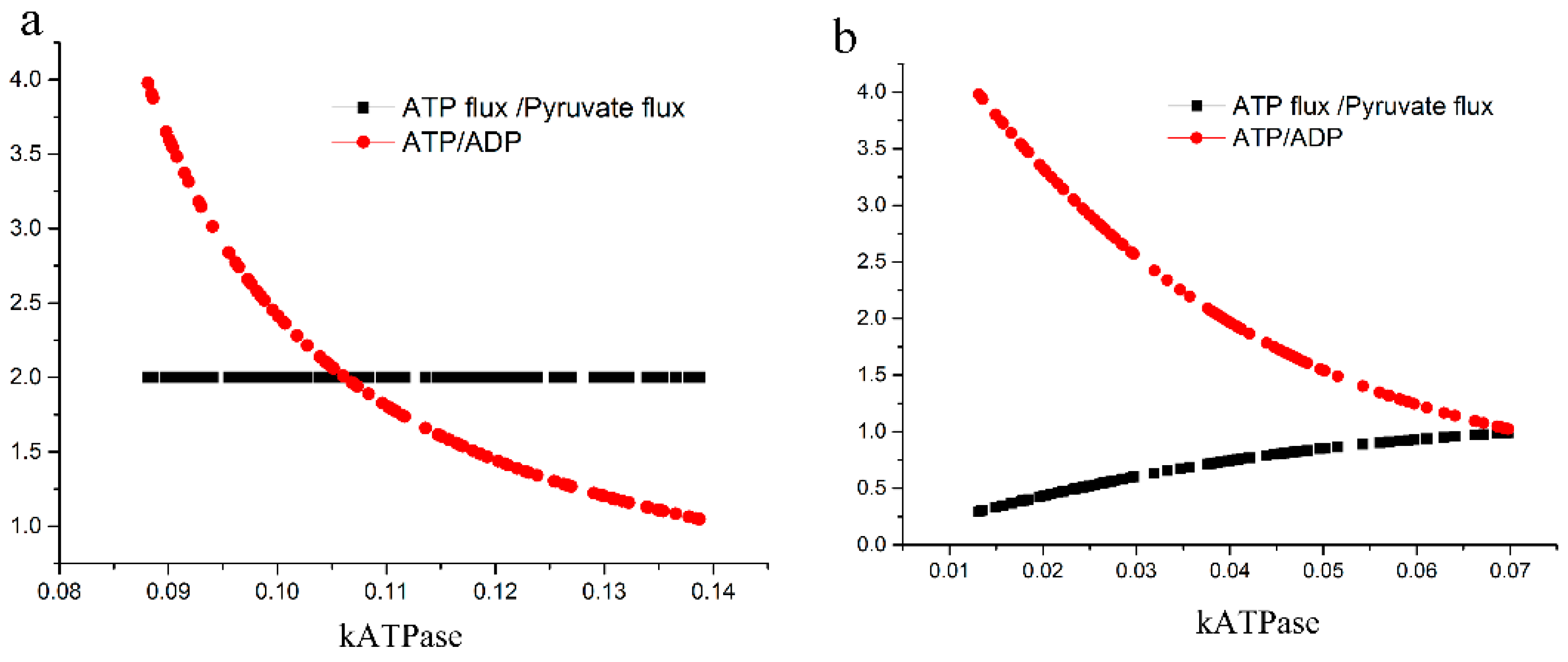
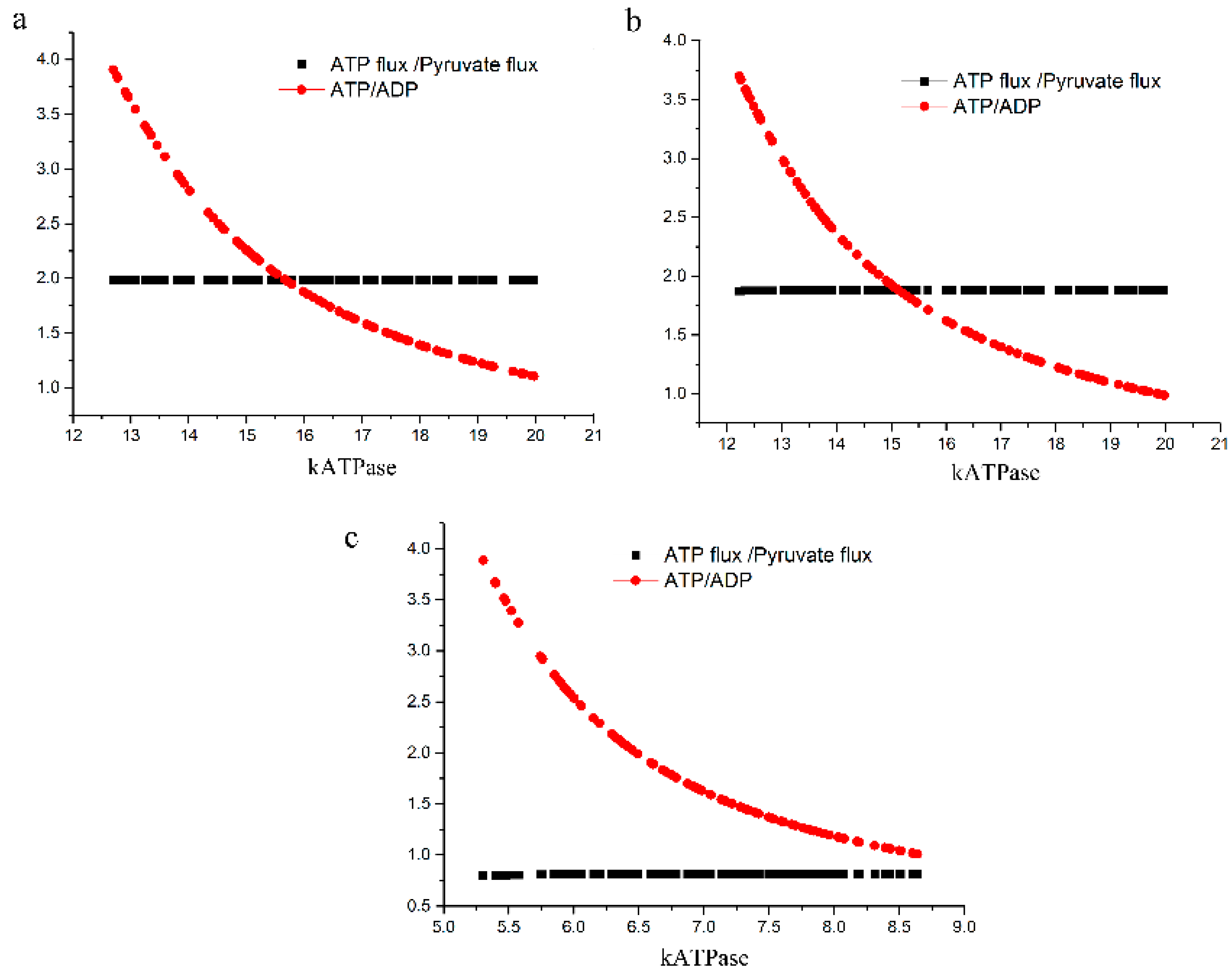
2.3.2. Test of the Second Hypothesis: Would There Be an Excessive ATP Loss?
2.3.3. Test of the Third Hypothesis: Is Traffic Bidirectional?
3. Discussion
4. Materials and Methods
4.1. Model Construction
4.2. Hypotheses Tests and Simulations
4.2.1. Test of the First Hypothesis; Thermodynamically Unfavorable GAPDH
4.2.2. Test of the Second Hypothesis; Excessive Loss of ATP
4.2.3. Test of the Third Hypothesis; Bidirectional Traffic through GAPDH Plus PGK and GAPDH
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chaban, B.; Ng, S.Y.M.; Jarrell, K.F. Archaeal habitats from the extreme to the ordinary. Can. J. Microbiol. 2006, 116, 73–116. [Google Scholar] [CrossRef] [PubMed]
- Stetter, K.O. Extremophiles and their adaptation to hot environments. FEBS Lett. 1999, 452, 22–25. [Google Scholar] [CrossRef]
- Daniel, R.M.; Cowan, D.A. Biomolecular stability and life at high temperatures. Cell Mol. Life Sci. 2000, 57, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Manaka, T. Molecular bases of thermophily in hyperthermophiles. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2011, 87, 587–602. [Google Scholar] [CrossRef]
- Ettema, T.J.; Ahmed, H.; Geerling, A.C.; van der Oost, J.; Siebers, B. The non-phosphorylating glyceraldehyde-3-phosphate dehydrogenase (GAPN) of Sulfolobus solfataricus: A key-enzyme of the semi-phosphorylative branch of the Entner–Doudoroff pathway. Extremophiles 2008, 12, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Bräsen, C.; Esser, D.; Rauch, B.; Siebers, B. Carbohydrate metabolism in Archaea: Current insights into unusual enzymes and pathways and their regulation. Microbiol. Mol. Biol. Rev. 2014, 78, 89–175. [Google Scholar] [CrossRef] [PubMed]
- Kouril, T.; Esser, D.; Kort, J.; Westerhoff, H.V.; Siebers, B.; Snoep, J.L. Intermediate instability at high temperature leads to low pathway efficiency for an in vitro reconstituted system of gluconeogenesis in Sulfolobus solfataricus. FEBS J. 2013, 280, 4666–4680. [Google Scholar] [CrossRef] [PubMed]
- Alberghina, L.; Westerhoff, H.V. (Eds.) Systems Biology: Definitions and Perspectives; Springer: New York, NY, USA, 2005. [Google Scholar]
- Kitano, H. Systems biology: A brief overview. Science 2002, 295, 1662–1665. [Google Scholar] [CrossRef] [PubMed]
- Haanstra, J.R.; van Tuijl, A.; Kessler, P.; Reijnders, W.; Michels, P.A.M.; Westerhoff, H.V.; Parsons, M.; Bakker, B.M. Compartmentation prevents a lethal turbo-explosion of glycolysis in trypanosomes. Proc. Natl. Acad. Sci. USA 2008, 105, 17718–17723. [Google Scholar] [CrossRef] [PubMed]
- Teusink, B.; Walsh, M.C.; van Dam, K.; Westerhoff, H.V. The danger of metabolic pathways with turbo design. Trends Biochem. Sci. 1998, 23, 162–169. [Google Scholar] [CrossRef]
- Smallbone, K.; Messiha, H.L.; Carroll, K.M.; Winder, C.L.; Malys, N.; Dunn, W.B.; Murabito, E.; Swainston, N.; Dada, J.O.; Khan, F.; et al. A model of yeast glycolysis based on a consistent kinetic characterisation of all its enzymes. FEBS Lett. 2013, 587, 2832–2841. [Google Scholar] [CrossRef] [PubMed]
- Bruggeman, F.J.; Boogerd, F.C.; Westerhoff, H.V. The multifarious short-term regulation of ammonium assimilation of Escherichia coli: Dissection using an in silico replica. FEBS J. 2005, 272, 1965–1985. [Google Scholar] [CrossRef] [PubMed]
- Hoops, S.; Sahle, S.; Gauges, R.; Lee, C.; Simus, N.; Singhal, M.; Kummer, U. Systems biology COPASI—A COmplex PAthway Simulator. Bioinformatics 2006, 22, 3067–3074. [Google Scholar] [CrossRef] [PubMed]
- Snoep, J.L.; Bruggeman, F.; Olivier, B.G.; Westerhoff, H.V. Towards building the silicon cell: A modular approach. BioSystems 2006, 83, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Westerhoff, H.V.; Simonetti, A.L.M.; van Dam, K. The hypothesis of localized chemiosmosis is unsatisfactory. Biochem. J. 1981, 200, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Freytag, B.; Steffen, M.; Ludwig, H.G.; Wedemeyer-Bohm, S.; Schaffenberger, W.; Steiner, O. Simulation of stellar convection with CO5BOLD. J. Comput. Phys. 2012, 231, 919–959. [Google Scholar] [CrossRef]
- Kolodkin, A.N.; Bruggeman, F.J.; Moné, M.J.; Bakker, B.M.; Campbell, M.J.; van Leeuwen, J.P.T.M.; Carlberg, C.; Westerhoff, H.V. Design principles of nuclear receptor signalling: How complex networking improves signal transduction. Mol. Syst. Biol. 2010, 6, 446. [Google Scholar] [CrossRef] [PubMed]
- Kouril, T.; Kolodkin, A.; Zaparty, M.; Steuer, R.; Ruoff, P.; Westerhoff, H.V.; Snoep, J.; Siebers, B. Sulfosys consortium. In Sulfolobus Systems Biology: Cool Hot Design for Metabolic Pathways. Systems Microbiology Current Topics and Applications; Robertson, B.D., Brendan, W., Eds.; Caister Academic Press: Norfolk, UK, 2012; Volume 8, pp. 153–170. [Google Scholar]
- Westerhoff, H.V.; van Dam, K. Thermodynamics and Control of Biological Free-Energy Transduction; Elsevier Science Ltd.: Amsterdam, The Netherlands, 1987. [Google Scholar]
- Tristan, C.; Shahani, N.; Sedlak, T.W.; Sawa, A. The diverse functions of GAPDH: Views from different subcellular compartments. Cell Signal. 2011, 23, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Teusink, B.; Passarge, J.; Reijenga, C.A.; Esgalhado, E.; van der Weijden, C.C.; Schepper, M.; Walsh, M.C.; Bakker, B.M.; van Dam, K.; Westerhoff, H.V. Can yeast glycolysis be understood in terms of in vitro kinetics of the constituent enzymes? Testing biochemistry. Eur. J. Biochem. 2000, 267, 5313–5329. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Yokooji, Y.; Atomi, H.; Imanaka, T. Biochemical and genetic characterization of the three metabolic routes in Thermococcus kodakarensis linking glyceraldehyde 3-phosphate and 3-phosphoglycerate. Mol. Microbiol. 2011, 81, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- Di Talia, S.; Skotheim, J.M.; Bean, J.M.; Siggia, E.D.; Cross, F.R. The effects of molecular noise and size control on variability in the budding yeast cell cycle. Nature 2007, 448, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Zaparty, M.; Esser, D.; Gertig, S.; Haferkamp, P.; Kouril, T.; Manica, A.; Pham, T.K.; Reimann, J.; Schreiber, K.; Sierocinski, P.; et al. Hot standards for the thermoacidophilic archaeon Sulfolobus solfataricus. Extremophiles 2010, 14, 119–142. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, N.; Dipasquale, L.; d’Ippolito, G.; Fontana, A.; Panico, A.; Lens, P.N.; Pirozzi, F.; Esposito, G. Kinetic modeling of fermentative hydrogen production by Thermotoga Neapolitana. Int. J. Hydrog. Energy 2016, 41, 4931–4940. [Google Scholar] [CrossRef]
- Van Eunen, K.; Bouwman, J.; Daran-Lapujade, P.; Postmus, J.; Canelas, A.B.; Mensonides, F.I.; Orij, R.; Tuzun, I.; van den Brink, J.; Smits, G.J.; et al. Measuring enzyme activities under standardized in vivo-like conditions for systems biology. FEBS J. 2010, 277, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Westerhoff, H.V.; Melandri, B.A.; Venturoli, G.; Azzone, G.F.; Kell, D.B. Mosaic protonic coupling hypothesis for free energy transduction. FEBS Lett. 1984, 165, 1–5. [Google Scholar] [CrossRef]
- Richard, P.; Teusink, B.; Hemker, M.B.; van Dam, K.; Westerhoff, H.V. Sustained oscillations in free-energy state and hexose phosphates in yeast. Yeast 1996, 12, 731–740. [Google Scholar] [CrossRef]
- Rohwer, J.M.; Postma, P.W.; Kholodenko, B.N.; Westerhoff, H.V. Implications of macromolecular crowding for signal transduction and metabolite channeling. Proc. Natl. Acad. Sci. USA 1998, 95, 10547–10552. [Google Scholar] [CrossRef] [PubMed]
- Hornberg, J.J.; Bruggeman, F.J.; Westerhoff, H.V.; Lankelma, J. Cancer: A systems biology disease. Biosystems 2006, 83, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Bakker, B.M.; Westerhoff, H.V.; Opperdoes, F.R.; Michels, P.A. Metabolic control analysis of glycolysis in trypanosomes as an approach to improve selectivity and effectiveness of drugs. Mol. Biochem. Parasitol. 2000, 106, 1–10. [Google Scholar] [CrossRef]
- Van Eunen, K.; Kiewiet, J.A.; Westerhoff, H.V.; Bakker, B.M. Testing biochemistry revisited: How in vivo metabolism can be understood from in vitro enzyme kinetics. PLoS Comput. Biol. 2012, 8, e1002483. [Google Scholar] [CrossRef] [PubMed]
- Kouril, T. Challenging Metabolic Networks at High Temperature: The Central Carbohydrate Metabolism of Sulfolobus solfataricus. Ph.D. Thesis, Vrije Universiteit Amsterdam, Amsterdam, The Netherlands, 2012. [Google Scholar]
- Kouril, T.; Eicher, J.J.; Siebers, B.; Snoep, J.L. Phosphoglycerate kinase acts as a futile cycle at high temperature. 2017; Unpublished. [Google Scholar]
- Byers, L.D. Glyceradehyde-3-phosphate dehydrogenase from yeast. Methods Enzymol. 1982, 89, 326–335. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | GAPDH | PGK | GAPDH | PGK |
|---|---|---|---|---|
| Temperature | 70 °C | 70 °C | 30 °C | 30 °C |
| Keq | 0.18 M−1 | 3793 | 3.6 M−1 | 3200 |
| ∆G°′′ (Pi = 1 mM) | 24.4 kJ/mol | −23.5 kJ/mol | 14.2 kJ/mol | −20.3 kJ/mol |
| ∆G°′′ (Pi = 10 mM) | 17.9 kJ/mol | −23.5 kJ/mol | 8.4 kJ/mol | −20.3 kJ/mol |
| ∆G°′′ (Pi = 100 mM) | 11.4 kJ/mol | −23.5 kJ/mol | 2.6 kJ/mol | −20.3 kJ/mol |
| ∆G°′′ (Pi = 1 M) ≡ ∆G0′ | 4.9 kJ/mol | −23.5 kJ/mol | −3.2 kJ/mol | −20.3 kJ/mol |
| Reactions between GAP and 3PG Included | S. cerevisiae (30 °C) | S. solfataricus (70 °C) | ||
|---|---|---|---|---|
| PGK, GAPDH | PGK, GAPDH | PGK and 1000 Times GAPDH | PGK, GAPDH, GAPN | |
| JGAPDH | 236 | 0.136 | 7.4 | −0.232 |
| JPGK | 236 | 0.136 | 7.4 | −0.234 |
| JGAPN | --- | --- | --- | 16.82 |
| BPG | 0.007 | 6.9 × 10−6 | 8.1 × 10−4 | 2.4 × 10−3 |
| CJGAPDH | 0.11 | 0.99 | 0.03 | −0.011 |
| CJPGK | 0.01 | 0.005 | 0.40 | 0 |
| Flux (mM/min) | GAPDH | GAPN | PGK | ENO | PGM |
|---|---|---|---|---|---|
| S. cerevisiae | 235 | --- | 235 | 235 | 235 |
| S. solfataricus | −0.23 | 16.82 | −0.23 | 16.59 | 16.59 |
| Flux (mM/min) | GAPDH | GAPN | PGK | ENO | PGM |
|---|---|---|---|---|---|
| S. cerevisiae | −107 | --- | −107 | −107 | −107 |
| S. solfataricus | −1.6 | 0.7 | −1.6 | −0.83 | −0.83 |
| Enzymes | Rate Laws | Parameters |
|---|---|---|
| GAPDH for S.cerevisiae | VmGAPDH = 1859 mM/min [33] | |
| KGAPDHGAP = 0.21 mM [12] | ||
| KeqGAPDH = 0.0054 [12] | ||
| KGAPDHNAD = 0. 09 mM [12] | ||
| KGAPDHNADH = 0.06 mM [12] | ||
| KGAPDHBPG = 0.0098 mM [12] | ||
| KGAPDHPi = 1.5 mM [12] | ||
| PGK for S.cerevisiae | 1 | VmrPGK = 2670 mM/min [33] |
| KPGKADP = 0.2 mM [12] | ||
| KPGKATP = 0.3 mM [12] | ||
| KPGKBPG = 0.003 mM [12] | ||
| KPGK3PG = 0.53 mM [12] | ||
| KeqPGK = 3200 [12] | ||
| PGMA | VmPGMA for S.cerevisiae = 856 mM/min [33] | |
| VmPGMA for S. solfataricus = 56 mM/min [34] | ||
| KmPGMA2PG = 0.08 mM [12] | ||
| KmPGMA3PG = 1.2 mM [12] | ||
| KeqPGMA = 0.19 mM [12] | ||
| ENO | VmENO for S.cerevisiae = 357 mM/min [33] | |
| VmENO for S. solfataricus = 20.5 mM/min 2 | ||
| KmENO2PG = 0.04 mM [12] | ||
| KmENOPEP = 0.5 mM [12] | ||
| KeqENO = 6.7 mM [12] | ||
| PYK | VmPYK for S.cerevisiae = 559 mM/min [34] | |
| VmPYK for S. solfataricus = 76 mM/min [34] | ||
| KmPYKADP = 0.53 mM [12] | ||
| KmPYKATP = 1.5 mM [12] | ||
| KmPYKPEP = 0.14 mM [12] | ||
| KmPYKPYR = 21 mM [12] | ||
| KeqPYK = 6500 mM [12] | ||
| GAPDH for S. solfataricus | VmGAPDHr = 66 mM [6] | |
| KmGAPDHGAP = 3.10 mM [35] | ||
| KmGAPDHNADP = 0.20 mM [35] | ||
| KmGAPDHNADPH = 0.094 mM [35] | ||
| KmGAPDHBPG = 0.089 mM [35] | ||
| KmGAPDHhospahte = 108.52 mM [35] | ||
| PGK for S. solfataricus | VmrPGK = 73 mM/min [34] | |
| KmPGKADP = 0.374 mM [35] | ||
| KmPGKATP = 9.303 mM [35] | ||
| KmPGKBPG = 0.008 mM [35] | ||
| KmPGK3PG = 0.567 mM [35] | ||
| KiADP = 1.14 mM [35] | ||
| KeqPGK = 3793 mM 3 | ||
| GAPN for S. solfataricus | VmGAPN = 20 mM/min [34] | |
| KmGAP = 0.02 mM [34] 4 | ||
| KmNADP = 0.09 mM [34] 4 | ||
| BPG degradation | KdBPG = 1.058 min−1 [7] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Kouril, T.; Snoep, J.L.; Siebers, B.; Barberis, M.; Westerhoff, H.V. The Peculiar Glycolytic Pathway in Hyperthermophylic Archaea: Understanding Its Whims by Experimentation In Silico. Int. J. Mol. Sci. 2017, 18, 876. https://doi.org/10.3390/ijms18040876
Zhang Y, Kouril T, Snoep JL, Siebers B, Barberis M, Westerhoff HV. The Peculiar Glycolytic Pathway in Hyperthermophylic Archaea: Understanding Its Whims by Experimentation In Silico. International Journal of Molecular Sciences. 2017; 18(4):876. https://doi.org/10.3390/ijms18040876
Chicago/Turabian StyleZhang, Yanfei, Theresa Kouril, Jacky L. Snoep, Bettina Siebers, Matteo Barberis, and Hans V. Westerhoff. 2017. "The Peculiar Glycolytic Pathway in Hyperthermophylic Archaea: Understanding Its Whims by Experimentation In Silico" International Journal of Molecular Sciences 18, no. 4: 876. https://doi.org/10.3390/ijms18040876
APA StyleZhang, Y., Kouril, T., Snoep, J. L., Siebers, B., Barberis, M., & Westerhoff, H. V. (2017). The Peculiar Glycolytic Pathway in Hyperthermophylic Archaea: Understanding Its Whims by Experimentation In Silico. International Journal of Molecular Sciences, 18(4), 876. https://doi.org/10.3390/ijms18040876