State of the Art on Functional Virgin Olive Oils Enriched with Bioactive Compounds and Their Properties



 ,
,  , ,
, , 
Abstract
:1. Market Needs for Functional Oils
2. Sources of Natural Bioactive Ingredients to Enrich Oils
- Hot pepper, garlic, oregano and rosemary [53]
- Rosemary, lavender, sage, menthe, basil, lemon and thyme [54]
- Garlic, lemon, oregano, hot pepper, and rosemary [55]
- Basil [56]
- Lemon and thyme [57]
- Thyme [36]
- Thyme [60]
- Garlic, hot chili peppers, laurel, oregano and pepper [61].
- Sweet lemon and sweet orange peels [62]
- Basil, chili and garlic [63]
- Caraway [64]
- Thyme and oregano [65]
3. Optimization of the Development of Novel Functional Oils
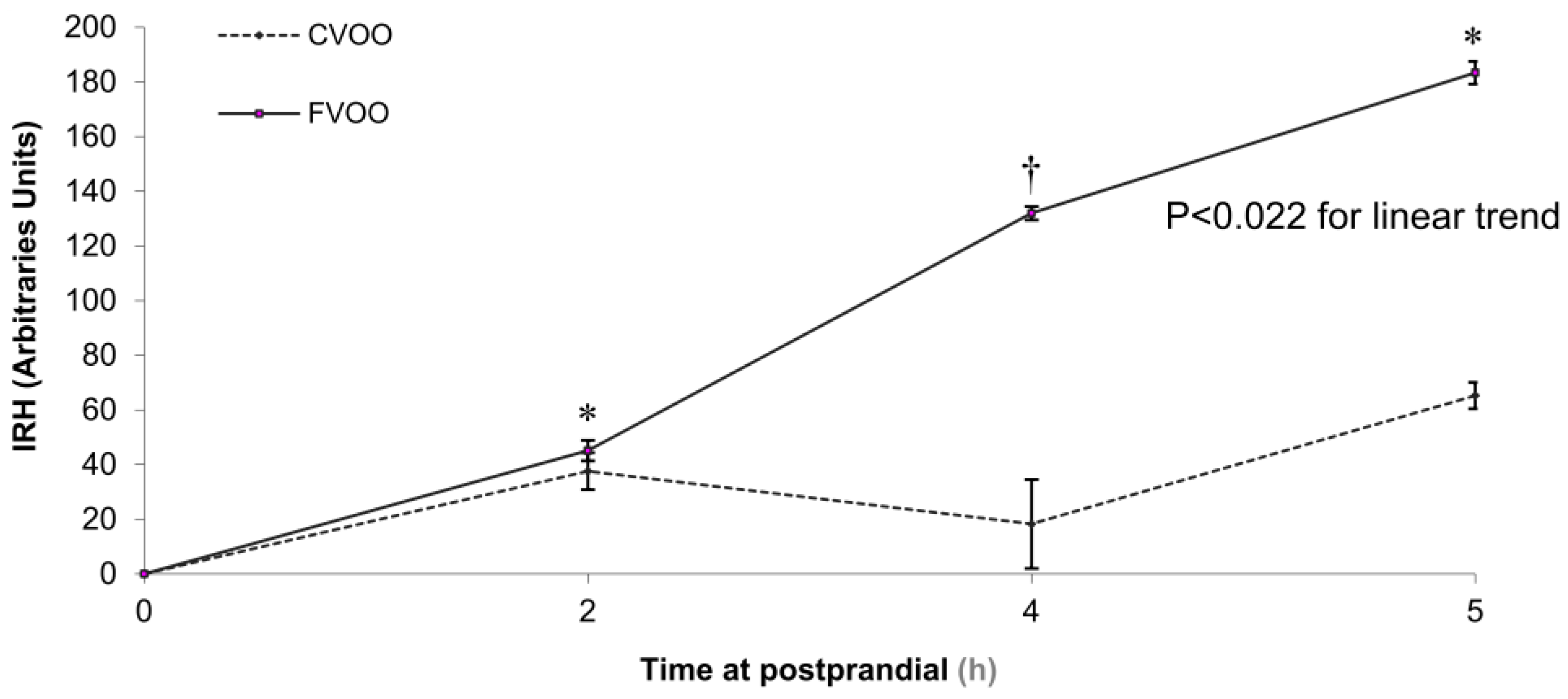
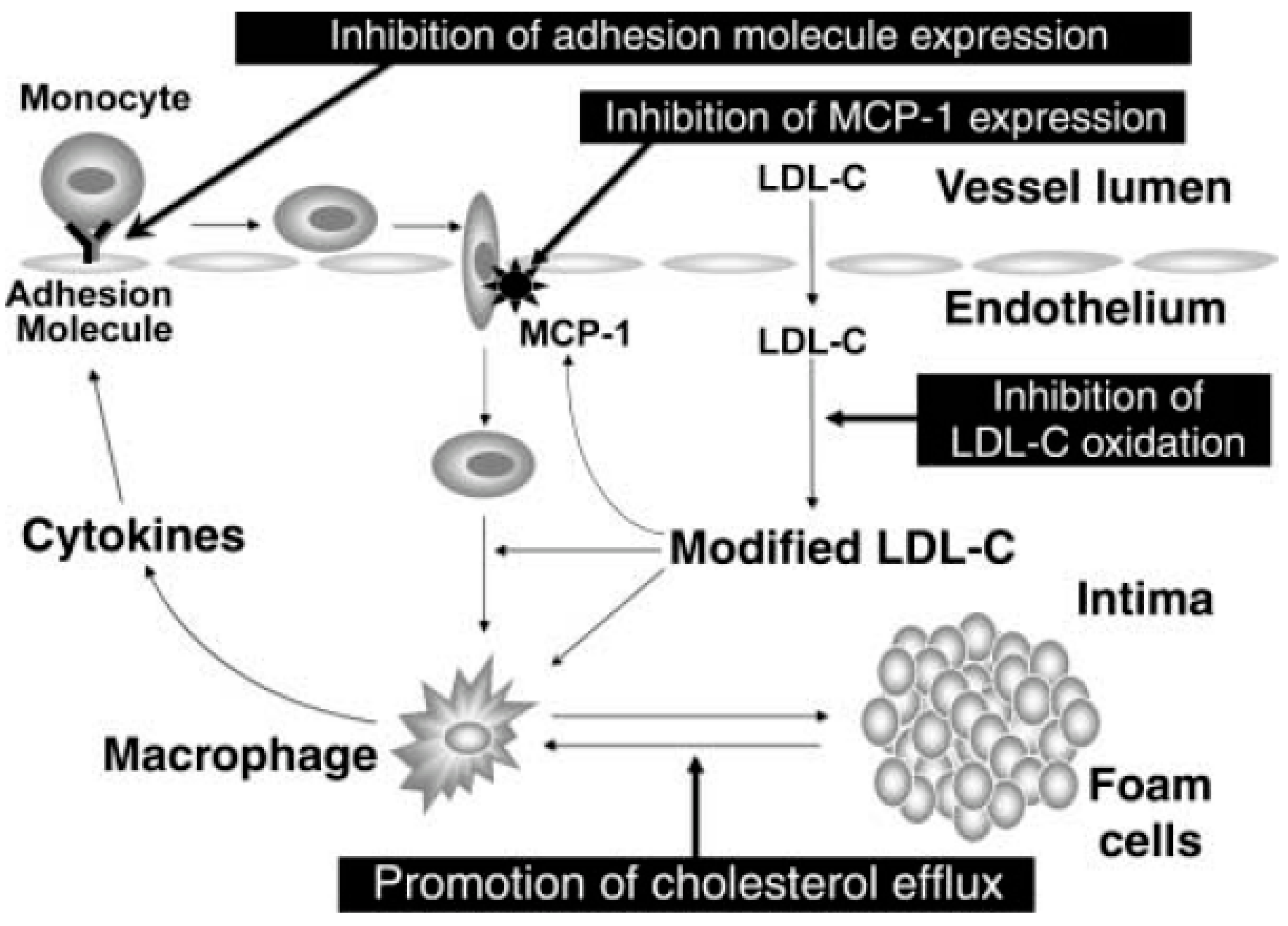
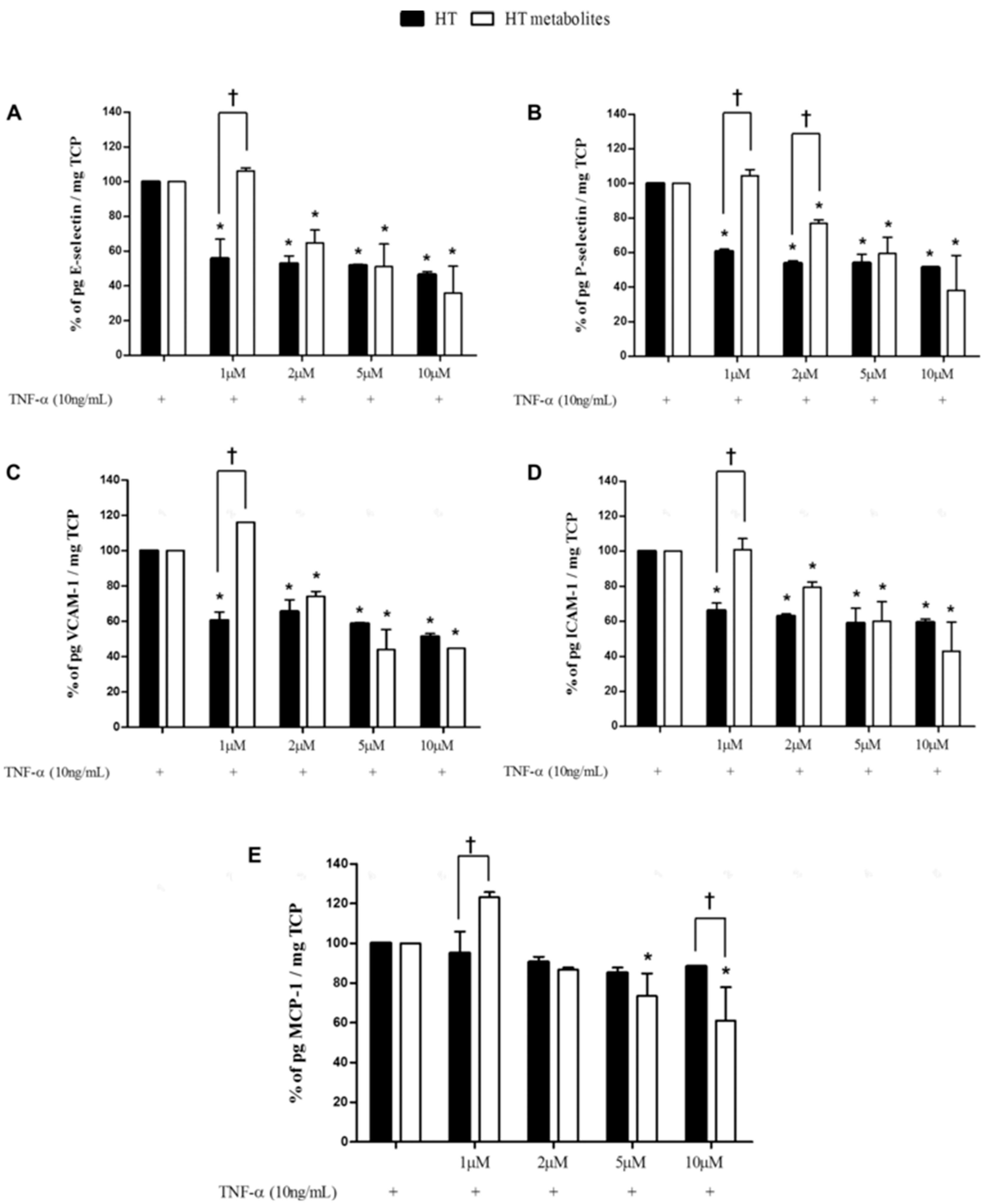
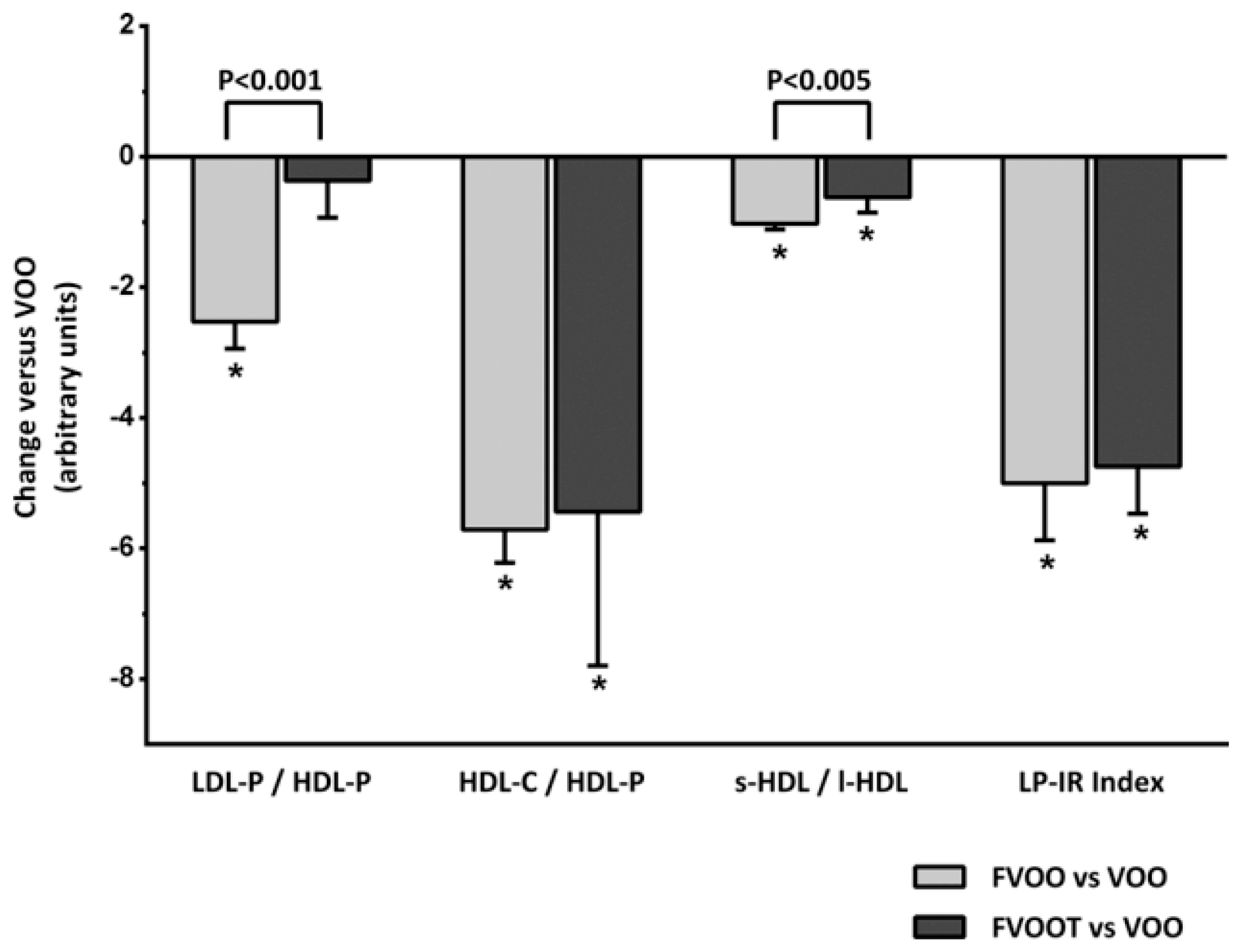
4. Positive Outcomes in Prevention of Disease and Management of Health
5. Future Directions and Opportunities in Oils Technology Research
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ABCA1/G1 | ATP binding cassette transporter A1 or G1 |
| ABTS | 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid) |
| Apo AI | Apolipoprotein AI |
| ASE | Accelerated extraction system |
| CAT | Catalase |
| CE | Cholesteryl ester |
| CETP | Cholesteryl ester transfer protein |
| CHD | Coronary heart disease |
| CRP | C-reactive protein |
| CVD | Cardiovascular disease |
| DNA | Deoxyribonucleic acid |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| ED | Endothelial dysfunction |
| EFSA | European Food Safety Authority |
| En-VOO | Enriched VOO |
| EVOO | Extra virgin olive oil |
| EUROLIVE | The Effect of Olive Oil on Oxidative Damage on European Populations |
| FOSHU | Foods for Specified Health Use |
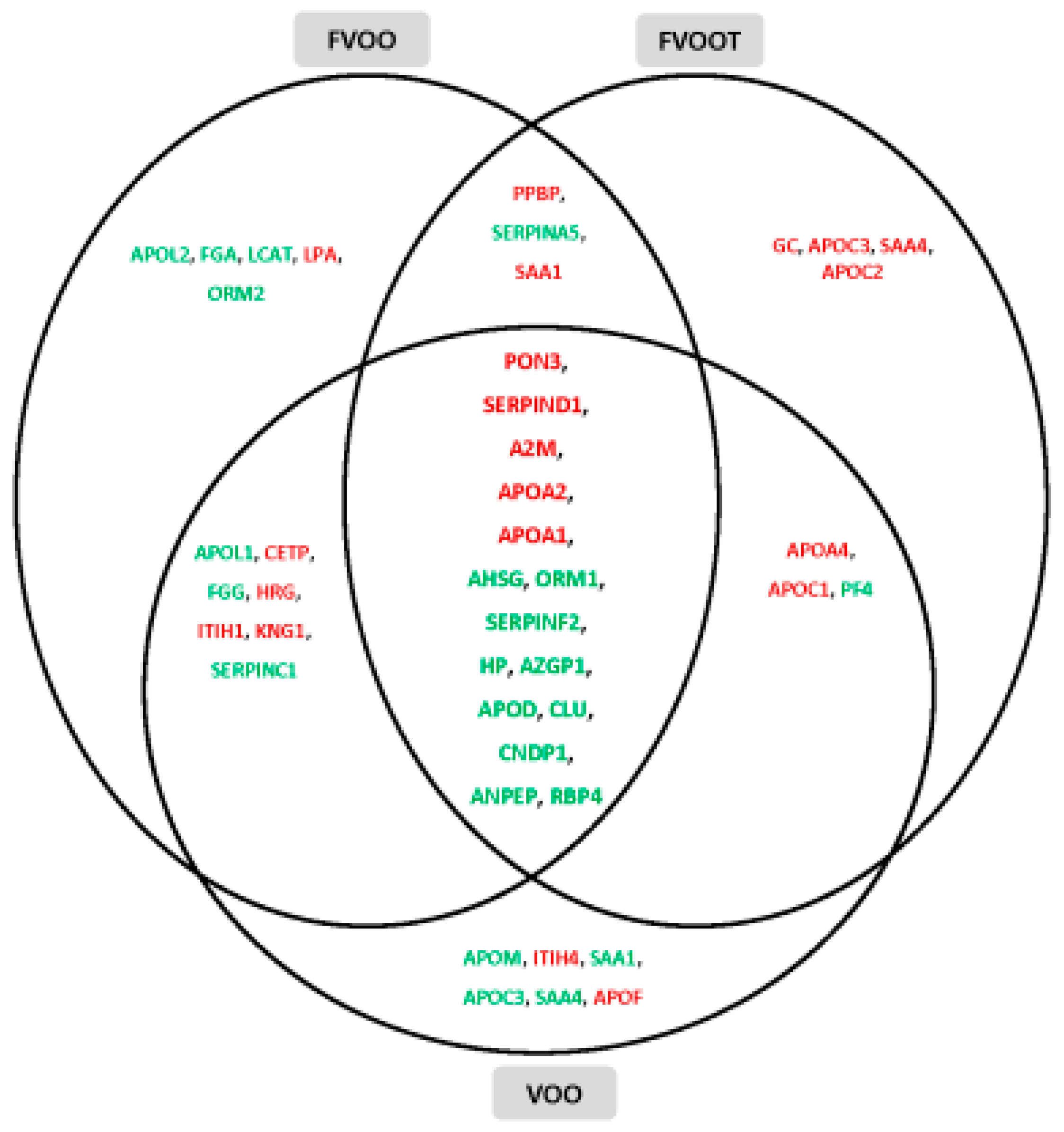
| FVOO | Functional virgin olive oil enriched with its phenolic compounds (500 ppm) |
| FVOOT | Functional virgin olive enriched with its phenolic compounds (250 ppm) and those from thyme (250 ppm) |
| GALT | Gut-associated lymphoid tissue |
| GCL | Glutamate-cysteine ligase |
| GPx | Glutathione peroxidase |
| GR | Glutathione reductase |
| GSH | Glutathione |
| GSSG | Oxidized glutathione |
| HAEC | Human aortic endothelial cells |
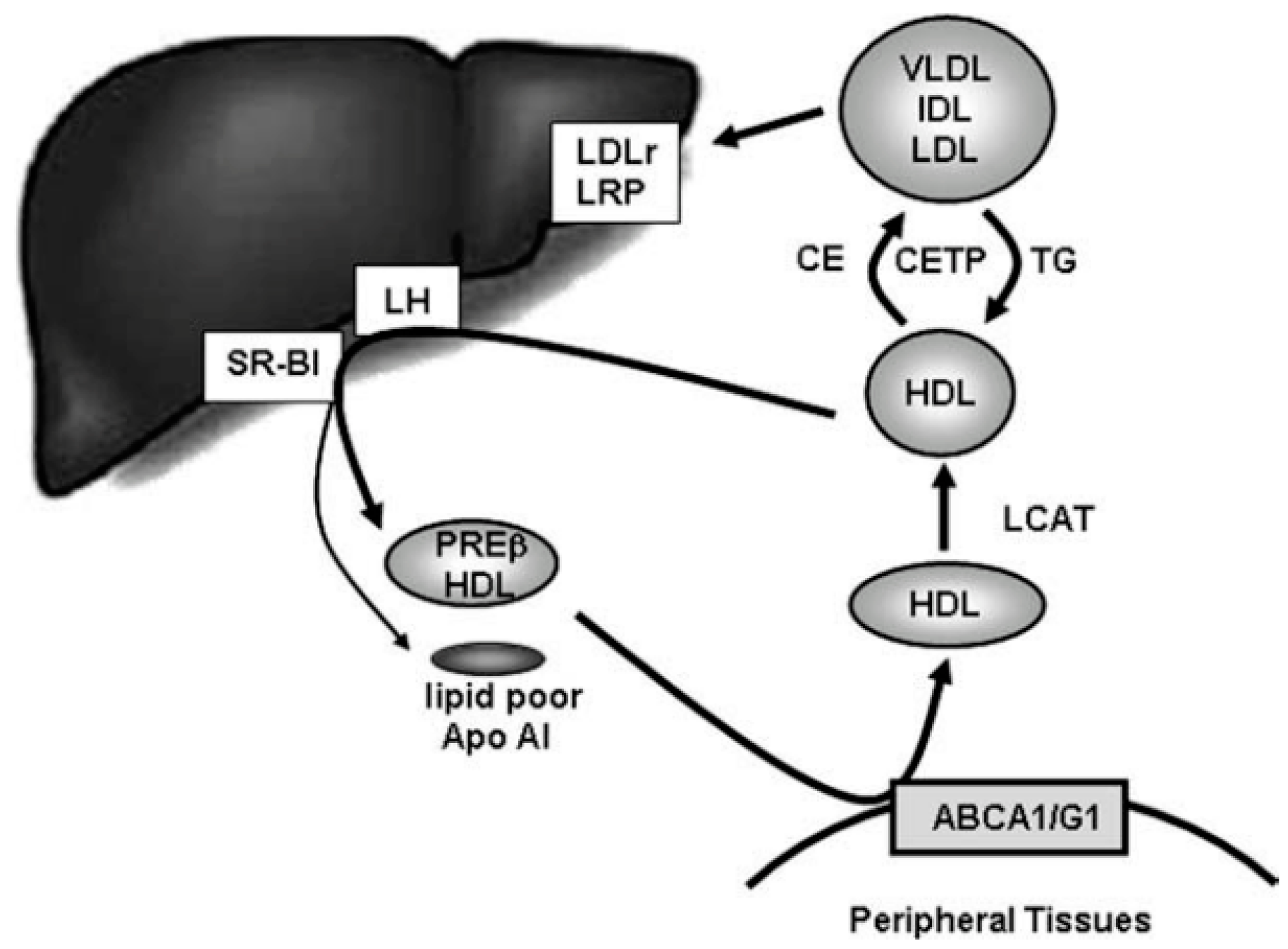
| HDL | High-density lipoprotein |
| HDL-C | HDL-cholesterol |
| HDL-CE | High-density lipoprotein- cholesteryl ester |
| HDL-P | HDL-particles |
| HDL-TG | High-density lipoprotein- triglycerides |
| HPLC | High performance liquid chromatography |
| HTyr/HT | Hydroxytyrosol |
| IBD | Inflammatory bowel disease |
| ICAM-1 | Intercellular adhesion molecule-1 |
| IDL | Intermediate-density lipoproteins |
| IgA | Immunoglobulin A |
| IRH | Ischemic reactive hyperemia |
| 8-iso PGF2α | F2-isoprostane, 8-isoprostaglandin |
| Keap1 | Kelch-like ECH-associated protein 1 |
| LCAT | Lecithin cholesterol acyl transferase |
| LDL | Low-density lipoprotein |
| LDL-C | LDL-cholesterol |
| LDL-P | LDL-particles |
| LDLr | LDL receptor |
| LH | Hepatic lipase |
| l-HDL | Large HDL |
| LLE | Liquid–liquid extraction |
| LP-IR | Lipoprotein insulin resistance index |
| LRP | LDL receptor related proteins |
| LXR | Liver X receptor |
| MAE | Microwave assisted extraction |
| MCFA | Medium chain fatty acids |
| MCP-1 | Monocyte chemotactic protein-1 |
| Met | Methionine |
| MetSO | Methionine sulfoxide |
| NMR | Nuclear magnetic resonance |
| NO | Nitric oxide |
| Nrf2 | Nuclear factor, erythroid 2-like 2 |
| 8-OHdG | 8-hydroxy-2′-deoxyguanosine |
| OO | Olive oil |
| ORAC | Oxygen radical antioxidant capacity |
| OSI | Oxidative stability index |
| PC | Phenolic compounds |
| PREDIMED | Prevention with Mediterranean diet |
| PV | Peroxide value |
| RCT | Reverse cholesterol transport |
| ROS | Reactive oxygen species |
| RXR | Retinoid X receptor |
| SEM | Standard error of the mean |
| SFE | Supercritical fluid extraction |
| SLE | Solid-liquid extraction |
| s-HDL | Small HDL |
| SOD | Superoxide dismutase |
| SRBI | Scavenger receptor class B type I |
| TC | Total cholesterol |
| TCP | Tissue culture plate |
| TG | Triglycerides |
| TNF-α | Tumour necrosis factor-α |
| Tyr | Tyrosol |
| USAE | Ultrasound assisted extraction |
| VCAM-1 | Vascular cell adhesion molecule-1 |
| VLDL | Very low-density lipoprotein |
| VOO | Virgin olive oil |
References
- Martirosyam, D.M.; Singh, J. A new definition of functional food by FFC: What makes a new definition unique? Func. Food Health Dis. 2015, 5, 209–213. [Google Scholar]
- Willett, W.C.; Sacks, F.; Trichopoulou, A.; Drescher, G.; Ferro-Luzzi, A.; Helsing, E.; Trichopoulos, D. Mediterranean diet pyramid: A cultural model for healthy eating. Am. J. Clin. Nutr. 1995, 61 (Suppl. S6), 1402S–1406S. [Google Scholar] [PubMed]
- Casas, R.; Sacanella, E.; Urpí-Sardà, M.; Chiva-Blanch, G.; Ros, E.; Martínez-González, M.-A.; Covas, M.-I.; Lamuela-Raventos, R.M.; Salas-Salvadó, J.; Fiol, M.; et al. The effects of the Mediterranean diet on biomarkers of vascular wall inflammation and plaque vulnerability in subjects with high risk for cardiovascular disease. A randomized trial. PLoS ONE 2014, 9, e100084. [Google Scholar] [CrossRef] [PubMed]
- Díaz-López, A.; Bulló, M.; Martínez-González, M.Á.; Guasch-Ferré, M.; Ros, E.; Basora, J.; Covas, M.-I.; Del Carmen López-Sabater, M.; Salas-Salvadó, J. Effects of mediterranean diets on kidney function: A report from the PREDIMED trial. Am. J. Kidney Dis. 2012, 60, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Fitó, M.; Estruch, R.; Salas-Salvadó, J.; Martínez-Gonzalez, M.A.; Arós, F.; Vila, J.; Corella, D.; Díaz, O.; Sáez, G.; De La Torre, R.; et al. Effect of the Mediterranean diet on heart failure biomarkers: A randomized sample from the PREDIMED trial. Eur. J. Heart Fail. 2014, 16, 543–550. [Google Scholar] [CrossRef] [PubMed]
- García-López, M.; Toledo, E.; Beunza, J.J.; Aros, F.; Estruch, R.; Salas-Salvadó, J.; Corella, D.; Ros, E.; Covas, M.I.; Gómez-Gracia, E.; et al. Mediterranean diet and heart rate: The PREDIMED randomised trial. Int. J. Cardiol. 2014, 171, 299–301. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Bulló, M.; Babio, N.; Martínez-González, M.A.; Estruch, R.; Covas, M.-I.; Wärnberg, J.; Arós, F.; Lapetra, J.; Serra-Majem, L.; et al. Mediterranean diet and risk of hyperuricemia in elderly participants at high cardiovascular risk. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Martínez-González, M.A.; García-Arellano, A.; Toledo, E.; Salas-Salvadó, J.; Buil-Cosiales, P.; Corella, D.; Covas, M.I.; Schröder, H.; Arós, F.; Gómez-Gracia, E.; et al. A 14-item mediterranean diet assessment tool and obesity indexes among high-risk subjects: The PREDIMED trial. PLoS ONE 2012, 7, e43134. [Google Scholar] [CrossRef] [PubMed]
- Mayneris-Perxachs, J.; Sala-Vila, A.; Chisaguano, M.; Castellote, A.I.; Estruch, R.; Covas, M.I.; Fitó, M.; Salas-Salvadó, J.; Martínez-González, M.A.; Lamuela-Raventós, R.; et al. Effects of 1-year intervention with a Mediterranean diet on plasma fatty acid composition and metabolic syndrome in a population at high cardiovascular risk. PLoS ONE 2014, 9, e85202. [Google Scholar] [CrossRef] [PubMed]
- Mitjavila, M.T.; Fandos, M.; Salas-Salvadó, J.; Covas, M.-I.; Borrego, S.; Estruch, R.; Lamuela-Raventós, R.; Corella, D.; Martínez-Gonzalez, M.T.; Sánchez, J.M.; et al. The Mediterranean diet improves the systemic lipid and DNA oxidative damage in metabolic syndrome individuals. A randomized, controlled, trial. Clin. Nutr. 2013, 32, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rejón, A.I.; Castro-Quezada, I.; Ruano-Rodríguez, C.; Ruiz-López, M.D.; Sánchez-Villegas, A.; Toledo, E.; Artacho, R.; Estruch, R.; Salas-Salvadó, J.; Covas, M.I.; et al. Effect of a Mediterranean Diet Intervention on Dietary Glycemic Load and Dietary Glycemic Index: The PREDIMED Study. J. Nutr. Metab. 2014, 2014, 985373. [Google Scholar] [CrossRef] [PubMed]
- Servili, M.; Montedoro, G.F. Contribution of phenolic compounds to virgin olive oil quality. Eur. J. Lipid Sci. Technol. 2002, 104, 602–613. [Google Scholar] [CrossRef]
- Catalán, U.; López de las Hazas, M.C.; Rubió, L.; Fernández-Castillejo, S.; Pedret, A.; de la Torre, R.; Motilva, M.J.; Solà, R. Protective effect of hydroxytyrosol and its predominant plasmatic human metabolites against endothelial dysfunction in human aortic endothelial cells. Mol. Nutr. Food. Res. 2015, 59, 2523–2536. [Google Scholar] [CrossRef] [PubMed]
- Covas, M.-I.; De La Torre, K.; Farré-Albaladejo, M.; Kaikkonen, J.; Fitó, M.; López-Sabater, C.; Pujadas-Bastardes, M.A.; Joglar, J.; Weinbrenner, T.; Lamuela-Raventós, R.M.; et al. Postprandial LDL phenolic content and LDL oxidation are modulated by olive oil phenolic compounds in human. Free Radic. Biol. Med. 2006, 40, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Oliveras-López, M.J.; Berná, G.; Jurado-Ruiz, E.; López-García de la Serrana, H.; Martín, F. Consumption of extra-virgin olive oil rich in phenolic compounds has beneficial antioxidant effects in healthy human adults. J. Funct. Foods 2014, 10, 475–484. [Google Scholar] [CrossRef]
- Martín-Peláez, S.; Mosele, J.I.; Pizarro, N.; Farràs, M.; de la Torre, R.; Subirana, I.; Pérez-Cano, F.J.; Castañer, O.; Solà, R.; Fernandez-Castillejo, S.; et al. Effect of virgin olive oil and thyme phenolic compounds on blood lipid profile: Implications of human gut microbiota. Eur. J. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Romeu, M.; Rubió, L.; Sánchez-Martos, V.; Castañer, O.; De La Torre, R.; Valls, R.M.; Ras, R.; Pedret, A.; Catalán, Ú.; López De Las Hazas, M.D.C.; et al. Virgin olive oil enriched with its own phenols or complemented with thyme phenols improves DNA protection against oxidation and antioxidant enzyme activity in hyperlipidemic subjects. J. Agric. Food Chem. 2016, 64, 1879–1888. [Google Scholar] [CrossRef] [PubMed]
- Rubió, L.; Serra, A.; Oliver Chen, C.-Y.; Macià, A.; Covas, M.I.; Sola, R.; Motilva, M.J. Effect of the co-occurring components from olive oil and thyme extracts on the antioxidant status and its bioavailability in an acute ingestion in rats. Food Funct. 2014, 5, 740–747. [Google Scholar] [CrossRef] [PubMed]
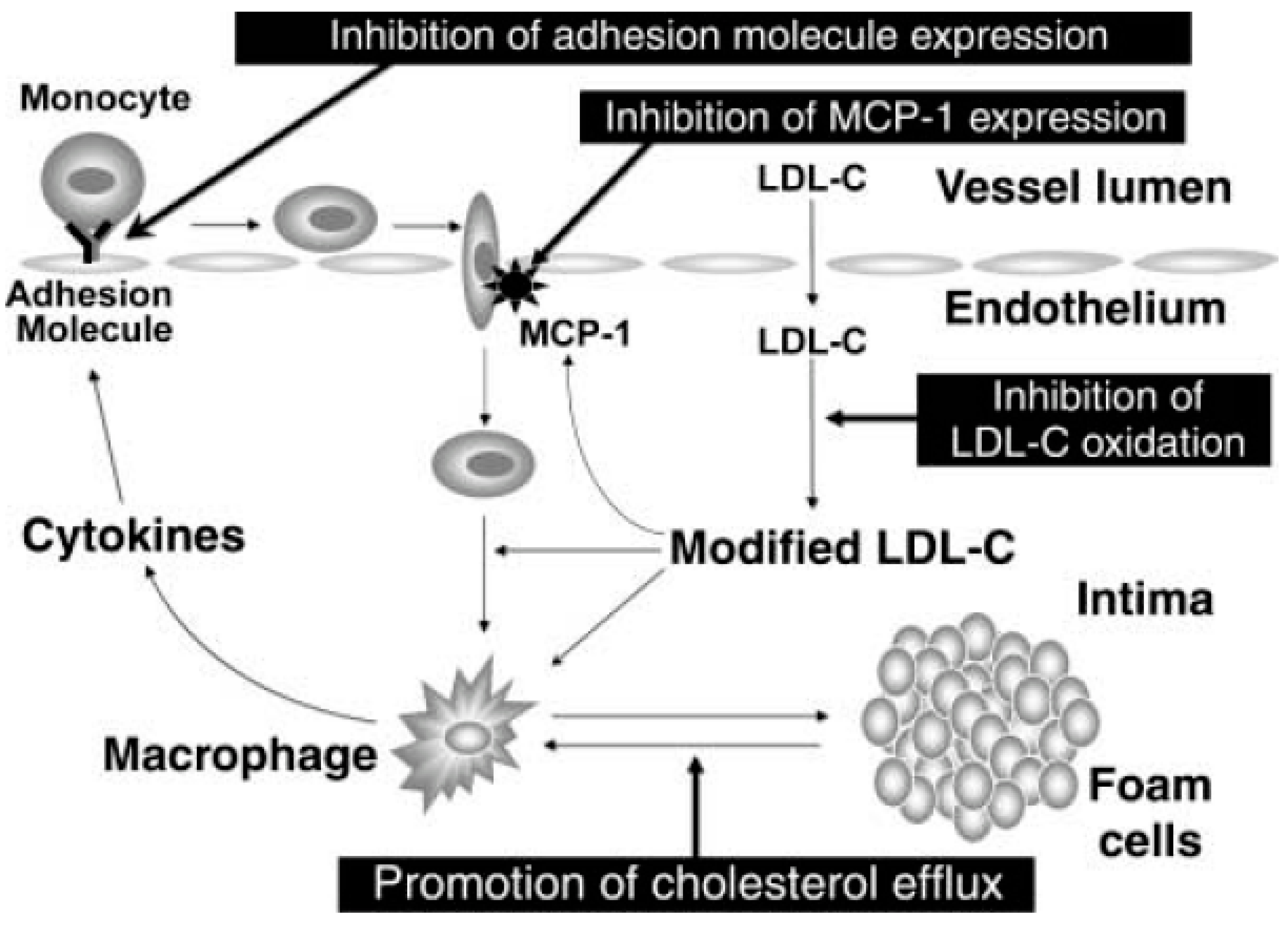
- Covas, M.-I. Olive oil and the cardiovascular system. Pharmacol. Res. 2007, 55, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Fitó, M.; De La Torre, R.; Covas, M.-I. Olive oil and oxidative stress. Mol. Nutr. Food. Res. 2007, 51, 1215–1224. [Google Scholar] [CrossRef] [PubMed]
- Covas, M.-I.; Nyyssönen, K.; Poulsen, H.E.; Kaikkonen, J.; Zunft, H.-J.F.; Kiesewetter, H.; Gaddi, A.; De La Torre, R.; Mursu, J.; Bäumler, H.; et al. The effect of polyphenols in olive oil on heart disease risk factors: A randomized trial. Ann. Intern. Med. 2006b, 145, 333–341. [Google Scholar] [CrossRef]
- Gómez, M.; Vila, J.; Elosua, R.; Molina, L.; Bruguera, J.; Sala, J.; Masià, R.; Covas, M.I.; Marrugat, J.; Fitó, M. Relationship of lipid oxidation with subclinical atherosclerosis and 10-year coronary events in general population. Atherosclerosis 2014, 232, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Hernáez, A.; Fernández-Castillejo, S.; Farràs, M.; Catalán, U.; Subirana, I.; Montes, R.; Solà, R.; Muñoz-Aguayo, D.; Gelabert-Gorgues, A.; Díaz-Gil, O.; et al. Olive oil polyphenols enhance high-density lipoprotein function in humans: A randomized controlled trial. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2115–2119. [Google Scholar] [CrossRef] [PubMed]
- Commission Regulation (EU). No. 432/2012 of 16 May 2012 Establishing a List of Permitted Health Claims Made on Foods, Other than Those Referring to the Reduction of Disease Risk and to Children’s Development and Health; Commission Regulation (EU): London, UK, 2012. [Google Scholar]
- Covas, M.I.; Konstantinidou, V.; Fitó, M. Olive oil and cardiovascular health. J. Cardiovasc. Pharmacol. 2009, 54, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Covas, M.-I.; De La Torre, R.; Fitó, M. Virgin olive oil: A key food for cardiovascular risk protection. Br. J. Nutr. 2015, 113, S19–S28. [Google Scholar] [CrossRef] [PubMed]
- Covas, M.-I.; Fitó, M.; De La Torre, R. Minor bioactive olive oil components and health: Key data for their role in providing health benefits in humans. In Olive and Olive Oil Bioactive Constituents; Boskou, B., Ed.; AOCS Press: Urbana, IL, USA, 2015; pp. 31–52. [Google Scholar]
- Hernáez, A.; Remaley, A.T.; Farràs, M.; Fernández-Castillejo, S.; Subirana, I.; Schröder, H.; Fernández-Mampel, M.; Muñoz-Aguayo, D.; Sampson, M.; Solà, R.; et al. Olive oil polyphenols decrease LDL concentrations and LDL atherogenicity in men in a randomized controlled trial. J. Nutr. 2015, 145, 1692–1697. [Google Scholar] [CrossRef] [PubMed]
- Martín-Peláez, S.; Covas, M.I.; Fitó, M.; Kusar, A.; Pravst, I. Health effects of olive oil polyphenols: Recent advances and possibilities for the use of health claims. Mol. Nutr. Food Res. 2013, 57, 760–771. [Google Scholar] [CrossRef] [PubMed]
- Medina-Remón, A.; Tresserra-Rimbau, A.; Pons, A.; Tur, J.A.; Martorell, M.; Ros, E.; Buil-Cosiales, P.; Sacanella, E.; Covas, M.I.; Corella, D.; et al. Effects of total dietary polyphenols on plasma nitric oxide and blood pressure in a high cardiovascular risk cohort. The PREDIMED randomized trial. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Segade, M.; Bermejo, R.; Silva, A.; Paiva-Martins, F.; Gil-Longo, J.; Campos-Toimil, M. Involvement of endothelium in the vasorelaxant effects of 3,4-DHPEA and 3,4-DHPEA-EDA, two major functional bioactives in olive oil. J. Funct. Foods 2016, 23, 637–646. [Google Scholar] [CrossRef]
- Achat, S.; Tomao, V.; Madani, K.; Chibane, M.; Elmaataoui, M.; Dangles, O.; Chemad, F. Direct enrichment of olive oil in oleuropein by ultrasound-assisted maceration at laboratory and pilot plant scale. Ultrason. Sonochem. 2012, 19, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Baba, N.H.; Antoniades, K.; Habbal, Z. Effects of dietary canola, olive, and linolenic acid enriched olive oils on plasma lipids, lipid peroxidation and lipoprotein lipase activity in rats. Nutr. Res. 1999, 19, 601–612. [Google Scholar] [CrossRef]
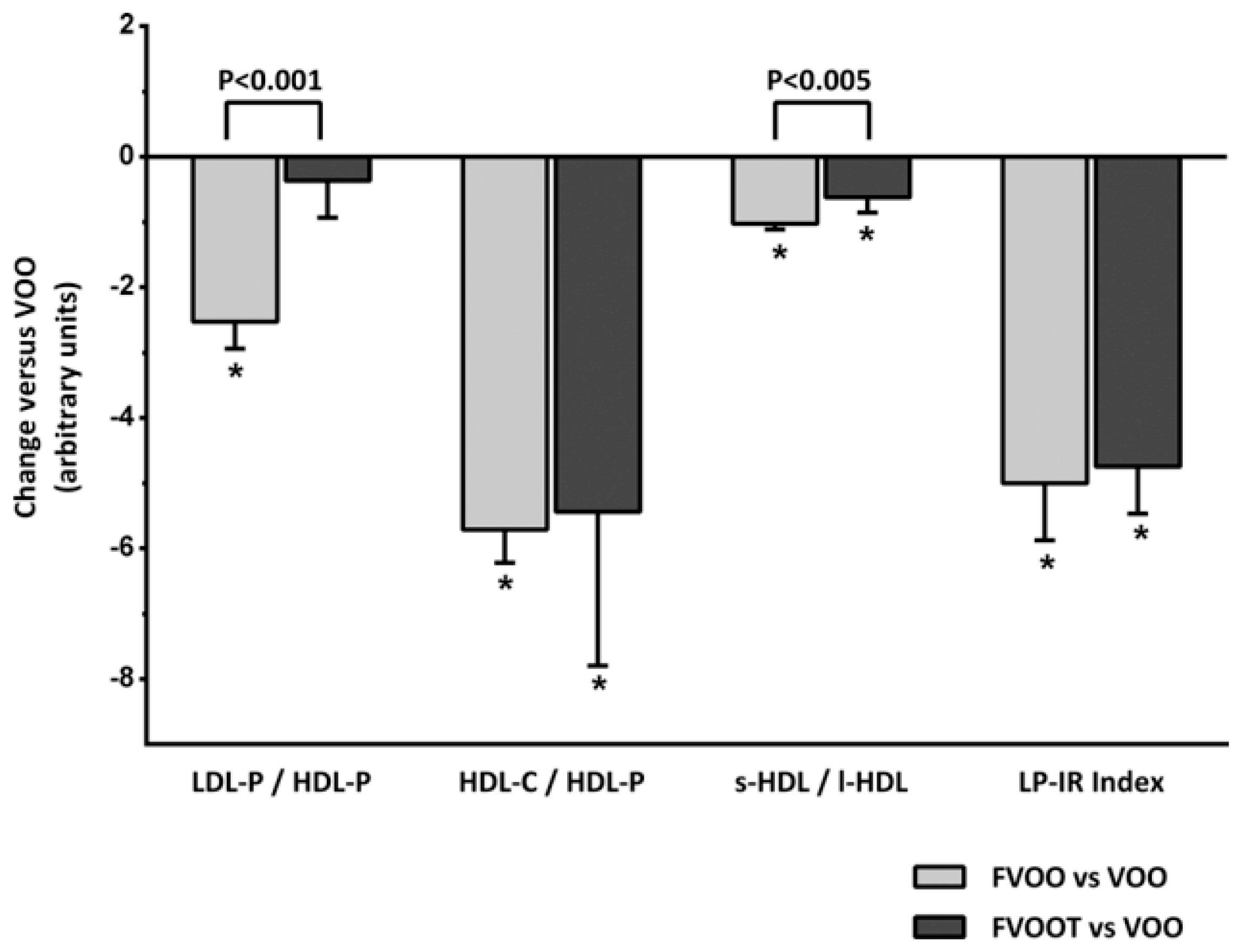
- Fernández-Castillejo, S.; Valls, R.-M.; Castañer, O.; Rubió, L.; Catalán, Ú.; Pedret, A.; Macià, A.; Sampson, M.L.; Covas, M.-I.; Fitó, M.; et al. Polyphenol rich olive oils improve lipoprotein particle atherogenic ratios and subclasses profile: A randomized, crossover, controlled trial. Mol. Nutr. Food Res. 2016, 60, 1544–1554. [Google Scholar] [CrossRef] [PubMed]
- Garrido, M.; González-Flores, D.; Marchena, A.M.; Prior, E.; García-Parra, J.; Barriga, C.; Rodríguez Moratinos, A.B. A lycopene-enriched virgin olive enhances antioxidant status in humans. J. Sci. Food Agric. 2013, 93, 1820–1826. [Google Scholar] [CrossRef] [PubMed]
- Rubió, L.; Motilva, M.-J.; Macià, A.; Ramo, T.; Romero, M.-P. Development of a phenol-enriched olive oil with both its own phenolic compounds and complementary phenols from thyme. J. Agric. Food Chem. 2012, 60, 3105–3112. [Google Scholar] [CrossRef] [PubMed]
- Suárez, M.; Macià, A.; Romero, M.-P.; Motilva, M.-J. Improved liquid chromatography tandem mass spectrometry method for the determination of phenolic compounds in virgin olive oil. J. Chromatogr. A 2008, 1214, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Suárez, M.; Romero, M.-P.; Motilva, M.-J. Development of a phenol-enriched olive oil with phenolic compounds from olive cake. J. Agric. Food Chem. 2010, 58, 10396–10403. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, M.; Volkova, N.; Coleman, R.; Almagor, Y.; Aviram, M. Antiatherogenicity of extra virgin olive oil and its enrichment with green tea polyphenols in the atherosclerotic apolipoprotein-E-deficient mice: Enhanced macrophage cholesterol efflux. J. Nutr. Biochem. 2008, 19, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Japón-Luján, R.; Janeiro, P.; Luque de Castro, M.D. Solid-liquid transfer of biophenols from olive leaves for the enrichment of edible oils by a dynamic ultrasound-assisted approach. J. Agric. Food Chem. 2008, 56, 7231–7235. [Google Scholar] [CrossRef] [PubMed]
- Japón-Luján, R.; Luque de Castro, M.D. Liquid–liquid extraction for the enrichment of edible oils with phenols from olive leaf extracts. J. Agric. Food Chem. 2008a, 56, 2505–2511. [Google Scholar] [CrossRef] [PubMed]
- Sánchez de Medina, V.; Priego-Capote, F.; Luque de Castro, M.D. Characterization of refined edible oils enriched with phenolic extracts from olive leaves and pomace. J. Agric. Food Chem. 2012, 60, 5866–5873. [Google Scholar]
- Sanchez de Medina, V.; Priego-Capote, F.; Jimenez-Ot, C.; Luque de Castro, M.D. Quality and Stability of Edible Oils Enriched with Hydrophilic Antioxidants from the Olive Tree: The Role of Enrichment Extracts and Lipid Composition. J. Agric. Food Chem. 2011, 59, 11432–11441. [Google Scholar] [CrossRef] [PubMed]
- Chiou, A.; Kalogeropoulos, N.; Salta, F.N.; Efstathiou, P.; Andrikopoulos, N.K. Pan-frying of French fries in three different edible oils enriched with olive leaf extract: Oxidative stability and fate of microconstituents. LWT Food Sci. Technol. 2009, 42, 1090–1097. [Google Scholar] [CrossRef]
- Zribi, A.; Gargouri, B.; Jabeur, H.; Rebaï, A.; Abdelhedi, R.; Bouaziz, M. Enrichment of pan-frying refined oils with olive leaf phenolic-rich extract to extend the usage life. Eur. J. Lipid Sci. Technol. 2013, 115, 1443–1453. [Google Scholar] [CrossRef]
- Inarejos-García, A.M.; Salvador, M.D.; Fregapane, G. Effect of crushing on olive paste and virgin olive oil minor components. Eur. Food Res. Technol. 2011, 232, 441–451. [Google Scholar] [CrossRef]
- Suarez, M.; Romero, M.P.; Ramo, T.; Motilva, M.J. Stability of a phenol-enriched olive oil during storage. Eur. J. Lipid Sci. Technol. 2011, 113, 894–903. [Google Scholar] [CrossRef]
- Delgado-Adamez, J.; Baltasar, M.N.F.; Yuste, M.C.A.; Martin-Vertedor, D. Oxidative stability, phenolic compounds and antioxidant potential of a virgin olive oil enriched with natural bioactive compounds. J. Oleo Sci. 2014, 63, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Artajo, L.S.; Romero, M.P.; Morelló, J.R.; Motilva, M.J. Enrichment of refined olive oil with phenolic compounds: Evaluation of their antioxidant activity and their effect on the bitter index. J. Agric. Food Chem. 2006, 54, 6079–6088. [Google Scholar] [CrossRef] [PubMed]
- Fregapane, G.; Salvador, M.D. Production of superior quality extra virgin olive oil modulating the content and profile of its minor components. Food Res. Int. 2013, 54, 1907–1914. [Google Scholar] [CrossRef]
- Caporaso, N.; Paduano, A.; Nicoletti, G.; Sacchi, R. Capsaicinoids, antioxidant activity, and volatile compounds in olive oil flavored with dried chili pepper (Capsicum annuum). Eur. J. Lipid Sci. Technol. 2013, 115, 1434–1442. [Google Scholar] [CrossRef]
- Gouveia, A.F.; Duarte, C.; Beirão da Costa, M.L.; Bernardo-Gil, M.G.; Moldão-Martins, M. Oxidative stability of olive oil flavoured by Capsicum frutescens supercritical fluid extracts. Eur. J. Lipid Sci. Technol. 2006, 108, 421–428. [Google Scholar] [CrossRef]
- Gambacorta, G.; Faccia, M.; Pati, S.; Lamacchia, C.; Baiano, A.; La Notte, E. Changes in the chemical and sensorial profile of extra virgin olive oils flavored with herbs and spices during storage. J. Food Lipids 2007, 14, 202–215. [Google Scholar] [CrossRef]
- Ayadi, M.A.; Grati-Kamoun, N.; Attia, H. Physico-chemical change and heat stability of extra virgin olive oils flavoured by selected Tunisian aromatic plants. Food Chem. Toxicol. 2009, 47, 2613–2619. [Google Scholar] [CrossRef] [PubMed]
- Baiano, A.; Terracone, C.; Gambacorta, G.; La Notte, E. Changes in quality indices, phenolic content and antioxidant activity of flavored olive oils during storage. J. Am. Oil Chem. Soc. 2009, 86, 1083–1092. [Google Scholar] [CrossRef]
- Veillet, S.; Tomao, V.; Chemat, F. Ultrasound assisted maceration: An original procedure for direct aromatisation of olive oil with basil. Food Chem. 2010, 123, 905–911. [Google Scholar] [CrossRef]
- Issaoui, M.; Flamini, G.; Hajaij, M.E.; Cioni, P.L.; Hammami, M. Oxidative evolution of virgin and flavored olive oils under thermo-oxidation processes. J. Am. Oil Chem. Soc. 2011, 88, 1339–1350. [Google Scholar] [CrossRef]
- Asensio, C.M.; Nepote, V.; Grosso, N.R. Consumers’ acceptance and quality stability of olive oil flavoured with essential oils of different oregano species. Int. J. Food Sci. Technol. 2013, 48, 2417–2428. [Google Scholar] [CrossRef]
- Peñalvo, G.C.; Robledo, V.R.; Callado, C.S.-C.; Santander-Ortega, M.J.; Castro-Vázquez, L.; Victoria Lozano, M.; Arroyo-Jiménez, M.M. Improving green enrichment of virgin olive oil by oregano. Effects on antioxidants. Food Chem. 2016, 197, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.G.; Antunes, M.D.; Rohaim, A.; Figueiredo, A.C.; Pedro, L.G.; Barroso, J.G. Stability of fried olive and sunflower oils enriched with Thymbra capitata essential oil. Czech J. Food Sci. 2014, 32, 102–108. [Google Scholar]
- Sousa, A.; Casal, S.; Malheiro, R.; Lamas, H.; Bento, A.; Pereira, J.A. Aromatized olive oils: Influence of flavouring in quality, composition, stability, antioxidants, and antiradical potential. LWT Food Sci. Technol. 2015, 60, 22–28. [Google Scholar] [CrossRef]
- Khemakhem, I.; Yaiche, C.; Ayadi, M.A.; Bouaziz, M. Impact of aromatization by Citrus limetta and Citrus sinensis peels on olive oil quality, chemical composition and heat stability. J. Am. Oil Chem. Soc. 2015, 92, 701–708. [Google Scholar] [CrossRef]
- Caponio, F.; Durante, V.; Varva, G.; Silletti, R.; Previtali, M.A.; Viggiani, I.; Squeo, G.; Summo, C.; Pasqualone, A.; Gomes, T.; Baiano, A. Effect of infusion of spices into the oil vs. combined malaxation of olive paste and spices on quality of naturally flavoured virgin olive oils. Food Chem. 2016, 202, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Assami, K.; Chemat, S.; Meklati, B.Y.; Chemat, F. Ultrasound-assisted aromatisation with condiments as an enabling technique for olive oil flavouring and shelf life enhancement. Food Anal. Methods 2016, 9, 982–990. [Google Scholar] [CrossRef]
- Clodoveo, M.L.; Dipalmo, T.; Crupi, P.; Durante, V.; Pesce, V.; Maiellaro, I.; Lovece, A.; Mercurio, A.; Laghezza, A.; Corbo, F.; et al. Comparison between different flavored olive oil production techniques: Healthy value and process efficiency. Plant Foods Hum. Nutr. 2016, 71, 81–87. [Google Scholar] [CrossRef] [PubMed]
- González-Gamallo, S.; Gómez-Alonso, S.; Salvador, M.D.; Fregapane, G. Sensory profile of virgin olive oils co-processed with citrus fruits Poster P2-113. In Proceedings of the 4th European Conference on Sensory and Consumer Research—A Sense of Quality, Vitoria-Gasteiz, Spain, 5–8 September 2010.
- Salvador, M.D.; González-Gamallo, S.; Gómez-Alonso, S.; Fregapane, G. Phenolic compounds of virgin olive oils co-processed with citrus fruits Poster OO-007 (Book of Abstracts page 281). In Proceedings of the 9th Euro Fed Lipid Congress, Rotterdam, The Netherlands, 18–21 September 2011.
- González-Gamallo, S.; Gómez-Alonso, S.; Fregapane, G.; Salvador, M.D. New virgin olive oil based products using fruits and spices. Poster P2-147. In Proceedings of the 4th European Conference on Sensory and Consumer Research—A Sense of Quality, Vitoria-Gasteiz, Spain, 5–8 September 2010.
- Eduardo, S.R. Method of Preparing Lycopene-Enriched Formulations That Are Free of Organic Solvents, Formulations thus Obtained, Compositions Comprising Said Formulations and Use of Same. WO Patent 2006/111591, 26 October 2006. [Google Scholar]
- Bendini, A.; Di Lecce, G.; Valli, E.; Barbieri, S.; Tesini, F.; Toschi, T.G. Olive oil enriched in lycopene from tomato by-product through a co-milling process. Int. J. Food Sci. Nutr. 2015, 66, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Frankel, E.; Bakhouche, A.; Lozano-Sánchez, J.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Literature review on production process to obtain extra virgin olive oil enriched in bioactive compounds. Potential use of byproducts as alternative sources of polyphenols. J. Agric. Food Chem. 2013, 61, 5179–5188. [Google Scholar] [CrossRef] [PubMed]
- Moldao-Martins, M.; Beirao-da-Costa, S.; Neves, C.; Cavaleiro, C.; Salgueiro, L.; Beirao-da-Costa, M.L. Olive oil flavoured by the essential oils of Mentha × piperita and Thymus mastichina L. Food Qual. Prefer. 2004, 15, 447–452. [Google Scholar] [CrossRef]
- Aidoud, A.; Ammouche, A.; Garrido, M.; Rodriguez, A.M. Effect of lycopene-enriched olive and argan oils upon lipid serum parameters in Wistar rats. J. Sci. Food Agric. 2014, 94, 2943–2950. [Google Scholar] [CrossRef] [PubMed]
- Zunin, P.; Leardi, R.; Bisio, A.; Boggia, R.; Romussi, G. Oxidative stability of virgin olive oil enriched with carnosic acid. Food Res. Int. 2010, 43, 1511–1516. [Google Scholar] [CrossRef]
- Koprivnjak, O.; Škevin, D.; Valic, S.; Majetic, V.; Petricevic, S.; Ljubenkov, I. The antioxidant capacity and oxidative stability of virgin olive oil enriched with phospholipids. Food Chem. 2008, 111, 121–126. [Google Scholar] [CrossRef]
- Nunes, P.A.; Pires-Cabral, P.; Ferreira-Dias, S. Production of olive oil enriched with medium chain fatty acids catalysed by commercial immobilised lipases. Food Chem. 2011, 127, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Rico, A.; Fregapane, G.; Salvador, M.D. Effect of cultivar and ripening on minor components in Spanish olive fruits and their corresponding virgin olive oils. Food Res. Int. 2008, 41, 433–440. [Google Scholar] [CrossRef]
- Gómez-Rico, A.; Inarejos-Garcia, A.M.; Salvador, M.D.; Fregapane, G. Effect of malaxation conditions on phenol and volatile profiles in olive paste and the corresponding virgin olive oils (Olea europaea L. Cv. cornicabra). J. Agric. Food Chem. 2009, 57, 3587–3595. [Google Scholar] [CrossRef] [PubMed]
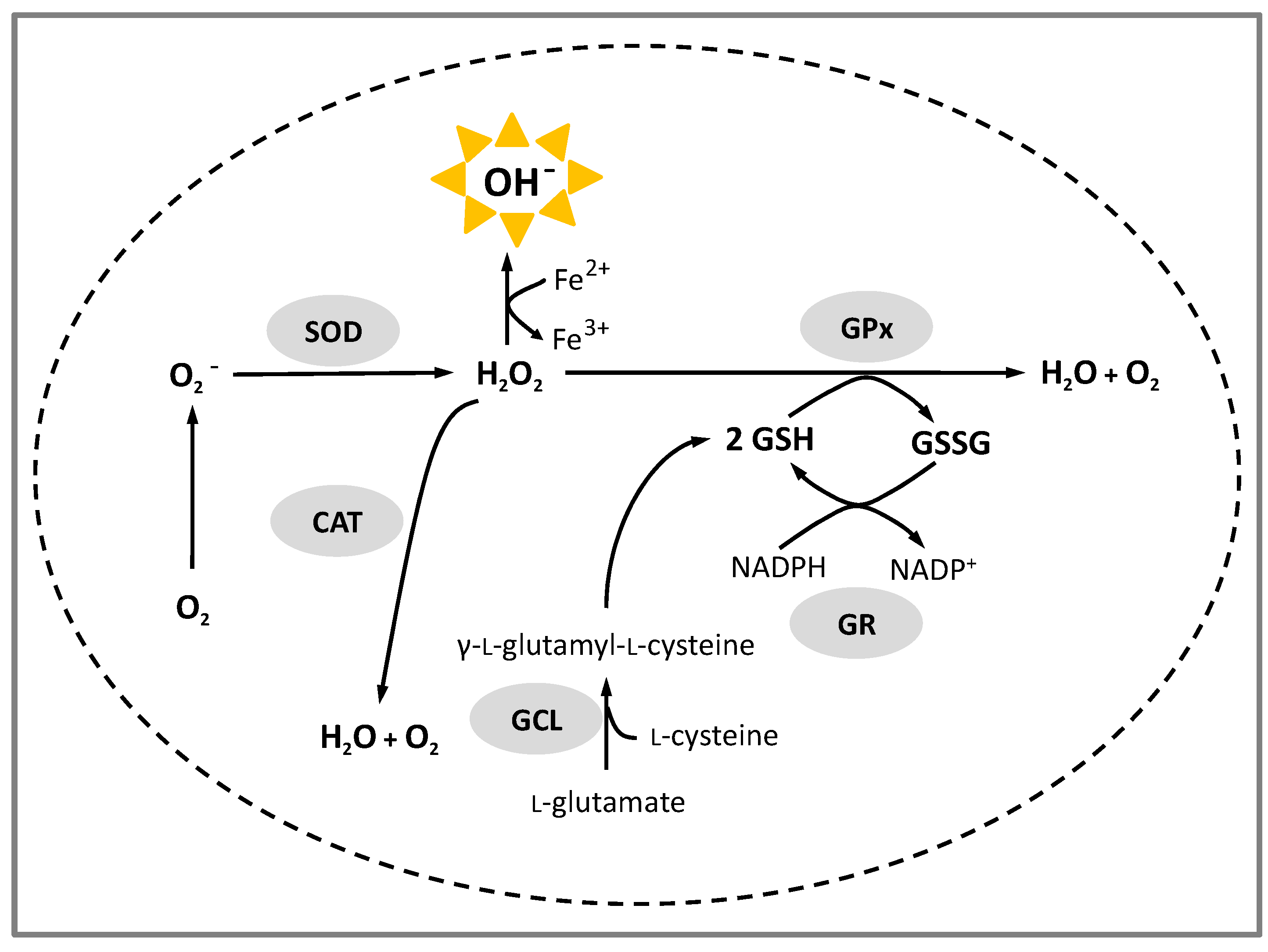
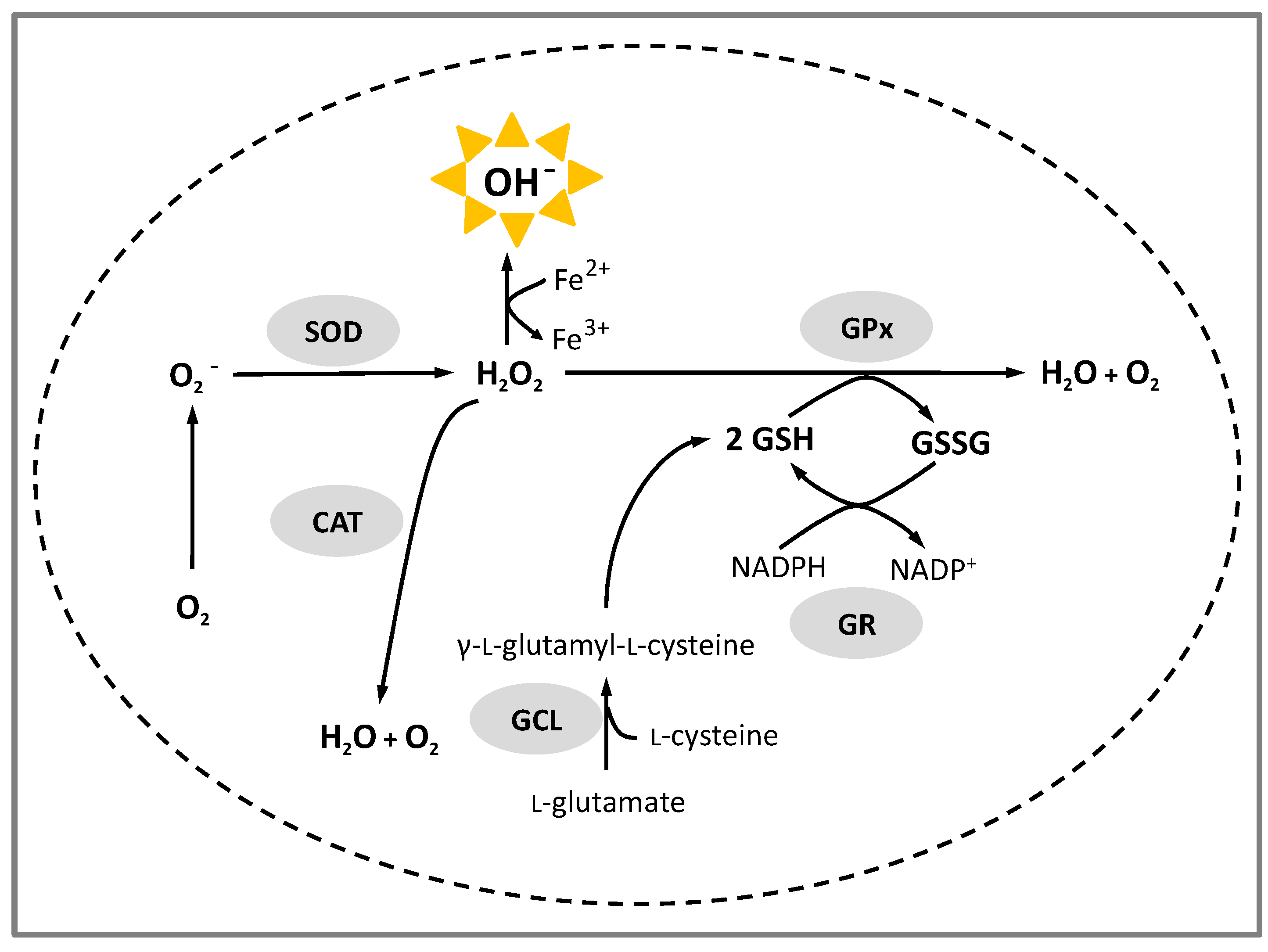
- Espinosa-Díez, C.; Miguel, V.; Mennerich, D.; Kietzmann, T.; Sánchez-Pérez, P.; Cadenas, S.; Lamas, S. Antioxidant responses and cellular adjustments to oxidative stress. Redox Biol. 2015, 6, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signalling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J. Chapter: Transcriptional regulation of cellular antioxidant defense mechanisms. In Oxidative Stress, Inflammation and Heath Book; Surh, Y.J., Packer, L., Eds.; Taylor and Francis Group: Boca Raton, FL, USA, 2005. [Google Scholar]
- Forman, H.J.; Davies, K.J.A.; Ursini, F. How Do Nutritional Antioxidants Really Work: Nucleophilic Tone and Para-Hormesis Versus Free Radical Scavenging in vivo. Free Radic. Biol. Med. 2014, 66, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Farràs, M.; Castañer, O.; Martín-Peláez, S.; Hernáez, A.; Schröder, H.; Subirana, I.; Muñoz-Aguayo, D.; Gaixas, S.; Torre, R.D.L.; Farré, M.; et al. Complementary phenol-enriched olive oil improves HDL characteristics in hypercholesterolemic subjects. A randomized, double-blind, crossover, controlled trial. The VOHF study. Mol. Nutr. Food Res. 2015, 59, 1758–1770. [Google Scholar] [CrossRef] [PubMed]
- Rubió, L.; Macià, A.; Castell-Auví, A.; Pinent, M.; Blay, M.T.; Ardévol, A.; Romero, M.-P.; Motilva, M.-J. Effect of the co-occurring olive oil and thyme extracts on the phenolic bioaccesibility and bioavailability assessed by in vitro digestion and cell models. Food Chem. 2014, 149, 277–284. [Google Scholar] [CrossRef] [PubMed]
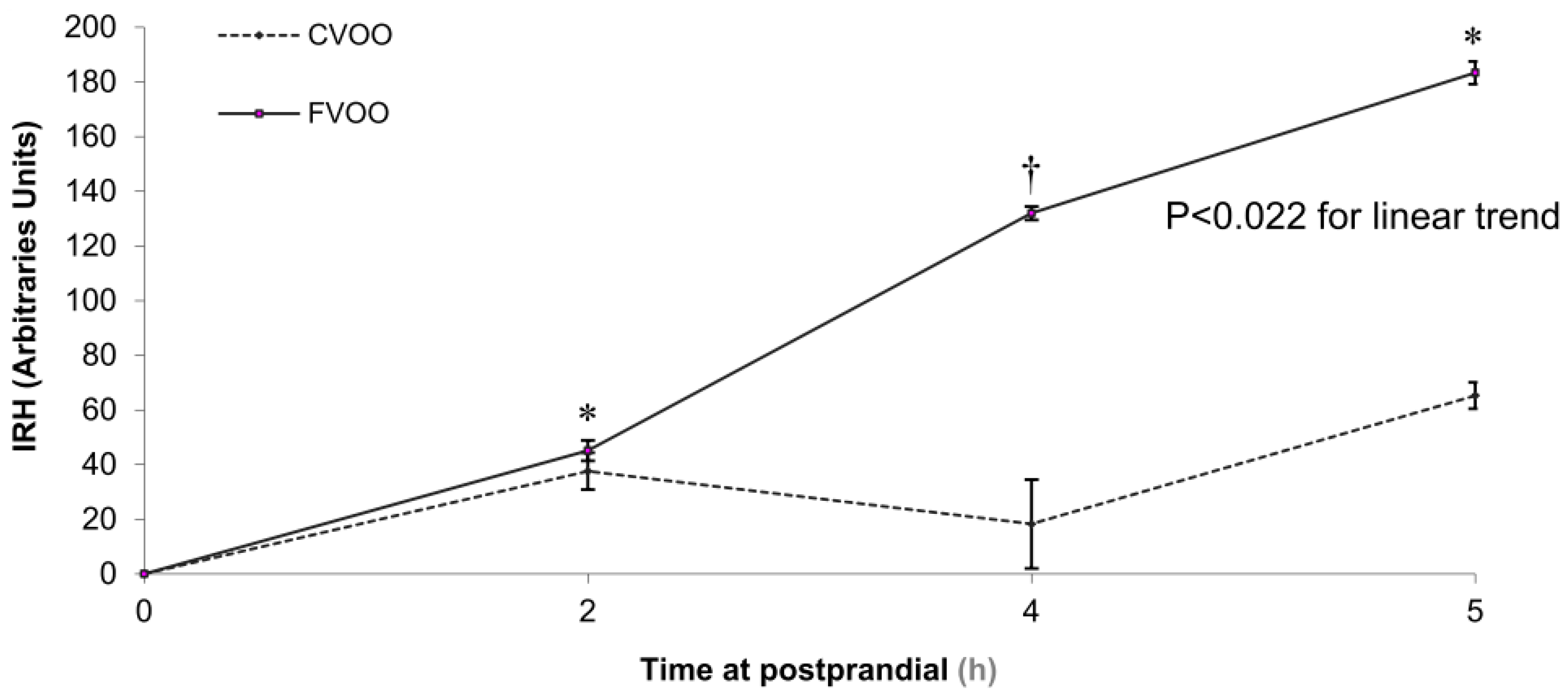
- Valls, R.-M.; Farràs, M.; Suárez, M.; Fernández-Castillejo, S.; Fitó, M.; Konstantinidou, V.; Fuentes, F.; López-Miranda, J.; Giralt, M.; Covas, M.-I.; et al. Effects of functional olive oil enriched with its own phenolic compounds on endothelial function in hypertensive patients. A randomised controlled trial. Food Chem. 2015, 167, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Fernández, J.M.; Fuentes-Jiménez, F.; López-Miranda, J. Función endotelial y ejercicio físico. RAMD 2009, 2, 61–69. Available online: http://www.redalyc.org/articulo.oa?id=323327658005 (accessed on 10 October 2016). [Google Scholar]
- Landmesser, U.; Drexler, H. Endothelial function and hypertension. Curr. Opin. Cardiol. 2007, 22, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Harrison, D.G. Endothelial dysfunction in cardiovascular diseases: The role of oxidant stress. Circ. Res. 2000, 87, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Kojda, G.; Harrison, D. Interactions between NO and reactive oxygen species: Pathophysiological importance in atherosclerosis, hypertension, diabetes and heart failure. Cardiovasc. Res. 1999, 43, 562–571. [Google Scholar] [CrossRef]
- Barter, P. The role of HDL-cholesterol in preventing atherosclerotic disease. Eur. Heart J. Suppl. 2005, 7 (Suppl. F), F4–F8. [Google Scholar] [CrossRef]
- Hadi, H.A.; Carr, C.S.; Al Suwaidi, J. Endothelial Dysfunction: Cardiovascular Risk Factors, Therapy, and Outcome. Vasc. Health Risk Manag. 2005, 1, 183–198. [Google Scholar] [PubMed]
- Meurman, J.H.; Sanz, M.; Janket, S.-J. Oral health, atherosclerosis, and cardiovascular disease. Crit. Rev. Oral. Biol. Med. 2004, 15, 403–413. [Google Scholar] [CrossRef] [PubMed]
- López de las Hazas, M.-C.; Piñol, C.; Macià, A.; Romero, M.-P.; Pedret, A.; Solà, R.; Rubió, L.; Motilva, M.-J. Differential absorption and metabolism of hydroxytyrosol and its precursors oleuropein and secoiridoids. J. Funct. Foods 2016, 22, 52–63. [Google Scholar] [CrossRef]
- Arts, E.; Fransen, J.; Lemmers, H.; Stalenhoef, A.; Joosten, L.; van Riel, P.; Popa, C.D. High-density lipoprotein cholesterol subfractions HDL2 and HDL3 are reduced in women with rheumatoid arthritis and may augment the cardiovascular risk of women with RA: A cross-sectional study. Arthritis Res. Ther. 2012, 14, R116. [Google Scholar] [CrossRef] [PubMed]
- Cooney, M.T.; Dudina, A.; De Bacquer, D.; Wilhelmsen, L.; Sans, S.; Menotti, A.; De Backer, G.; Jousilahti, P.; Keil, U.; Thomsen, T.; et al. HDL cholesterol protects against cardiovascular disease in both genders, at all ages and at all levels of risk. Atherosclerosis 2009, 206, 611–616. [Google Scholar] [CrossRef] [PubMed]
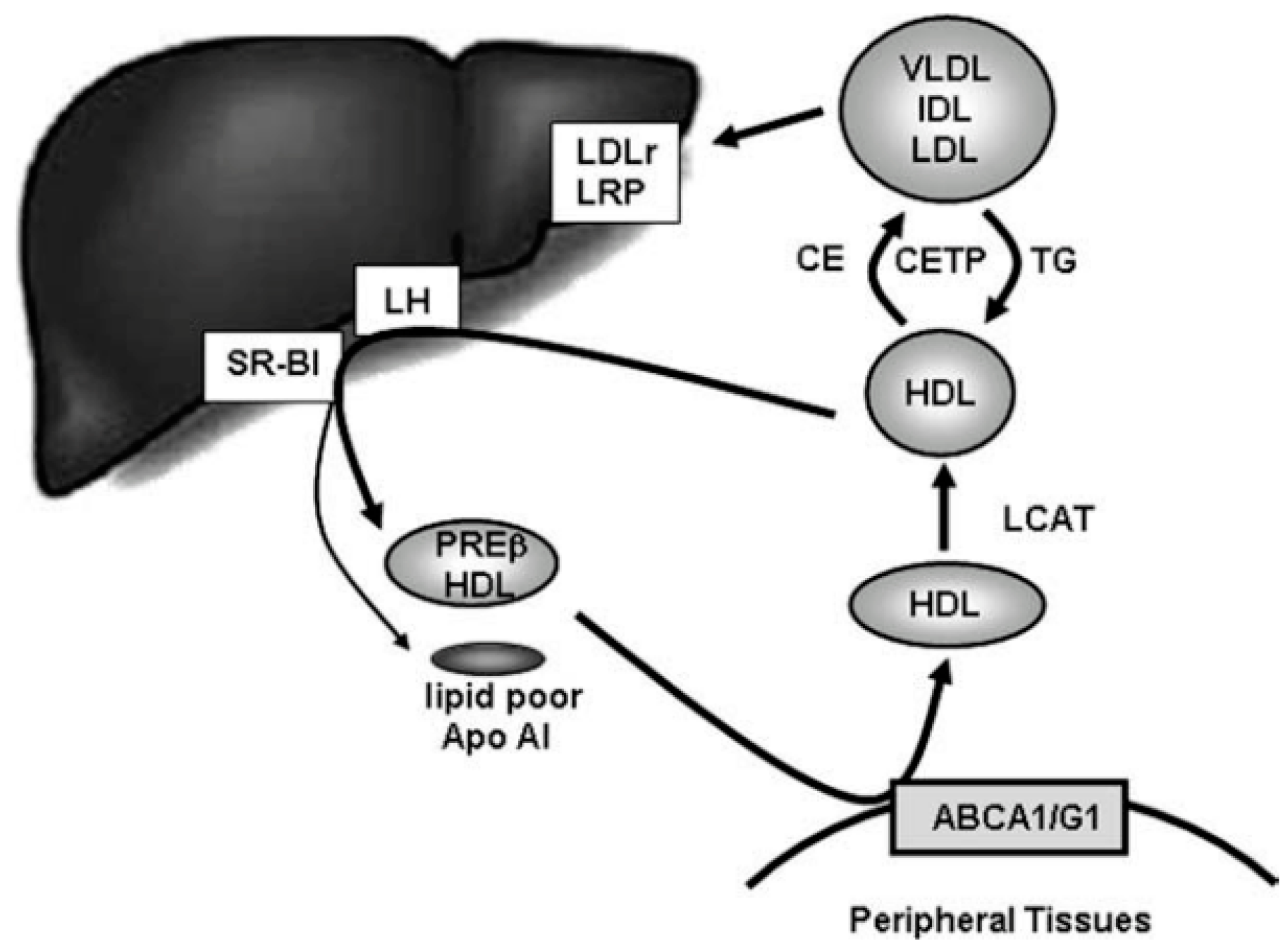
- Rothblat, G.H.; Phillips, M.C. High-density lipoprotein heterogeneity and function in reverse cholesterol transport. Curr. Opin. Lipidol. 2010, 21, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Pekkanen, J.; Linn, S.; Heiss, G.; Suchindran, C.M.; Leon, A.; Rifkind, B.M.; Tyroler, H.A. Ten-year mortality from cardiovascular disease in relation to cholesterol level among men with and without preexisting cardiovascular disease. N. Eng. J. Med. 1990, 322, 1700–1707. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.C.F.; De Faria, E.C. Cholesteryl ester transfer protein: The controversial relation to atheroclerosis and emerging new biological roles. IUBMB Life 2011, 63, 248–257. [Google Scholar] [CrossRef] [PubMed]
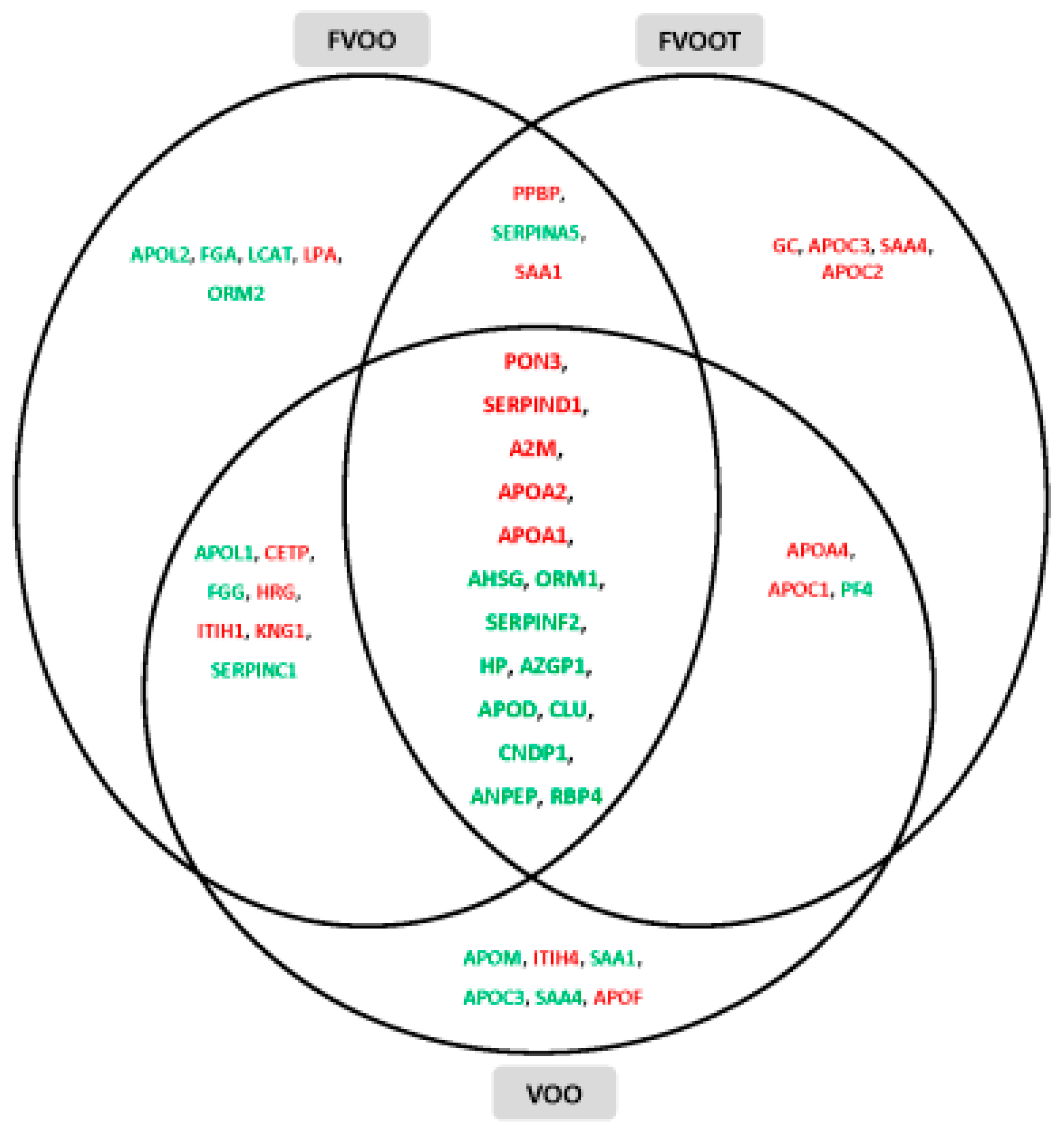
- Pedret, A.; Catalán, Ú.; Fernández-Castillejo, S.; Farràs, M.; Valls, R.-M.; Rubió, L.; Canela, N.; Aragonés, G.; Romeu, M.; Castañer, O.; et al. Impact of virgin olive oil and phenol-enriched virgin olive oils on the HDL proteome in hypercholesterolemic subjects: A double blind, randomized, controlled, cross-over clinical trial (VOHF study). PLoS ONE 2015, 10, e0129160. [Google Scholar] [CrossRef] [PubMed]
- Littman, D.R.; Pamer, E.G. Role of the commensal microbiota in normal and pathogenic host immune responses. Cell Host Microbe 2011, 10, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Borruel, N. Interacciones bacterianas con el sistema inmunológico intestinal: Inmunomodulación. Gastroenterol. Hepatol. 2003, 26 (Suppl. S1), 13–22. [Google Scholar] [CrossRef]
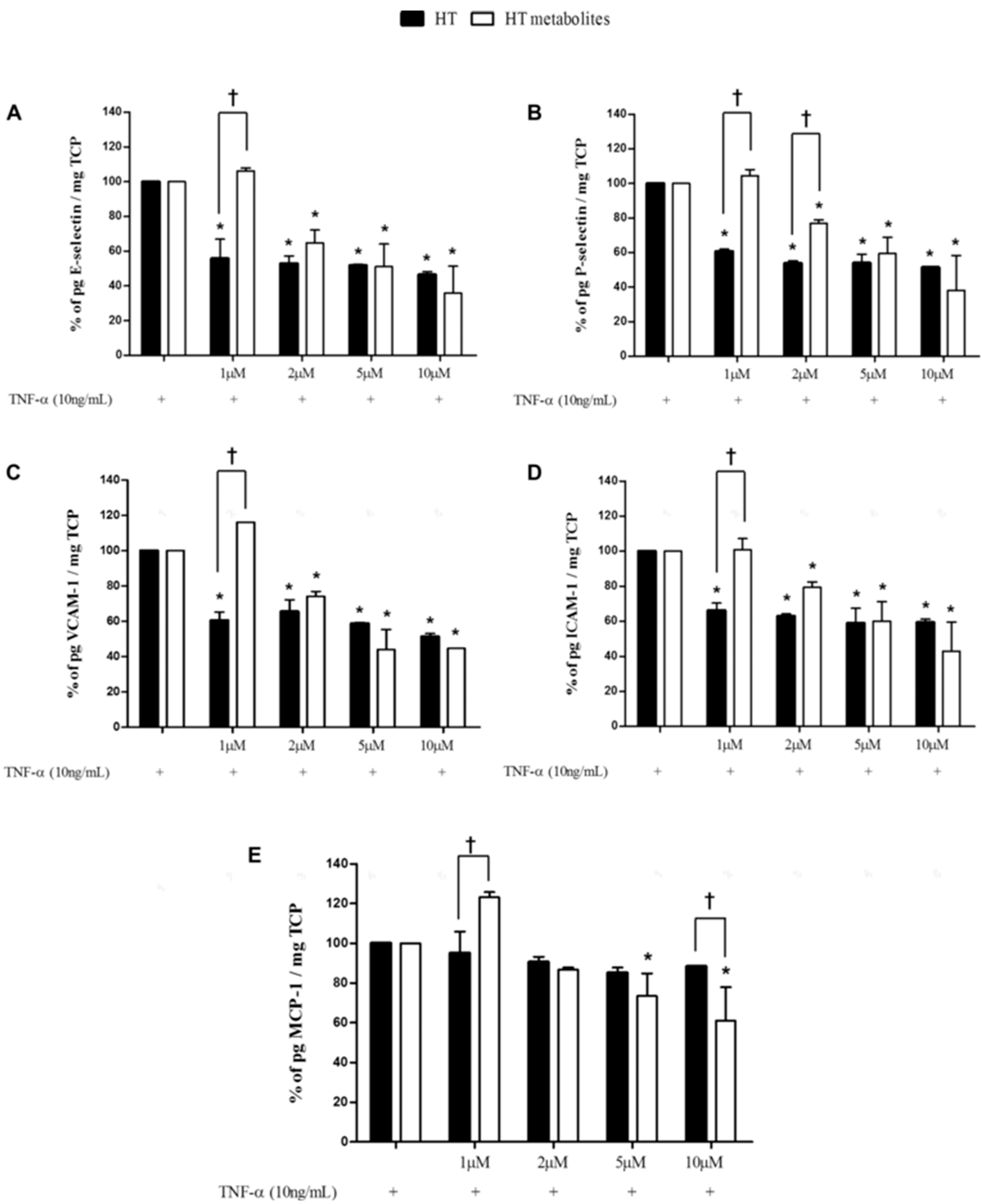
- Martín-Peláez, S.; Castañer, O.; Solà, R.; Motilva, M.J.; Castell, M.; Pérez-Cano, F.J.; Fitó, M. Influence of phenol-enriched olive oils on human intestinal immune function. Nutrients 2016, 8, 213. [Google Scholar] [CrossRef] [PubMed]
- Vollaard, E.J.; Clasener, H.A.L. Colonization resistance. Antimicrob. Agents Chemother. 1994, 38, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Fitó, M.; Cladellas, M.; de la Torre, R.; Marti, J.; Munoz, D.; Schroder, H.; Alcantara, M.; Pujadas-Bastardes, M.; Marrugat, J.; Lopez-Sabater, M.C.; et al. Anti-inflammatory effect of virgin olive oil in stable coronary disease patients: A randomized, crossover, controlled trial. Eur. J. Clin. Nutr. 2008, 62, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Walnutolive. Available online: http://www.walnutolive.com/index.html (accessed on 11 March 2017).
- iO Antioxidante Natural. Available online: http://www.ioantioxidantenatural.es/io-antioxidante-natural/ (accessed on 11 March 2017).
- AOVE Omega 3. Available online: http://www.laespanolaaceites.com/es/aceite-de-oliva-virgen-extra-omega-3/ (accessed on 11 March 2017).
- Un Aceite de Oliva con Sabor a Mar. Available online: http://espana.gastronomia.com/noticia/3175/un-aceite-de-oliva-con-sabor-a-mar (accessed on 11 March 2017).
- Limón, P.; Malheiro, R.; Casal, S.; Acién-Fernández, F.G.; Fernández-Sevilla, J.M.; Rodriges, N.; Cruz, R.; Bermejo, R.; Pereira, J.A. Improvement of stability and carotenoids fraction of virgin olive oils by addition of microalgae Scenedesmus almeriensis extracts. Food Chem. 2015, 175, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Kussmann, M.; Van Bladeren, P.J. The extended nutrigenomics—Understanding the interplay between the genomes of food, gut microbes, and human host. Front. Genet. 2011, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R. Nutrigenomics and Nutrigenetics in Functional Foods and Personal Nutrition; Ferguson, L.R., Ed.; Taylor and Francis Group: Boca Raton, FL, USA, 2013. [Google Scholar]
- Constantin, N.; Wahli, W. Nutrigenomic foods. What will be eating tomorrow? Nutrafoods 2013, 12, 3–12. [Google Scholar] [CrossRef]
- Giordano, E.; Dávalos, A.; Visioli, F. Chronic hydroxytyrosol feeding modulates glutathione-mediated oxido-reduction pathways in adipose tissue: A nutrigenomic study. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1144–1150. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source/Ingredient | Extraction Method and Conditions | Observed Functional Effect in Enriched Olive Oil | Reference |
|---|---|---|---|
| Olive leaves | SLE: directly in the oil (organic-solvent free), dynamic extraction chamber, USAE (25 °C, 20 min) | Increased oxidative stability | [40] |
| SLE and LLE with ethanol, MAE (8–10 min, 200–400 W) | Increased oxidative stability | [41,42,43] | |
| Olive pomace | SLE: ethanol/water (70:30), overnight stirring | Increased oxidative stability | [44] |
| SLE: methanol and 1,2-propanediol, hydrolyzate (HCl 2 M, 100 °C for 1 h) | Increased phenolic content in fried foodstuffs | [45] | |
| LLE with ethyl acetate (for vegetative water) and SLE with ethanol/water (80:20; for solid residue) | Increased oxidative stability and antioxidant capacity | [38] | |
| ASE with ethanol/water (80:20 at 80 °C) | Increased oxidative stability and antioxidant capacity | [42] | |
| ASE with ethanol/water (80:20 at 80 °C), enrichment with 0.3% of emulsifier (lecithin or monoglycerides) and USAE | Lecithin is more effective increasing oxidative stability and antioxidant capacity | [47] | |
| Olive leaf and pomace | Aqueous extracts with or without the use of lecithin | Increased oxidative stability | [48] |
| Olive oil | Semi-preparative HPLC | Increased antioxidant capacity | [49] |
| Technological conditions during crushing (milling intensity) and malaxation (temperature and time) | Increased oxidative stability and antioxidant capacity | [50] | |
| Red pepper | Infusion: 10%–20% up to 30 days | Reduced oxidative stability | [51] |
| SFE at 40 °C and 15–23 MPa | Reduced oxidative stability | [52] | |
| Hot pepper, garlic, oregano and rosemary | Infusion: 20–40 g/L up to 7 months | Increased oxidative stability | [53] |
| Rosemary, lavender, sage, menthe, basil, lemon and thyme | Infusion: 5% for 15 days | Increased oxidative stability for rosemary > thyme and > lemon | [54] |
| Garlic, lemon, oregano, hot pepper, and rosemary | Co-processing (pressing, crushing and malaxation): 3%–20% into olives | Reduced stability, expect garlic Higher antioxidant activity (rosemary) | [55] |
| Basil | Infusion: 15% with USAE (1 W/cm2) for 15 min | Not determined | [56] |
| Lemon and thyme | Infusion: 20% during 2 months | Unchanged oxidative stability | [57] |
| Thyme and olive pomace | ASE: ethanol/water (80:20) at 80 °C and up to 1500 psi | Increased antioxidant capacity | [36] |
| Oregano | Addition of 0.05% of essential oil | Increased oxidative stability | [58] |
| Infusion by stirring at 1000 rpm for 3 h | Increased oxidative stability | [59] | |
| Thyme | Addition of 200 mg/L of essential oil | Not clear effect on oxidative stability | [60] |
| Garlic, hot chili peppers, laurel, oregano and pepper | Infusion: 10 g/L during three months at room temperature | Increased oxidative stability | [61] |
| Sweet lemon and sweet orange peels | Infusion: 1%–5% at 60 °C for 40 days | Increased antioxidant capacity. Reduced oxidative stability | [62] |
| Basil, chili and garlic | Infusion: 10%–20%, 7 days stirring at 15–18 °C vs. combined malaxation | Antioxidant activity was significantly lower in the oils obtained by infusion | [63] |
| Caraway | 1.5% at room temperature, infusion for 6 h vs. USAE for 30 min | Increased oxidative stability (OSI, but not for PV or Ks) | [64] |
| Thyme and oregano | Infusion (10 g/L for 15 days at room temperature) vs. co-malaxation (10 g/kg with or without 6 min USAE, before kneading) | Increased antioxidant capacity (especially with thyme) | [65] |
| Citrus fruits or peel | 0.5%–5% by co-processing (milling and/or malaxation) | Increased antioxidant capacity | [66,67] |
| Fruits (apple, lemon and orange), spices (rosemary, thyme, basil and oregano) and leaves (rocket) | 0.5%–5% by co-processing (milling and/or malaxation) | Statistically significant differences in sensory testing, except in apple and rocket | [68] |
| Lycopene | SLE with tomato pulp and VOO at high mechanical mixing (patented) | Enhanced antioxidant status in humans | [35,69] |
| Co-milling of tomato seed or skin and olives | Significant enrichment in carotenoids, especially in lycopene | [70] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reboredo-Rodríguez, P.; Figueiredo-González, M.; González-Barreiro, C.; Simal-Gándara, J.; Salvador, M.D.; Cancho-Grande, B.; Fregapane, G. State of the Art on Functional Virgin Olive Oils Enriched with Bioactive Compounds and Their Properties. Int. J. Mol. Sci. 2017, 18, 668. https://doi.org/10.3390/ijms18030668
Reboredo-Rodríguez P, Figueiredo-González M, González-Barreiro C, Simal-Gándara J, Salvador MD, Cancho-Grande B, Fregapane G. State of the Art on Functional Virgin Olive Oils Enriched with Bioactive Compounds and Their Properties. International Journal of Molecular Sciences. 2017; 18(3):668. https://doi.org/10.3390/ijms18030668
Chicago/Turabian StyleReboredo-Rodríguez, Patricia, María Figueiredo-González, Carmen González-Barreiro, Jesús Simal-Gándara, María Desamparados Salvador, Beatriz Cancho-Grande, and Giuseppe Fregapane. 2017. "State of the Art on Functional Virgin Olive Oils Enriched with Bioactive Compounds and Their Properties" International Journal of Molecular Sciences 18, no. 3: 668. https://doi.org/10.3390/ijms18030668
APA StyleReboredo-Rodríguez, P., Figueiredo-González, M., González-Barreiro, C., Simal-Gándara, J., Salvador, M. D., Cancho-Grande, B., & Fregapane, G. (2017). State of the Art on Functional Virgin Olive Oils Enriched with Bioactive Compounds and Their Properties. International Journal of Molecular Sciences, 18(3), 668. https://doi.org/10.3390/ijms18030668