
Comprehensive Analysis of Rice Laccase Gene (OsLAC) Family and Ectopic Expression of OsLAC10 Enhances Tolerance to Copper Stress in Arabidopsis

Abstract
:1. Introduction
2. Results
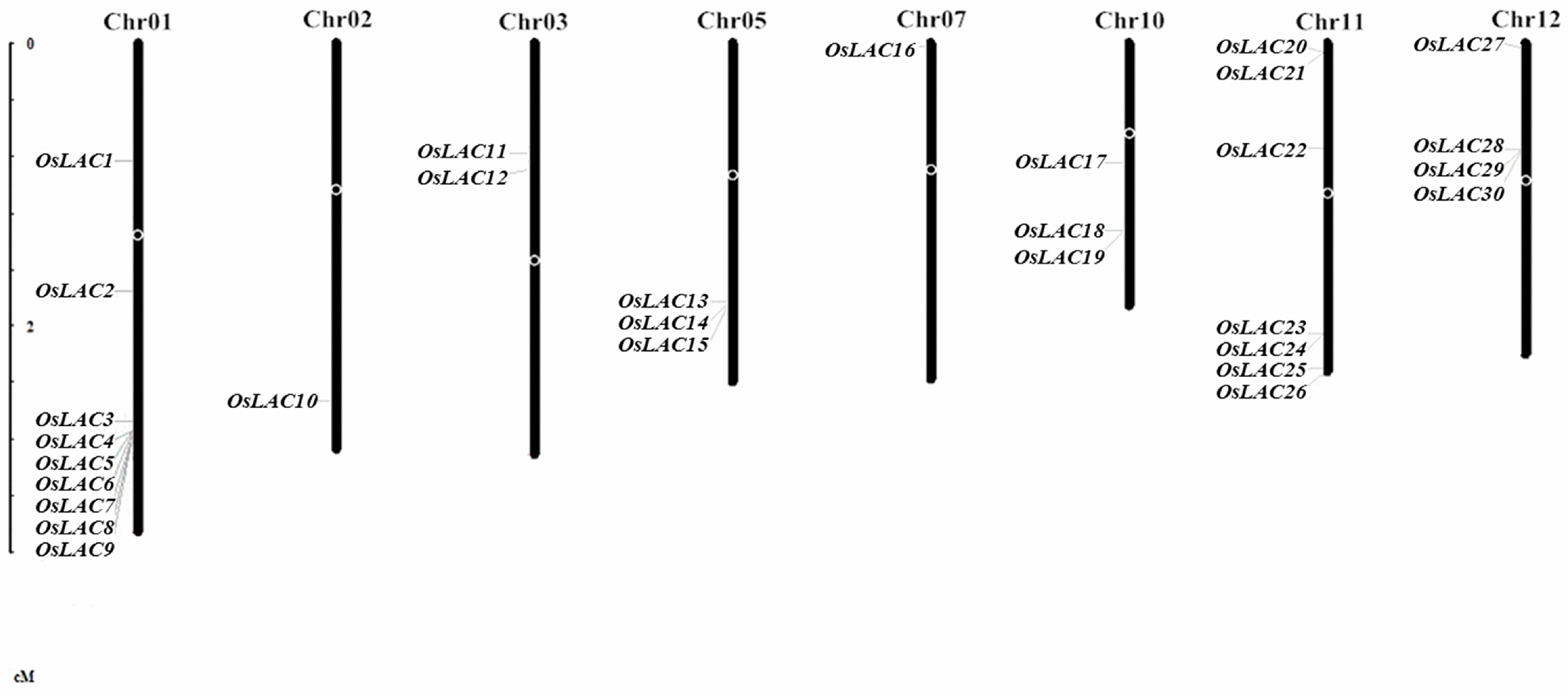
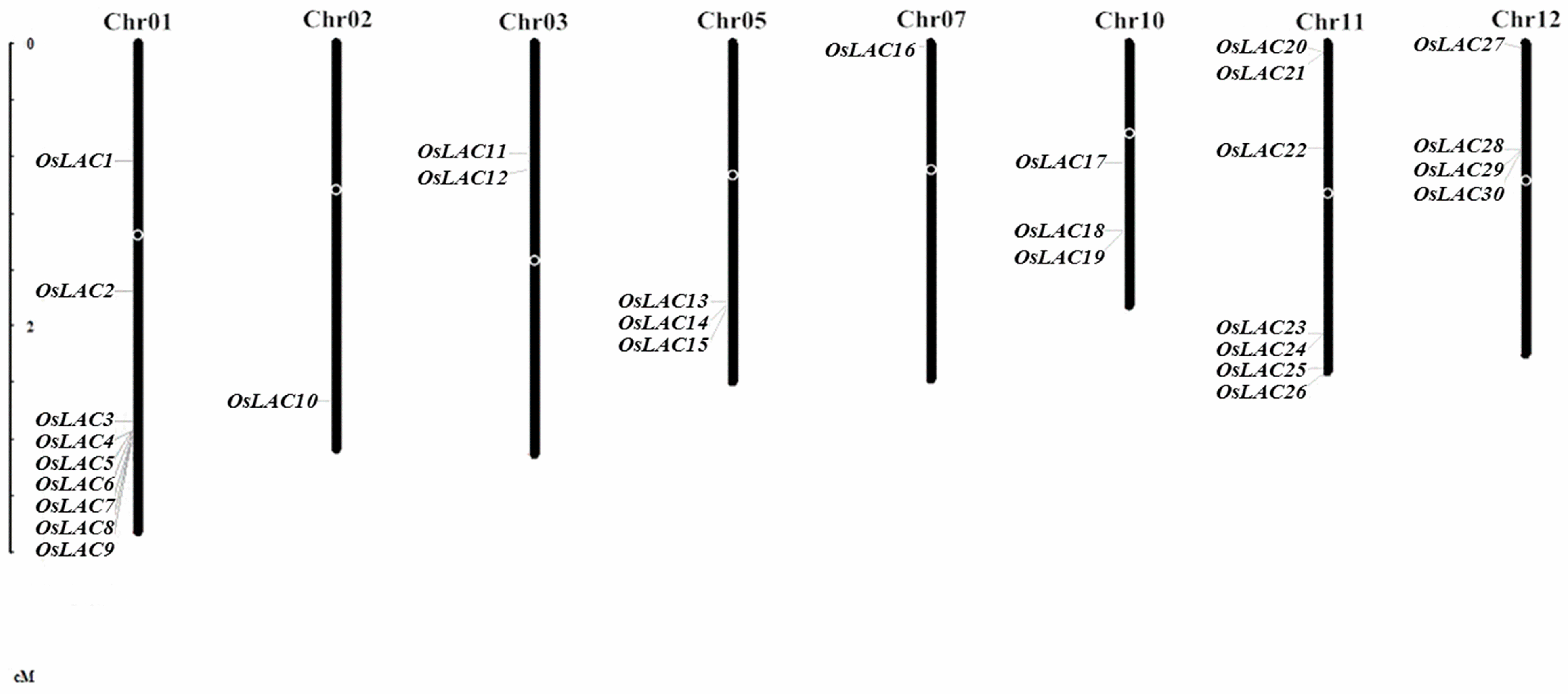
2.1. General Information on the Rice Laccase Gene Family
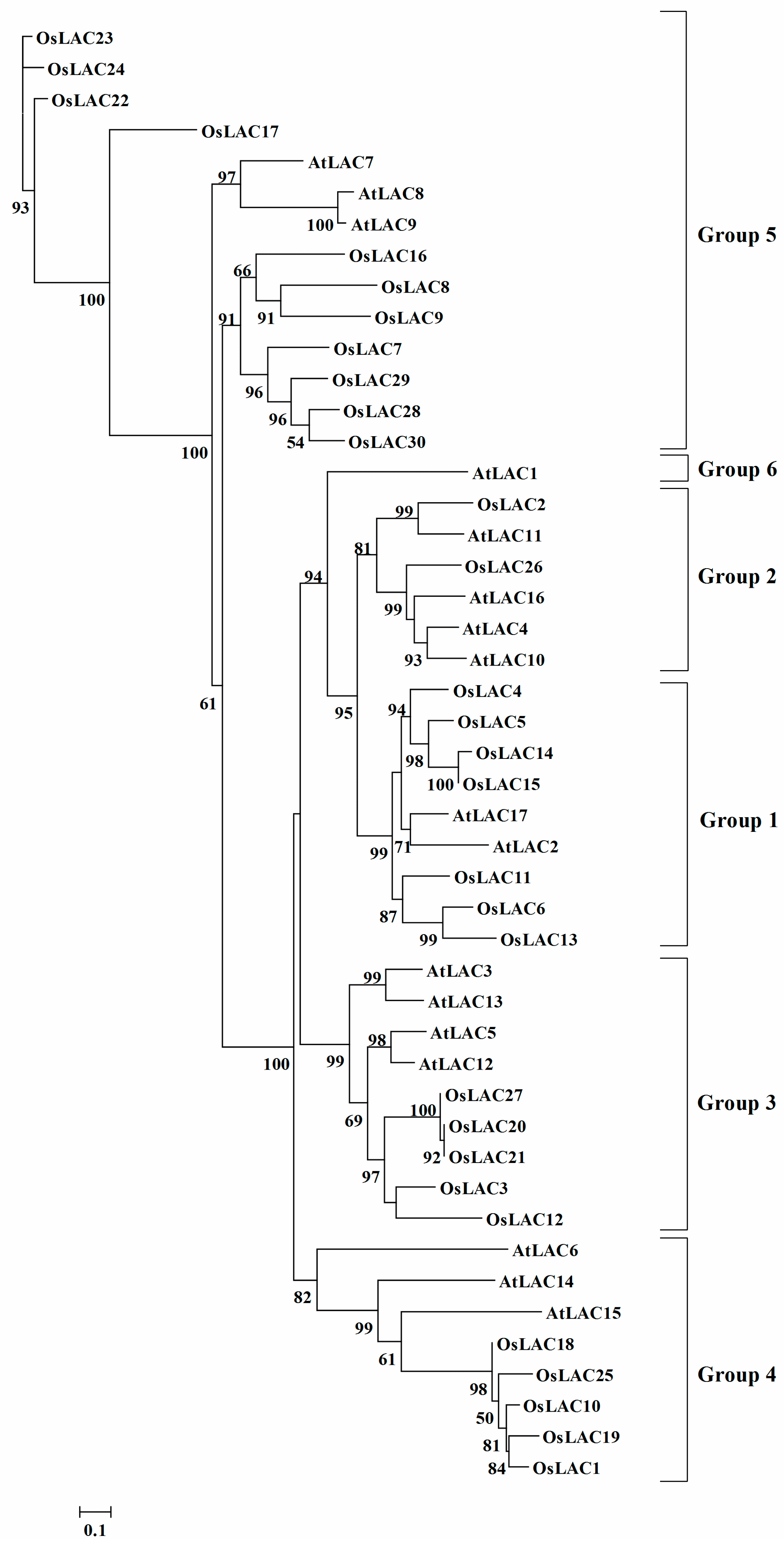
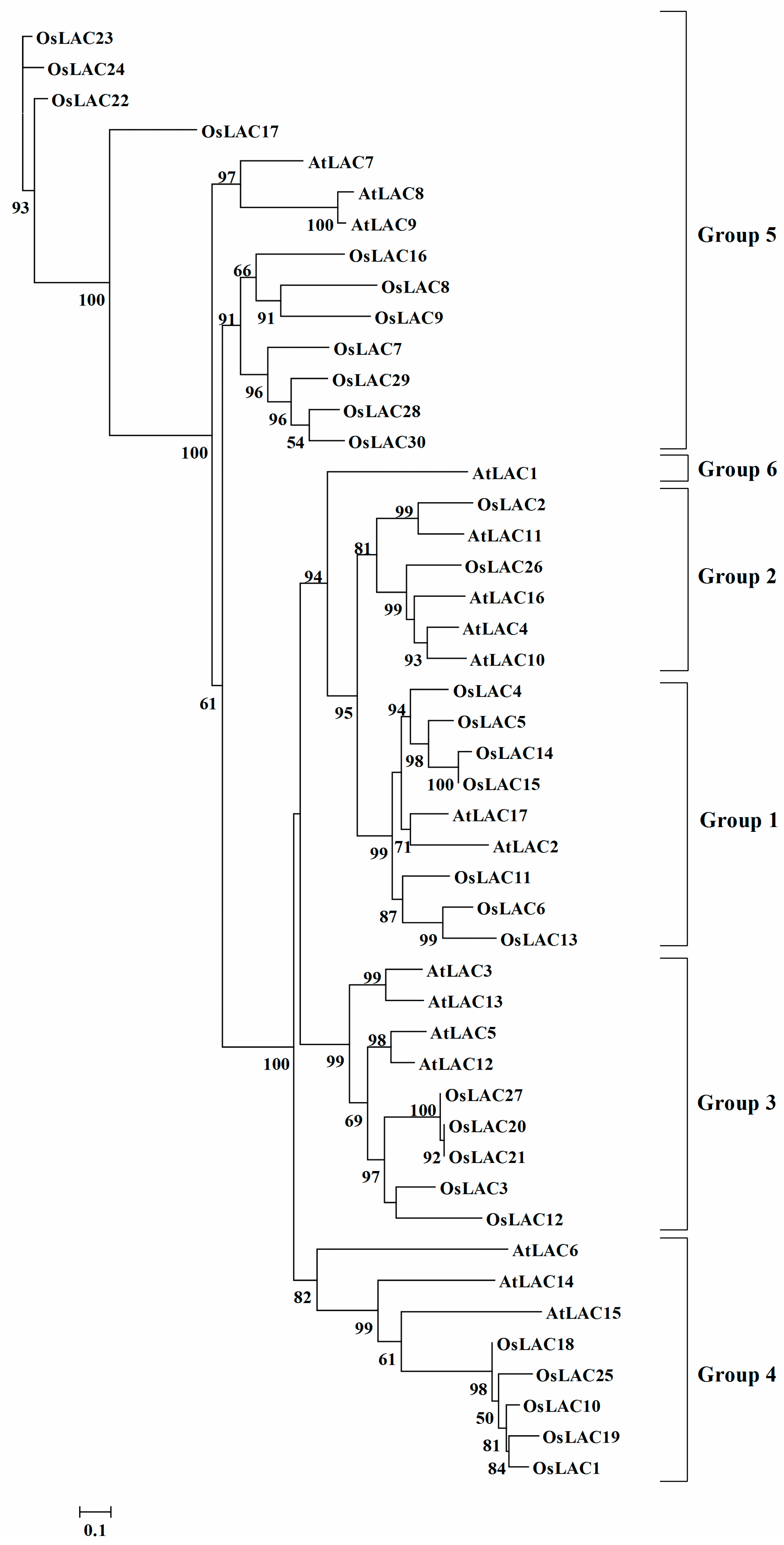
2.2. Phylogenetic Analysis of the Rice Laccase Family
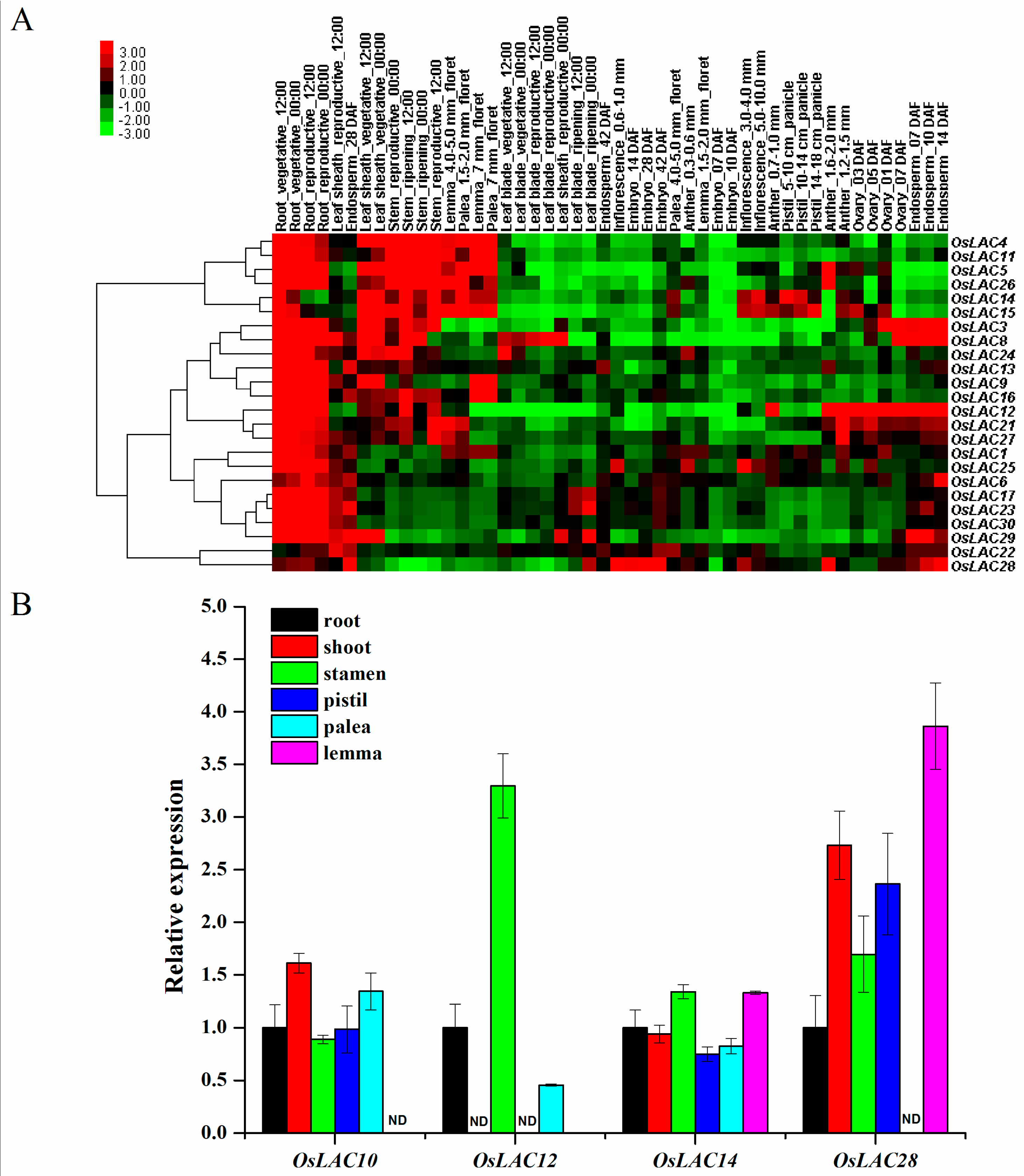
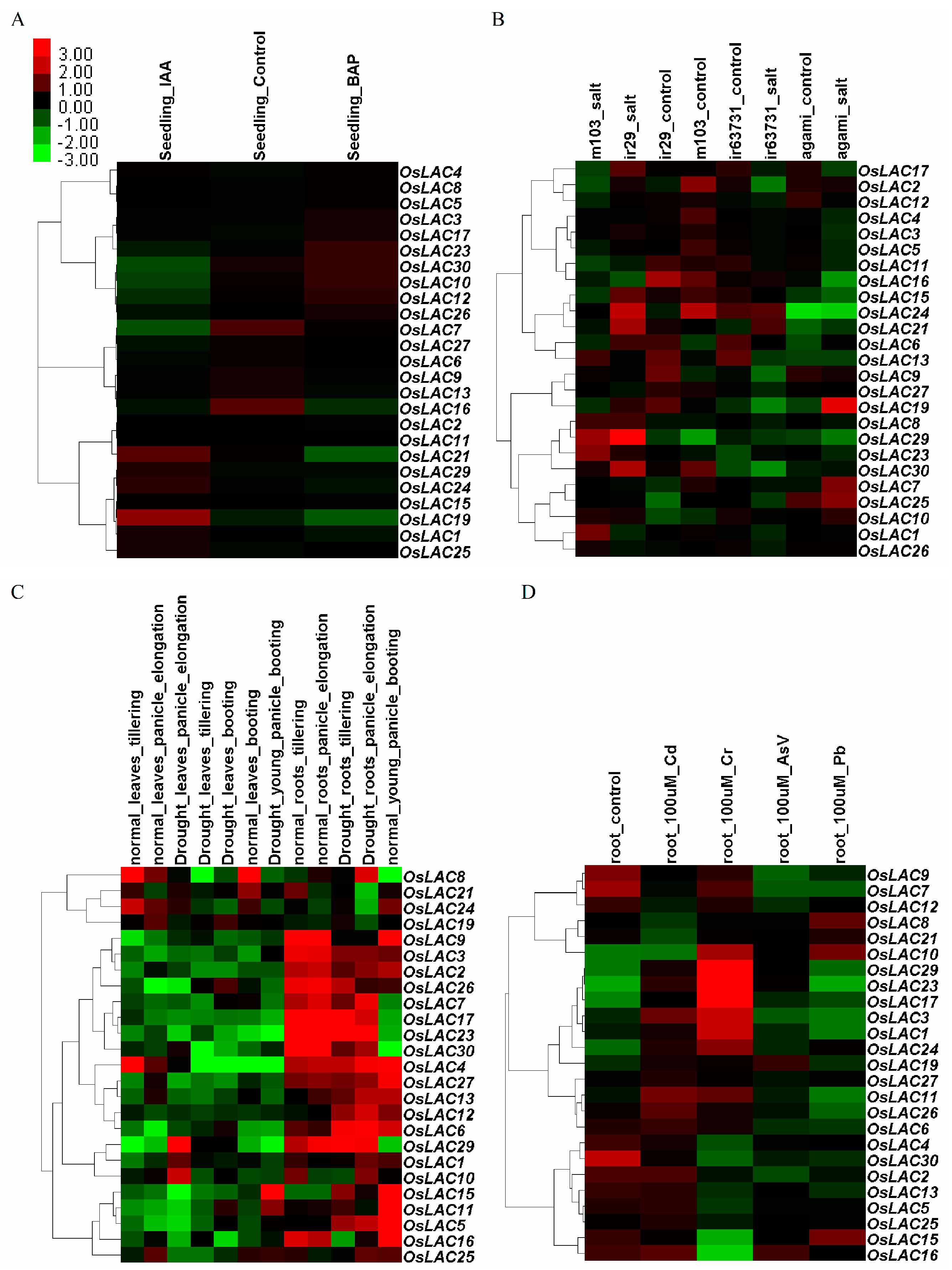
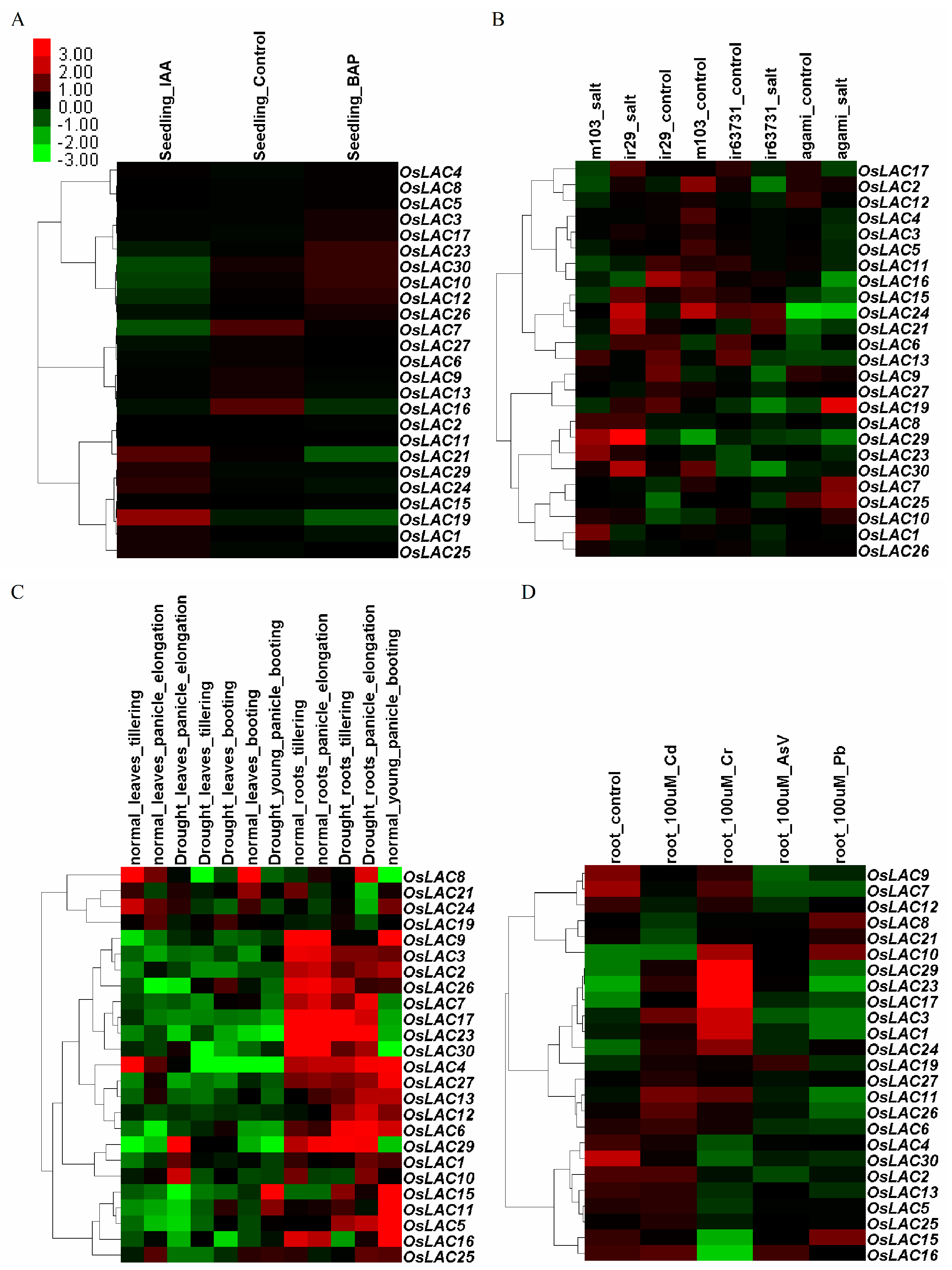
2.3. Spatiotemporal and Abiotic Stress-Inducible Expression Patterns of Rice Laccase Genes
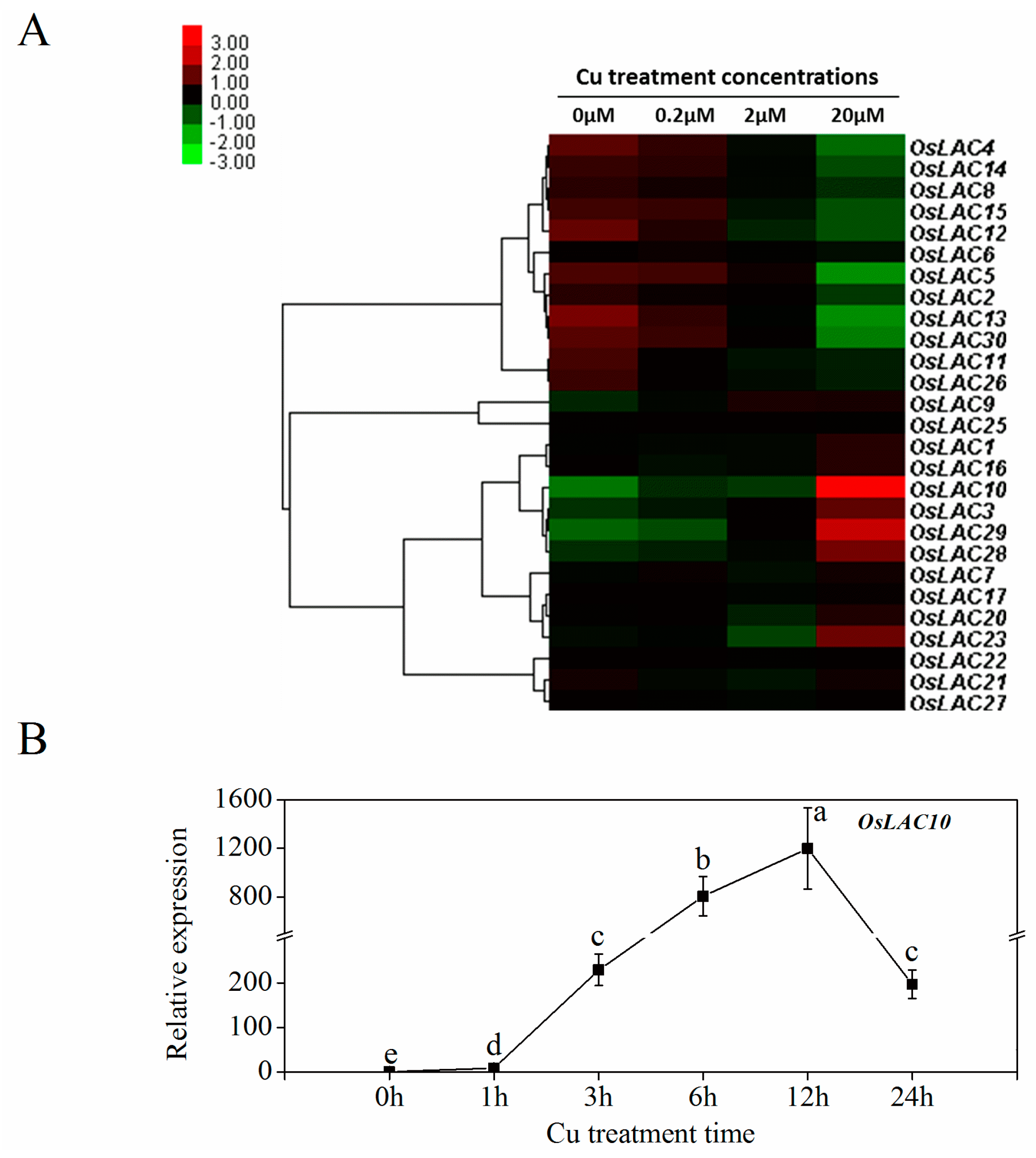
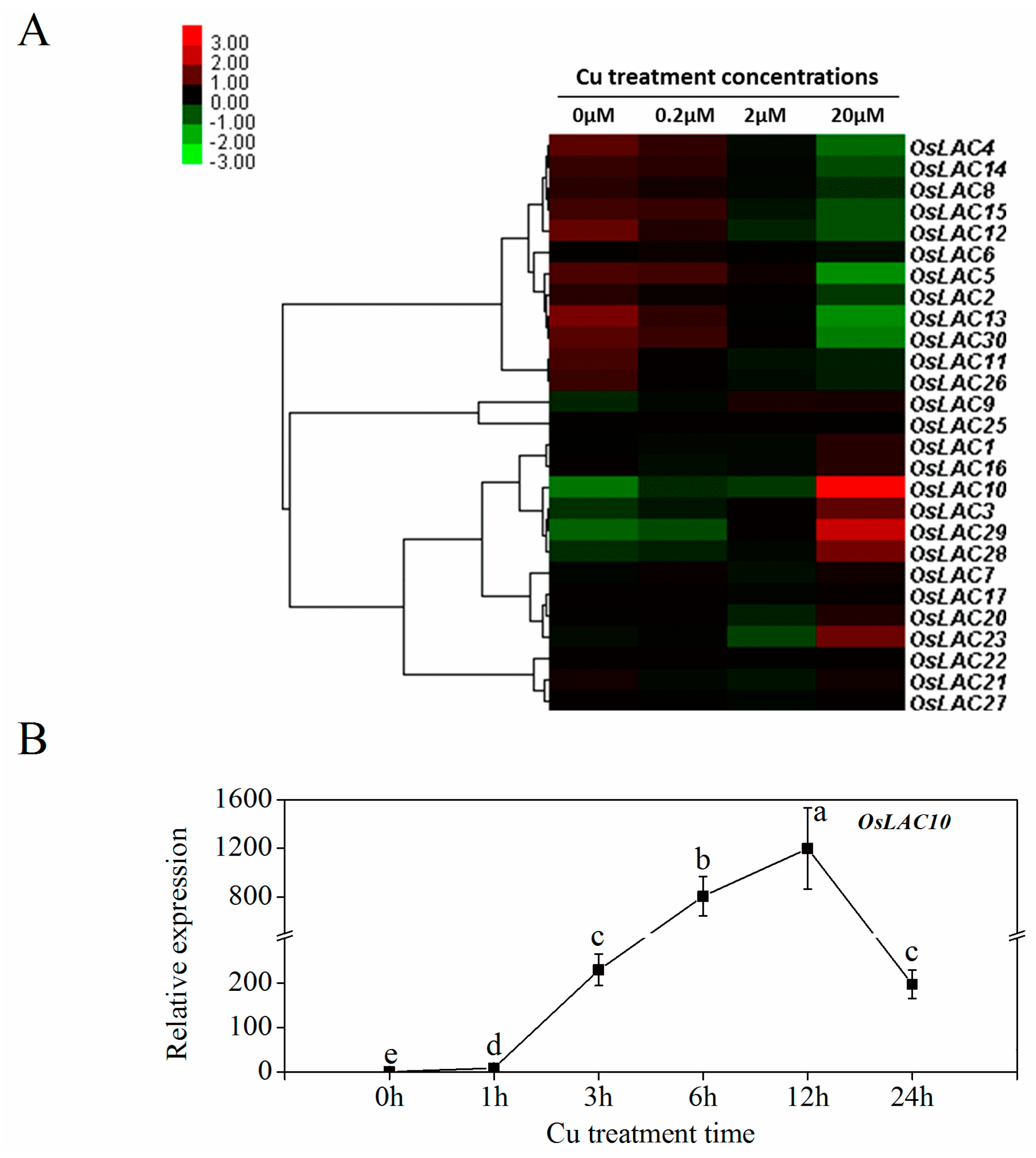
2.4. Cu-Inducible Expression Patterns of Rice Laccase Genes
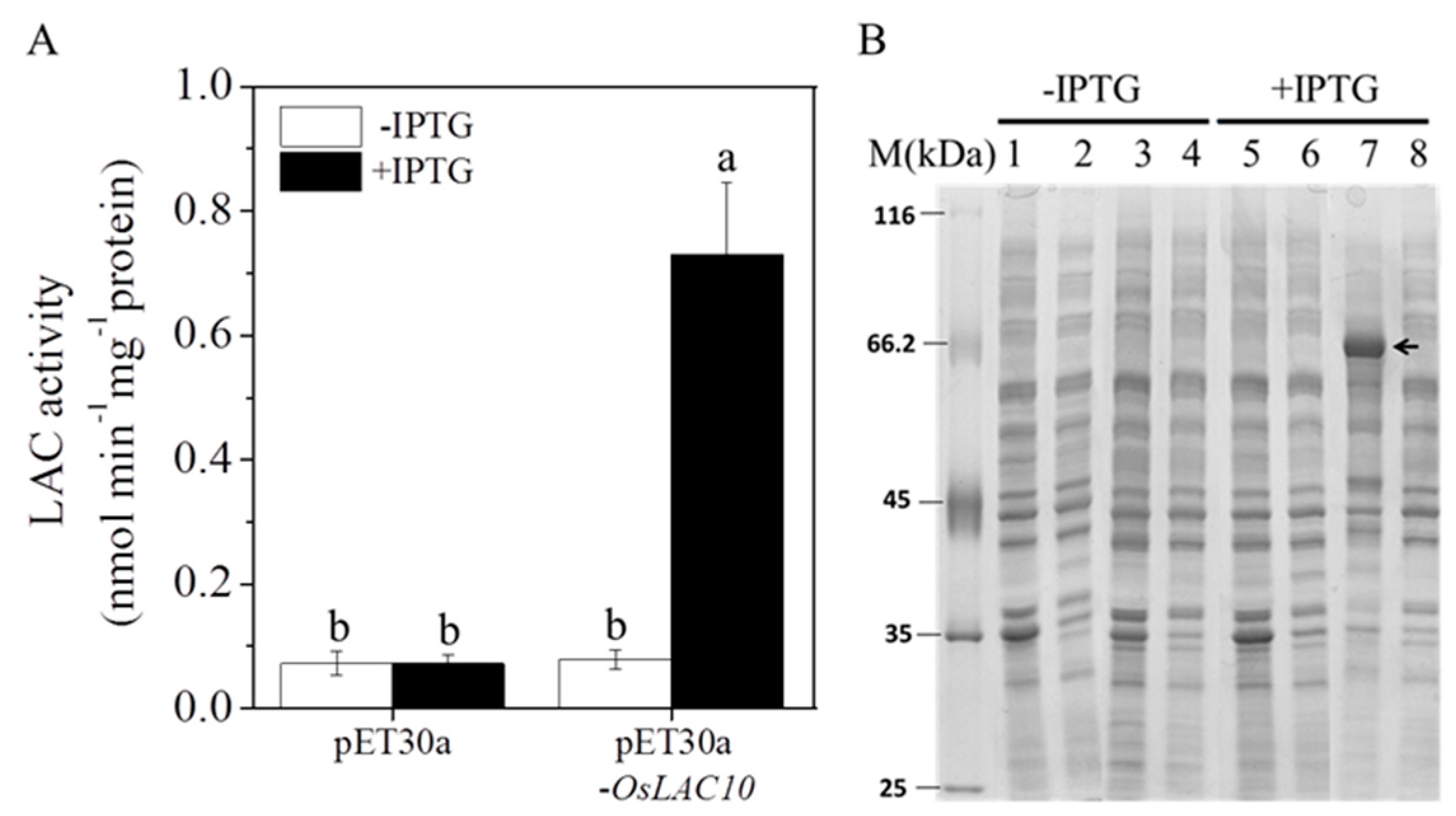
2.5. Expression of the Rice Laccase Protein OsLAC10 in Escherichia coli
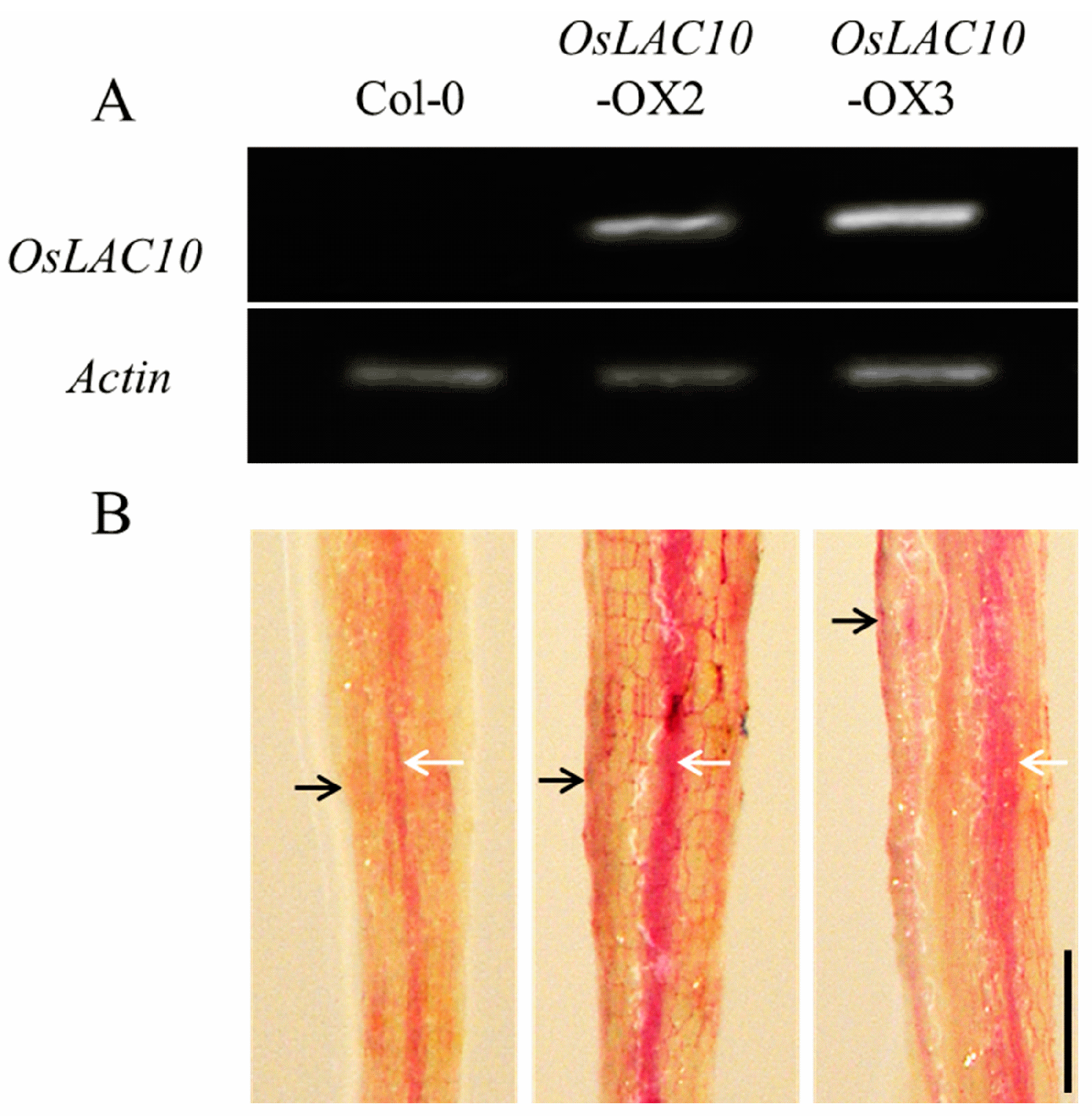
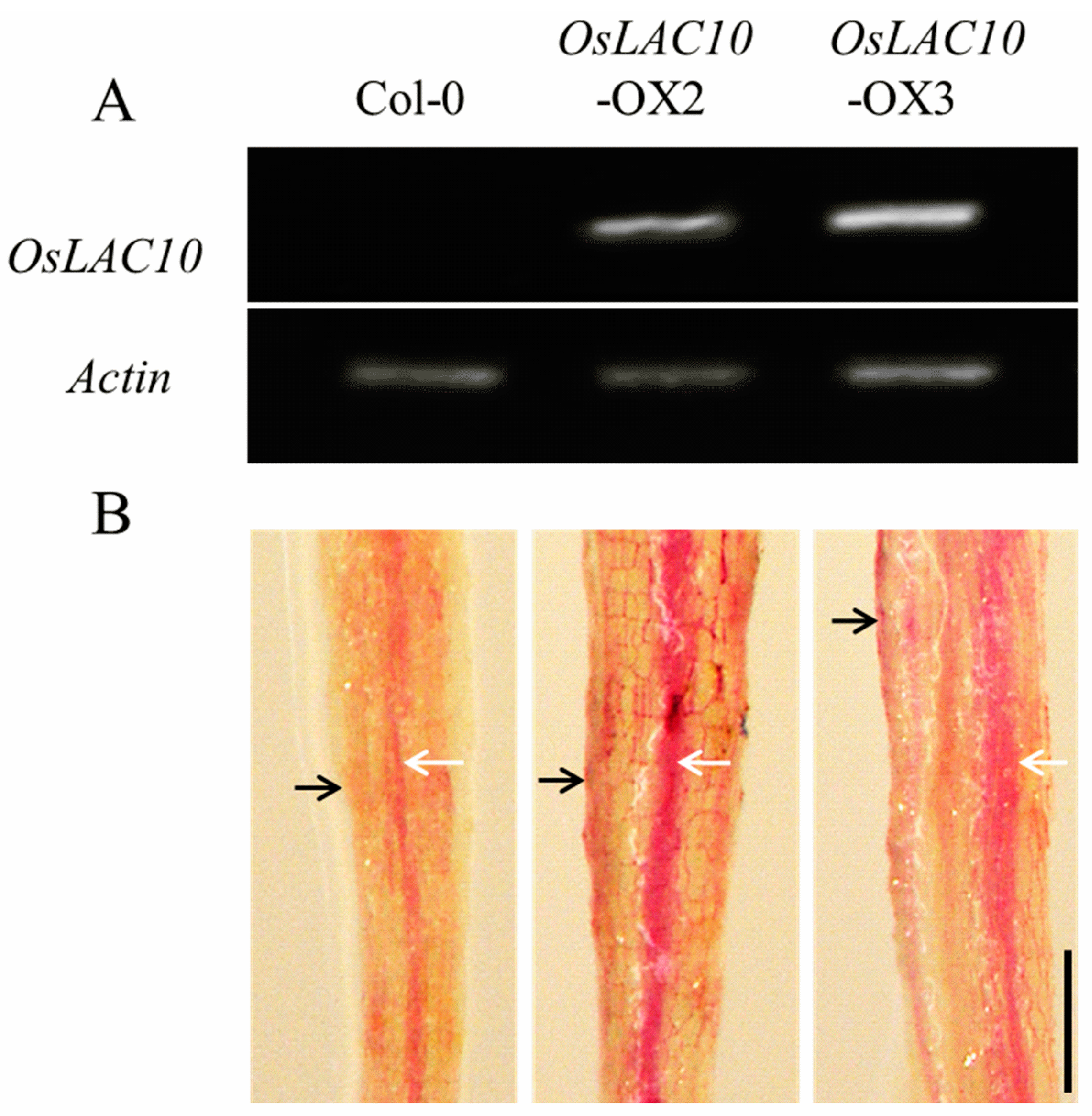
2.6. Ectopic Expression of OsLAC10 in Arabidopsis
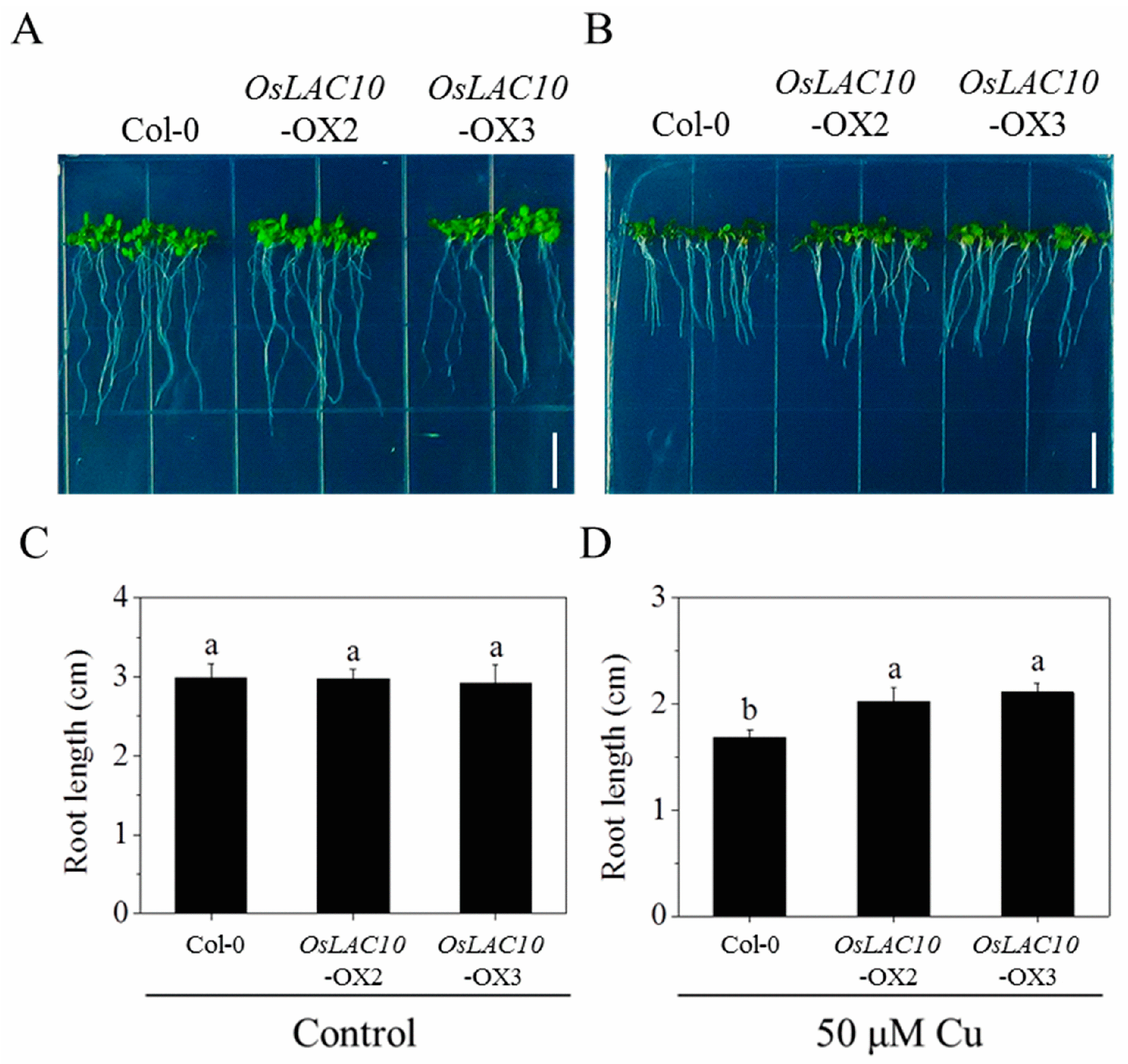
2.7. OsLAC10-Overexpressing Arabidopsis Shows Enhanced Cu Tolerance
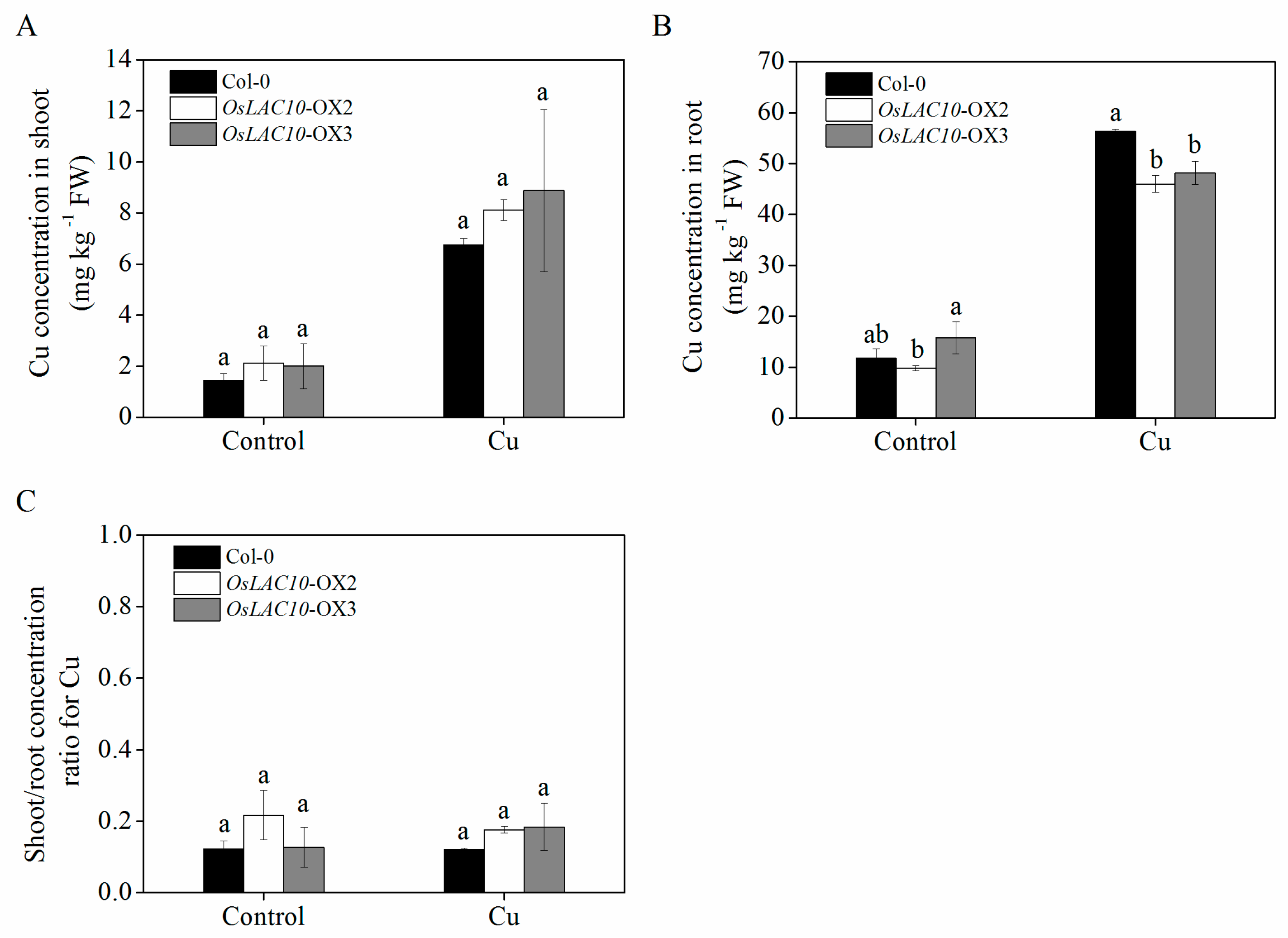
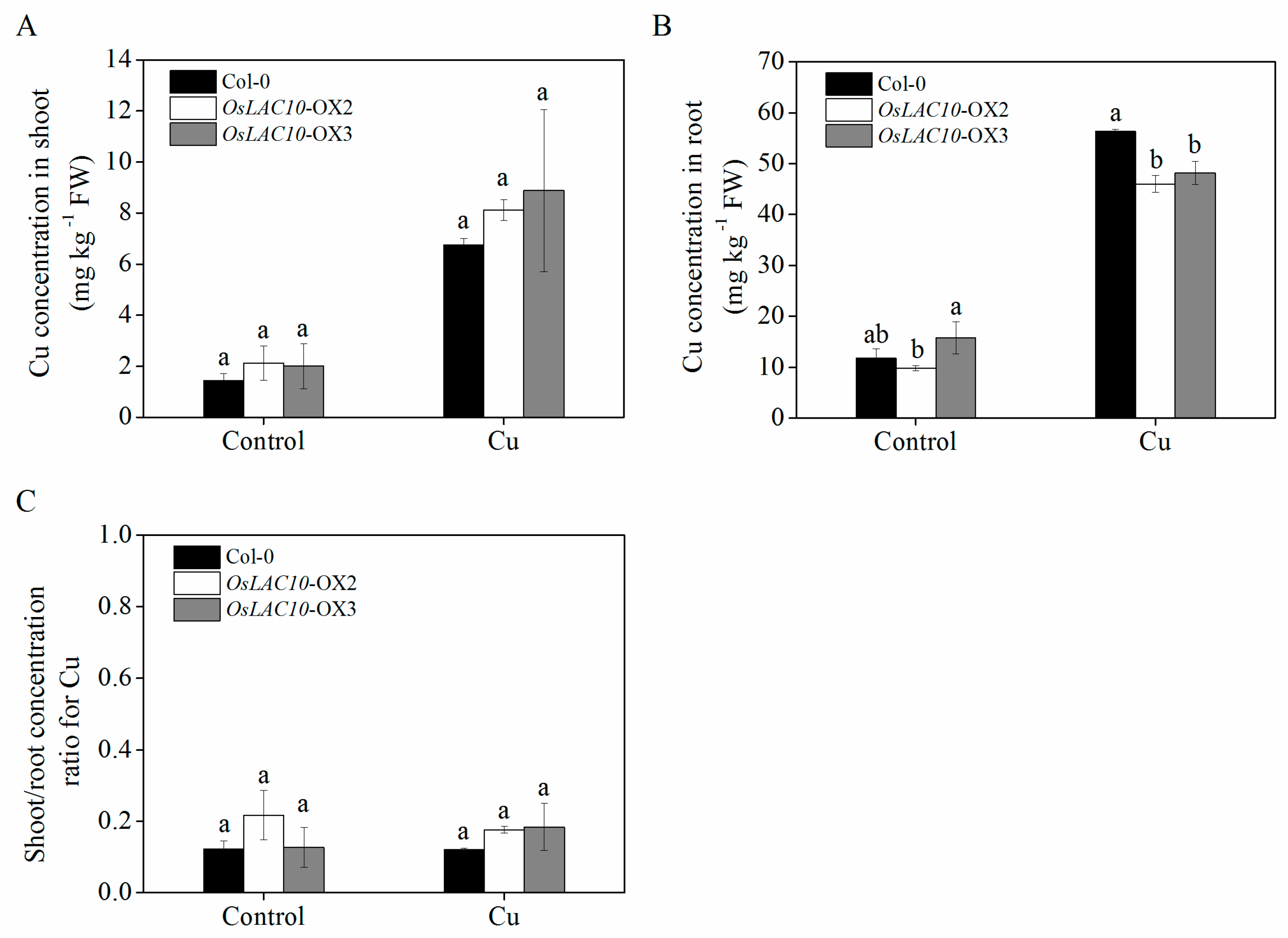
2.8. Analysis of Cu Uptake and Translocation in Transgenic OsLAC10 Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Identification of the Rice Laccase Genes (OsLACs)
4.3. Sequence Analysis and Phylogenetic Tree
4.4. Expression Analysis of Rice Laccase Family Genes
4.5. Quantitative RT-PCR Analysis
4.6. Plasmid Construction and Generation of Transgenic E. coli and Arabidopsis
4.7. Transgenic E. coli Laccase Activity Measurement and SDS-PAGE of Recombinant Proteins
4.8. Lignin Staining and Determination of Cu Concentrations of Transgenic Arabidopsis
4.9. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yoshida, H. Chemistry of lacquer (urushi). Part I. J. Chem. Soc. 1883, 43, 472–486. [Google Scholar] [CrossRef]
- Martínez-Alvarez, O.; Montero, P.; Gómez-Guillén, C. Evidence of an active laccase-like enzyme in deepwater pink shrimp (Parapenaeus longirostris). Food Chem. 2008, 108, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Diamantidis, G.; Effosse, A.; Potier, P.; Bally, R. Purification and characterization of the first bacterial laccase in the rhizospheric bacterium Azospirillum lipoferum. Soil Biol. Biochem. 2000, 32, 919–927. [Google Scholar] [CrossRef]
- Kramer, K.J.; Kanost, M.R.; Hopkins, T.L.; Jiang, H.; Zhu, Y.C.; Xu, R.; Kerwind, J.L.; Tureceke, F. Oxidative conjugation of catechols with proteins in insect skeletal systems. Tetrahedron 2001, 57, 385–392. [Google Scholar] [CrossRef]
- Sterjiades, R.; Dean, J.F.; Eriksson, K.E.L. Laccase from sycamore maple (Acer pseudoplatanus) polymerizes monolignols. Plant Physiol. 1992, 99, 1162–1168. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.; McDougall, G.J. A laccase-type polyphenol oxidase from lignifying xylem of tobacco. Phytochemistry 1997, 44, 229–235. [Google Scholar] [CrossRef]
- LaFayette, P.R.; Eriksson, K.E.L.; Dean, J.F. Characterization and heterologous expression of laccase cDNAs from xylem tissues of yellow-poplar (Liriodendron tulipifera). Plant Mol. Biol. 1999, 40, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Gavnholt, B.; Larsen, K.; Rasmussen, S.K. Isolation and characterisation of laccase cDNAs from meristematic and stem tissues of ryegrass (Lolium perenne). Plant Sci. 2002, 162, 873–885. [Google Scholar] [CrossRef]
- Ranocha, P.; Chabannes, M.; Chamayou, S.; Danoun, S.; Jauneau, A.; Boudet, A.M.; Goffner, D. Laccase down-regulation causes alterations in phenolic metabolism and cell wall structure in poplar. Plant Physiol. 2002, 129, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.D.; Li, Q.J.; Luo, B.; Chen, X.Y. Ex planta phytoremediation of trichlorophenol and phenolic allelochemicals via an engineered secretory laccase. Nat. Biotechnol. 2004, 22, 893–897. [Google Scholar] [CrossRef] [PubMed]
- McCaig, B.C.; Meagher, R.B.; Dean, J.F. Gene structure and molecular analysis of the laccase-like multicopper oxidase (LMCO) gene family in Arabidopsis thaliana. Planta 2005, 221, 619–636. [Google Scholar] [CrossRef] [PubMed]
- Hüttermann, A.; Mai, C.; Kharazipour, A. Modification of lignin for the production of new compounded materials. Appl. Microbiol. Biotechnol. 2001, 55, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; O’malley, D.M.; Whetten, R.; Sederoff, R.R. A laccase associated with lignification in loblolly pine xylem. Science 1993, 260, 672. [Google Scholar] [CrossRef] [PubMed]
- Berthet, S.; Demont-Caulet, N.; Pollet, B.; Bidzinski, P.; Cézard, L.; Le Bris, P.; Borrega, N.; Hervé, J.; Blondet, E.; Balzergue, S.; et al. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems. Plant Cell 2011, 23, 1124–1137. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Nakashima, J.; Chen, F.; Yin, Y.; Fu, C.; Yun, J.; Shao, H.; Wang, X.; Wang, Z.; Dixon, R.A. Laccase is necessary and nonredundant with peroxidase for lignin polymerization during vascular development in Arabidopsis. Plant Cell 2013, 25, 3976–3987. [Google Scholar] [CrossRef] [PubMed]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Turlapati, P.V.; Kim, K.W.; Davin, L.B.; Lewis, N.G. The laccase multigene family in Arabidopsis thaliana: Towards addressing the mystery of their gene function (s). Planta 2011, 233, 439–470. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Davis, E.J.; Ballif, J.; Liang, M.; Bushman, E.; Haroldsen, V.; Torabinejad, J.; Wu, Y. Mutant identification and characterization of the laccase gene family in Arabidopsis. J. Exp. Bot. 2006, 57, 2563–2569. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Davis, E.; Gardner, D.; Cai, X.; Wu, Y. Involvement of AtLAC15 in lignin synthesis in seeds and in root elongation of Arabidopsis. Planta 2006, 224, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.Y.; Lee, C.; Hwang, S.G.; Park, Y.C.; Lim, H.L.; Jang, C.S. Overexpression of the OsChI1 gene, encoding a putative laccase precursor, increases tolerance to drought and salinity stress in transgenic Arabidopsis. Gene 2014, 552, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Haroldsen, V.; Cai, X.; Wu, Y. Expression of a putative laccase gene, ZmLAC1, in maize primary roots under stress. Plant Cell Environ. 2006, 29, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Moura, J.; Bonine, C.; Viana, J.; Dornelas, M.; Mazzafera, P. Abiotic and Biotic Stresses and Changes in the Lignin Content and Composition in Plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; p. 651. [Google Scholar]
- Memon, A.R.; Schröder, P. Implications of metal accumulation mechanisms to phytoremediation. Environ. Sci. Pollut. Res. Int. 2009, 16, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Tester, M.; Leigh, R.A. Partitioning of nutrient transport processes in roots. J. Exp. Bot. 2001, 52, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Puig, S.; Thiele, D.J. Molecular mechanisms of copper uptake and distribution. Curr. Opin. Chem. Biol. 2002, 6, 171–180. [Google Scholar] [CrossRef]
- Hall, J.L.; Williams, L.E. Transition metal transporters in plants. J. Exp. Bot. 2003, 54, 2601–2613. [Google Scholar] [CrossRef] [PubMed]
- Sancenón, V.; Puig, S.; Mira, H.; Thiele, D.J.; Peñarrubia, L. Identification of a copper transporter family in Arabidopsis thaliana. Plant Mol. Biol. 2003, 51, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Colás, N.; Sancenón, V.; Rodríguez-Navarro, S.; Mayo, S.; Thiele, D.J.; Ecker, J.R.; Puig, S.; Peñarrubia, L. The Arabidopsis heavy metal P-type ATPase HMA5 interacts with metallochaperones and functions in copper detoxification of roots. Plant J. 2006, 45, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Yamaji, N.; Yokosho, K.; Ma, J.F. YSL16 is a phloem-localized transporter of the copper-nicotianamine complex that is responsible for copper distribution in rice. Plant Cell 2012, 24, 3767–3782. [Google Scholar] [CrossRef] [PubMed]
- Schmidke, I.; Stephan, U.W. Transport of metal micronutrients in the phloem of castor bean (Ricinus communis) seedlings. Physiol. Plant. 1995, 95, 147–153. [Google Scholar] [CrossRef]
- Arduini, I.; Godbold, D.L.; Onnis, A. Cadmium and copper uptake and distribution in Mediterranean tree seedlings. Physiol. Plant. 1996, 97, 111–117. [Google Scholar] [CrossRef]
- Liu, Q.; Zheng, L.; He, F.; Zhao, F.J.; Shen, Z.; Zheng, L. Transcriptional and physiological analyses identify a regulatory role for hydrogen peroxide in the lignin biosynthesis of copper-stressed rice roots. Plant Soil. 2015, 387, 323–336. [Google Scholar] [CrossRef]
- Ceriotti, A.; Duranti, M.; Bollini, R. Effects of N-glycosylation on the folding and structure of plant proteins. J. Exp. Bot. 1998, 49, 1091–1103. [Google Scholar] [CrossRef]
- Kang, J.S.; Frank, J.; Kang, C.H.; Kajiura, H.; Vikram, M.; Ueda, A.; Kim, S.; Bahk, J.D.; Triplett, B.; Fujiyama, K.; et al. Salt tolerance of Arabidopsis thaliana requires maturation of N-glycosylated proteins in the Golgi apparatus. Proc. Natl. Acad. Sci. USA 2008, 105, 5933–5938. [Google Scholar] [CrossRef] [PubMed]
- Caparrós-Ruiz, D.; Fornalé, S.; Civardi, L.; Puigdomènech, P.; Rigau, J. Isolation and characterisation of a family of laccases in maize. Plant Sci. 2006, 171, 217–225. [Google Scholar] [CrossRef]
- Sato, Y.; Wuli, B.; Sederoff, R.; Whetten, R. Molecular Cloning and Expression of Eight Laccase cDNAs in Loblolly Pine (Pinus taeda). J. Plant Res. 2001, 114, 147–155. [Google Scholar] [CrossRef]
- Birnbaum, K.; Shasha, D.E.; Wang, J.Y.; Jung, J.W.; Lambert, G.M.; Galbraith, D.W.; Benfey, P.N. A gene expression map of the Arabidopsis root. Science 2003, 302, 1956–1960. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Ghany, S.E.; Pilon, M. MicroRNA-mediated systemic down-regulation of copper protein expression in response to low copper availability in Arabidopsis. J. Biol. Chem. 2008, 283, 15932–15945. [Google Scholar] [CrossRef] [PubMed]
- Pourcel, L.; Routaboul, J.M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef] [PubMed]
- Nitta, K.; Kataoka, K.; Sakurai, T. Primary structure of a Japanese lacquer tree laccase as a prototype enzyme of multicopper oxidases. J. Inorg. Biochem. 2002, 91, 125–131. [Google Scholar] [CrossRef]
- Koutaniemi, S.; Malmberg, H.A.; Simola, L.K.; Teeri, T.H.; Kärkönen, A. Norway spruce (Picea abies) laccases: Characterization of a laccase in a lignin-forming tissue culture. J. Integr. Plant Biol. 2015, 57, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Bonawitz, N.D.; Chapple, C. The genetics of lignin biosynthesis: Connecting genotype to phenotype. Annu. Rev. Genet. 2010, 44, 337–363. [Google Scholar] [CrossRef] [PubMed]
- Alejandro, S.; Lee, Y.; Tohge, T.; Sudre, D.; Osorio, S.; Park, J.; Bovet, L.; Lee, Y.; Geldner, N.; Fernie, A.R.; et al. AtABCG29 is a monolignol transporter involved in lignin biosynthesis. Curr. Biol. 2012, 22, 1207–1212. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Jain, M.; Khurana, J.P. Transcript profiling reveals diverse roles of auxin-responsive genes during reproductive development and abiotic stress in rice. FEBS J. 2009, 276, 3148–3162. [Google Scholar] [CrossRef] [PubMed]
- Walia, H.; Wilson, C.; Zeng, L.; Ismail, A.M.; Condamine, P.; Close, T.J. Genome-wide transcriptional analysis of salinity stressed japonica and indica rice genotypes during panicle initiation stage. Plant Mol. Biol. 2007, 63, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Pan, Y.; Zhao, X.; Zhu, L.; Fu, B.; Li, Z. Genome-wide temporal-spatial gene expression profiling of drought responsiveness in rice. BMC Genom. 2011, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Dubey, S.; Misra, P.; Dwivedi, S.; Chatterjee, S.; Bag, S.K.; Mantri, S.; Asif, M.H.; Rai, A.; Kumar, S.; Shri, M.; et al. Transcriptomic and metabolomic shifts in rice roots in response to Cr (VI) stress. BMC Genom. 2010, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Liu, Q.; Zheng, L.; Cui, Y.; Shen, Z.; Zheng, L. RNA-Seq analysis of rice roots reveals the involvement of post-transcriptional regulation in response to cadmium stress. Front. Plant Sci. 2015, 6, 1136. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Deng, X.; Quan, L.; Xia, Y.; Shen, Z. Metallothioneins BcMT1 and BcMT2 from Brassica campestris enhance tolerance to cadmium and copper and decrease production of reactive oxygen species in Arabidopsis thaliana. Plant Soil. 2013, 367, 507–519. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Zhu, M.; Yu, Y.; Zhang, Y.; Wei, Z. Generation and characterization of transgenic poplar plants overexpressing a cotton laccase gene. Plant Cell Tissue Org. 2008, 93, 303–310. [Google Scholar] [CrossRef]
- Lequeux, H.; Hermans, C.; Lutts, S.; Verbruggen, N. Response to copper excess in Arabidopsis thaliana: Impact on the root system architecture, hormone distribution, lignin accumulation and mineral profile. Plant Physiol. Biochem. 2010, 48, 673–682. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus Number | Amino Acids Length | Predicted Target Site | Signal Peptide Length | Cleavage Site | Number of N-glycosyl Sites |
|---|---|---|---|---|---|---|
| OsLAC1 | Os01g0374600 | 599 | Secretory | 28 | AHG-AK | 7 |
| OsLAC2 | Os01g0634500 | 562 | Secretory | 26 | AHA-DV | 13 |
| OsLAC3 | Os01g0827300 | 567 | Secretory | 24 | AGA-EV | 8 |
| OsLAC4 | Os01g0842400 | 579 | Secretory | 28 | AQG-IT | 14 |
| OsLAC5 | Os01g0842500 | 577 | Secretory | 27 | AKG-DI | 11 |
| OsLAC6 | Os01g0843800 | 547 | Mitochondrion | 35 | TAG-LT | 10 |
| OsLAC7 | Os01g0850550 | 580 | Secretory | 22 | AQA-DV | 6 |
| OsLAC8 | Os01g0850700 | 559 | Secretory | 28 | ADA-AT | 15 |
| OsLAC9 | Os01g0850800 | 554 | Secretory | 21 | ASA-AV | 5 |
| OsLAC10 | Os02g0749700 | 579 | Secretory | 21 | ALA-VN | 6 |
| OsLAC11 | Os03g0273200 | 578 | Secretory | 29 | AGA-AT | 15 |
| OsLAC12 | Os03g0297900 | 646 | Secretory | 33 | AVA-EE | 3 |
| OsLAC13 | Os05g0458300 | 513 | - | - | - | - |
| OsLAC14 | Os05g0458500 | 549 | Secretory | 27 | AEA-IT | 11 |
| OsLAC15 | Os05g0458600 | 574 | Secretory | 27 | AEA-IT | 12 |
| OsLAC16 | Os07g0101000 | 583 | Secretory | 30 | VDA-AI | 11 |
| OsLAC17 | Os10g0346300 | 599 | Secretory | - | - | - |
| OsLAC18 | LOC_Os10g30120 | 69 | Secretory | 24 | TNY-TR | 2 |
| OsLAC19 | Os10g0437400 | 467 | Mitochondrion | 28 | VDQ-PR | 5 |
| OsLAC20 | Os11g0108650 | 201 | Secretory | 22 | AAA-KE | 1 |
| OsLAC21 | Os11g0108700 | 326 | - | - | - | 8 |
| OsLAC22 | Os11g0264000 | 595 | Secretory | 29 | GEA-AV | 8 |
| OsLAC23 | Os11g0641500 | 590 | Secretory | 28 | GEA-AV | 8 |
| OsLAC24 | Os11g0641800 | 580 | Secretory | 23 | GEA-GV | 8 |
| OsLAC25 | Os11g0696900 | 583 | Secretory | 29 | AHG-GR | 8 |
| OsLAC26 | Os11g0708100 | 586 | Chloroplast | - | - | - |
| OsLAC27 | Os12g0108000 | 567 | Secretory | 22 | AAA-KE | 11 |
| OsLAC28 | Os12g0257600 | 571 | Secretory | 18 | AHG-AV | 13 |
| OsLAC29 | Os12g0258700 | 579 | Secretory | 23 | AQA-AV | 14 |
| OsLAC30 | Os12g0259800 | 577 | Secretory | 22 | AQA-AV | 10 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Luo, L.; Wang, X.; Shen, Z.; Zheng, L. Comprehensive Analysis of Rice Laccase Gene (OsLAC) Family and Ectopic Expression of OsLAC10 Enhances Tolerance to Copper Stress in Arabidopsis. Int. J. Mol. Sci. 2017, 18, 209. https://doi.org/10.3390/ijms18020209
Liu Q, Luo L, Wang X, Shen Z, Zheng L. Comprehensive Analysis of Rice Laccase Gene (OsLAC) Family and Ectopic Expression of OsLAC10 Enhances Tolerance to Copper Stress in Arabidopsis. International Journal of Molecular Sciences. 2017; 18(2):209. https://doi.org/10.3390/ijms18020209
Chicago/Turabian StyleLiu, Qingquan, Le Luo, Xiaoxiao Wang, Zhenguo Shen, and Luqing Zheng. 2017. "Comprehensive Analysis of Rice Laccase Gene (OsLAC) Family and Ectopic Expression of OsLAC10 Enhances Tolerance to Copper Stress in Arabidopsis" International Journal of Molecular Sciences 18, no. 2: 209. https://doi.org/10.3390/ijms18020209
APA StyleLiu, Q., Luo, L., Wang, X., Shen, Z., & Zheng, L. (2017). Comprehensive Analysis of Rice Laccase Gene (OsLAC) Family and Ectopic Expression of OsLAC10 Enhances Tolerance to Copper Stress in Arabidopsis. International Journal of Molecular Sciences, 18(2), 209. https://doi.org/10.3390/ijms18020209