The FKBP51 Glucocorticoid Receptor Co-Chaperone: Regulation, Function, and Implications in Health and Disease




Abstract
1. Introduction
2. Glucocorticoid Receptor Co-Chaperones, Focus on FKBP51
2.1. Complexity of GR Regulating Chaperones and Co-Chaperones
2.2. Hsp70 and Hsp90 Cofactors
3. Structure and Biochemical Function of FKBP51
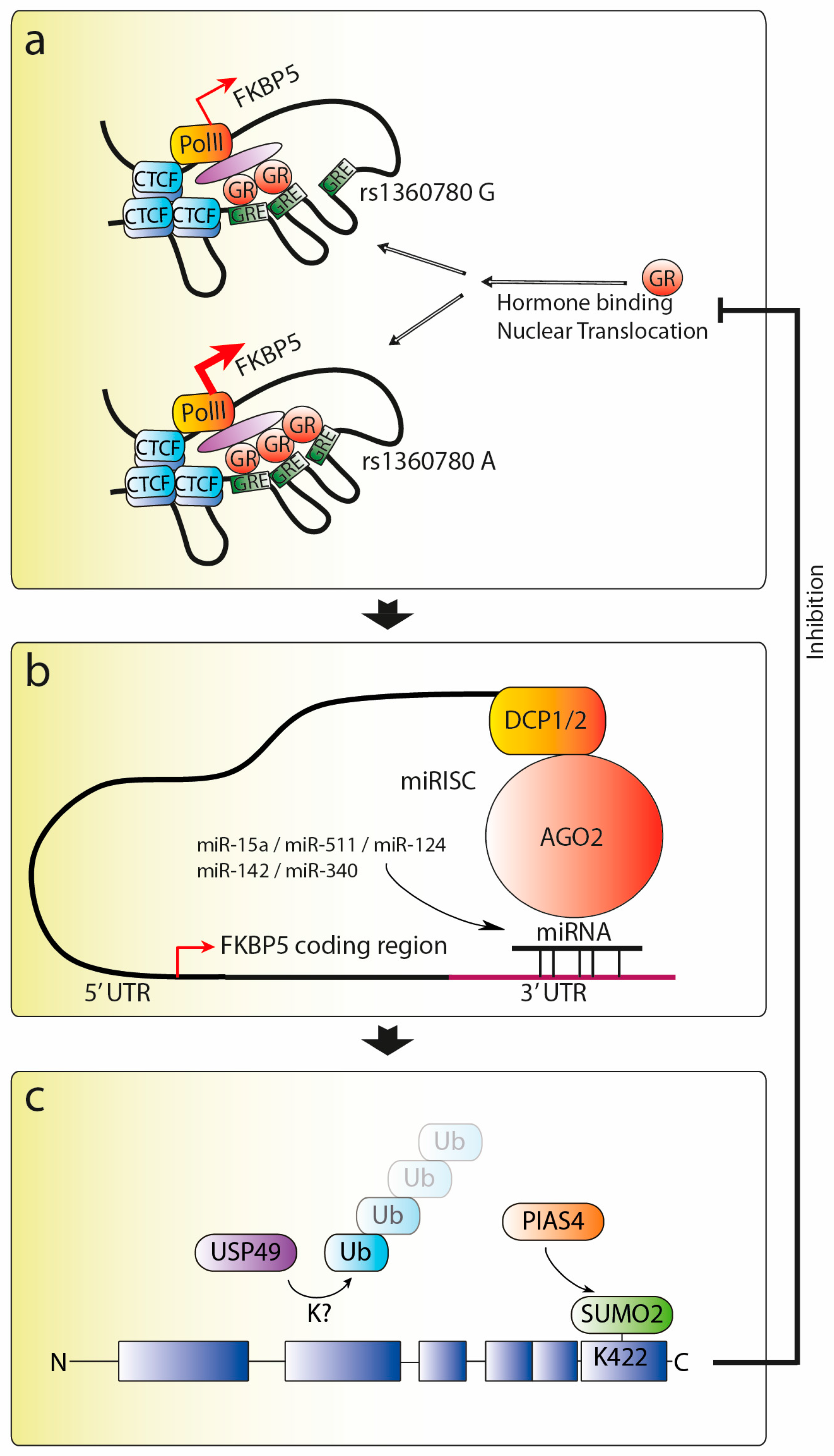
4. Regulation of FKBP51
4.1. Transcriptional Regulation
4.2. Post-Transcriptional Regulation
4.3. Post-Translational Regulation
5. Functions of FKBP51
5.1. Regulation of Immune Pathways
5.2. Regulation of the AKT Pathway
5.3. Regulation of Microtubule Dynamics
5.4. Modulation of Autophagy
5.5. Regulation of DNA Methylation
5.6. Regulation of Metabolism
5.7. Implications in Diseases: Focus on Depression
6. Drugs and FKBP51
6.1. FKBP51 Influences the Effects of Other Drugs
6.2. FKBP51 as A Drug Target
7. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| BAG1 | Bcl2-associated athanogene 1 |
| CDK | cyclin-dependent kinase |
| DNMT1 | DNA methyltransferase 1 |
| FKBP | FK506 binding protein |
| GR | glucocorticoid receptor |
| GRE | glucocorticoid response element |
| HPA | hypothalamus–pituitary–adrenals |
| HSP | heat shock protein |
| HSPBP1 | Hsp70 binding protein 1 |
| MDD | major depressive disorder |
| NFκB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NR | nuclear receptor |
| PHLPP | pleckstrin homology domain leucine-rich repeat protein phosphatase |
| PPIase | peptidylprolylisomerase |
| PTM | post translational modification |
| RISC | RNA-induced gene silencing complex |
| SNP | single nucleotide polymorphisms |
| SR | steroid receptor |
| SUMO | small ubiquitin-like modifier |
| TPR | tetratricopeptide repeat |
References
- Smith, D.F.; Faber, L.E.; Toft, D.O. Purification of unactivated progesterone receptor and identification of novel receptor-associated proteins. J. Biol. Chem. 1990, 265, 3996–4003. [Google Scholar] [PubMed]
- Grad, I.; Picard, D. The glucocorticoid responses are shaped by molecular chaperones. Mol. Cell. Endocrinol. 2007, 275, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, P.C.; Picard, D. Molecular chaperones, essential partners of steroid hormone receptors for activity and mobility. Biochim. Biophys. Acta 2010, 1803, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Pratt, W.B.; Toft, D.O. Steroid receptor interactions with heat shock protein and immunophilin chaperones. Endocr. Rev. 1997, 18, 306–360. [Google Scholar] [CrossRef] [PubMed]
- Pratt, W.B.; Galigniana, M.D.; Harrell, J.M.; DeFranco, D.B. Role of hsp90 and the hsp90-binding immunophilins in signalling protein movement. Cell. Signal. 2004, 16, 857–872. [Google Scholar] [CrossRef] [PubMed]
- Rein, T. FK506 binding protein 51 integrates pathways of adaptation: FKBP51 shapes the reactivity to environmental change. Bioessays 2016, 38, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Wochnik, G.M.; Rüegg, J.; Abel, G.A.; Schmidt, U.; Holsboer, F.; Rein, T. FK506-binding proteins 51 and 52 differentially regulate dynein interaction and nuclear translocation of the glucocorticoid receptor in mammalian cells. J. Biol. Chem. 2005, 280, 4609–4616. [Google Scholar] [CrossRef] [PubMed]
- Denny, W.B.; Valentine, D.L.; Reynolds, P.D.; Smith, D.F.; Scammell, J.G. Squirrel monkey immunophilin FKBP51 is a potent inhibitor of glucocorticoid receptor binding. Endocrinology 2000, 141, 4107–4113. [Google Scholar] [CrossRef] [PubMed]
- Riggs, D.L.; Roberts, P.J.; Chirillo, S.C.; Cheung-Flynn, J.; Prapapanich, V.; Ratajczak, T.; Gaber, R.; Picard, D.; Smith, D.F. The Hsp90-binding peptidylprolyl isomerase FKBP52 potentiates glucocorticoid signaling in vivo. EMBO J. 2003, 22, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Scammell, J.G.; Denny, W.B.; Valentine, D.L.; Smith, D.F. Overexpression of the FK506-binding immunophilin FKBP51 is the common cause of glucocorticoid resistance in three New World primates. Gen. Comp. Endocrinol. 2001, 124, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Holsboer, F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology 2000, 23, 477–501. [Google Scholar] [CrossRef]
- Binder, E.B.; Salyakina, D.; Lichtner, P.; Wochnik, G.M.; Ising, M.; Pütz, B.; Papiol, S.; Seaman, S.; Lucae, S.; Kohli, M.A.; et al. Polymorphisms in FKBP5 are associated with increased recurrence of depressive episodes and rapid response to antidepressant treatment. Nat. Genet. 2004, 36, 1319–1325. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, J.A. Historical overview of nuclear receptors. J. Steroid Biochem. Mol. Biol. 2016, 157, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Gronemeyer, H.; Gustafsson, J.A.; Laudet, V. Principles for modulation of the nuclear receptor superfamily. Nat. Rev. Drug Discov. 2004, 3, 950–964. [Google Scholar] [CrossRef] [PubMed]
- Sladek, F.M. What are nuclear receptor ligands? Mol. Cell. Endocrinol. 2011, 334, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Wrange, O.; Gustafsson, J.A. Separation of the hormone- and DNA-binding sites of the hepatic glucocorticoid receptor by means of proteolysis. J. Biol. Chem. 1978, 253, 856–865. [Google Scholar] [PubMed]
- Bain, D.L.; Heneghan, A.F.; Connaghan-Jones, K.D.; Miura, M.T. Nuclear receptor structure: Implications for function. Annu. Rev. Physiol. 2007, 69, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Germain, P.; Staels, B.; Dacquet, C.; Spedding, M.; Laudet, V. Overview of nomenclature of nuclear receptors. Pharmacol. Rev. 2006, 58, 685–704. [Google Scholar] [CrossRef] [PubMed]
- Garabedian, M.J.; Harris, C.A.; Jeanneteau, F. Glucocorticoid receptor action in metabolic and neuronal function. F1000 Res. 2017, 6, 1208. [Google Scholar] [CrossRef] [PubMed]
- Weikum, E.R.; Knuesel, M.T.; Ortlund, E.A.; Yamamoto, K.R. Glucocorticoid receptor control of transcription: Precision and plasticity via allostery. Nat. Rev. Mol. Cell Biol. 2017, 18, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Caratti, G.; Matthews, L.; Poolman, T.; Kershaw, S.; Baxter, M.; Ray, D. Glucocorticoid receptor function in health and disease. Clin. Endocrinol. 2015, 83, 441–448. [Google Scholar] [CrossRef] [PubMed]
- King, H.A.; Trotter, K.W.; Archer, T.K. Chromatin remodeling during glucocorticoid receptor regulated transactivation. Biochim. Biophys. Acta 2012, 1819, 716–726. [Google Scholar] [CrossRef] [PubMed]
- Picard, D.; Khursheed, B.; Garabedian, M.J.; Fortin, M.G.; Lindquist, S.; Yamamoto, K.R. Reduced levels of hsp90 compromise steroid receptor action in vivo. Nature 1990, 348, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, E.R.; Toft, D.O.; Schlesinger, M.J.; Pratt, W.B. Evidence that the 90-kDa phosphoprotein associated with the untransformed L-cell glucocorticoid receptor is a murine heat shock protein. J. Biol. Chem. 1985, 260, 12398–12401. [Google Scholar] [PubMed]
- Welch, W.J.; Feramisco, J.R. Purification of the major mammalian heat shock proteins. J. Biol. Chem. 1982, 257, 14949–14959. [Google Scholar] [PubMed]
- Schopf, F.H.; Biebl, M.M.; Buchner, J. The HSP90 chaperone machinery. Nat. Rev. Mol. Cell Biol. 2017, 18, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Davey, M.; Hsu, Y.C.; Kaplanek, P.; Tong, A.; Parsons, A.B.; Krogan, N.; Cagney, G.; Mai, D.; Greenblatt, J.; et al. Navigating the chaperone network: An integrative map of physical and genetic interactions mediated by the hsp90 chaperone. Cell 2005, 120, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Taipale, M.; Krykbaeva, I.; Koeva, M.; Kayatekin, C.; Westover, K.D.; Karras, G.I.; Lindquist, S. Quantitative analysis of HSP90-client interactions reveals principles of substrate recognition. Cell 2012, 150, 987–1001. [Google Scholar] [CrossRef] [PubMed]
- Pratt, W.B.; Dittmar, K.D. Studies with Purified Chaperones Advance the Understanding of the Mechanism of Glucocorticoid Receptor-hsp90 Heterocomplex Assembly. Trends Endocrinol. Metab. 1998, 9, 244–252. [Google Scholar] [CrossRef]
- Hernandez, M.P.; Chadli, A.; Toft, D.O. HSP40 binding is the first step in the HSP90 chaperoning pathway for the progesterone receptor. J. Biol. Chem. 2002, 277, 11873–11881. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Yahara, I.; Lindquist, S. Role of the protein chaperone YDJ1 in establishing Hsp90-mediated signal transduction pathways. Science 1995, 268, 1362–1365. [Google Scholar] [CrossRef] [PubMed]
- Laufen, T.; Mayer, M.P.; Beisel, C.; Klostermeier, D.; Mogk, A.; Reinstein, J.; Bukau, B. Mechanism of regulation of hsp70 chaperones by DnaJ cochaperones. Proc. Natl. Acad. Sci. USA 1999, 96, 5452–5457. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.; Smith, D.F. Molecular chaperone interactions with steroid receptors: An update. Mol. Endocrinol. 2000, 14, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Kirschke, E.; Goswami, D.; Southworth, D.; Griffin, P.R.; Agard, D.A. Glucocorticoid receptor function regulated by coordinated action of the Hsp90 and Hsp70 chaperone cycles. Cell 2014, 157, 1685–1697. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.J. Yeast molecular chaperones and the mechanism of steroid hormone action. Trends Endocrinol. Metab. 1997, 8, 271–275. [Google Scholar] [CrossRef]
- Bracher, A.; Verghese, J. The nucleotide exchange factors of Hsp70 molecular chaperones. Front Mol. Biosci. 2015, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Craig, E.A.; Marszalek, J. How Do J-Proteins Get Hsp70 to Do So Many Different Things? Trends Biochem. Sci. 2017, 42, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Duncan, E.J.; Cheetham, M.E.; Chapple, J.P.; van der, S.J. The role of HSP70 and its co-chaperones in protein misfolding, aggregation and disease. Subcell. Biochem. 2015, 78, 243–273. [Google Scholar] [PubMed]
- Sahasrabudhe, P.; Rohrberg, J.; Biebl, M.M.; Rutz, D.A.; Buchner, J. The Plasticity of the Hsp90 Co-chaperone System. Mol. Cell. 2017, 67, 947–961. [Google Scholar] [CrossRef] [PubMed]
- Felts, S.J.; Toft, D.O. p23, a simple protein with complex activities. Cell Stress Chaperones. 2003, 8, 108–113. [Google Scholar] [CrossRef]
- Young, J.C.; Hartl, F.U. Polypeptide release by Hsp90 involves ATP hydrolysis and is enhanced by the co-chaperone p23. EMBO J. 2000, 19, 5930–5940. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Weikl, T.; Bugl, H.; Buchner, J. Chaperone function of Hsp90-associated proteins. Science 1996, 274, 1715–1717. [Google Scholar] [CrossRef] [PubMed]
- Freeman, B.C.; Toft, D.O.; Morimoto, R.I. Molecular chaperone machines: Chaperone activities of the cyclophilin Cyp-40 and the steroid aporeceptor-associated protein p23. Science 1996, 274, 1718–1720. [Google Scholar] [CrossRef] [PubMed]
- Tanioka, T.; Nakatani, Y.; Semmyo, N.; Murakami, M.; Kudo, I. Molecular identification of cytosolic prostaglandin E2 synthase that is functionally coupled with cyclooxygenase-1 in immediate prostaglandin E2 biosynthesis. J. Biol. Chem. 2000, 275, 32775–32782. [Google Scholar] [CrossRef] [PubMed]
- Freeman, B.C.; Yamamoto, K.R. Disassembly of transcriptional regulatory complexes by molecular chaperones. Science 2002, 296, 2232–2235. [Google Scholar] [CrossRef] [PubMed]
- Freeman, B.C.; Felts, S.J.; Toft, D.O.; Yamamoto, K.R. The p23 molecular chaperones act at a late step in intracellular receptor action to differentially affect ligand efficacies. Genes Dev. 2000, 14, 422–434. [Google Scholar] [PubMed]
- Wochnik, G.M.; Young, J.C.; Schmidt, U.; Holsboer, F.; Hartl, F.U.; Rein, T. Inhibition of GR-mediated transcription by p23 requires interaction with Hsp90. FEBS Lett. 2004, 560, 35–38. [Google Scholar] [CrossRef]
- Daugaard, M.; Rohde, M.; Jaattela, M. The heat shock protein 70 family: Highly homologous proteins with overlapping and distinct functions. FEBS Lett. 2007, 581, 3702–3710. [Google Scholar] [CrossRef] [PubMed]
- Kominek, J.; Marszalek, J.; Neuveglise, C.; Craig, E.A.; Williams, B.L. The complex evolutionary dynamics of Hsp70s: A genomic and functional perspective. Genome Biol. Evol. 2013, 5, 2460–2477. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.B.; Shao, Y.M.; Miao, S.; Wang, L. The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cell Mol. Life Sci. 2006, 63, 2560–2570. [Google Scholar] [CrossRef] [PubMed]
- Powers, E.T.; Balch, W.E. Diversity in the origins of proteostasis networks—A driver for protein function in evolution. Nat. Rev. Mol. Cell. Biol. 2013, 14, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Höhfeld, J.; Jentsch, S. GrpE-like regulation of the hsc70 chaperone by the anti-apoptotic protein BAG-1. EMBO J. 1997, 16, 6209–6216. [Google Scholar] [CrossRef] [PubMed]
- Shomura, Y.; Dragovic, Z.; Chang, H.C.; Tzvetkov, N.; Young, J.C.; Brodsky, J.L.; Guerriero, V.; Hartl, F.U.; Bracher, A. Regulation of Hsp70 function by HspBP1: Structural analysis reveals an alternate mechanism for Hsp70 nucleotide exchange. Mol. Cell. 2005, 17, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, U.; Wochnik, G.M.; Rosenhagen, M.C.; Young, J.C.; Hartl, F.U.; Holsboer, F.; Rein, T. Essential role of the unusual DNA-binding motif of BAG-1 for inhibition of the glucocorticoid receptor. J. Biol. Chem. 2003, 278, 4926–4931. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, U.; Holsboer, F.; Rein, T. Role of the hsp70 cochaperone BAG1 in glucocorticoid receptor function and stress-related diseases. Proc. Natl. Acad. Sci. USA 2008, 105, E101. [Google Scholar] [CrossRef] [PubMed]
- Schneikert, J.; Hubner, S.; Langer, G.; Petri, T.; Jaattela, M.; Reed, J.; Cato, A.C. Hsp70-RAP46 interaction in downregulation of DNA binding by glucocorticoid receptor. EMBO J. 2000, 19, 6508–6516. [Google Scholar] [CrossRef] [PubMed]
- Schneikert, J.; Hubner, S.; Martin, E.; Cato, A.C. A nuclear action of the eukaryotic cochaperone RAP46 in downregulation of glucocorticoid receptor activity. J. Cell Biol. 1999, 146, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Knapp, R.T.; Wong, M.J.; Kollmannsberger, L.K.; Gassen, N.C.; Kretzschmar, A.; Zschocke, J.; Hafner, K.; Young, J.C.; Rein, T. Hsp70 cochaperones HspBP1 and BAG-1M differentially regulate steroid hormone receptor function. PLoS ONE 2014, 9, e85415. [Google Scholar] [CrossRef] [PubMed]
- Höhfeld, J.; Minami, Y.; Hartl, F.U. Hip, a novel cochaperone involved in the eukaryotic Hsc70/Hsp40 reaction cycle. Cell 1995, 83, 589–598. [Google Scholar] [CrossRef]
- Kanelakis, K.C.; Murphy, P.J.; Galigniana, M.D.; Morishima, Y.; Takayama, S.; Reed, J.C.; Toft, D.O.; Pratt, W.B. Hsp70 interacting protein Hip does not affect glucocorticoid receptor folding by the hsp90-based chaperone machinery except to oppose the effect of BAG-1. Biochemistry 2000, 39, 14314–14321. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.M.; Prapapanich, V.; Carrigan, P.E.; Roberts, P.J.; Riggs, D.L.; Smith, D.F. The heat shock protein 70 cochaperone hip enhances functional maturation of glucocorticoid receptor. Mol. Endocrinol. 2004, 18, 1620–1630. [Google Scholar] [CrossRef] [PubMed]
- Dittmar, K.D.; Banach, M.; Galigniana, M.D.; Pratt, W.B. The role of DnaJ-like proteins in glucocorticoid receptor.hsp90 heterocomplex assembly by the reconstituted hsp90.p60.hsp70 foldosome complex. J. Biol. Chem. 1998, 273, 7358–7366. [Google Scholar] [CrossRef] [PubMed]
- Mandal, A.K.; Gibney, P.A.; Nillegoda, N.B.; Theodoraki, M.A.; Caplan, A.J.; Morano, K.A. Hsp110 chaperones control client fate determination in the hsp70-Hsp90 chaperone system. Mol. Biol. Cell 2010, 21, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D.; Morano, K.A.; Thiele, D.J. The yeast Hsp110 family member, Sse1, is an Hsp90 cochaperone. J. Biol. Chem. 1999, 274, 26654–26660. [Google Scholar] [CrossRef] [PubMed]
- Harst, A.; Lin, H.; Obermann, W.M. Aha1 competes with Hop, p50 and p23 for binding to the molecular chaperone Hsp90 and contributes to kinase and hormone receptor activation. Biochem. J. 2005, 387, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Lotz, G.P.; Lin, H.; Harst, A.; Obermann, W.M. Aha1 binds to the middle domain of Hsp90, contributes to client protein activation, and stimulates the ATPase activity of the molecular chaperone. J. Biol. Chem. 2003, 278, 17228–17235. [Google Scholar] [CrossRef] [PubMed]
- Panaretou, B.; Siligardi, G.; Meyer, P.; Maloney, A.; Sullivan, J.K.; Singh, S.; Millson, S.H.; Clarke, P.A.; Naaby-Hansen, S.; Stein, R.; et al. Activation of the ATPase activity of hsp90 by the stress-regulated cochaperone aha1. Mol. Cell. 2002, 10, 1307–1318. [Google Scholar] [CrossRef]
- Marsh, J.A.; Kalton, H.M.; Gaber, R.F. Cns1 is an essential protein associated with the hsp90 chaperone complex in Saccharomyces cerevisiae that can restore cyclophilin 40-dependent functions in cpr7Delta cells. Mol. Cell. Biol. 1998, 18, 7353–7359. [Google Scholar] [CrossRef] [PubMed]
- Tesic, M.; Marsh, J.A.; Cullinan, S.B.; Gaber, R.F. Functional interactions between Hsp90 and the co-chaperones Cns1 and Cpr7 in Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 32692–32701. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Periyasamy, S.; Wolf, I.M.; Hinds, T.D., Jr.; Yong, W.; Shou, W.; Sanchez, E.R. Control of glucocorticoid and progesterone receptor subcellular localization by the ligand-binding domain is mediated by distinct interactions with tetratricopeptide repeat proteins. Biochemistry 2008, 47, 10471–10480. [Google Scholar] [CrossRef] [PubMed]
- Schülke, J.P.; Wochnik, G.M.; Lang-Rollin, I.; Gassen, N.C.; Knapp, R.T.; Berning, B.; Yassouridis, A.; Rein, T. Differential impact of tetratricopeptide repeat proteins on the steroid hormone receptors. PLoS ONE 2010, 5, e11717. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, T.; Ward, B.K.; Minchin, R.F. Immunophilin chaperones in steroid receptor signalling. Curr. Top. Med. Chem. 2003, 3, 1348–1357. [Google Scholar] [CrossRef] [PubMed]
- Duina, A.A.; Marsh, J.A.; Kurtz, R.B.; Chang, H.C.; Lindquist, S.; Gaber, R.F. The peptidyl-prolyl isomerase domain of the CyP-40 cyclophilin homolog Cpr7 is not required to support growth or glucocorticoid receptor activity in Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 10819–10822. [Google Scholar] [CrossRef] [PubMed]
- Riggs, D.L.; Cox, M.B.; Tardif, H.L.; Hessling, M.; Buchner, J.; Smith, D.F. Noncatalytic role of the FKBP52 peptidyl-prolyl isomerase domain in the regulation of steroid hormone signaling. Mol. Cell. Biol. 2007, 27, 8658–8669. [Google Scholar] [CrossRef] [PubMed]
- Galigniana, M.D.; Radanyi, C.; Renoir, J.M.; Housley, P.R.; Pratt, W.B. Evidence that the peptidylprolyl isomerase domain of the hsp90-binding immunophilin FKBP52 is involved in both dynein interaction and glucocorticoid receptor movement to the nucleus. J. Biol. Chem. 2001, 276, 14884–14889. [Google Scholar] [CrossRef] [PubMed]
- McKeen, H.D.; McAlpine, K.; Valentine, A.; Quinn, D.J.; McClelland, K.; Byrne, C.; O’Rourke, M.; Young, S.; Scott, C.J.; McCarthy, H.O.; et al. A novel FK506-like binding protein interacts with the glucocorticoid receptor and regulates steroid receptor signaling. Endocrinology 2008, 149, 5724–5734. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, K.A.; Stancato, L.F.; Owens-Grillo, J.K.; Johnson, J.L.; Krishna, P.; Toft, D.O.; Pratt, W.B. The 23-kDa acidic protein in reticulocyte lysate is the weakly bound component of the hsp foldosome that is required for assembly of the glucocorticoid receptor into a functional heterocomplex with hsp90. J. Biol. Chem. 1995, 270, 18841–18847. [Google Scholar] [CrossRef] [PubMed]
- Grad, I.; McKee, T.A.; Ludwig, S.M.; Hoyle, G.W.; Ruiz, P.; Wurst, W.; Floss, T.; Miller, C.A., III; Picard, D. The Hsp90 cochaperone p23 is essential for perinatal survival. Mol. Cell. Biol. 2006, 26, 8976–8983. [Google Scholar] [CrossRef] [PubMed]
- Soroka, J.; Wandinger, S.K.; Mausbacher, N.; Schreiber, T.; Richter, K.; Daub, H.; Buchner, J. Conformational switching of the molecular chaperone Hsp90 via regulated phosphorylation. Mol. Cell 2012, 45, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Wandinger, S.K.; Suhre, M.H.; Wegele, H.; Buchner, J. The phosphatase Ppt1 is a dedicated regulator of the molecular chaperone Hsp90. EMBO J. 2006, 25, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.S.; Silverstein, A.M.; Pratt, W.B.; Chinkers, M. The tetratricopeptide repeat domain of protein phosphatase 5 mediates binding to glucocorticoid receptor heterocomplexes and acts as a dominant negative mutant. J. Biol. Chem. 1996, 271, 32315–32320. [Google Scholar] [CrossRef] [PubMed]
- Pazdrak, K.; Straub, C.; Maroto, R.; Stafford, S.; White, W.I.; Calhoun, W.J.; Kurosky, A. Cytokine-Induced Glucocorticoid Resistance from Eosinophil Activation: Protein Phosphatase 5 Modulation of Glucocorticoid Receptor Phosphorylation and Signaling. J. Immunol. 2016, 197, 3782–3791. [Google Scholar] [CrossRef] [PubMed]
- Laenger, A.; Lang-Rollin, I.; Kozany, C.; Zschocke, J.; Zimmermann, N.; Rüegg, J.; Holsboer, F.; Hausch, F.; Rein, T. XAP2 inhibits glucocorticoid receptor activity in mammalian cells. FEBS Lett. 2009, 583, 1493–1498. [Google Scholar] [CrossRef] [PubMed]
- Connell, P.; Ballinger, C.A.; Jiang, J.; Wu, Y.; Thompson, L.J.; Höhfeld, J.; Patterson, C. The co-chaperone CHIP regulates protein triage decisions mediated by heat-shock proteins. Nat. Cell. Biol. 2001, 3, 93–96. [Google Scholar] [PubMed]
- Alvira, S.; Cuellar, J.; Rohl, A.; Yamamoto, S.; Itoh, H.; Alfonso, C.; Rivas, G.; Buchner, J.; Valpuesta, J.M. Structural characterization of the substrate transfer mechanism in Hsp70/Hsp90 folding machinery mediated by Hop. Nat. Commun. 2014, 5, 5484. [Google Scholar] [CrossRef] [PubMed]
- Brychzy, A.; Rein, T.; Winklhofer, K.F.; Hartl, F.U.; Young, J.C.; Obermann, W.M. Cofactor Tpr2 combines two TPR domains and a J domain to regulate the Hsp70/Hsp90 chaperone system. EMBO J. 2003, 22, 3613–3623. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Garcia, Y.A.; Zierer, B.; Patwardhan, C.; Gutierrez, O.; Hildenbrand, Z.; Harris, D.C.; Balsiger, H.A.; Sivils, J.C.; Johnson, J.L.; et al. The cochaperone SGTA (small glutamine-rich tetratricopeptide repeat-containing protein alpha) demonstrates regulatory specificity for the androgen, glucocorticoid, and progesterone receptors. J. Biol. Chem. 2014, 289, 15297–15308. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, T.; Cluning, C.; Ward, B.K. Steroid Receptor-Associated Immunophilins: A Gateway to Steroid Signalling. Clin. Biochem. Rev. 2015, 36, 31–52. [Google Scholar] [PubMed]
- Scheufler, C.; Brinker, A.; Bourenkov, G.; Pegoraro, S.; Moroder, L.; Bartunik, H.; Hartl, F.U.; Moarefi, I. Structure of TPR domain-peptide complexes: Critical elements in the assembly of the Hsp70-Hsp90 multichaperone machine. Cell 2000, 101, 199–210. [Google Scholar] [CrossRef]
- Hinds, T.D., Jr.; Stechschulte, L.A.; Cash, H.A.; Whisler, D.; Banerjee, A.; Yong, W.; Khuder, S.S.; Kaw, M.K.; Shou, W.; Najjar, S.M.; et al. Protein phosphatase 5 mediates lipid metabolism through reciprocal control of glucocorticoid receptor and peroxisome proliferator-activated receptor-gamma (PPARgamma)1. J. Biol. Chem. 2011, 286, 42911–42922. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; DeFranco, D.B. Chromatin recycling of glucocorticoid receptors: Implications for multiple roles of heat shock protein 90. Mol. Endocrinol. 1999, 13, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Haché, R.J.; Tse, R.; Reich, T.; Savory, J.G.; Lefebvre, Y.A. Nucleocytoplasmic trafficking of steroid-free glucocorticoid receptor. J. Biol. Chem. 1999, 274, 1432–1439. [Google Scholar] [CrossRef] [PubMed]
- Tago, K.; Tsukahara, F.; Naruse, M.; Yoshioka, T.; Takano, K. Regulation of nuclear retention of glucocorticoid receptor by nuclear Hsp90. Mol. Cell. Endocrinol. 2004, 213, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Elbi, C.; Walker, D.A.; Romero, G.; Sullivan, W.P.; Toft, D.O.; Hager, G.L.; DeFranco, D.B. Molecular chaperones function as steroid receptor nuclear mobility factors. Proc. Natl. Acad. Sci. USA 2004, 101, 2876–2881. [Google Scholar] [CrossRef] [PubMed]
- Stavreva, D.A.; Müller, W.G.; Hager, G.L.; Smith, C.L.; McNally, J.G. Rapid glucocorticoid receptor exchange at a promoter is coupled to transcription and regulated by chaperones and proteasomes. Mol. Cell. Biol. 2004, 24, 2682–2697. [Google Scholar] [CrossRef] [PubMed]
- Storer, C.L.; Dickey, C.A.; Galigniana, M.D.; Rein, T.; Cox, M.B. FKBP51 and FKBP52 in signaling and disease. Trends Endocrinol. Metab. 2011, 22, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Pirkl, F.; Buchner, J. Functional analysis of the Hsp90-associated human peptidyl prolyl cis/trans isomerases FKBP51, FKBP52 and Cyp40. J. Mol. Biol. 2001, 308, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Sinars, C.R.; Cheung-Flynn, J.; Rimerman, R.A.; Scammell, J.G.; Smith, D.F.; Clardy, J. Structure of the large FK506-binding protein FKBP51, an Hsp90-binding protein and a component of steroid receptor complexes. Proc. Natl. Acad. Sci. USA 2003, 100, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Brinker, A.; Scheufler, C.; Von Der, M.F.; Fleckenstein, B.; Herrmann, C.; Jung, G.; Moarefi, I.; Hartl, F.U. Ligand discrimination by TPR domains. Relevance and selectivity of EEVD-recognition in Hsp70 x Hop x Hsp90 complexes. J. Biol. Chem. 2002, 277, 19265–19275. [Google Scholar] [CrossRef] [PubMed]
- Ebong, I.O.; Beilsten-Edmands, V.; Patel, N.A.; Morgner, N.; Robinson, C.V. The interchange of immunophilins leads to parallel pathways and different intermediates in the assembly of Hsp90 glucocorticoid receptor complexes. Cell Discov. 2016, 2, 16002. [Google Scholar] [CrossRef] [PubMed]
- Barent, R.L.; Nair, S.C.; Carr, D.C.; Ruan, Y.; Rimerman, R.A.; Fulton, J.; Zhang, Y.; Smith, D.F. Analysis of FKBP51/FKBP52 chimeras and mutants for Hsp90 binding and association with progesterone receptor complexes. Mol. Endocrinol. 1998, 12, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Li, P.; Liu, Y.; Lou, Z.; Ding, Y.; Shu, C.; Ye, S.; Bartlam, M.; Shen, B.; Rao, Z. 3D structure of human FK506-binding protein 52: Implications for the assembly of the glucocorticoid receptor/Hsp90/immunophilin heterocomplex. Proc. Natl. Acad. Sci. USA 2004, 101, 8348–8353. [Google Scholar] [CrossRef] [PubMed]
- Taipale, M.; Jarosz, D.F.; Lindquist, S. HSP90 at the hub of protein homeostasis: Emerging mechanistic insights. Nat. Rev. Mol. Cell. Biol. 2010, 11, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Taipale, M.; Tucker, G.; Peng, J.; Krykbaeva, I.; Lin, Z.Y.; Larsen, B.; Choi, H.; Berger, B.; Gingras, A.C.; Lindquist, S. A quantitative chaperone interaction network reveals the architecture of cellular protein homeostasis pathways. Cell 2014, 158, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Klengel, T.; Mehta, D.; Anacker, C.; Rex-Haffner, M.; Pruessner, J.C.; Pariante, C.M.; Pace, T.W.; Mercer, K.B.; Mayberg, H.S.; Bradley, B.; et al. Allele-specific FKBP5 DNA demethylation mediates gene-childhood trauma interactions. Nat. Neurosci. 2013, 16, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Paakinaho, V.; Makkonen, H.; Jaaskelainen, T.; Palvimo, J.J. Glucocorticoid receptor activates poised FKBP51 locus through long-distance interactions. Mol. Endocrinol. 2010, 24, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Pelleymounter, L.L.; Moon, I.; Johnson, J.A.; Laederach, A.; Halvorsen, M.; Eckloff, B.; Abo, R.; Rossetti, S. A novel application of pattern recognition for accurate SNP and indel discovery from high-throughput data: Targeted resequencing of the glucocorticoid receptor co-chaperone FKBP5 in a Caucasian population. Mol. Genet. Metab. 2011, 104, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Hubler, T.R.; Scammell, J.G. Intronic hormone response elements mediate regulation of FKBP5 by progestins and glucocorticoids. Cell. Stress Chaperones 2004, 9, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Galat, A. A note on clustering the functionally-related paralogues and orthologues of proteins: A case of the FK506-binding proteins (FKBPs). Comput. Biol. Chem. 2004, 28, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Scharf, S.H.; Liebl, C.; Binder, E.B.; Schmidt, M.V.; Müller, M.B. Expression and regulation of the Fkbp5 gene in the adult mouse brain. PLoS ONE 2011, 6, e16883. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.T.; Corces, V.G. CTCF: An architectural protein bridging genome topology and function. Nat. Rev. Genet. 2014, 15, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Oshida, T.; Miyauchi, J.; Yamada, M.; Miyashita, T. Identification of novel direct transcriptional targets of glucocorticoid receptor. Leukemia 2004, 18, 1850–1856. [Google Scholar]
- Magee, J.A.; Chang, L.W.; Stormo, G.D.; Milbrandt, J. Direct, androgen receptor-mediated regulation of the FKBP5 gene via a distal enhancer element. Endocrinology 2006, 147, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.S.; Tamashiro, K.L.; Yang, X.; Purcell, R.H.; Harvey, A.; Willour, V.L.; Huo, Y.; Rongione, M.; Wand, G.S.; Potash, J.B. Chronic corticosterone exposure increases expression and decreases deoxyribonucleic acid methylation of Fkbp5 in mice. Endocrinology 2010, 151, 4332–4343. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Shelton, R.C.; Dwivedi, Y. DNA methylation and expression of stress related genes in PBMC of MDD patients with and without serious suicidal ideation. J. Psychiatr. Res. 2017, 89, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Han, K.M.; Won, E.; Sim, Y.; Kang, J.; Han, C.; Kim, Y.K.; Kim, S.H.; Joe, S.H.; Lee, M.S.; Tae, W.S.; et al. Influence of FKBP5 polymorphism and DNA methylation on structural changes of the brain in major depressive disorder. Sci. Rep. 2017, 7, 42621. [Google Scholar] [CrossRef] [PubMed]
- Sabbagh, J.J.; O’Leary, J.C., III; Blair, L.J.; Klengel, T.; Nordhues, B.A.; Fontaine, S.N.; Binder, E.B.; Dickey, C.A. Age-associated epigenetic upregulation of the FKBP5 gene selectively impairs stress resiliency. PLoS ONE 2014, 9, e107241. [Google Scholar] [CrossRef] [PubMed]
- Fries, G.R.; Carvalho, A.F.; Quevedo, J. The miRNome of bipolar disorder. J. Affect. Disord. 2017, in press. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Chandradoss, S.D.; Depken, M.; Joo, C. Why Argonaute is needed to make microRNA target search fast and reliable. Semin. Cell Dev. Biol. 2017, 65, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Volk, N.; Pape, J.C.; Engel, M.; Zannas, A.S.; Cattane, N.; Cattaneo, A.; Binder, E.B.; Chen, A. Amygdalar MicroRNA-15a Is Essential for Coping with Chronic Stress. Cell Rep. 2016, 17, 1882–1891. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Sabbagh, J.J.; Blair, L.J.; Darling, A.L.; Wen, X.; Dickey, C.A. MicroRNA-511 Binds to FKBP5 mRNA, Which Encodes a Chaperone Protein, and Regulates Neuronal Differentiation. J. Biol. Chem. 2016, 291, 17897–17906. [Google Scholar] [CrossRef] [PubMed]
- Aghajan, M.; Jonai, N.; Flick, K.; Fu, F.; Luo, M.; Cai, X.; Ouni, I.; Pierce, N.; Tang, X.; Lomenick, B.; et al. Chemical genetics screen for enhancers of rapamycin identifies a specific inhibitor of an SCF family E3 ubiquitin ligase. Nat. Biotechnol. 2010, 28, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.B.; Riggs, D.L.; Hessling, M.; Schumacher, F.; Buchner, J.; Smith, D.F. FK506-binding protein 52 phosphorylation: A potential mechanism for regulating steroid hormone receptor activity. Mol. Endocrinol. 2007, 21, 2956–2967. [Google Scholar] [CrossRef] [PubMed]
- Antunica-Noguerol, M.; Budziñski, M.L.; Druker, J.; Gassen, N.C.; Proto-Cassina, L.; Senin, S.; Aprile-Garcia, F.; Holsboer, F.; Rein, T.; Liberman, A.C.; et al. The activity of the glucocorticoid receptor is regulated by SUMO conjugation to FKBP51. Cell Death Differ. 2016, 23, 1579–1591. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, K.A.; Henley, J.M. Mechanisms, regulation and consequences of protein SUMOylation. Biochem. J. 2010, 428, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, F.; Dikic, I. Atypical ubiquitin chains: New molecular signals. ‘Protein Modifications: Beyond the Usual Suspects’ review series. EMBO Rep. 2008, 9, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.; Li, Y.; Yin, Y.; Li, L.; Wu, C.; Chen, Y.; Nowsheen, S.; Hu, Q.; Zhang, L.; Lou, Z.; et al. USP49 negatively regulates tumorigenesis and chemoresistance through FKBP51-AKT signaling. EMBO J. 2017, 36, 1434–1446. [Google Scholar] [CrossRef] [PubMed]
- Bekhbat, M.; Rowson, S.A.; Neigh, G.N. Checks and balances: The glucocorticoid receptor and NFkB in good times and bad. Front. Neuroendocrinol. 2017, 46, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.; Shah, S.; Altonsy, M.O.; Gerber, A.N. Glucocorticoid and cytokine crosstalk: Feedback, feedforward, and co-regulatory interactions determine repression or resistance. J. Biol. Chem. 2017, 292, 7163–7172. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Mallardo, M.; Romano, M.F. FKBP51 and the NF-kappaB regulatory pathway in cancer. Curr. Opin. Pharmacol. 2011, 11, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, T.; Bauch, A.; Ruffner, H.; Angrand, P.O.; Bergamini, G.; Croughton, K.; Cruciat, C.; Eberhard, D.; Gagneur, J.; Ghidelli, S.; et al. A physical and functional map of the human TNF-alpha/NF-kappa B signal transduction pathway. Nat. Cell Biol. 2004, 6, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.M.; Wang, Q.; Sun, W.B. Silencing of FKBP51 alleviates the mechanical pain threshold, inhibits DRG inflammatory factors and pain mediators through the NF-kappaB signaling pathway. Gene 2017, 627, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Li, T.K.; Baksh, S.; Cristillo, A.D.; Bierer, B.E. Calcium- and FK506-independent interaction between the immunophilin FKBP51 and calcineurin. J. Cell Biochem. 2002, 84, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Giraudier, S.; Chagraoui, H.; Komura, E.; Barnache, S.; Blanchet, B.; LeCouedic, J.P.; Smith, D.F.; Larbret, F.; Taksin, A.L.; Moreau-Gachelin, F.; et al. Overexpression of FKBP51 in idiopathic myelofibrosis regulates the growth factor independence of megakaryocyte progenitors. Blood 2002, 100, 2932–2940. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, T.; Shiraishi, T.; Qin, J.; Konno, H.; Akiyama, N.; Shinzawa, M.; Miyauchi, M.; Takizawa, N.; Yanai, H.; Ohashi, H.; et al. Mitochondria-nucleus shuttling FK506-binding protein 51 interacts with TRAF proteins and facilitates the RIG-I-like receptor-mediated expression of type I IFN. PLoS ONE 2014, 9, e95992. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, Y.J.; Lee, J.M.; Kim, E.K.; Park, Y.J.; Choe, S.K.; Ko, H.J.; Kang, C.Y. Functional changes in myeloid-derived suppressor cells (MDSCs) during tumor growth: FKBP51 contributes to the regulation of the immunosuppressive function of MDSCs. J. Immunol. 2012, 188, 4226–4234. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [PubMed]
- Wang, L. FKBP51 regulation of AKT/protein kinase B phosphorylation. Curr. Opin. Pharmacol. 2011, 11, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Fabian, A.K.; Marz, A.; Neimanis, S.; Biondi, R.M.; Kozany, C.; Hausch, F. InterAKTions with FKBPs--mutational and pharmacological exploration. PLoS ONE 2013, 8, e57508. [Google Scholar] [CrossRef] [PubMed]
- Gassen, N.C.; Hartmann, J.; Zschocke, J.; Stepan, J.; Hafner, K.; Zellner, A.; Kirmeier, T.; Kollmannsberger, L.; Wagner, K.V.; Dedic, N.; et al. Association of FKBP51 with Priming of Autophagy Pathways and Mediation of Antidepressant Treatment Response: Evidence in Cells, Mice, and Humans. PLoS Med. 2014, 11, e1001755. [Google Scholar] [CrossRef] [PubMed]
- Jinwal, U.K.; Koren, J., III; Borysov, S.I.; Schmid, A.B.; Abisambra, J.F.; Blair, L.J.; Johnson, A.G.; Jones, J.R.; Shults, C.L.; O’Leary, J.C., III; et al. The Hsp90 cochaperone, FKBP51, increases Tau stability and polymerizes microtubules. J. Neurosci. 2010, 30, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, D.L.; Hubler, T.R.; Scammell, J.G. Organization and function of the FKBP52 and FKBP51 genes. Curr. Opin. Pharmacol. 2011, 11, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Chambraud, B.; Belabes, H.; Fontaine-Lenoir, V.; Fellous, A.; Baulieu, E.E. The immunophilin FKBP52 specifically binds to tubulin and prevents microtubule formation. FASEB J. 2007, 21, 2787–2797. [Google Scholar] [CrossRef] [PubMed]
- Quinta, H.R.; Maschi, D.; Gomez-Sanchez, C.; Piwien-Pilipuk, G.; Galigniana, M.D. Subcellular rearrangement of hsp90-binding immunophilins accompanies neuronal differentiation and neurite outgrowth. J. Neurochem. 2010, 115, 716–734. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; D’Angelillo, A.; Pacelli, R.; Staibano, S.; De, L.E.; Bisogni, R.; Eskelinen, E.L.; Mascolo, M.; Cali, G.; Arra, C.; et al. Role of FK506-binding protein 51 in the control of apoptosis of irradiated melanoma cells. Cell Death. Differ. 2010, 17, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Gassen, N.C.; Hartmann, J.; Schmidt, M.V.; Rein, T. FKBP5/FKBP51 enhances autophagy to synergize with antidepressant action. Autophagy 2015, 11, 578–580. [Google Scholar] [CrossRef] [PubMed]
- Laane, E.; Tamm, K.P.; Buentke, E.; Ito, K.; Kharaziha, P.; Oscarsson, J.; Corcoran, M.; Bjorklund, A.C.; Hultenby, K.; Lundin, J.; et al. Cell death induced by dexamethasone in lymphoid leukemia is mediated through initiation of autophagy. Cell Death. Differ. 2009, 16, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Zhi, X.; Feng, W.; Rong, Y.; Liu, R. Anatomy of autophagy: From the beginning to the end. Cell. Mol. Life Sci. 2017, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, V.; Verstreken, P. Autophagy in the presynaptic compartment in health and disease. J. Cell Biol. 2017, 216, 1895–1906. [Google Scholar] [CrossRef] [PubMed]
- Gassen, N.C.; Fries, G.R.; Zannas, A.S.; Hartmann, J.; Zschocke, J.; Hafner, K.; Carrillo-Roa, T.; Steinbacher, J.; Preissinger, S.N.; Hoeijmakers, L.; et al. Chaperoning epigenetics: FKBP51 decreases the activity of DNMT1 and mediates epigenetic effects of the antidepressant paroxetine. Sci. Signal. 2015, 8, ra119. [Google Scholar] [CrossRef] [PubMed]
- Lyko, F. The DNA methyltransferase family: A versatile toolkit for epigenetic regulation. Nat. Rev. Genet. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, G.; St-Pierre, Y. Phosphorylation of human DNMT1: Implication of cyclin-dependent kinases. Biochem. Biophys. Res. Commun. 2011, 409, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Ota, M.; Hori, H.; Hattori, K.; Teraishi, T.; Matsuo, J.; Kinoshita, Y.; Ishida, I.; Nagashima, A.; Kunugi, H. The common functional FKBP5 variant rs1360780 is associated with altered cognitive function in aged individuals. Sci. Rep. 2014, 4, 6696. [Google Scholar] [CrossRef] [PubMed]
- Mamdani, F.; Rollins, B.; Morgan, L.; Myers, R.M.; Barchas, J.D.; Schatzberg, A.F.; Watson, S.J.; Akil, H.; Potkin, S.G.; Bunney, W.E.; et al. Variable telomere length across post-mortem human brain regions and specific reduction in the hippocampus of major depressive disorder. Transl. Psychiatr. 2015, 5, e636. [Google Scholar] [CrossRef] [PubMed]
- Lagadari, M.; Zgajnar, N.R.; Gallo, L.I.; Galigniana, M.D. Hsp90-binding immunophilin FKBP51 forms complexes with hTERT enhancing telomerase activity. Mol. Oncol. 2016, 10, 1086–1098. [Google Scholar] [CrossRef] [PubMed]
- Zannas, A.S.; Balsevich, G.; Gassen, N.C. The emerging role of FKBP5 in the regulation of metabolism and body weight. Surg. Obes. Relat. Dis. 2016, 12, 1560–1561. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.J.; Palming, J.; Svensson, M.K.; Rizell, M.; Dalenback, J.; Hammar, M.; Fall, T.; Sidibeh, C.O.; Svensson, P.A.; Eriksson, J.W. FKBP5 expression in human adipose tissue increases following dexamethasone exposure and is associated with insulin resistance. Metabolism 2014, 63, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Fichna, M.; Krzysko-Pieczka, I.; Zurawek, M.; Skowronska, B.; Januszkiewicz-Lewandowska, D.; Fichna, P. FKBP5 polymorphism is associated with insulin resistance in children and adolescents with obesity. Obes. Res. Clin. Pract. 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, I.B.; Fries, G.R.; Bucker, J.; Scotton, E.; von, D.L.; Kauer-Sant’Anna, M. The FKBP5 polymorphism rs1360780 is associated with lower weight loss after bariatric surgery: 26 months of follow-up. Surg. Obes. Relat. Dis. 2016, 12, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Balsevich, G.; Uribe, A.; Wagner, K.V.; Hartmann, J.; Santarelli, S.; Labermaier, C.; Schmidt, M.V. Interplay between diet-induced obesity and chronic stress in mice: Potential role of FKBP51. J. Endocrinol. 2014, 222, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Isoda, F.; Yen, K.; Kleopoulos, S.P.; Janssen, W.; Fan, X.; Mastaitis, J.; Dunn-Meynell, A.; Levin, B.; McCrimmon, R.; et al. Hypothalamic Fkbp51 is induced by fasting, and elevated hypothalamic expression promotes obese phenotypes. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E987–E991. [Google Scholar] [CrossRef] [PubMed]
- Stechschulte, L.A.; Qiu, B.; Warrier, M.; Hinds, T.D., Jr.; Zhang, M.; Gu, H.; Xu, Y.; Khuder, S.S.; Russo, L.; Najjar, S.M.; et al. FKBP51 Null Mice Are Resistant to Diet-Induced Obesity and the PPARgamma Agonist Rosiglitazone. Endocrinology 2016, 157, 3888–3900. [Google Scholar] [CrossRef] [PubMed]
- Stechschulte, L.A.; Hinds, T.D., Jr.; Khuder, S.S.; Shou, W.; Najjar, S.M.; Sanchez, E.R. FKBP51 controls cellular adipogenesis through p38 kinase-mediated phosphorylation of GRalpha and PPARgamma. Mol. Endocrinol. 2014, 28, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Qiu, B.; Wang, T.; Wang, J.; Liu, M.; Xu, Y.; Wang, C.; Deng, R.; Williams, K.; Yang, Z.; et al. Loss of FKBP5 impedes adipocyte differentiation under both normoxia and hypoxic stress. Biochem. Biophys. Res. Commun. 2017, 485, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Dogan, M.V.; Lei, M.K.; Beach, S.R.; Brody, G.H.; Philibert, R.A. Alcohol and tobacco consumption alter hypothalamic pituitary adrenal axis DNA methylation. Psychoneuroendocrinology 2016, 66, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Levran, O.; Randesi, M.; Li, Y.; Rotrosen, J.; Ott, J.; Adelson, M.; Kreek, M.J. Drug addiction and stress-response genetic variability: Association study in African Americans. Ann. Hum. Genet. 2014, 78, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Crider, A.; Pandya, C.D.; Ahmed, A.O.; Pillai, A. Altered mRNA Levels of Glucocorticoid Receptor, Mineralocorticoid Receptor, and Co-Chaperones (FKBP5 and PTGES3) in the Middle Frontal Gyrus of Autism Spectrum Disorder Subjects. Mol. Neurobiol. 2016, 53, 2090–2099. [Google Scholar] [CrossRef] [PubMed]
- Seifuddin, F.; Pirooznia, M.; Judy, J.T.; Goes, F.S.; Potash, J.B.; Zandi, P.P. Systematic review of genome-wide gene expression studies of bipolar disorder. BMC Psychiatry 2013, 13, 213. [Google Scholar] [CrossRef] [PubMed]
- Fries, G.R.; Moreno, M.P.V.; Gubert, C.; Santos, B.T.M.Q.; Sartori, J.; Eisele, B.; Ferrari, P.; Fijtman, A.; Rüegg, J.; Gassen, N.C.; et al. Hypothalamic-pituitary-adrenal axis dysfunction and illness progression in bipolar disorder. Int. J. Neuropsychopharmacol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Misiak, B.; Stramecki, F.; Gaweda, L.; Prochwicz, K.; Sasiadek, M.M.; Moustafa, A.A.; Frydecka, D. Interactions Between Variation in Candidate Genes and Environmental Factors in the Etiology of Schizophrenia and Bipolar Disorder: A Systematic Review. Mol. Neurobiol. 2017, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Fani, N.; King, T.Z.; Shin, J.; Srivastava, A.; Brewster, R.C.; Jovanovic, T.; Bradley, B.; Ressler, K.J. Structural and functional connectivity in posttraumatic stress disorder: Associations with fkbp5. Depress. Anxiety 2016, 33, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Watkins, L.E.; Han, S.; Harpaz-Rotem, I.; Mota, N.P.; Southwick, S.M.; Krystal, J.H.; Gelernter, J.; Pietrzak, R.H. FKBP5 polymorphisms, childhood abuse, and PTSD symptoms: Results from the National Health and Resilience in Veterans Study. Psychoneuroendocrinology 2016, 69, 98–105. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz-Cano, E. Association between FKBP5 and CRHR1 genes with suicidal behavior: A systematic review. Behav. Brain Res. 2017, 317, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Isaksson, J.; Allen, M.; Nilsson, K.W.; Lindblad, F. Polymorphisms in the FK506 binding protein 5 gene are associated with attention deficit hyperactivity disorder and diurnal cortisol levels. Acta Paediatr. 2015, 104, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Minelli, A.; Maffioletti, E.; Cloninger, C.R.; Magri, C.; Sartori, R.; Bortolomasi, M.; Congiu, C.; Bignotti, S.; Segala, M.; Giacopuzzi, M.; et al. Role of allelic variants of FK506-binding protein 51 (FKBP5) gene in the development of anxiety disorders. Depress. Anxiety 2013, 30, 1170–1176. [Google Scholar] [CrossRef] [PubMed]
- Ising, M.; Depping, A.M.; Siebertz, A.; Lucae, S.; Unschuld, P.G.; Kloiber, S.; Horstmann, S.; Uhr, M.; Müller-Myhsok, B.; Holsboer, F. Polymorphisms in the FKBP5 gene region modulate recovery from psychosocial stress in healthy controls. Eur. J. Neurosci. 2008, 28, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.; Enoch, M.A.; Gorodetsky, E.; Akhtar, L.; Schuebel, K.; Roy, A.; Goldman, D. The influence of FKBP5 genotype on expression of FKBP5 and other glucocorticoid-regulated genes, dependent on trauma exposure. Genes Brain Behav. 2017, 16, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Lessard, J.; Holman, E.A. FKBP5 and CRHR1 polymorphisms moderate the stress-physical health association in a national sample. Health Psychol. 2014, 33, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Maric, N.P.; Adzic, M. Pharmacological modulation of HPA axis in depression—New avenues for potential therapeutic benefits. Psychiatr. Danub. 2013, 25, 299–305. [Google Scholar] [PubMed]
- Dean, J.; Keshavan, M. The neurobiology of depression: An integrated view. Asian J. Psychiatry 2017, 27, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Pariante, C.M.; Lightman, S.L. The HPA axis in major depression: Classical theories and new developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Lukic, I.; Mitic, M.; Soldatovic, I.; Jovicic, M.; Maric, N.; Radulovic, J.; Adzic, M. Accumulation of Cytoplasmic Glucocorticoid Receptor Is Related to Elevation of FKBP5 in Lymphocytes of Depressed Patients 3573. J. Mol. Neurosci. 2014, 55, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Tatro, E.T.; Everall, I.P.; Masliah, E.; Hult, B.J.; Lucero, G.; Chana, G.; Soontornniyomkij, V.; Achim, C.L. Differential expression of immunophilins FKBP51 and FKBP52 in the frontal cortex of HIV-infected patients with major depressive disorder. J. Neuroimmune Pharmacol. 2009, 4, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Frodl, T.; Carballedo, A.; Frey, E.M.; O’Keane, V.; Skokauskas, N.; Morris, D.; Gill, M.; Hughes, M.M.; Harkin, A.; Connor, T. Expression of glucocorticoid inducible genes is associated with reductions in cornu ammonis and dentate gyrus volumes in patients with major depressive disorder. Dev. Psychopathol. 2014, 26, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Menke, A.; Arloth, J.; Putz, B.; Weber, P.; Klengel, T.; Mehta, D.; Gonik, M.; Rex-Haffner, M.; Rubel, J.; Uhr, M.; et al. Dexamethasone stimulated gene expression in peripheral blood is a sensitive marker for glucocorticoid receptor resistance in depressed patients. Neuropsychopharmacology 2012, 37, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Menke, A.; Klengel, T.; Rubel, J.; Bruckl, T.; Pfister, H.; Lucae, S.; Uhr, M.; Holsboer, F.; Binder, E.B. Genetic variation in FKBP5 associated with the extent of stress hormone dysregulation in major depression. Genes Brain Behav. 2013, 12, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Cordova-Palomera, A.; de Reus, M.A.; Fatjo-Vilas, M.; Falcon, C.; Bargallo, N.; van den Heuvel, M.P.; Fananas, L. FKBP5 modulates the hippocampal connectivity deficits in depression: A study in twins. Brain Imaging Behav. 2017, 11, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Tozzi, L.; Carballedo, A.; Wetterling, F.; McCarthy, H.; O’Keane, V.; Gill, M.; Morris, D.; Fahey, C.; Meaney, J.; Frodl, T. Single-Nucleotide Polymorphism of the FKBP5 Gene and Childhood Maltreatment as Predictors of Structural Changes in Brain Areas Involved in Emotional Processing in Depression. Neuropsychopharmacology 2016, 41, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Zobel, A.; Schuhmacher, A.; Jessen, F.; Hofels, S.; Von, W.O.; Metten, M.; Pfeiffer, U.; Hanses, C.; Becker, T.; Rietschel, M.; et al. DNA sequence variants of the FKBP5 gene are associated with unipolar depression. Int. J. Neuropsychopharmacol. 2010, 13, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Lahti, J.; la-Mikkula, H.; Kajantie, E.; Haljas, K.; Eriksson, J.G.; Raikkonen, K. Associations Between Self-Reported and Objectively Recorded Early Life Stress, FKBP5 Polymorphisms, and Depressive Symptoms in Midlife. Biol. Psychiatry 2016, 80, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Brückl, T.; Nocon, A.; Pfister, H.; Binder, E.B.; Uhr, M.; Lieb, R.; Moffitt, T.E.; Caspi, A.; Holsboer, F.; et al. Interaction of variants in the FKBP5 gene and adverse life events in predicting the first depression onset: Results from a ten-year prospective community study. Am. J. Psychiatry 2011, 168, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Appel, K.; Schwahn, C.; Mahler, J.; Schulz, A.; Spitzer, C.; Fenske, K.; Stender, J.; Barnow, S.; John, U.; Teumer, A.; et al. Moderation of adult depression by a polymorphism in the FKBP5 gene and childhood physical abuse in the general population. Neuropsychopharmacology 2011, 36, 1982–1991. [Google Scholar] [CrossRef] [PubMed]
- Dackis, M.N.; Rogosch, F.A.; Oshri, A.; Cicchetti, D. The role of limbic system irritability in linking history of childhood maltreatment and psychiatric outcomes in low-income, high-risk women: Moderation by FK506 binding protein 5 haplotype 3714. Dev. Psychopathol. 2012, 24, 1237–1252. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Yao, Y.; Ryan, J.; Li, T.; Wang, D.; Zheng, C.; Xu, Y.; Xu, Q. Common variants in FKBP5 gene and major depressive disorder (MDD) susceptibility: A comprehensive meta-analysis. Sci. Rep. 2016, 6, 32687. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, A.; Gennarelli, M.; Uher, R.; Breen, G.; Farmer, A.; Aitchison, K.J.; Craig, I.W.; Anacker, C.; Zunsztain, P.A.; McGuffin, P.; et al. Candidate genes expression profile associated with antidepressants response in the GENDEP study: Differentiating between baseline ‘predictors’ and longitudinal ‘targets’. Neuropsychopharmacology 2013, 38, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Kirchheiner, J.; Lorch, R.; Lebedeva, E.; Seeringer, A.; Roots, I.; Sasse, J.; Brockmoller, J. Genetic variants in FKBP5 affecting response to antidepressant drug treatment. Pharmacogenomics 2008, 9, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Horstmann, S.; Lucae, S.; Menke, A.; Hennings, J.M.; Ising, M.; Roeske, D.; Müller-Myhsok, B.; Holsboer, F.; Binder, E.B. Polymorphisms in GRIK4, HTR2A, and FKBP5 show interactive effects in predicting remission to antidepressant treatment. Neuropsychopharmacology 2010, 35, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Stamm, T.J.; Rampp, C.; Wiethoff, K.; Stingl, J.; Mossner, R.; Malley, O.; Ricken, R.; Seemüller, F.; Keck, M.; Fisher, R.; et al. The FKBP5 polymorphism rs1360780 influences the effect of an algorithm-based antidepressant treatment and is associated with remission in patients with major depression. J. Psychopharmacol. 2016, 30, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, K.A.; Moon, I.; Eckloff, B.W.; Fridley, B.L.; Jenkins, G.D.; Batzler, A.; Biernacka, J.M.; Abo, R.; Brisbin, A.; Ji, Y.; et al. FKBP5 genetic variation: Association with selective serotonin reuptake inhibitor treatment outcomes in major depressive disorder. Pharmacogenet. Genom. 2013, 23, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Lekman, M.; Laje, G.; Charney, D.; Rush, A.J.; Wilson, A.F.; Sorant, A.J.; Lipsky, R.; Wisniewski, S.R.; Manji, H.; McMahon, F.J.; et al. The FKBP5-gene in depression and treatment response—An association study in the Sequenced Treatment Alternatives to Relieve Depression (STAR*D) Cohort. Biol. Psychiatry 2008, 63, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Sarginson, J.E.; Lazzeroni, L.C.; Ryan, H.S.; Schatzberg, A.F.; Murphy, G.M., Jr. FKBP5 polymorphisms and antidepressant response in geriatric depression. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2010, 153B, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.J.; Hong, C.J.; Chen, T.J.; Yu, Y.W. Lack of supporting evidence for a genetic association of the FKBP5 polymorphism and response to antidepressant treatment 26. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2007, 144B, 1097–1098. [Google Scholar] [CrossRef] [PubMed]
- Niitsu, T.; Fabbri, C.; Bentini, F.; Serretti, A. Pharmacogenetics in major depression: A comprehensive meta-analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 45, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Gassen, N.C.; Hartmann, J.; Zannas, A.S.; Kretzschmar, A.; Zschocke, J.; Maccarrone, G.; Hafner, K.; Zellner, A.; Kollmannsberger, L.K.; Wagner, K.V.; et al. FKBP51 inhibits GSK3beta and augments the effects of distinct psychotropic medications. Mol. Psychiatry 2016, 21, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Katz, E.R.; Stowe, Z.N.; Newport, D.J.; Kelley, M.E.; Pace, T.W.; Cubells, J.F.; Binder, E.B. Regulation of mRNA expression encoding chaperone and co-chaperone proteins of the glucocorticoid receptor in peripheral blood: Association with depressive symptoms during pregnancy. Psychol. Med. 2012, 42, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Tatro, E.T.; Nguyen, T.B.; Bousman, C.A.; Masliah, E.; Grant, I.; Atkinson, J.H.; Everall, I.P. Correlation of major depressive disorder symptoms with FKBP5 but not FKBP4 expression in human immunodeficiency virus-infected individuals. J. Neurovirol. 2010, 16, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Perroud, N.; Bondolfi, G.; Uher, R.; Gex-Fabry, M.; Aubry, J.M.; Bertschy, G.; Malafosse, A.; Kosel, M. Clinical and genetic correlates of suicidal ideation during antidepressant treatment in a depressed outpatient sample. Pharmacogenomics 2011, 12, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, A.; Kondo, K.; Saito, T.; Esaki, K.; Otsuka, Y.; Mano, K.; Ikeda, M.; Iwata, N. A genetic variant in 12q13, a possible risk factor for bipolar disorder, is associated with depressive state, accounting for stressful life events. PLoS ONE 2014, 9, e115135. [Google Scholar] [CrossRef] [PubMed]
- Höhne, N.; Poidinger, M.; Merz, F.; Pfister, H.; Bruckl, T.; Zimmermann, P.; Uhr, M.; Holsboer, F.; Ising, M. FKBP5 Genotype-Dependent DNA Methylation and mRNA Regulation after Psychosocial Stress in Remitted Depression and Healthy Controls. Int. J. Neuropsychopharmacol. 2014, 18. [Google Scholar] [CrossRef] [PubMed]
- VanZomeren-Dohm, A.A.; Pitula, C.E.; Koss, K.J.; Thomas, K.; Gunnar, M.R. FKBP5 moderation of depressive symptoms in peer victimized, post-institutionalized children. Psychoneuroendocrinology 2015, 51, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Owens, M.; Harrison, A.J.; Burkhouse, K.L.; McGeary, J.E.; Knopik, V.S.; Palmer, R.H.; Gibb, B.E. Eye tracking indices of attentional bias in children of depressed mothers: Polygenic influences help to clarify previous mixed findings. Dev. Psychopathol. 2016, 28, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Szczepankiewicz, A.; Leszczynska-Rodziewicz, A.; Pawlak, J.; Narozna, B.; Rajewska-Rager, A.; Wilkosc, M.; Zaremba, D.; Maciukiewicz, M.; Twarowska-Hauser, J. FKBP5 polymorphism is associated with major depression but not with bipolar disorder. J. Affect. Disord. 2014, 164, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Suppli, N.P.; Bukh, J.D.; Moffitt, T.E.; Caspi, A.; Johansen, C.; Tjonneland, A.; Kessing, L.V.; Dalton, S.O. Genetic variants in 5-HTTLPR, BDNF, HTR1A, COMT, and FKBP5 and risk for treated depression after cancer diagnosis. Depress. Anxiety 2017, 34, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Engel, A.; Fasching, P.A.; Haberle, L.; Binder, E.B.; Voigt, F.; Grimm, J.; Faschingbauer, F.; Eichler, A.; Dammer, U.; et al. Genetic variants in the genes of the stress hormone signalling pathway and depressive symptoms during and after pregnancy. Biomed. Res. Int. 2014, 2014, 469278. [Google Scholar] [CrossRef] [PubMed]
- Velders, F.P.; Kuningas, M.; Kumari, M.; Dekker, M.J.; Uitterlinden, A.G.; Kirschbaum, C.; Hek, K.; Hofman, A.; Verhulst, F.C.; Kivimaki, M.; et al. Genetics of cortisol secretion and depressive symptoms: A candidate gene and genome wide association approach. Psychoneuroendocrinology 2011, 36, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Udina, M.; Navines, R.; Egmond, E.; Oriolo, G.; Langohr, K.; Gimenez, D.; Valdes, M.; Gomez-Gil, E.; Grande, I.; Gratacos, M.; et al. Glucocorticoid Receptors, Brain-Derived Neurotrophic Factor, Serotonin and Dopamine Neurotransmission are Associated with Interferon-Induced Depression. Int. J. Neuropsychopharmacol. 2016, 19. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.; Collishaw, S.; Harold, G.; Rice, F.; Thapar, A. Maternal depression and child and adolescent depression symptoms: An exploratory test for moderation by CRHR1, FKBP5 and NR3C1 gene variants. Behav. Genet. 2012, 42, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, G.; Jowsey, S.; Amer, H.; Biernacka, J.; Colby, C.; Walker, D.; Black, J.; Rundell, J.; Stegall, M.; Mrazek, D.A. Relationship between FKBP5 polymorphisms and depression symptoms among kidney transplant recipients. Depress. Anxiety 2011, 28, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Borelli, J.L.; Smiley, P.A.; Rasmussen, H.F.; Gomez, A.; Seaman, L.C.; Nurmi, E.L. Interactive effects of attachment and FKBP5 genotype on school-aged children’s emotion regulation and depressive symptoms. Behav. Brain Res. 2017, 325, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Matsumoto, Y.; Sadahiro, R.; Enokido, M.; Goto, K.; Otani, K. Relationship of the FKBP5 C/T polymorphism with dysfunctional attitudes predisposing to depression. Compr. Psychiatry 2014, 55, 1422–1425. [Google Scholar] [CrossRef] [PubMed]
- Sarubin, N.; Hilbert, S.; Naumann, F.; Zill, P.; Wimmer, A.M.; Nothdurfter, C.; Rupprecht, R.; Baghai, T.C.; Buhner, M.; Schule, C. The sex-dependent role of the glucocorticoid receptor in depression: Variations in the NR3C1 gene are associated with major depressive disorder in women but not in men. Eur. Arch. Psychiatry Clin. Neurosci. 2017, 267, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Lavebratt, C.; Aberg, E.; Sjoholm, L.K.; Forsell, Y. Variations in FKBP5 and BDNF genes are suggestively associated with depression in a Swedish population-based cohort. J. Affect. Disord. 2010, 125, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Arlt, S.; Demiralay, C.; Tharun, B.; Geisel, O.; Storm, N.; Eichenlaub, M.; Lehmbeck, J.T.; Wiedemann, K.; Leuenberger, B.; Jahn, H. Genetic risk factors for depression in Alzheimer’s disease patients. Curr. Alzheimer Res. 2013, 10, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Shelton, R.C.; Dwivedi, Y. Interaction between early-life stress and FKBP5 gene variants in major depressive disorder and post-traumatic stress disorder: A systematic review and meta-analysis. J. Affect. Disord. 2018, 225, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Papiol, S.; Arias, B.; Gastó, C.; Gutiérrez, B.; Catalán, R.; Fañanás, L. Genetic variability at HPA axis in major depression and clinical response to antidepressant treatment. J. Affect. Disord. 2007, 104, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, T. Steroid Receptor-Associated Immunophilins: Candidates for Diverse Drug-Targeting Approaches in Disease. Curr. Mol. Pharmacol. 2015, 9, 66–95. [Google Scholar] [CrossRef] [PubMed]
- Mazaira, G.I.; Camisay, M.F.; De, L.S.; Erlejman, A.G.; Galigniana, M.D. Biological relevance of Hsp90-binding immunophilins in cancer development and treatment. Int. J. Cancer 2016, 138, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Periyasamy, S.; Hinds, T., Jr.; Shemshedini, L.; Shou, W.; Sanchez, E.R. FKBP51 and Cyp40 are positive regulators of androgen-dependent prostate cancer cell growth and the targets of FK506 and cyclosporin A. Oncogene 2010, 29, 1691–1701. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Yang, C.S.; Gioeli, D.; Frierson, H.; Toft, D.O.; Paschal, B.M. FKBP51 promotes assembly of the Hsp90 chaperone complex and regulates androgen receptor signaling in prostate cancer cells. Mol. Cell. Biol. 2010, 30, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Pei, H.; Li, L.; Fridley, B.L.; Jenkins, G.D.; Kalari, K.R.; Lingle, W.; Petersen, G.; Lou, Z.; Wang, L. FKBP51 affects cancer cell response to chemotherapy by negatively regulating Akt. Cancer Cell 2009, 16, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.B.; Hong, Y.; Dhe-Paganon, S.; Yoon, H.S. FKBP family proteins: Immunophilins with versatile biological functions. Neurosignals 2008, 16, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Van Duyne, G.D.; Standaert, R.F.; Karplus, P.A.; Schreiber, S.L.; Clardy, J. Atomic structure of FKBP-FK506, an immunophilin-immunosuppressant complex. Science 1991, 252, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Farmer, J.D., Jr.; Lane, W.S.; Friedman, J.; Weissman, I.; Schreiber, S.L. Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. Cell 1991, 66, 807–815. [Google Scholar] [CrossRef]
- Allan, R.K.; Mok, D.; Ward, B.K.; Ratajczak, T. Modulation of chaperone function and cochaperone interaction by novobiocin in the C-terminal domain of Hsp90: Evidence that coumarin antibiotics disrupt Hsp90 dimerization. J. Biol. Chem. 2006, 281, 7161–7171. [Google Scholar] [CrossRef] [PubMed]
- McConnell, J.R.; Alexander, L.A.; McAlpine, S.R. A heat shock protein 90 inhibitor that modulates the immunophilins and regulates hormone receptors without inducing the heat shock response. Bioorg. Med. Chem. Lett. 2014, 24, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Kabouridis, P.S. Biological applications of protein transduction technology. Trends Biotechnol. 2003, 21, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Horibe, T.; Torisawa, A.; Kohno, M.; Kawakami, K. Molecular mechanism of cytotoxicity induced by Hsp90-targeted Antp-TPR hybrid peptide in glioblastoma cells. Mol. Cancer 2012, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Horibe, T.; Kohno, M.; Haramoto, M.; Ohara, K.; Kawakami, K. Designed hybrid TPR peptide targeting Hsp90 as a novel anticancer agent. J. Transl. Med. 2011, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Moche, M.; Winblad, B.; Pavlov, P.F. Combined X-ray crystallography and computational modeling approach to investigate the Hsp90 C-terminal peptide binding to FKBP51. Sci. Rep. 2017, 7, 14288. [Google Scholar] [CrossRef] [PubMed]
- Gaali, S.; Kirschner, A.; Cuboni, S.; Hartmann, J.; Kozany, C.; Balsevich, G.; Namendorf, C.; Fernandez-Vizarra, P.; Sippel, C.; Zannas, A.S.; et al. Selective inhibitors of the FK506-binding protein 51 by induced fit. Nat. Chem. Biol. 2015, 11, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Pomplun, S.; Hausch, F. Recent Progress in FKBP Ligand Development. Curr. Mol. Pharmacol. 2015, 9, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, J.; Wagner, K.V.; Gaali, S.; Kirschner, A.; Kozany, C.; Ruhter, G.; Dedic, N.; Hausl, A.S.; Hoeijmakers, L.; Westerholz, S.; et al. Pharmacological Inhibition of the Psychiatric Risk Factor FKBP51 Has Anxiolytic Properties. J. Neurosci. 2015, 35, 9007–9016. [Google Scholar] [CrossRef] [PubMed]
- Maiaru, M.; Tochiki, K.K.; Cox, M.B.; Annan, L.V.; Bell, C.G.; Feng, X.; Hausch, F.; Geranton, S.M. The stress regulator FKBP51 drives chronic pain by modulating spinal glucocorticoid signaling. Sci. Transl. Med. 2016, 8, 325ra19. [Google Scholar] [CrossRef] [PubMed]
- Pomplun, S.; Wang, Y.; Kirschner, A.; Kozany, C.; Bracher, A.; Hausch, F. Rational design and asymmetric synthesis of potent and neurotrophic ligands for FK506-binding proteins (FKBPs). Angew. Chem. Int. Ed. Engl. 2015, 54, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Bracher, A.; Kozany, C.; Thost, A.K.; Hausch, F. Structural characterization of the PPIase domain of FKBP51, a cochaperone of human Hsp90. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Bai, Q.; Zhou, S.; Liu, X.; Liu, H.; Yao, X. Molecular dynamics simulation, binding free energy calculation and unbinding pathway analysis on selectivity difference between FKBP51 and FKBP52: Insight into the molecular mechanism of isoform selectivity. Proteins 2017. [Google Scholar] [CrossRef] [PubMed]
- LeMaster, D.M.; Hernandez, G. Conformational Dynamics in FKBP Domains: Relevance to Molecular Signaling and Drug Design. Curr. Mol. Pharmacol. 2015, 9, 5–26. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Ota, M.; Hori, H.; Hattori, K.; Teraishi, T.; Sasayama, D.; Higuchi, T.; Kunugi, H. Association between the common functional FKBP5 variant (rs1360780) and brain structure in a non-clinical population. J. Psychiatr. Res. 2014, 58, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Akiyoshi, J.; Muronaga, M.; Tanaka, Y.; Ishitobi, Y.; Inoue, A.; Oshita, H.; Aizawa, S.; Masuda, K.; Higuma, H.; et al. FKBP5 is associated with amygdala volume in the human brain and mood state: A voxel-based morphometry (VBM) study. Int. J. Psychiatry Clin. Pract. 2016, 20, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Holz, N.E.; Buchmann, A.F.; Boecker, R.; Blomeyer, D.; Baumeister, S.; Wolf, I.; Rietschel, M.; Witt, S.H.; Plichta, M.M.; Meyer-Lindenberg, A.; et al. Role of FKBP5 in emotion processing: Results on amygdala activity, connectivity and volume. Brain Struct. Funct. 2015, 220, 1355–1368. [Google Scholar] [CrossRef] [PubMed]
- Bryant, R.A.; Felmingham, K.L.; Liddell, B.; Das, P.; Malhi, G.S. Association of FKBP5 polymorphisms and resting-state activity in a frontotemporal-parietal network. Transl. Psychiatry 2016, 6, e925. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Name | Description | Citations |
|---|---|---|
| Hsp70 cofactors | ||
| BAG1 (Bcl2 Associated Athanogene 1) | Nucleotide exchange factor for Hsp70 proteins. L- and M-forms inhibit GR in mammalian cells. | [52,54,56,58] |
| Hip (Hsp70 interacting protein, ST13) | Stabilizes the ADP bound state of Hsp70, opposes the inhibitory effect of BAG1, enhances GR function in the yeast model, possibly also independently of Hsp70. | [59,60,61] |
| Hsp40 (Heat shock protein 40) | Family of J-domain proteins, enhances Hsp70’s ATP hydrolysis, one of the 5 components in vitro reconstitution system, forms the initial contact in SR folding. | [29,30,62] |
| Hsp110 (Heat shock protein 110) | Sse1 in yeast, related to and nucleotide exchange factor of Hsp70, enhances GR maturation in the yeast model. | [63,64] |
| HSPBP1 (Hsp70 binding protein 1) | Nucleotide exchange factor of Hsp70. Overexpression in mammalian cells decreases GR-dependent transcription. | [53,58] |
| Hsp90 cofactors | ||
| Aha1 (Activator of Hsp 90 ATPase activity 1) | Stimulates the ATPase activity of Hsp90. Enhances GR function in the yeast model and in mammalian cells. | [65,66,67] |
| Cns1 (cyclophilin seven suppressor) | TPR domain containing protein. Evidence from yeast suggests enhancement of GR function. | [68,69] |
| Cyp40 (Cyclophilin 40, Cpr6 and Cpr7 in yeast) | TPR domain containing protein and PPIase. Influences GR in some yeast models, no association and effect shown in mammalian cells. | [70,71,72,73] |
| FKBP51 (FK506 binding protein 51) | TPR domain containing protein and PPIase. Potent inhibitor of GR in mammalian cells. Decreases hormone affinity and delays nuclear translocation. | [7,8] |
| FKBP52 (FK506 binding protein 52) | TPR domain containing protein and PPIase. Potentiates GR activity in the yeast model. Context-dependency in mammalian cells. | [7,9,74,75] |
| FKBPL (FK506 binding protein like, WISp39) | TPR domain containing protein with PPIase-like domain. Affects levels, nuclear translocation, and transcriptional activity of GR in a cell line dependent manner. | [76] |
| P23 (also known as PTGES3, prostaglandin E synthase 3) | Part of the basal in vitro reconstitution system. Promotes Hsp90’s ATP-bound state and substrate-interaction. Context-dependent action on GR activity, chromatin effects. | [2,45,46,47,77,78] |
| PP5 (protein phosphatase 5, Ppt1 in yeast) | Contains TPR domain protein, dephosphorylates Hsp90. Enhances Hsp90 and GR activity in the yeast model. Overexpression reduces GR activity in mammalian cells. | [71,79,80,81,82] |
| Tah1 (TPR-containing protein associated with Hsp90) | TPR domain containing protein, in complex with Pih1. Evidence from yeast suggests stimulatory effect on GR. | [27] |
| SGT1 (Suppressor of G2 allele of Skp1) | Enhances Hsp90 chaperone cycle and GR activity in the yeast model. | [39] |
| XAP2 (HBV X- Associated Protein 2 = FKBP37) | TPR domain containing protein with PPIase-like domain. Interferes with GR activity upon overexpression in mammalian cells. | [83] |
| Cofactors for both Hsp70 and Hsp90 | ||
| CHIP (Carboxy- terminus of Hsp70- binding protein | TPR domain containing protein, has E3 ubiquitin ligase activity. Reduces GR protein levels and function in mammalian cells. | [71,84] |
| Hop (Hsp70–Hsp90 organizing protein) | Part of the basal in vitro foldosome reconstitution system. Mediates the transfer from the Hsp70- to the Hsp90-based folding platform. | [2,34,85] |
| TPR2 (TPR repeat protein 2 = DNAJC7) | Features TPR- and J-domain. Stimulates ATP hydrolysis and polypeptide binding by Hsp70. Excess inhibits GR folding in vitro and activity in vivo. | [71,86] |
| SGTA (Small Gluta- mine-rich TPR-con- taining Protein α) | TPR domain containing protein. Knock-down and ectopic expression in yeast and mammalian cells evidence an inhibitory role on GR function. | [87] |
| Ref. | FKBP51 Measure | Sample | Finding |
|---|---|---|---|
| [183] | Protein levels in blood | Patients with MDD (n = 30) and healthy controls (n = 35) | Higher FKBP51 levels in the cytoplasm of blood cells from depressed patients. Significant correlation between cytoplasmic GR and FKBP5 levels. |
| [186] | mRNA levels in blood after dexamethasone administration | Two cohorts: (1) n = 18 cases/18 controls; (2) n = 11 cases/13 controls | Lower FKBP5 induction after dexamethasone in patients compared to controls. |
| [185] | mRNA levels in the hippocampus (cornu ammonis and dentate gyrus) | MDD patients (n = 43) and healthy controls (n = 43) | No difference between groups. No association between FKBP5 expression and hippocampal volumes. |
| [206] | mRNA levels in blood | Pregnant women with a lifetime history of mood or anxiety disorders (n = 106) | Effect of gestation trimester in upregulating FKBP5 expression. Reduced magnitude of upregulation of FKBP5 across pregnancy in depressed women. |
| [155] | mRNA levels in the hippocampus | MDD patients (n = 10) and controls (n = 10) | Increased FKBP5 mRNA levels in patients compared to controls. |
| [184] | Protein and mRNA levels in the frontal cortex | Controls (n = 12), MDD patients (n = 12), MDD with psychosis (n = 12), HIV+ (n = 11), and HIV+ with MDD (n = 11) | Elevated FKBP51 at both the transcript and protein levels correlated with MDD. Higher frequency of the rs3800373 CC genotype in the MDD and MDD/psychosis groups. |
| [207] | mRNA levels in blood and genotype (rs3800373) | MDD patients at first visit and euthymic at second visit (n = 56); euthymic patients at first visit and MDD at second visit (n = 30) | Correlation of change in the severity of depressive mood with FKBP5 gene expression in individuals homozygous for GG of the SNP rs3800373. |
| [115] | DNA methylation and mRNA levels in PBMCs | Controls (n = 20), MDD patients with (n = 14) or without serious suicidal ideation (n = 10) | Hypermethylation of FKBP5 gene in MDD patients with concomitant reduced expression of FKBP5 transcript variants 1, 2, and 3. Further analyses showed that differences were primarily seen in the MDD-suicide group compared to controls. |
| [116] | Genotype (rs1360780) and DNA methylation | MDD patients (n = 114) and healthy controls (n = 88) | Interaction between the FKBP5 genotype and MDD diagnosis on gray matter volumes of several brain regions. Allele-specific positive correlation of the FKBP5 gene methylation with thickness of the transverse frontopolar gyrus. |
| [191] | Genotype (rs1360780, rs9470080, rs9394309) | N = 1431 participants with available data on early life stress and depressive symptoms at midlife | FKBP5 SNPs interacted with early life stress exposure, but not with recent stressful life events, in predicting self-reported depressive symptoms in midlife. |
| [208] | Genotype (rs1360780) | N = 131 outpatients with moderate to severe depression | The FKBP5 rs1360780 T allele was significantly associated with antidepressant treatment increasing suicidal ideation, with a partly drug-specific pattern of association. |
| [209] | Genotype (rs1360780) | N = 922 hospital staff members (depressive state, n = 309; non-depressive state, n = 613) | Significant association of the FKBP5 SNP as main effect on depressive state. No significant association between the depressive state and the SNP x stressful life events interaction. |
| [190] | Genotype (several SNPs) | Inpatients with unipolar depression (n = 268) and controls (n = 284) | Association between SNPs (especially rs3800373 and rs4713916) in the FKBP5 gene with vulnerability to unipolar depression. Association of the FKBP5 genotype with HPA axis activity after citalopram treatment, as well as with right hippocampal volume. |
| [210] | Genotype, mRNA expression and DNA methylation in blood | Patients with remitted major depression (n = 61) and healthy controls (n = 55) | Genotype-dependent plasma cortisol response to psychosocial stress exposure in controls (not in patients). rs1360780 T-carrier controls responded with a blunted FKBP5 mRNA expression after psychosocial stress. Depression- and genotype-specific differences in FKBP5 methylation in intron 7. |
| [211] | Genotype (rs1360780) | N = 489 children | Significant three-way interaction between FKBP5 genotype, victimization, and child sex predicting depressive symptoms. |
| [212] | Genotype (rs1360780) | Children of mothers with (n = 81) and without (n = 81) a history of depression | Children of depressed mothers who carried the reactive genotype of FKBP5 rs1360780 exhibited less sustained attention to sad faces and more sustained attention to happy faces (information-processing bias). |
| [188] | Genotype (rs1360780) | N = 115 monozygotic twin pairs discordant or concordant for depression | Additive effect of right hippocampal connectivity alterations and FKBP5 genotype on depression risk (CC genotype carriers who have low nodal strength in the right hippocampus show higher depression risk). |
| [213] | Genotype (several SNPs) | Patients with MDD (n = 218) and controls (n = 742) | Association between five FKBP5 SNPs (rs1360780, rs9470080, rs4713916, rs9296158, and rs9394309) and MDD. Two haplotype combinations were significantly more frequent in MDD than in controls. |
| [214] | Genotype (rs1360780) | Patients with a first primary cancer diagnosis (n = 7,320) | No association between FKBP5 genetic variant and use of antidepressants or hospital contact for depression after diagnosis of cancer. |
| [215] | Genotype (rs1360780, rs9296158, rs3800373, rs9470080) | N = 361 pregnant women | No association between FKBP5 genotypes or haplotypes with depressive symptoms during and after pregnancy. |
| [187] | Genotype (rs1360780) | Inpatients in a depressive episode (n = 68) and healthy controls (n = 87) | Significant interaction between MDD and T allele on GR-stimulated FKBP5 mRNA expression (reduced induction of FKBP5 mRNA in T carriers). GR resistance in T carrier patients. |
| [216] | Genotype (several SNPs) | N = 2928 participants with genetic data and information about depressive symptoms | Significant association between minor alleles of 4 FKBP5 SNPs (rs9470080, rs9394309, rs7748266, and rs1360780) with decreased cortisol area under the curve and increased risk of depressive symptoms. |
| [217] | Genotype (rs1360780) | N = 344 outpatients with chronic hepatitis C initiating IFN-alpha and ribavirin therapy | No association between FKBP5 genetic variants and IFN-induced depression. |
| [192] | Genotype (rs3800373, rs1360780, rs4713916, rs9296158, rs9470080) | N = 884 participants (community sample) | No genetic main effect on major depressive episode. Interactions between the five SNPs and traumatic (but not separation) events (especially severe trauma). Homozygosity for the minor allele of selected FKBP5 SNPs suggested as a risk genotype for the development of a major depressive episode in subjects with prior trauma exposure. |
| [218] | Genotype (rs1360780, rs4713916, rs3800373) | N = 271 children/adolescents whose mothers had experiences at least two major depressive episodes and N = 165 controls | No differences in the frequency of the genotypes between the two groups. No association between FKBP5 genotypes and child and adolescent depression scores. |
| [12] | Genotype (several SNPs) and protein levels in blood | N = 294 depressed individuals and N = 339 controls | Individuals with the TT genotype (rs1360780) experienced more than twice as many depressive episodes before the index episode than the other genotypes. |
| [219] | Genotype (rs1360780, rs3800373, rs9296158, rs9470080) | N = 131 adult patients who have received a kidney transplant | FKBP5 rare alleles at three out of four SNPs in FKBP5 (rs1360780, rs9296158, and rs9470080) were associated with increased depressive symptoms. |
| [220] | Genotype (rs3800373) | N = 106 school-aged children | Attachment security was negatively associated with depressive symptoms among children with two minor alleles of the FKBP5 SNP. Maternal attachment anxiety was positively associated with depressive symptoms in these children. |
| [221] | Genotype (rs1360780) | N = 311 physically healthy subjects | Dysfunctional attitudes predisposing to depression were significantly higher in the group with the T-allele than in that without this allele (particularly in achievement and self-control). |
| [189] | Genotype (rs1360780) | Adult patients with MDD (n = 40) and healthy controls (n = 20) | T-allele carriers had a later onset of disease compared with CC homozygous patients. Patients expressing the T-allele exhibited functional and structural differences in areas involved in emotional perception and inhibition. Interaction between the risk allele and higher CTQ scores mediates structural alterations. |
| [222] | Genotype (rs1360780, rs4713916) | Depressed unrelated inpatients with a major depressive episode (n = 412) and controls (n = 634) | No difference in FKBP5 genotypes between groups. |
| [223] | Genotype (rs1360780) | Patients with depression (n = 457) and healthy controls (n = 2286) | T-allele and TT genotype were overrepresented in depression for men. |
| [176] | Genotype (rs7757037, rs1360780, rs4713902) | Patients with MDD (n = 657) and healthy controls (n = 462) | T-allele of rs1360780 was more frequent among patients with MDD with a comorbidity of anxiety disorders, compared to those without. |
| [193] | Genotype (rs1360780) | N = 2157 Caucasian subjects | Significant interaction of physical abuse with the TT genotype of rs1360780 was found to increase depressive symptoms. |
| [194] | Genotype (rs3800373, rs9296158, rs1360870, rs9470080) | N = 236 high-risk, low-income women | FKBP5 moderation of the indirect effects of maltreatment on depression and dissociation via limbic irritability. |
| [224] | Genotype (rs1360780) | N = 246 Alzheimer’s disease patients with or without major depressive disorder | Significant association of FKBP5 with MDD in Alzheimer’s disease. C-allele was associated with a higher risk of depression. |
| [195] | Genotype (rs1360780, rs4713916, rs3800373, rs755658) | Meta-analysis with a total sample size of 26,582, including 14,491 MDD patients and 14,091 controls | No significant association between any of the FKBP5 SNPs and MDD susceptibility when all samples were pooled. After removing one heterogeneous study, both the rs1360780 T-allele and the rs3800373 C-allele were significantly associated with MDD. |
| [225] | Genotype (rs1360780, rs3800373, rs9470080) | Meta-analysis with a total sample of 15,109 participants. | Individuals who carry T-allele of rs1360780, C-allele of rs3800373 or T-allele of rs9470080 exposed to early-life trauma had higher risks for depression or PTSD. |
| Ref. | FKBP5 Measure | Sample | Finding |
|---|---|---|---|
| [196] | mRNA levels in blood | Patients with MDD (n = 74) and healthy controls (n = 34) | Association between successful antidepressant response and reduction of FKBP5 mRNA levels after 8 weeks of treatment with escitalopram or nortriptyline. |
| [202] | Genotype (rs1360780, rs3800373) | N = 246 geriatric patients with depression | No association between FKBP5 and clinical outcomes after 8 weeks of treatment with paroxetine and mirtazapine. |
| [226] | Genotype (rs1360780) | Depressive outpatients (n = 159) and controls (n = 96) | T allele carriers showed a 2.10 increased risk for non-responding at 4th week to 12-week citalopram treatment (almost significant tendency). |
| [197] | Genoytpe (rs3800373, rs1360780) | N = 304 depressive inpatients with unipolar or bipolar depression | Carriers of the FKBP5 variants had a trend toward a higher chance to response (mainly in those treated with antidepressant drug combinations or with venlafaxine) |
| [203] | Genotype (rs1360780) | N = 125 outpatients with major depression (n = 119) or dysthymic disorders (n = 6) | No association between rs1360780 and short-term antidepressant treatment response (fluoxetine; 20 mg/day) or lifetime depressive episodes. |
| [12] | Genotype (several SNPs) | N = 233 depressed inpatients | Association between three SNPs in FKBP5 (rs1360780, rs1334894 and rs755658) with antidepressant response. In a replication sample, rs1360780 showed a significant association and rs3800373 showed a trend for an association with response to antidepressants. |
| [198] | Genotype (rs1360780) | N = 1953 STAR*D outpatients and N = 275 MARS in patients | Interaction between the TT genotype of FKBP5 rs1360780 and the GG genotype of GRIK4 (rs12800734) in the prediction of antidepressant response. |
| [199] | Genotype (rs1360780) | N = 298 inpatients with MDD | Interaction between FKBP5 genotype at rs1360780 and treatment mode. C-allele carriers had a significantly worse outcome when treated naturalistically. TT-genotype subjects showed a superior treatment response across both SSTR and TAU treatment conditions. |
| [200] | Genotype (several SNPs) | N = 529 MDD patients from the Mayo clinic and n = 96 from the STAR*D study | Association between the SNP rs352428 with 8-week SSRI treatment response in the Mayo study and 6-week treatment response in the STAR*D replication study. |
| [201] | Genotype (rs1360780, rs4713916) | Outpatients with non-psychotic MDD (n = 1809) and controls (n = 729) | rs1360780 was significantly associated with MDD in White non-Hispanics. Significant association between rs4713916 and remission. |
| [204] | Genotype (rs1360780, rs3800373) | Meta-analysis with a total sample of 2,194 subjects for rs1360780 and 2,049 for rs3800373 | rs1360780: In Caucasians T-allele carriers showed a marginal evidence of better response, while in the other/mixed ethnic subgroup a better response was seen in C/C homozygous subjects. rs3800373: In Caucasians C-allele carriers showed a trend of better response. |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fries, G.R.; Gassen, N.C.; Rein, T. The FKBP51 Glucocorticoid Receptor Co-Chaperone: Regulation, Function, and Implications in Health and Disease. Int. J. Mol. Sci. 2017, 18, 2614. https://doi.org/10.3390/ijms18122614
Fries GR, Gassen NC, Rein T. The FKBP51 Glucocorticoid Receptor Co-Chaperone: Regulation, Function, and Implications in Health and Disease. International Journal of Molecular Sciences. 2017; 18(12):2614. https://doi.org/10.3390/ijms18122614
Chicago/Turabian StyleFries, Gabriel R., Nils C. Gassen, and Theo Rein. 2017. "The FKBP51 Glucocorticoid Receptor Co-Chaperone: Regulation, Function, and Implications in Health and Disease" International Journal of Molecular Sciences 18, no. 12: 2614. https://doi.org/10.3390/ijms18122614
APA StyleFries, G. R., Gassen, N. C., & Rein, T. (2017). The FKBP51 Glucocorticoid Receptor Co-Chaperone: Regulation, Function, and Implications in Health and Disease. International Journal of Molecular Sciences, 18(12), 2614. https://doi.org/10.3390/ijms18122614