Aquaporins during Pregnancy: Their Function and Significance


Abstract
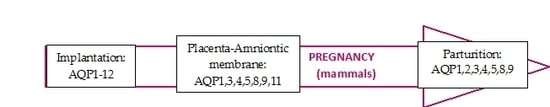
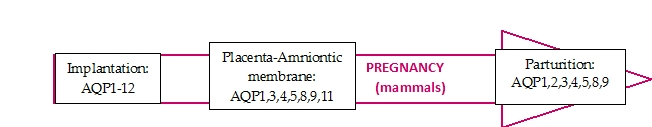
1. Introduction
2. Implantation
3. Maternal–Fetal Fluid Flow
3.1. Amniotic Membrane
3.2. Placenta
4. Parturition
4.1. Myometrial Contraction
4.2. Cervical Ripening
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ishibashi, K.; Kondo, S.; Hara, S.; Morishita, Y. The evolutionary aspects of aquaporin family. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R566–R576. [Google Scholar] [CrossRef] [PubMed]
- Madeira, A.; Moura, T.F.; Soveral, G. Aquaglyceroporins: Implications in adipose biology and obesity. Cell. Mol. Life Sci. 2015, 72, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Hara-Chikuma, M.; Verkman, A.S. Physiological roles of glycerol-transporting aquaporins: The aquaglyceroporins. Cell. Mol. Life Sci. 2006, 63, 1386–1392. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, R.; Bhattacharjee, H.; Rosen, B.P. Aquaglyceroporins: Generalized metalloid channels. Biochim. Biophys. Acta 2014, 1840, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Yakata, K.; Hiroaki, Y.; Ishibashi, K.; Sohara, E.; Sasaki, S.; Mitsuoka, K. Aquaporin-11 containing a divergent NPA motif has normal water channel activity. Biochim. Biophys. Acta Biomembr. 2007, 1768, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Rai, T.; Kuwahara, M.; Ko, S.B.H.; Uchida, S.; Sasaki, S.; Ishibashi, K. Identification of a novel aquaporin, AQP12, expressed in pancreatic acinar cells. Biochem. Biophys. Res. Commun. 2005, 330, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tan, Y.-J.; Qu, F.; Sheng, J.-Z.; Huang, H.-F. Functions of water channels in male and female reproductive systems. Mol. Aspects Med. 2012, 33, 676–690. [Google Scholar] [CrossRef] [PubMed]
- Tornroth-Horsefield, S.; Hedfalk, K.; Fischer, K.; Lindkvist-Petersson, G.; Neutze, R. Structural insights into eukaryotic aquaporin regulation. FEBS Lett. 2010, 584, 2580–2588. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, U.K.; Fischer, G.; Friemann, R.; Enkavi, G.; Tajkhorshid, E.; Neutze, R. Subangstrom resolution X-ray structure details aquaporin-water interactions. Science 2013, 340, 1346–1349. [Google Scholar] [CrossRef] [PubMed]
- Enders, A.; Schlafke, S. A morphological analysis of the early implantation stages in the rat. Am. J. Anat. 1967, 120, 185–225. [Google Scholar] [CrossRef]
- Lindsay, L.A.; Murphy, C.R. Aquaporin-1 increases in the rat myometrium during early pregnancy. J. Mol. Histol. 2004, 35, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, L.A.; Murphy, C.R. Redistribution of aquaporins 1 and 5 in the rat uterus is dependent on progesterone: A study with light and electron microscopy. Reproduction 2006, 131, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Jiang, W.; Zhang, W.; Shen, Q.; Chen, M.; Zhu, X. Expression and significance of aquaporins during pregnancy. Front. Biosci. 2013, 18, 1373–1383. [Google Scholar]
- He, R.H.; Sheng, J.Z.; Luo, Q.; Jin, F.; Wang, B.; Qian, Y.L.; Zhou, C.Y.; Sheng, X.; Huang, H.F. Aquaporin-2 expression in human endometrium correlates with serum ovarian steroid hormones. Life Sci. 2006, 79, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Wray, S.; Marples, D. Distribution of AQP2 and AQP3 water channels in human tissue microarrays. J. Mol. Histol. 2005, 36, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sponchiado, M.; Gomes, N.S.; Fontes, P.K.; Martins, T.; Del Collado, M.; Pastore, A.A.; Pugliesi, G.; Nogueira, M.F.G.; Binelli, M. Pre-hatching embryo-dependent and -independent programming of endometrial function in cattle. PLoS ONE 2017, 12, e0175954. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.L.; Zhang, J.; Fan, Y.; Ding, J.H.; Sha, J.H.; Hu, G. Aquaporin-4 deficiency induces subfertility in female mice. Fertil. Steril. 2009, 92, 1736–1743. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, L.A.; Murphy, C.R. Redistribution of aquaporins in uterine epithelial cells at the time of implantation in the rat. Acta Histochem. 2004, 106, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, L.A.; Murphy, C.R. Aquaporins are upregulated in glandular epithelium at the time of implantation in the rat. J. Mol. Histol. 2007, 38, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Richard, C.; Gao, J.; Brown, N.; Reese, J. Aquaporin water channel genes are differentially expressed and regulated by ovarian steroids during the periimplantation period in the mouse. Endocrinology 2003, 144, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Qiao, Y.; Yi, F.; Guan, X.; Zhang, D.; Zhang, S.; Hao, F.; Xiao, Y.; Zhang, H.; Guo, L.; et al. Increased female fertility in aquaporin 8-deficient mice. IUBMB Life 2010, 62, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jee, B.C.; Kim, S.K.; Kim, H.; Lee, J.R.; Suh, C.S.; Kim, S.H. Expressions of aquaporin family in human luteinized granulosa cells and their correlations with IVF outcomes. Hum. Reprod. 2016, 31, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.F.; Chen, L.Y.; Xu, K.H.; Yao, J.F.; Shi, Y.F.; Shanguan, X.J. Reduced expression of aquaporin 9 in tubal ectopic pregnancy. J. Mol. Histol. 2013, 44, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, Q.; Zhang, H.; Wang, Q.; Li, R.; Jin, Y.; Wang, H.; Ma, T.; Qiao, J.; Duan, E. Aquaporin-dependent excessive intrauterine fluid accumulation is a major contributor in hyper-estrogen induced aberrant embryo implantation. Cell Res. 2015, 25, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, H.; Giribabu, N.; Karim, K.; Kassim, N.; Muniandy, S.; Kumar, K.E.; Salleh, N. Quercetin interferes with the fluid volume and receptivity development of the uterus in rats during the peri-implantation period. Reprod. Toxicol. 2017, 71, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Marunaka, Y. Actions of quercetin, a flavonoid, on ion transporters: Its physiological roles. Ann. N. Y. Acad. Sci. 2017, 1398, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Skowronska, A.; Mlotkowska, P.; Majewski, M.; Nielsen, S.; Skowronski, M.T. Expression of aquaporin 1 and 5 and their regulation by ovarian hormones, arachidonic acid, forskolin and cAMP during implantation in pigs. Physiol. Res. 2016, 8, 637–650. [Google Scholar]
- Brace, R.A.; Vermin, M.L.; Huijssoon, E.I. Regulation of amniotic fluid volume: Intramembranous solute and volume fluxes in late gestation fetal sheep. Am. J. Obstet. Gynecol. 2004, 191, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Yasui, M. Cellular and subcellular localization of aquaporins 1, 3, 8, and 9 in amniotic membranes during pregnancy in mice. Cell Tissue Res. 2010, 342, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Prat, C.; Blanchon, L.; Borel, V.; Gallot, D.; Herbet, A.; Bouvier, D.; Marceau, G.; Sapin, V. Ontogeny of aquaporins in human fetal membranes. Biol. Reprod. 2012, 86, 48. [Google Scholar] [CrossRef] [PubMed]
- Sha, X.Y.; Xiong, Z.F.; Liu, H.S.; Di, X.D.; Ma, T.H. Maternal-fetal fluid balance and aquaporins: From molecule to physiology. Acta Pharmacol. Sin. 2011, 32, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Misaka, T.; Tanaka, Y.; Matsumoto, I.; Ishibashi, K.; Sasaki, S.; Abe, K. Aquaporin-11 knockout mice and polycystic kidney disease animals share a common mechanism of cyst formation. FASEB J. 2008, 22, 3672–3684. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, D.; Rouzaire, M.; Marceau, G.; Prat, C.; Pereira, B.; Lemarié, R.; Deruelle, P.; Fajardy, I.; Gallot, D.; Blanchon, L.; et al. Aquaporins and Fetal Membranes From Diabetic Parturient Women: Expression Abnormalities and Regulation by Insulin. J. Clin. Endocrinol. Metab. 2015, 100, E1270–E1279. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Koukoulas, I.; Ross, M.C.; Wang, S.; Wintour, E.M. Quantitative comparison of placental expression of three aquaporin genes. Placenta 2004, 25, 475–478. [Google Scholar] [CrossRef] [PubMed]
- De Falco, M.; Cobellis, L.; Torella, M.; Acone, G.; Varano, L.; Sellitti, A.; Ragucci, A.; Coppola, G.; Cassandro, R.; Laforgia, V.; et al. Down-regulation of aquaporin 4 in human placenta throughout pregnancy. In Vivo 2007, 21, 813–817. [Google Scholar] [PubMed]
- Escobar, J.; Gormaz, M.; Arduini, A.; Gosens, K.; Martinez, A.; Perales, A.; Escrig, R.; Tormos, E.; Roselló, M.; Orellana, C.; et al. Expression of aquaporins early in human pregnancy. Early Hum. Dev. 2012, 88, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Jiang, S.; Hu, Y.; Zheng, X.; Zou, S.; Wang, Y.; Zhu, X. The expression of aquaporin 8 and aquaporin 9 in fetal membranes and placenta in term pregnancies complicated by idiopathic polyhydramnios. Early Hum. Dev. 2010, 86, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.S.; Zhu, X.J.; Ding, S.D.; Wang, J.J.; Jiang, L.L.; Jiang, W.X.; Zhu, X.Q. Expression and localization of aquaporins 8 and 9 in term placenta with oligohydramnios. Reprod. Sci. 2012, 19, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Belkacemi, L.; Desai, M.; Beall, M.H.; Liu, Q.; Lin, J.T.; Nelson, D.M.; Ross, M.G. Early compensatory adaptations in maternal undernourished pregnancies in rats: Role of the aquaporins. J. Matern. Fetal Neonatal Med. 2011, 24, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Vilariño-García, T.; Pérez-Pérez, A.; Dietrich, V.; Guadix, P.; Dueñas, J.L.; Varone, C.L.; Damiano, A.E.; Sánchez-Margalet, V. Gynecol Endocrinol. Leptin upregulates aquaporin 9 expression in human placenta in vitro. Gynecol. Endocrinol. 2017, 23, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lin, L.; Lai, A. Expression and significance of aquaporin-2 and serum hormones in placenta of patients with preeclampsia. J. Obstet. Gynaecol. 2017, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Szpilbarg, N.; Castro-Parodi, M.; Reppetti, J.; Repetto, M.; Maskin, B.; Martinez, N.; Damiano, A.E. Placental programmed cell death: Insights into the role of aquaporins. Mol. Hum. Reprod. 2016, 22, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Ducza, E.; Seres, A.B.; Hajagos-Tóth, J.; Falkay, G.; Gáspár, R. Oxytocin regulates the expression of aquaporin 5 in the late-pregnant rat uterus. Mol. Reprod. Dev. 2014, 81, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Helguera, G.; Eghbali, M.; Sforza, D.; Minosyan, T.Y.; Toro, L.; Stefani, E. Changes in global gene expression in rat myometrium in transition from late pregnancy to parturition. Physiol. Genom. 2009, 8, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Csányi, A.; Bóta, J.; Falkay, G.; Gáspár, R.; Ducza, E. The Effects of Female Sexual Hormones on the Expression of Aquaporin 5 in the Late-Pregnant Rat Uterus. Int. J. Mol. Sci. 2016, 17, 1300. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Brown, N.; Mahendroo, M.S.; Reese, J. Utilization of Different Aquaporin Water Channels in the Mouse Cervix during Pregnancy and Parturition and in Models of Preterm and Delayed Cervical Ripening. Endocrinology 2006, 147, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Soh, Y.M.; Tiwari, A.; Mahendroo, M.; Conrad, K.P.; Parry, L.J. Relaxin regulates hyaluronan synthesis and aquaporins in the cervix of late pregnant mice. Endocrinology 2012, 153, 6054–6064. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Sites of Expression | AQP1 | AQP2 | AQP3 | AQP4 | AQP5 | AQP6 | AQP7 | AQP8 | AQP9 | AQP11 | AQP12 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Oocytes/blastocytes (implantation) | h-gc-R | h-gc-R | h-gc-R | h-gc-R | h-gc-R | h-gc-R | h-gc-R | m-icm-P | h-gc-R m-mt-P | h-gc-R | h-gc-R |
| Uterus (implantation) | r-mm-P m-mm-P+R p-P | h-em-P+R | h-em-P+R | m-mm-P+R c-em-R | m-mm-P+R r-mm-P p-P | - | - | m-mm-P | r-P+R p-P | - | - |
| Amnionic membrane | h-P+R o-P+R m-fc-P | - | h-P+R o-P+R m-ec, fc-P | - | - | - | - | h-P+R o-P+R m-ec-P | h-P+R o-P+R m-aec-P | h-P+R o-P+R | - |
| Placenta | h-pv-R r-bz, lz-P m-vec-P+R o-vec-P+R | - | h-pv-R r-R m-tec-P+R o-tec-P+R | h-pv-R h-st, ec-P | h-pv-R | - | - | h-pv-R r-bz, lz-P m-P+R o-tec-R | h-pv-R r-cp, bz, lz-P m-R | h-pv-R | - |
| Uterus (late pregnant) | r-P+R | r-P+R | r-P+R | r-P+R | - | - | r-P+R | r-P+R | - | - | |
| Cervix (late pregnant) | - | - | m-bcl-P+R | m-acl P+R | m-acl P+R | - | - | m-acl P+R | - | - | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ducza, E.; Csányi, A.; Gáspár, R. Aquaporins during Pregnancy: Their Function and Significance. Int. J. Mol. Sci. 2017, 18, 2593. https://doi.org/10.3390/ijms18122593
Ducza E, Csányi A, Gáspár R. Aquaporins during Pregnancy: Their Function and Significance. International Journal of Molecular Sciences. 2017; 18(12):2593. https://doi.org/10.3390/ijms18122593
Chicago/Turabian StyleDucza, Eszter, Adrienn Csányi, and Róbert Gáspár. 2017. "Aquaporins during Pregnancy: Their Function and Significance" International Journal of Molecular Sciences 18, no. 12: 2593. https://doi.org/10.3390/ijms18122593
APA StyleDucza, E., Csányi, A., & Gáspár, R. (2017). Aquaporins during Pregnancy: Their Function and Significance. International Journal of Molecular Sciences, 18(12), 2593. https://doi.org/10.3390/ijms18122593