Protective Effects of Red Ginseng Oil against Aβ25–35-Induced Neuronal Apoptosis and Inflammation in PC12 Cells


Abstract
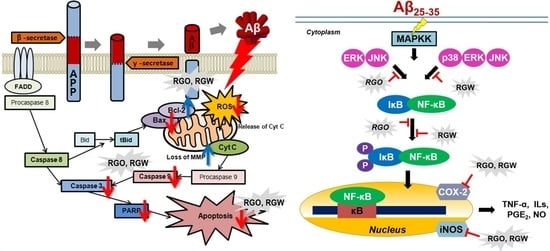
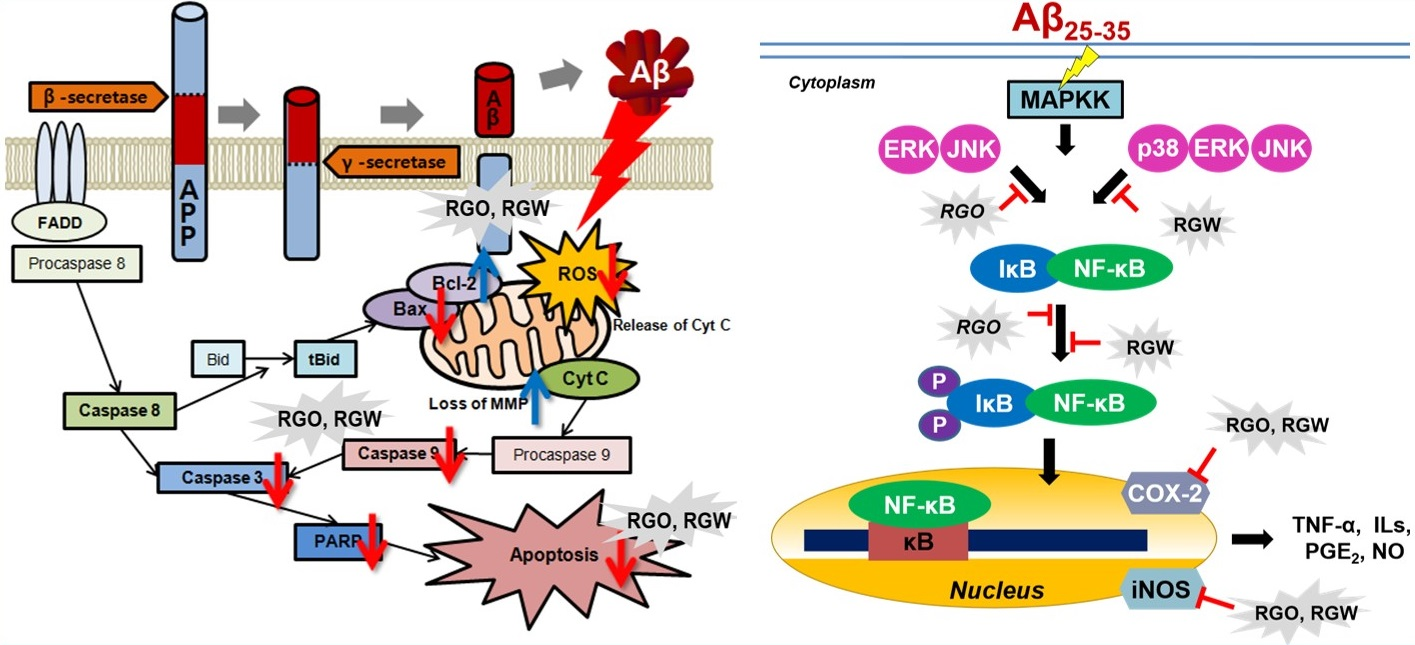
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
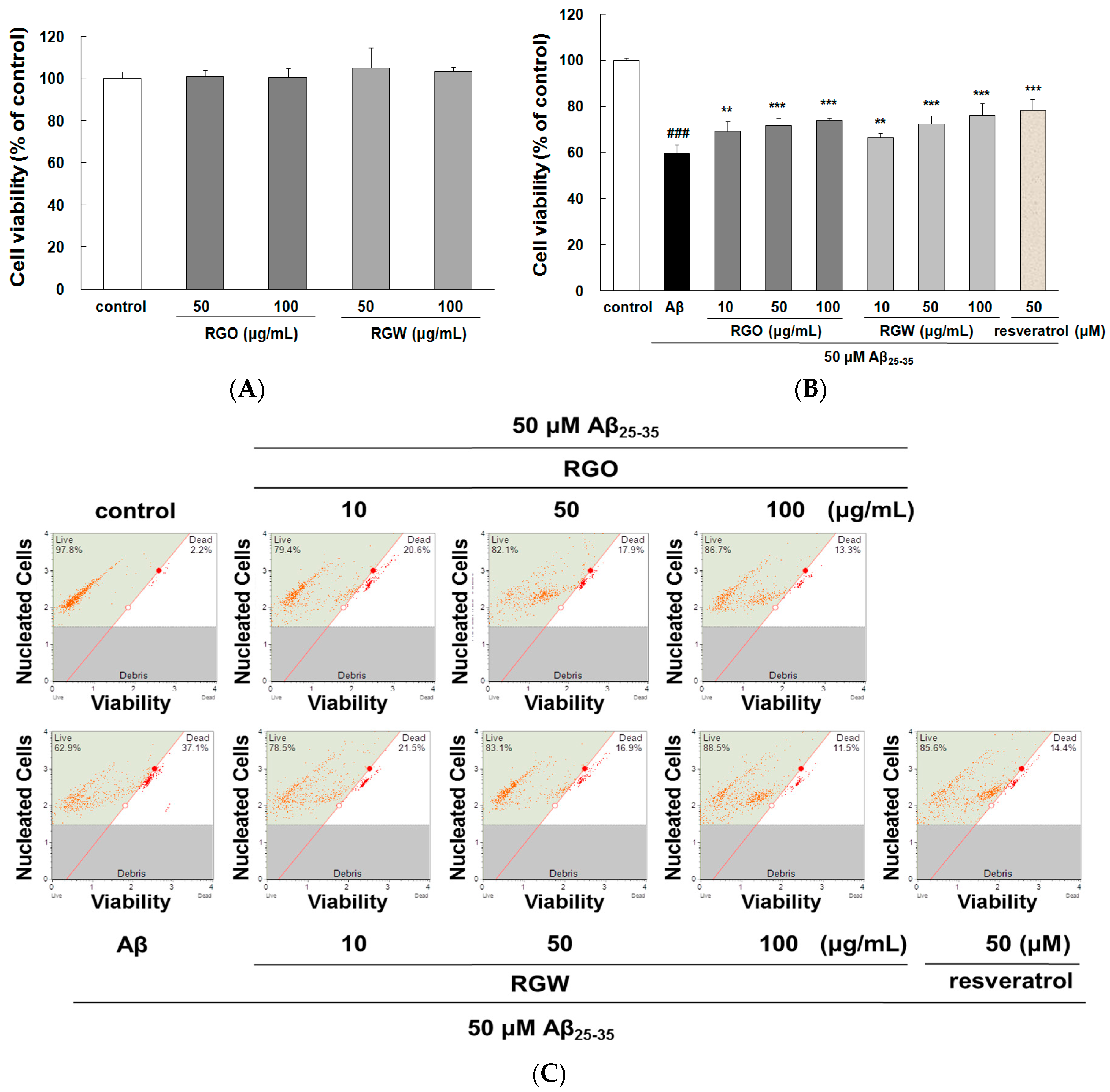
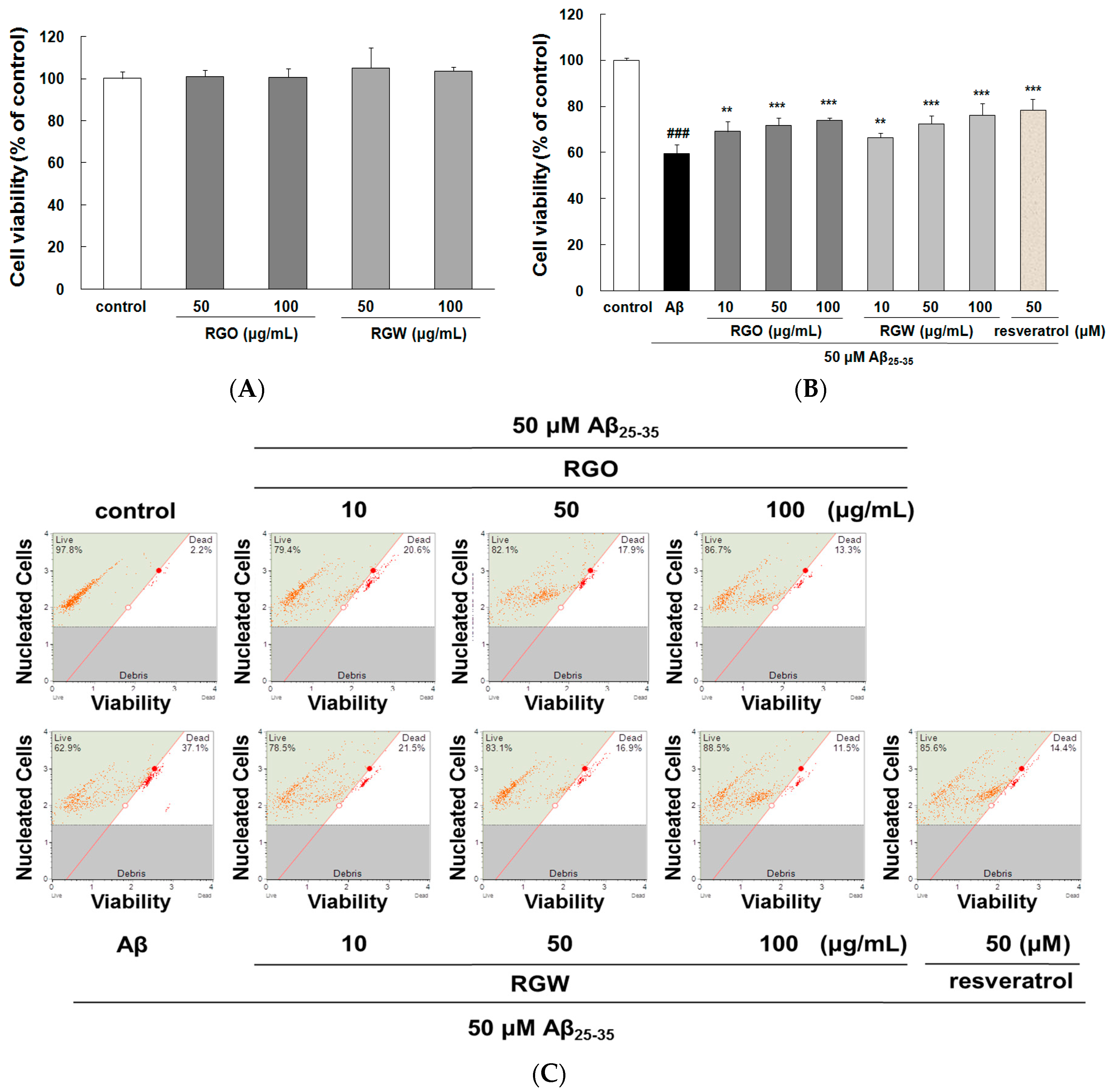
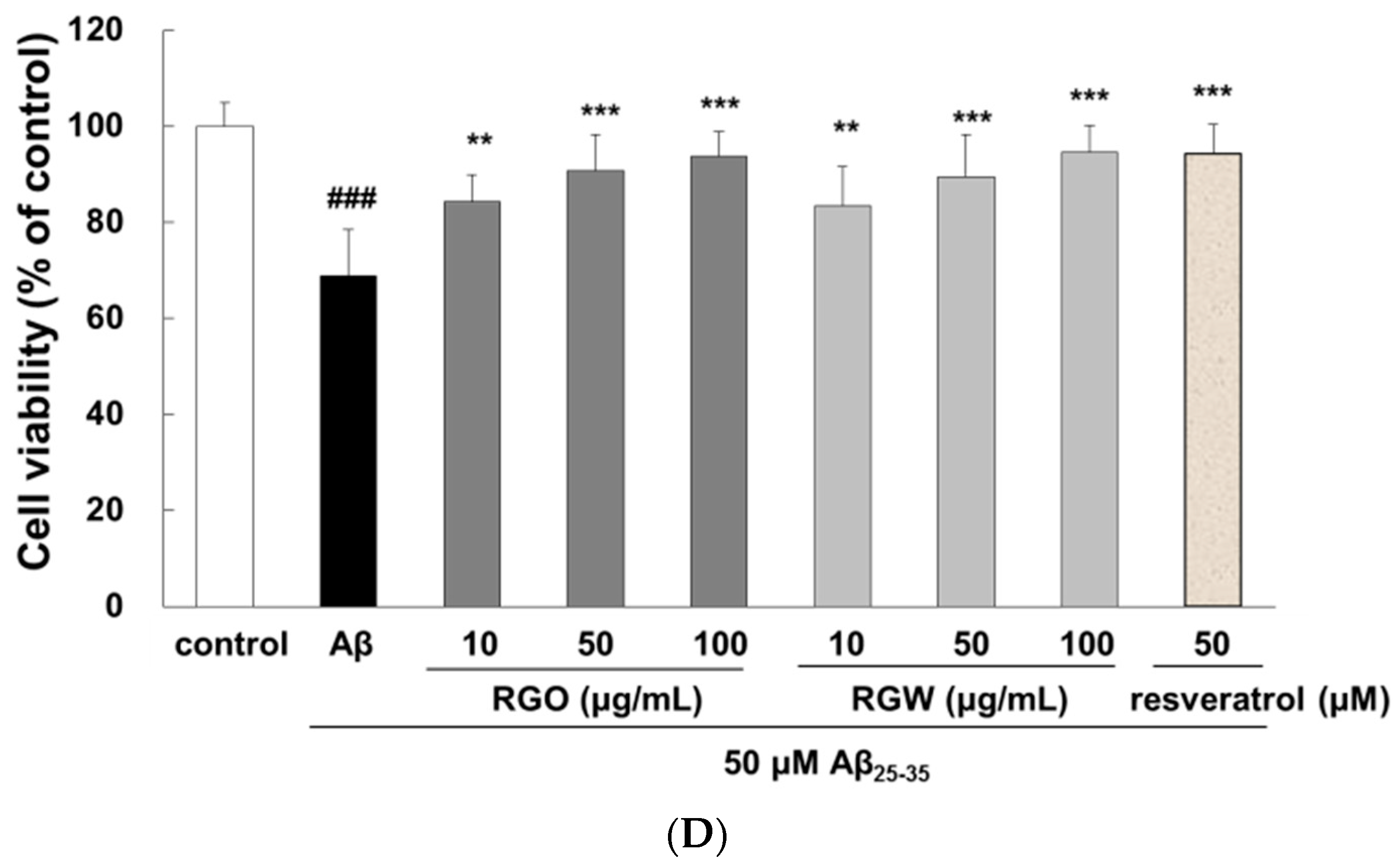
2.1. RGO on Aβ25–35-Stimulated PC12 Cell Death
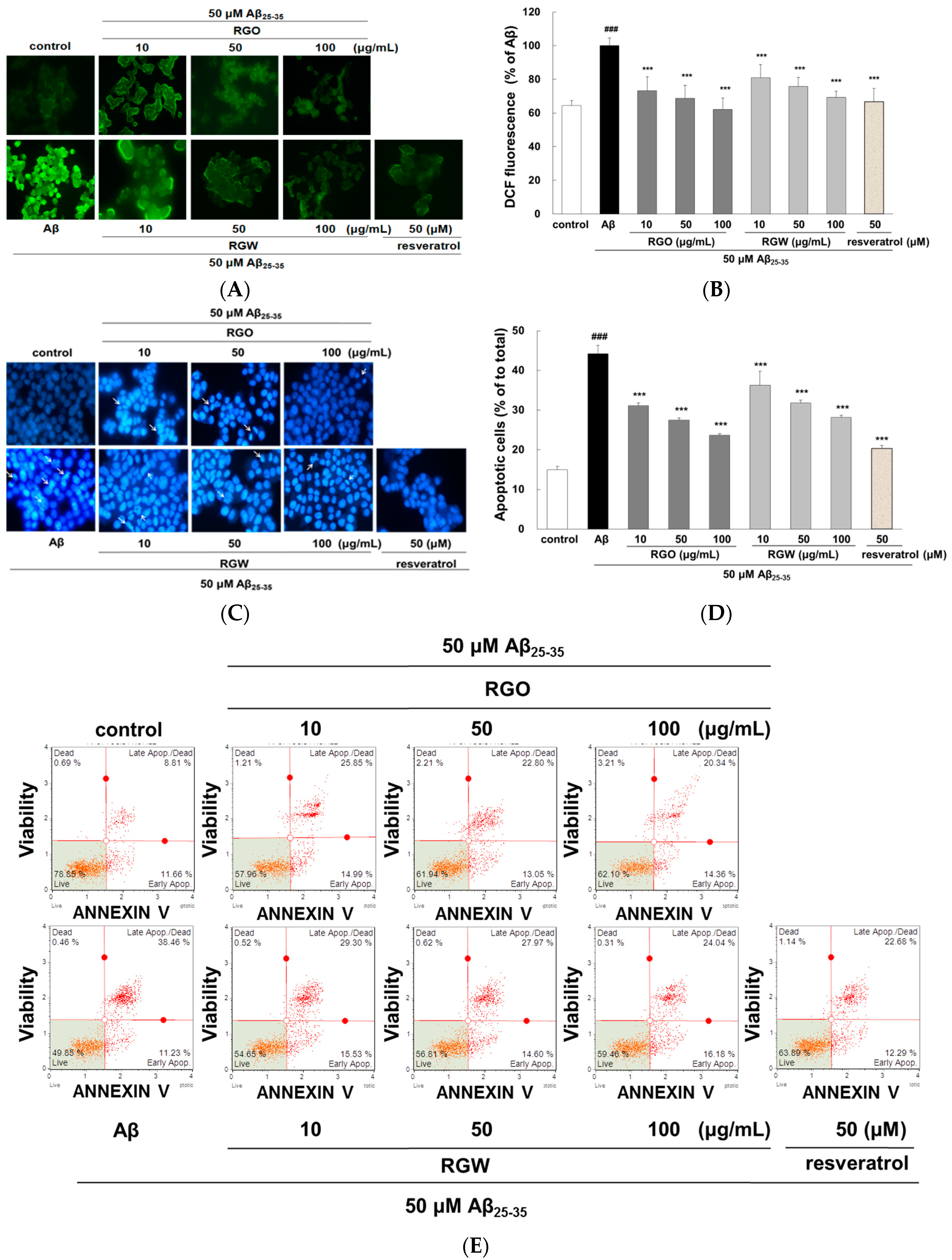
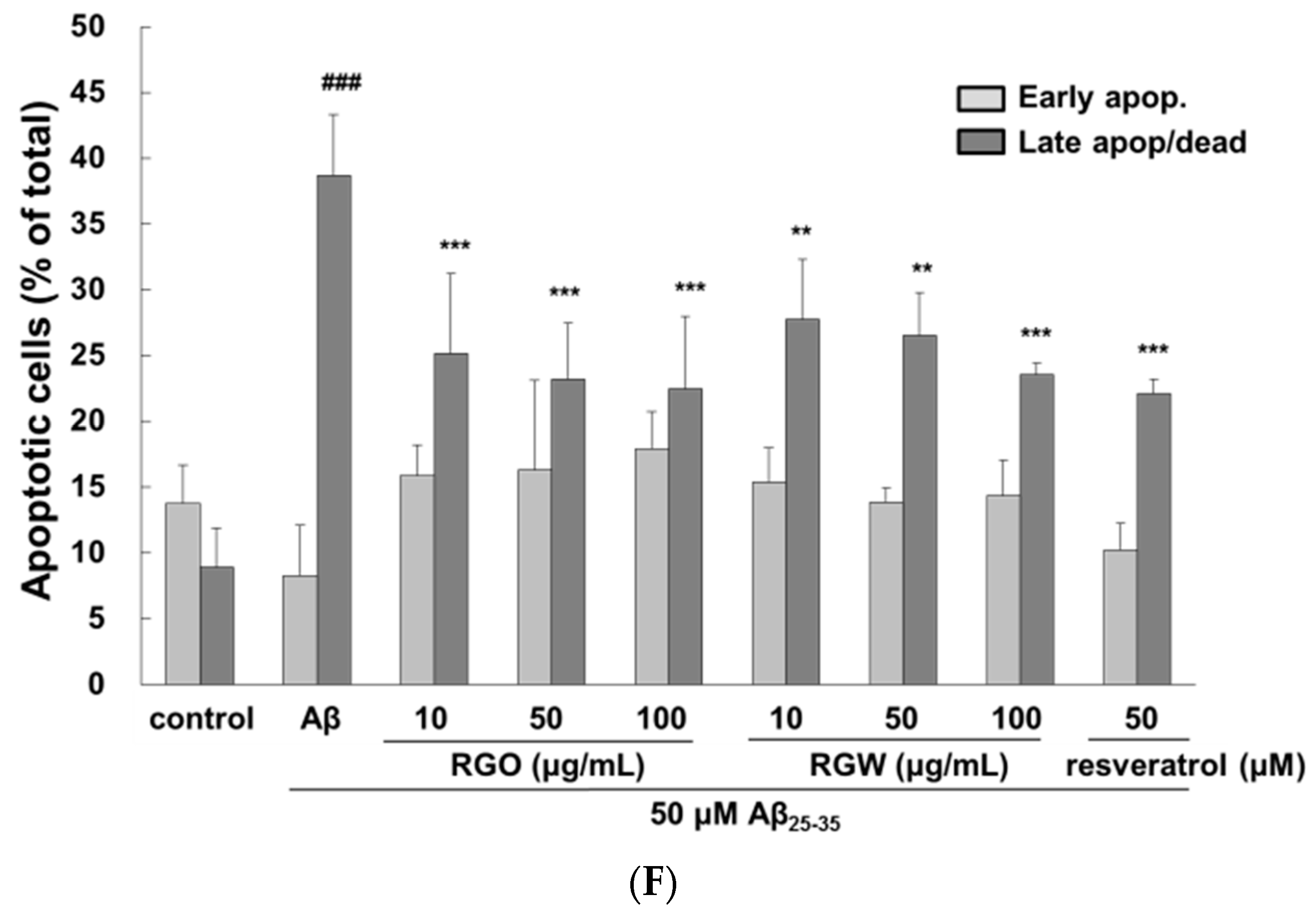
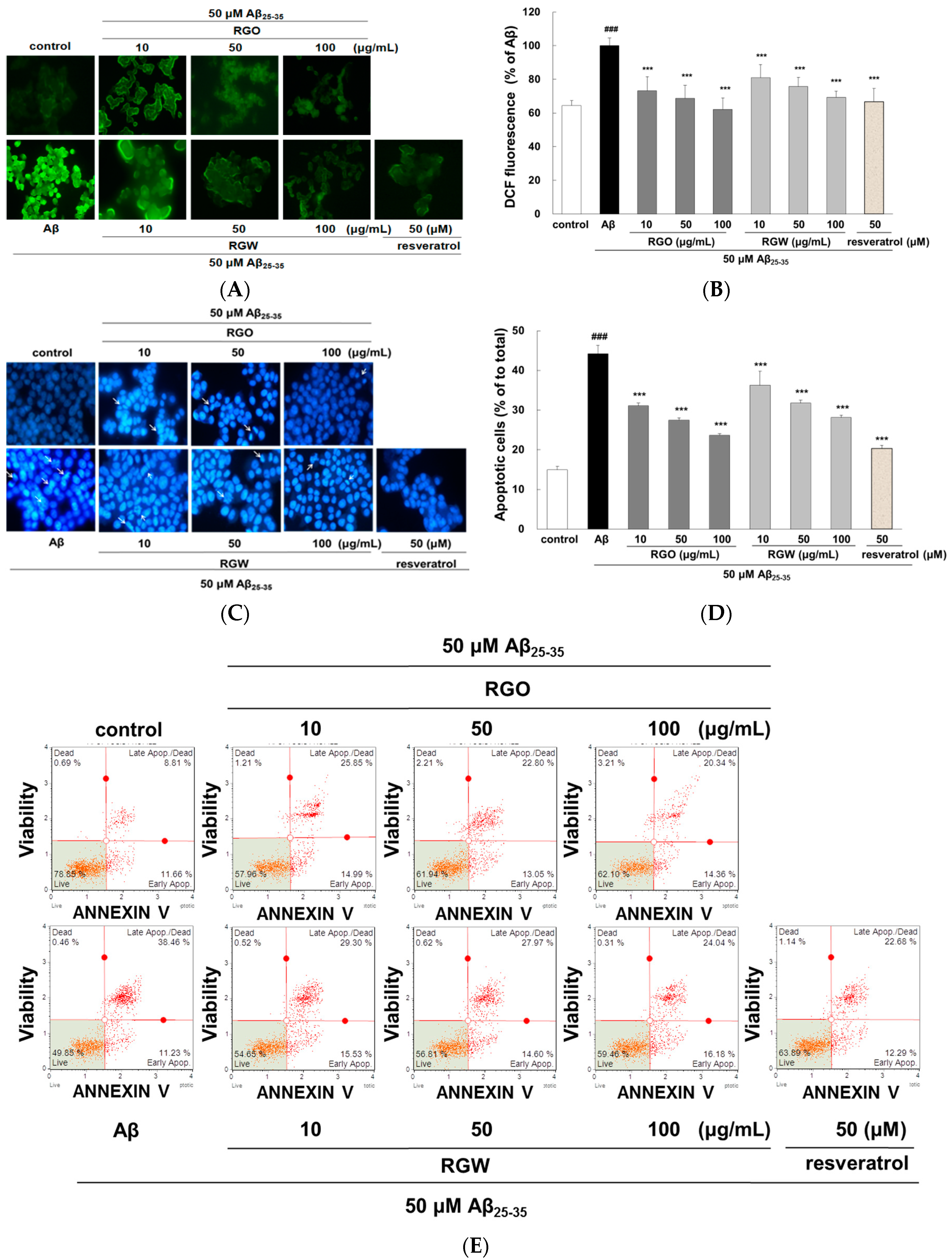
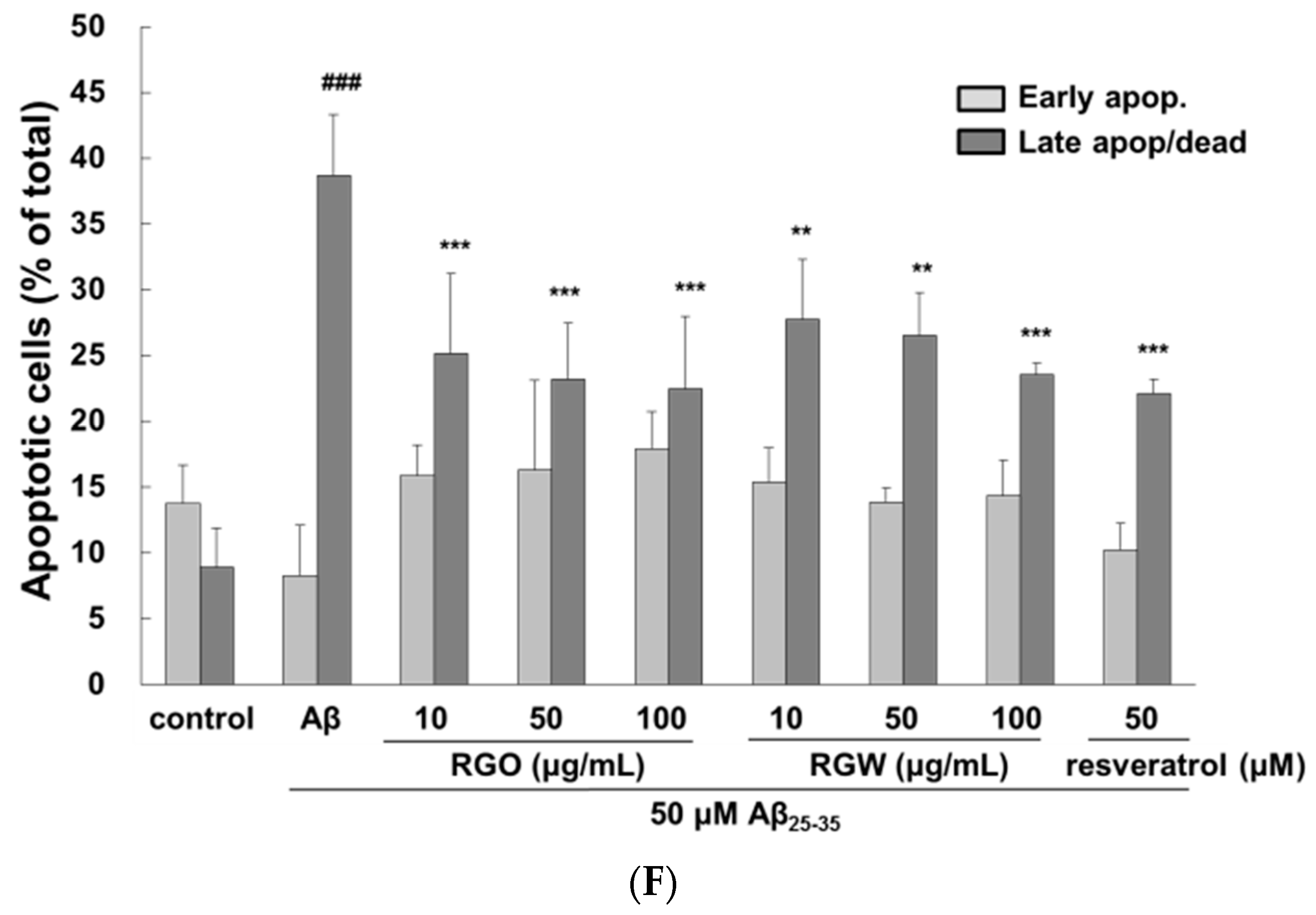
2.2. RGO on Aβ25–35-Stimulated Intracellular Reactive Oxygen Species (ROS) and Apoptosis
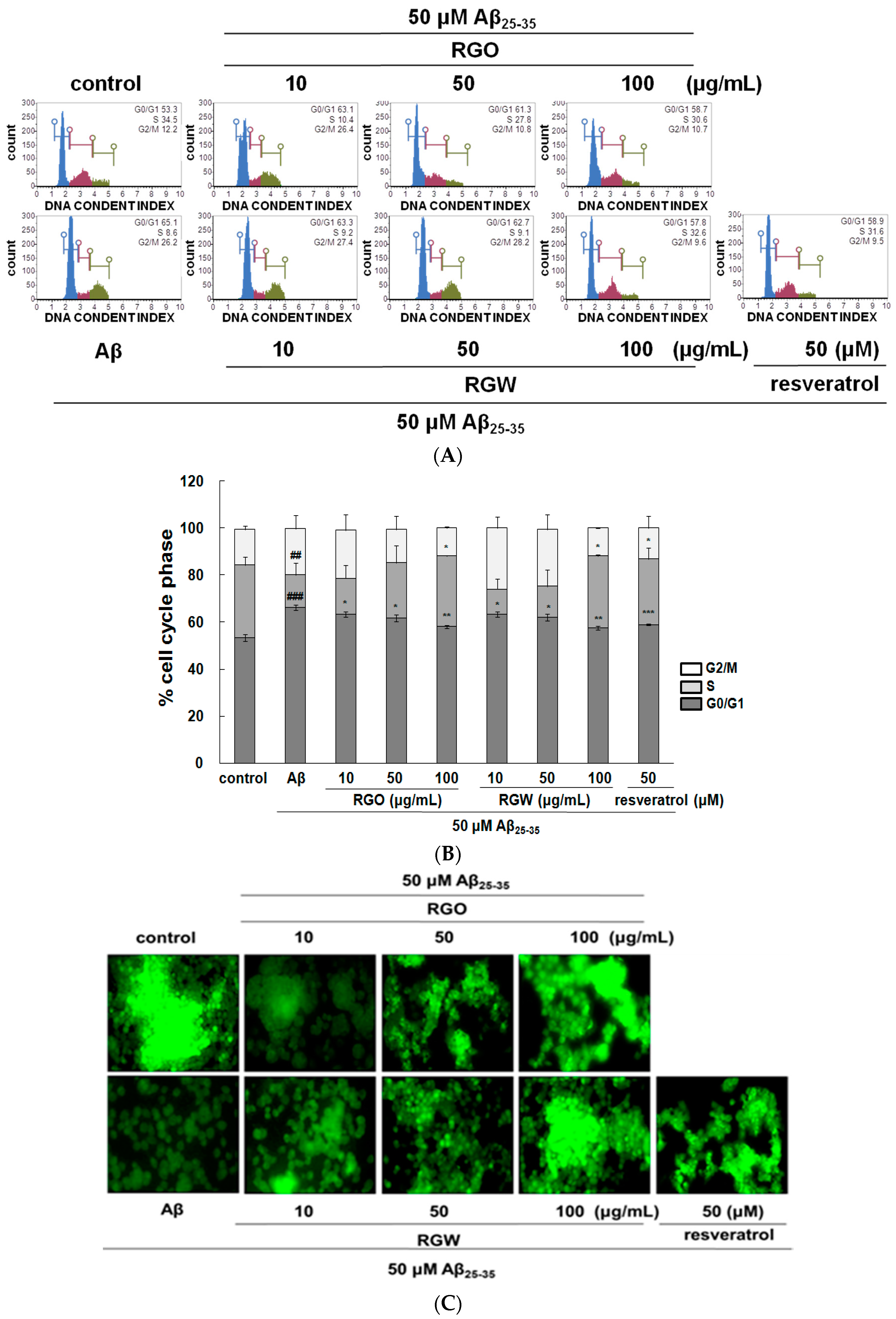
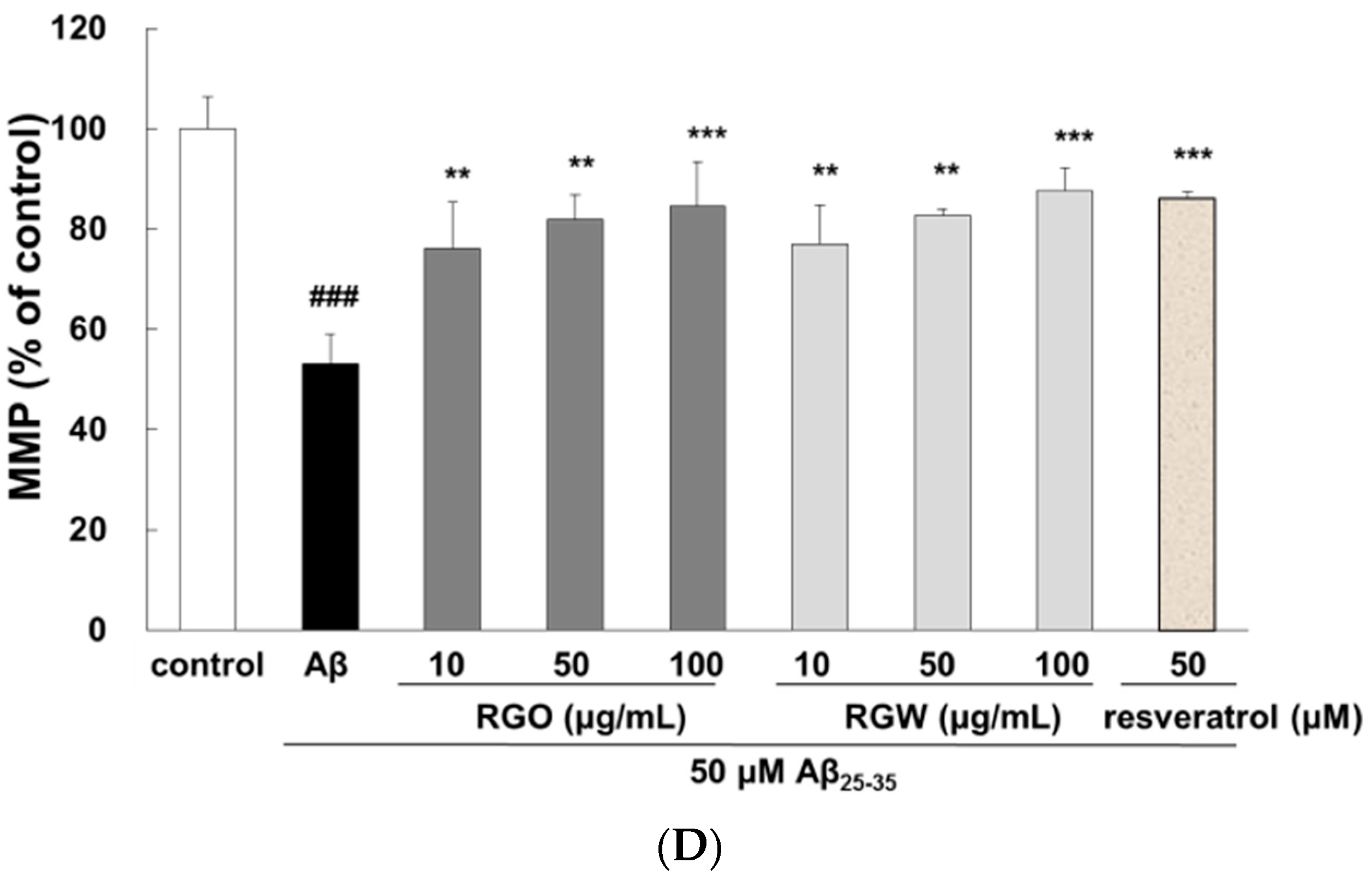
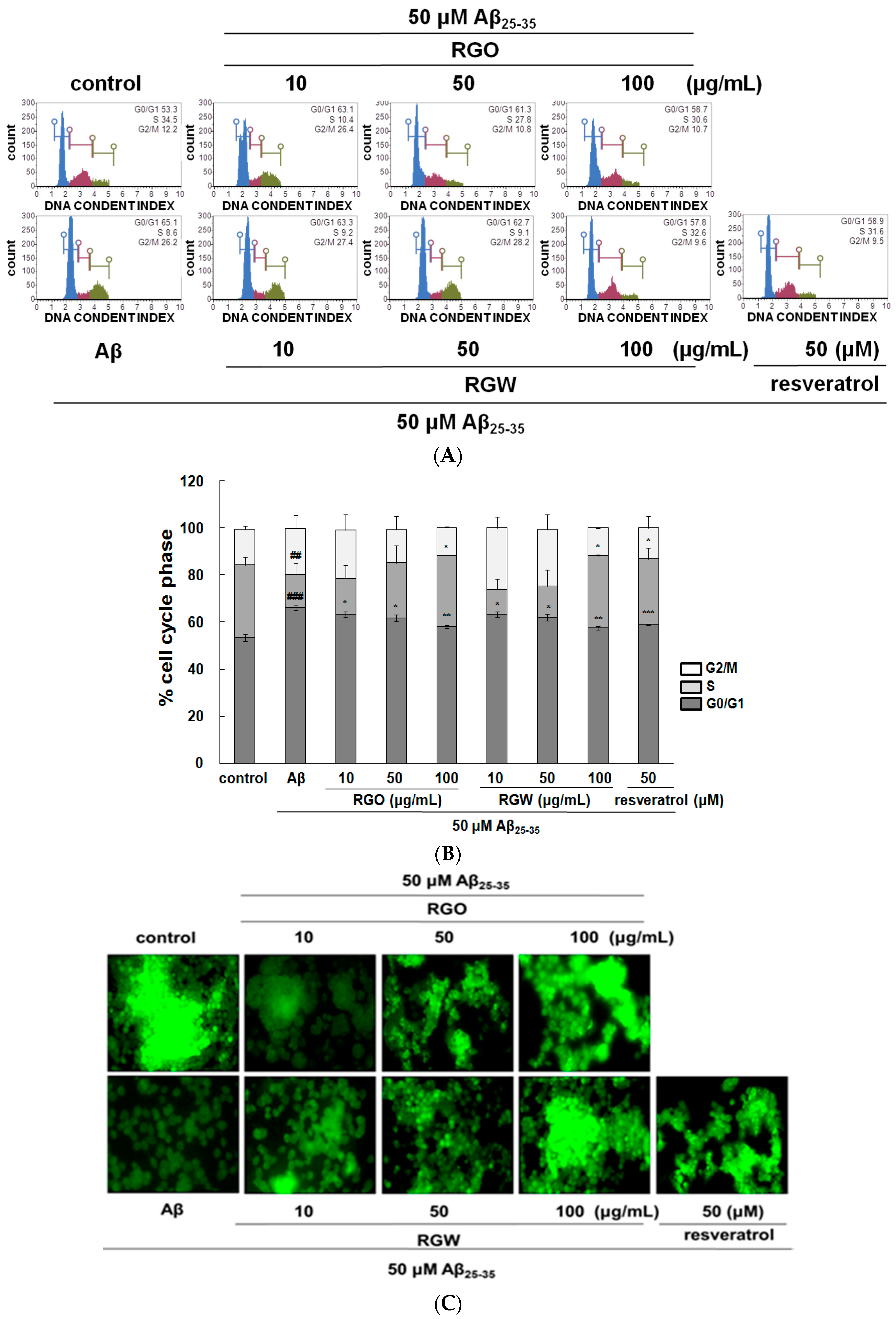
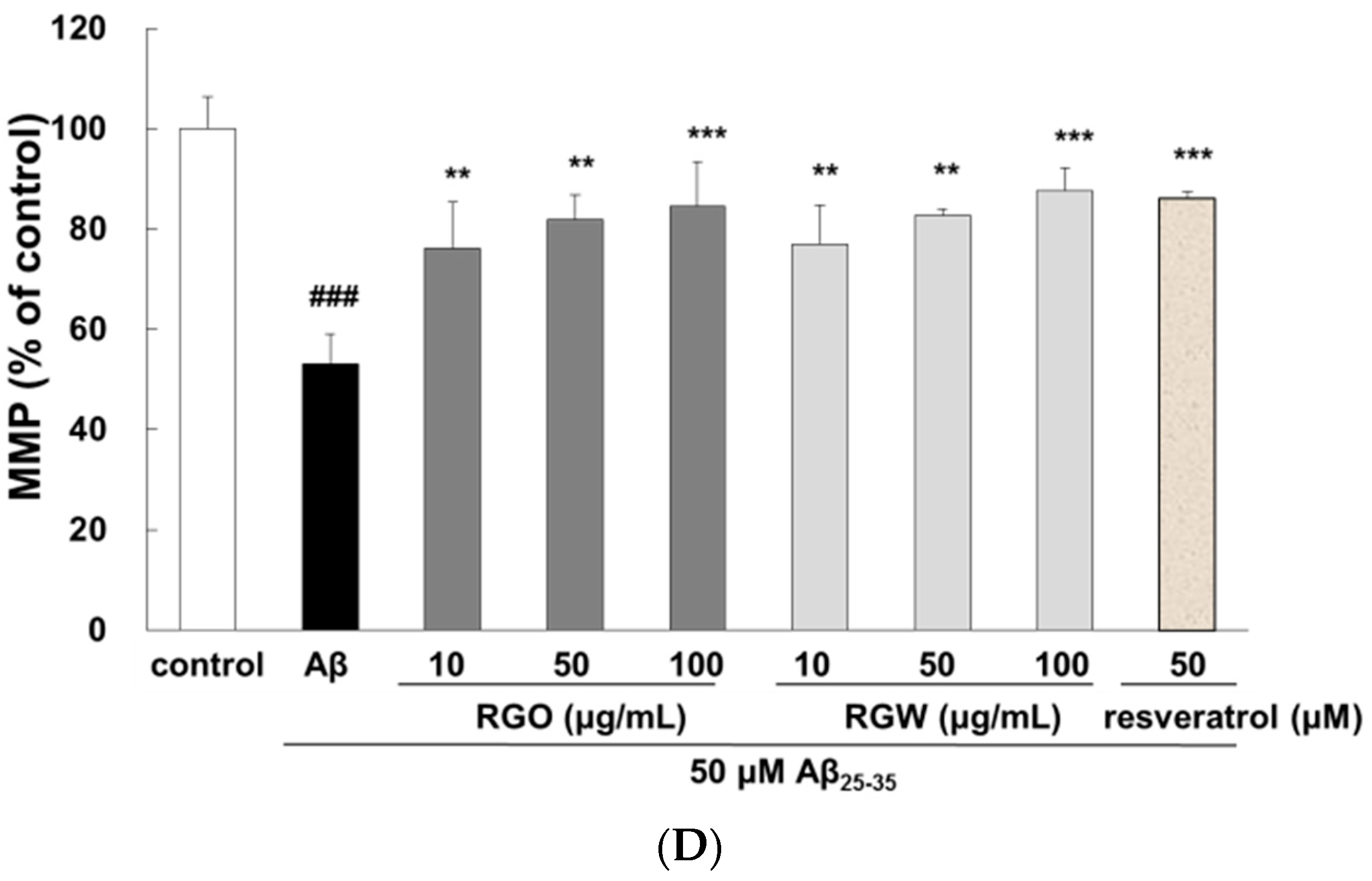
2.3. RGO on Aβ25–35-Stimulated Cell Cycle Changes and Mitochondrial Membrane Potential (MMP)
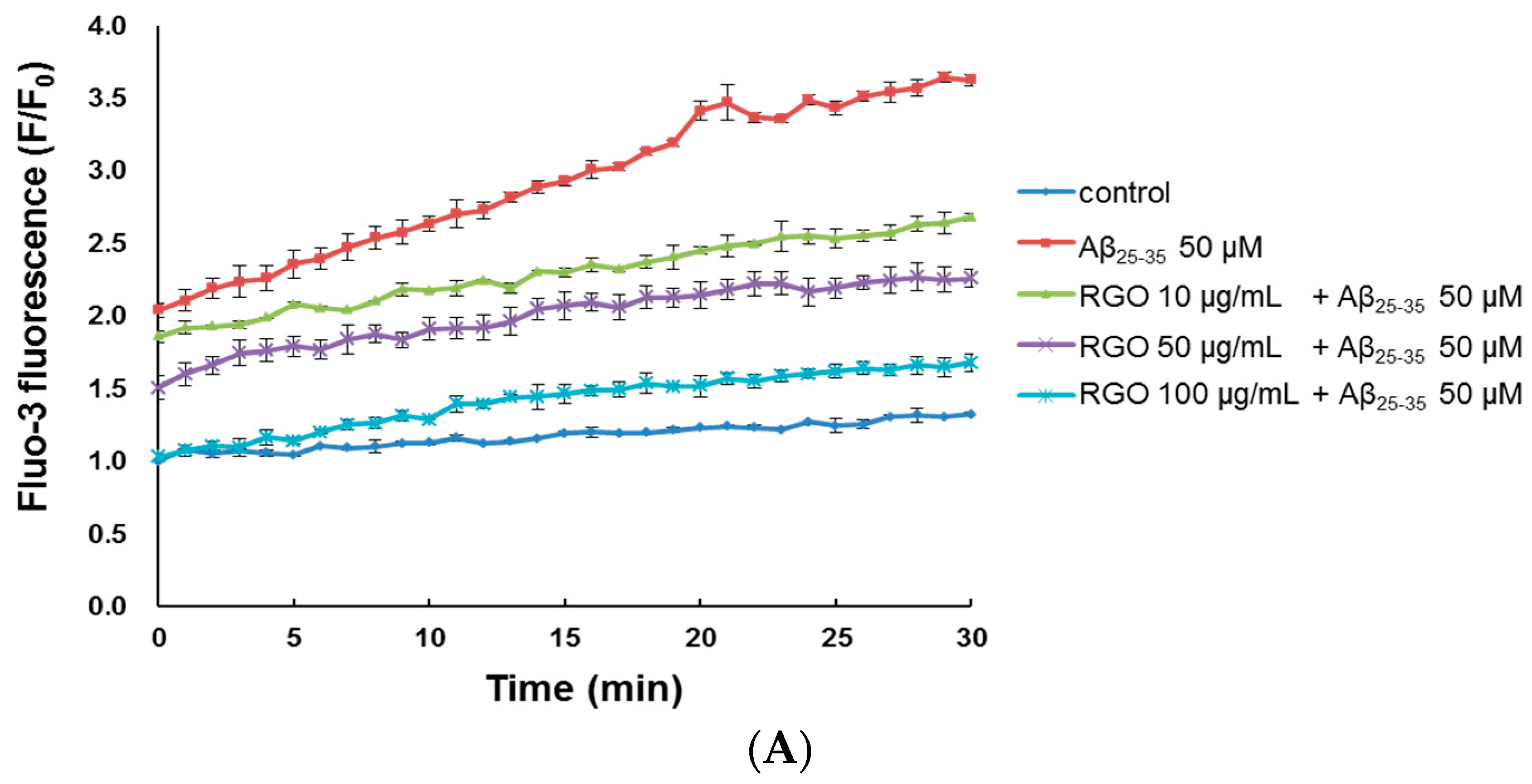
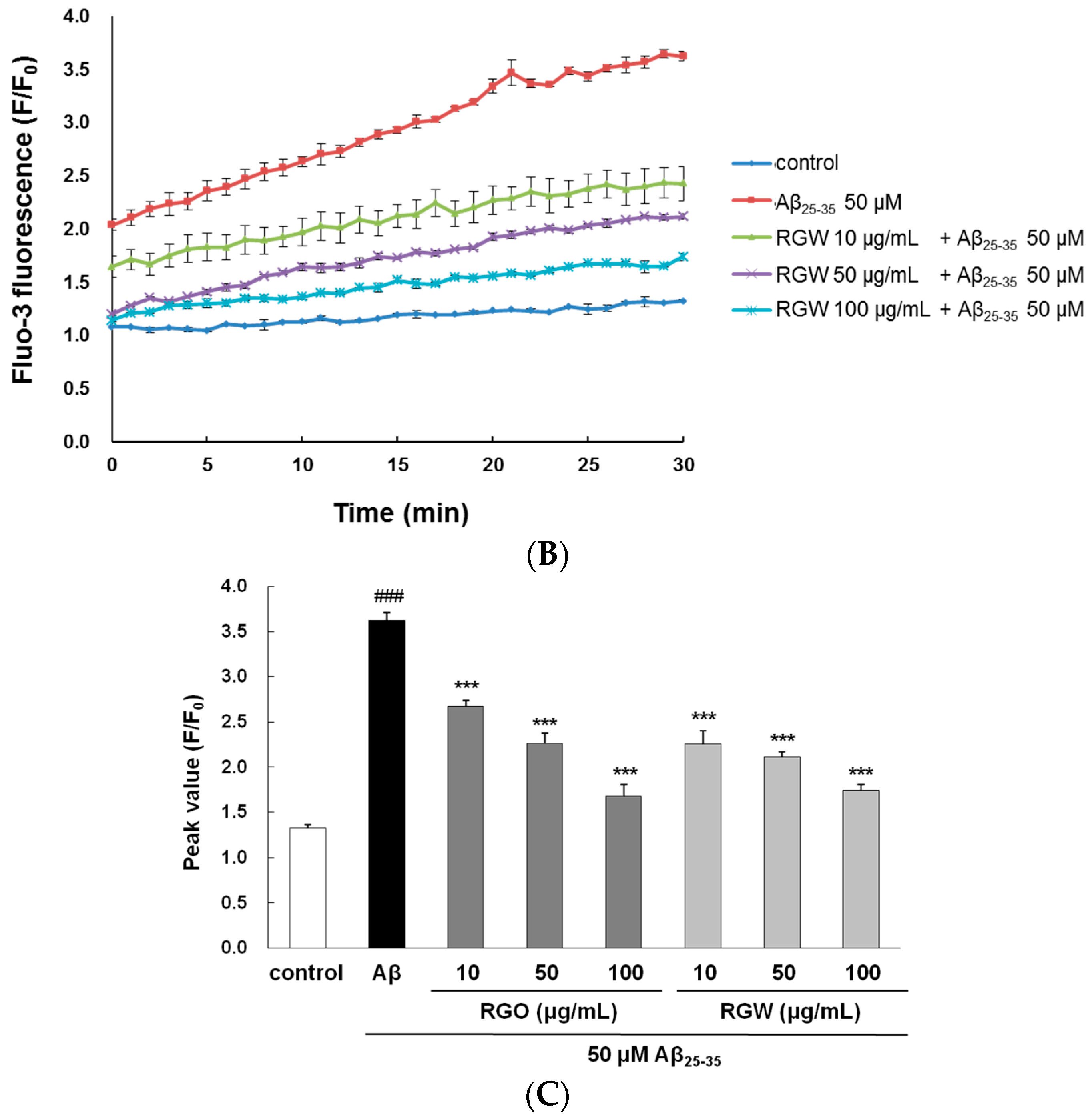
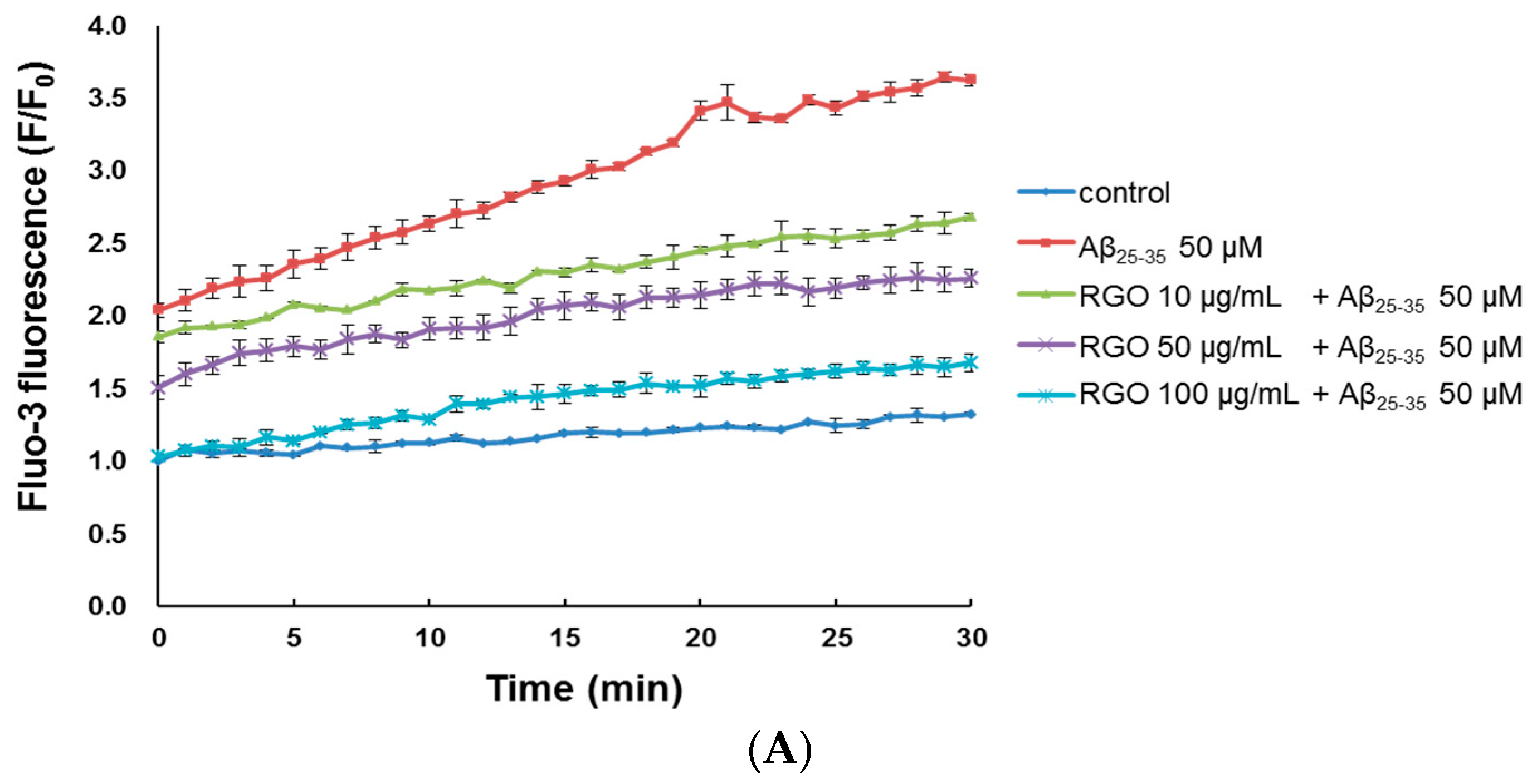
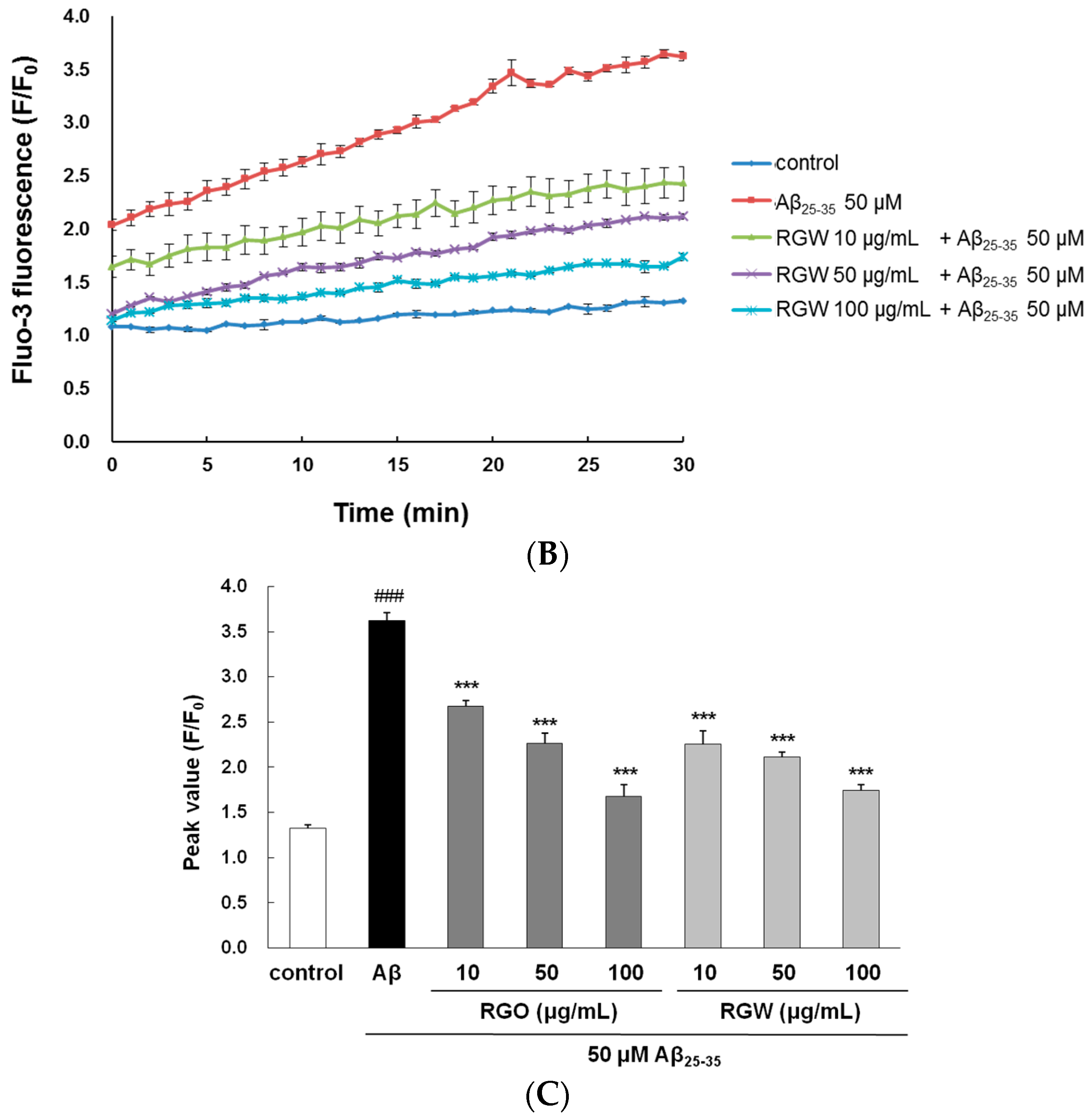
2.4. RGO on Aβ25–35-Induced Ca2+ Influx
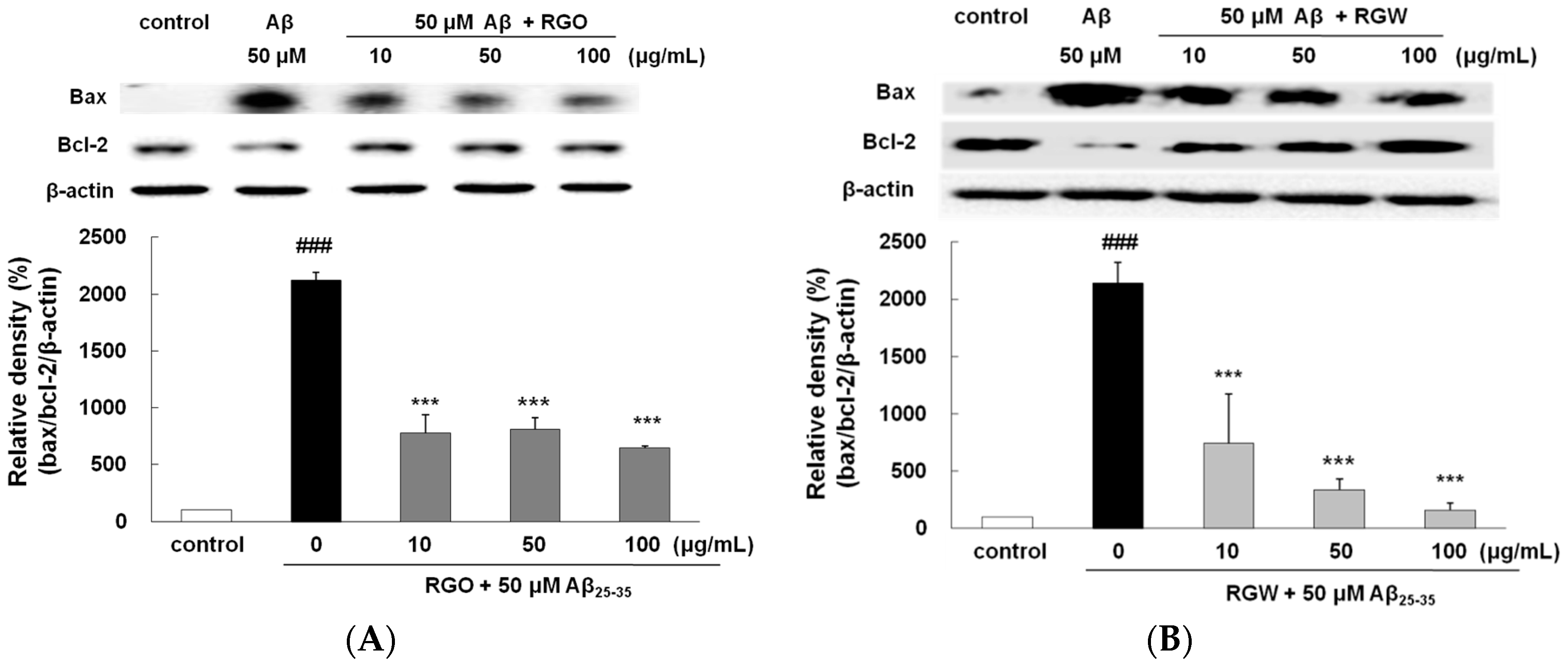
2.5. RGO on Aβ25–35-Stimulated Changes in Bcl-2 and Bax Expression
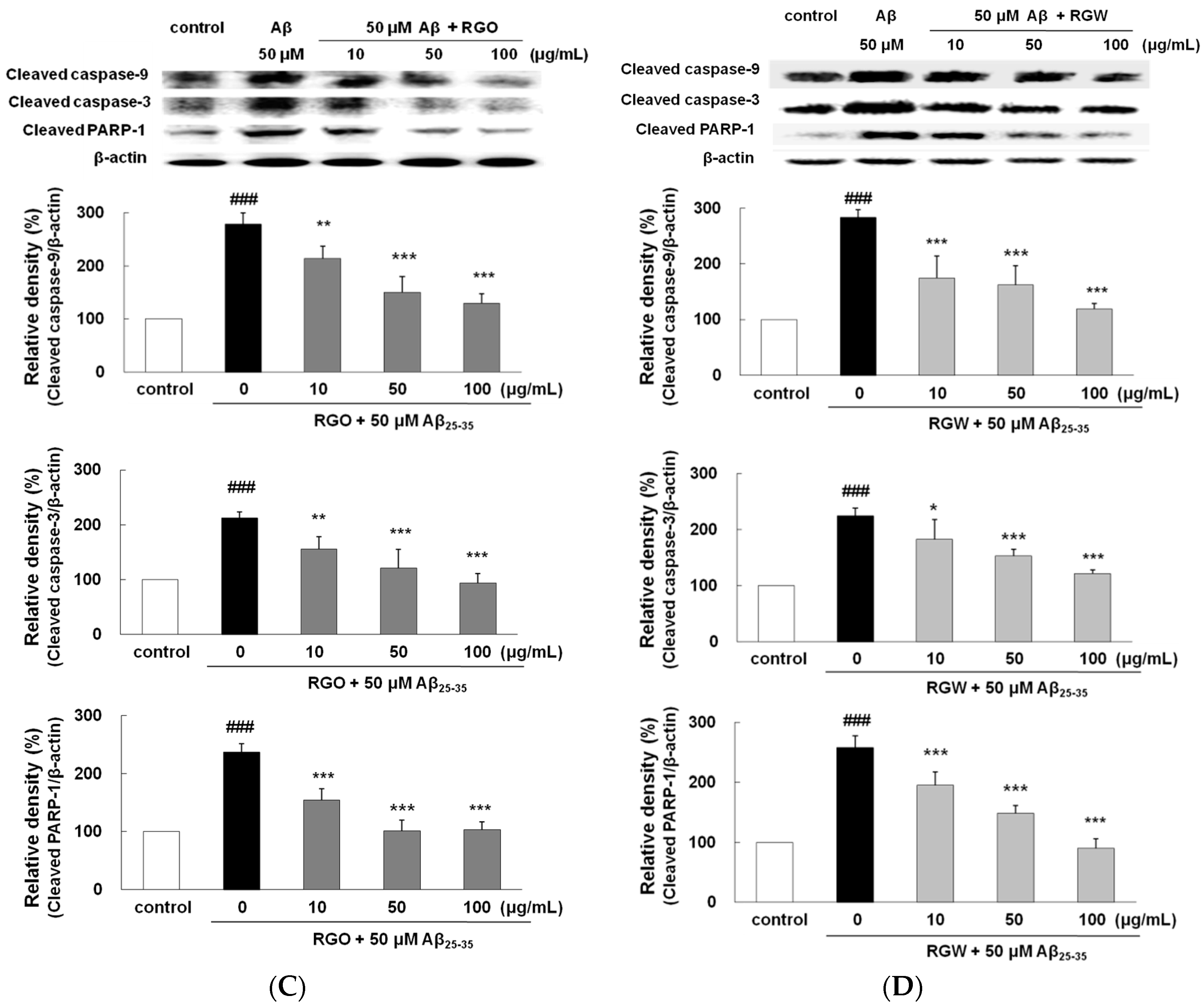
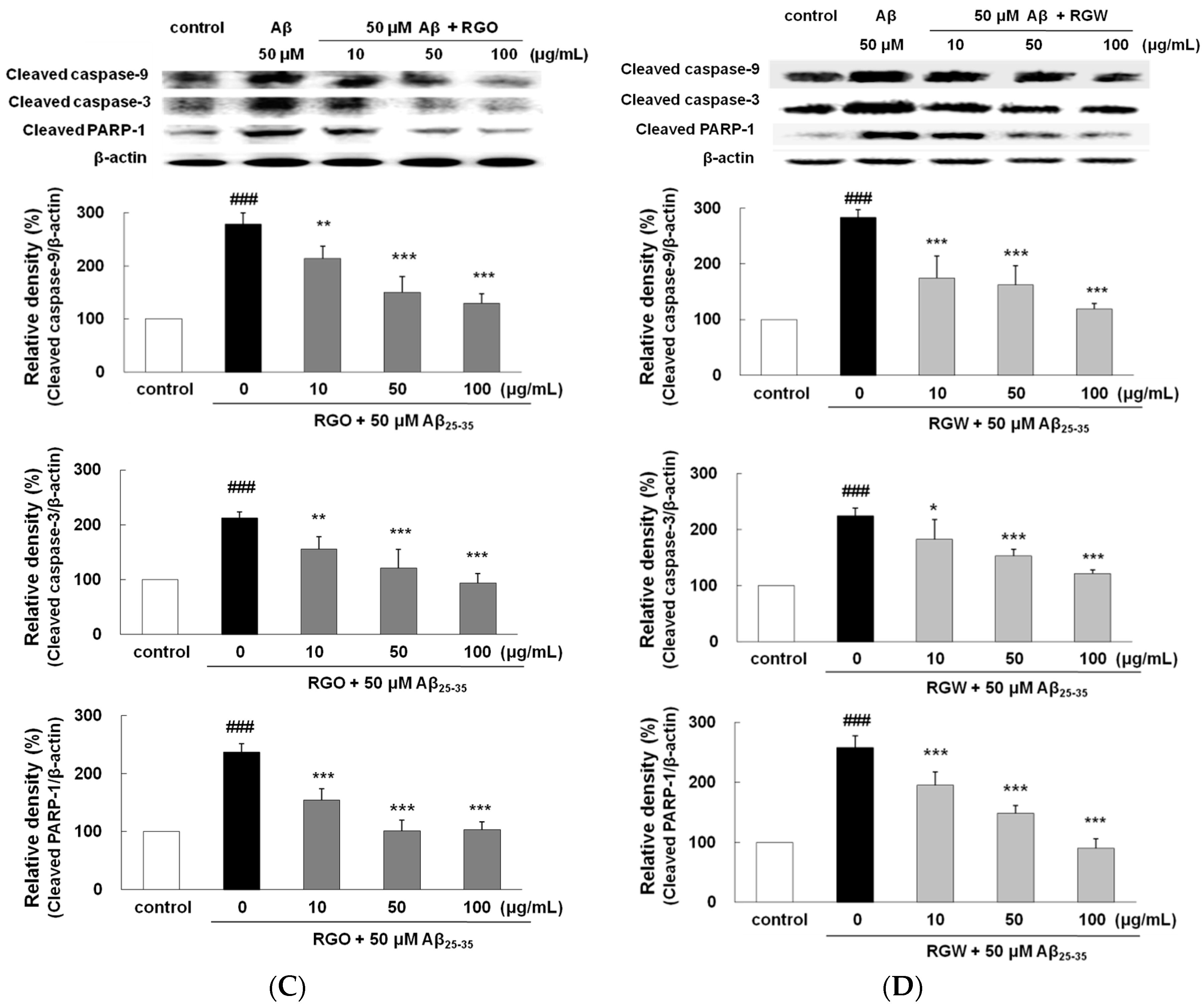
2.6. RGO on Aβ25–35-Stimulated Expression of Caspase-3, Caspase-9, and PARP-1
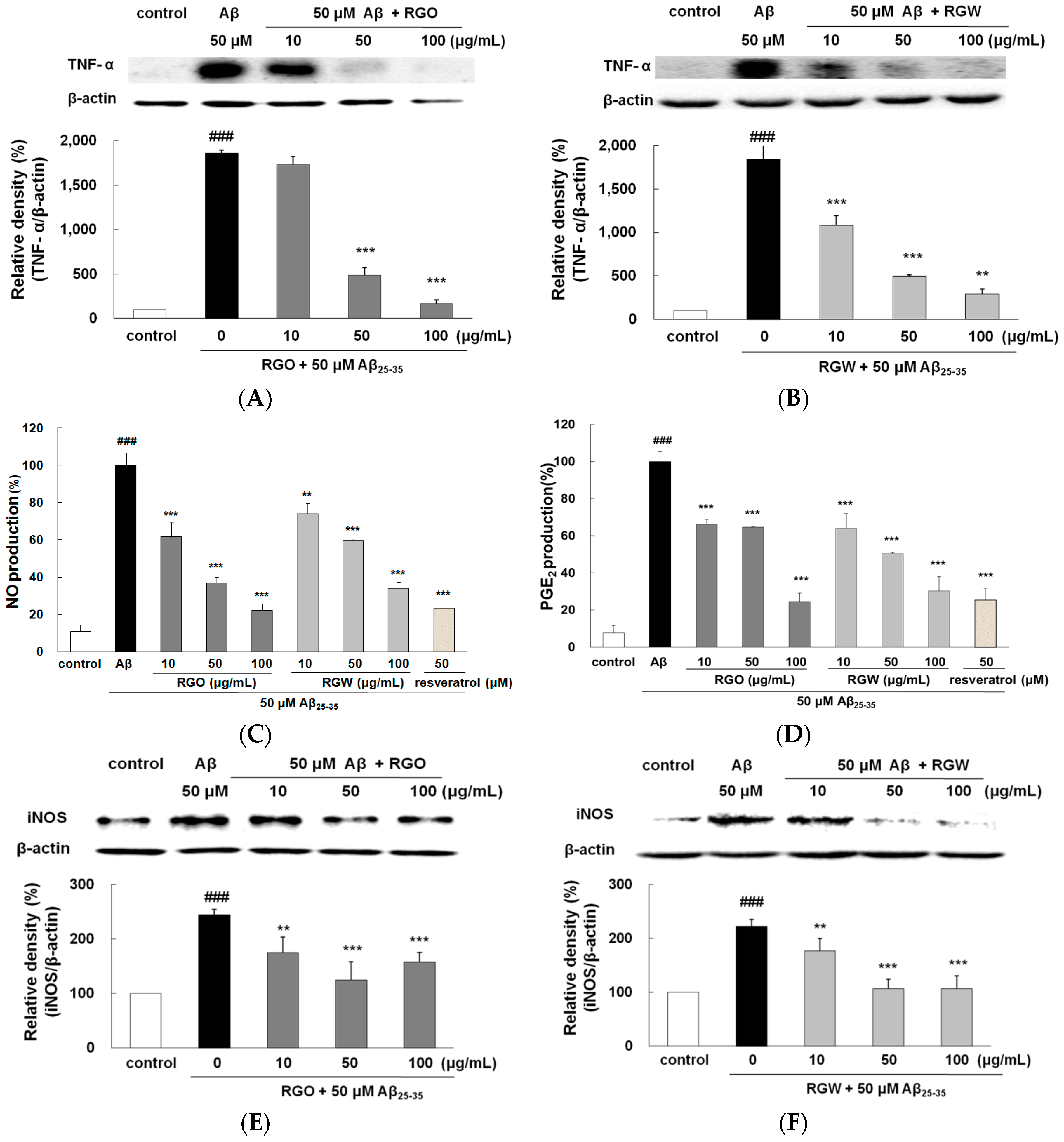
2.7. RGO on Aβ25–35-Stimulated TNF-α Production
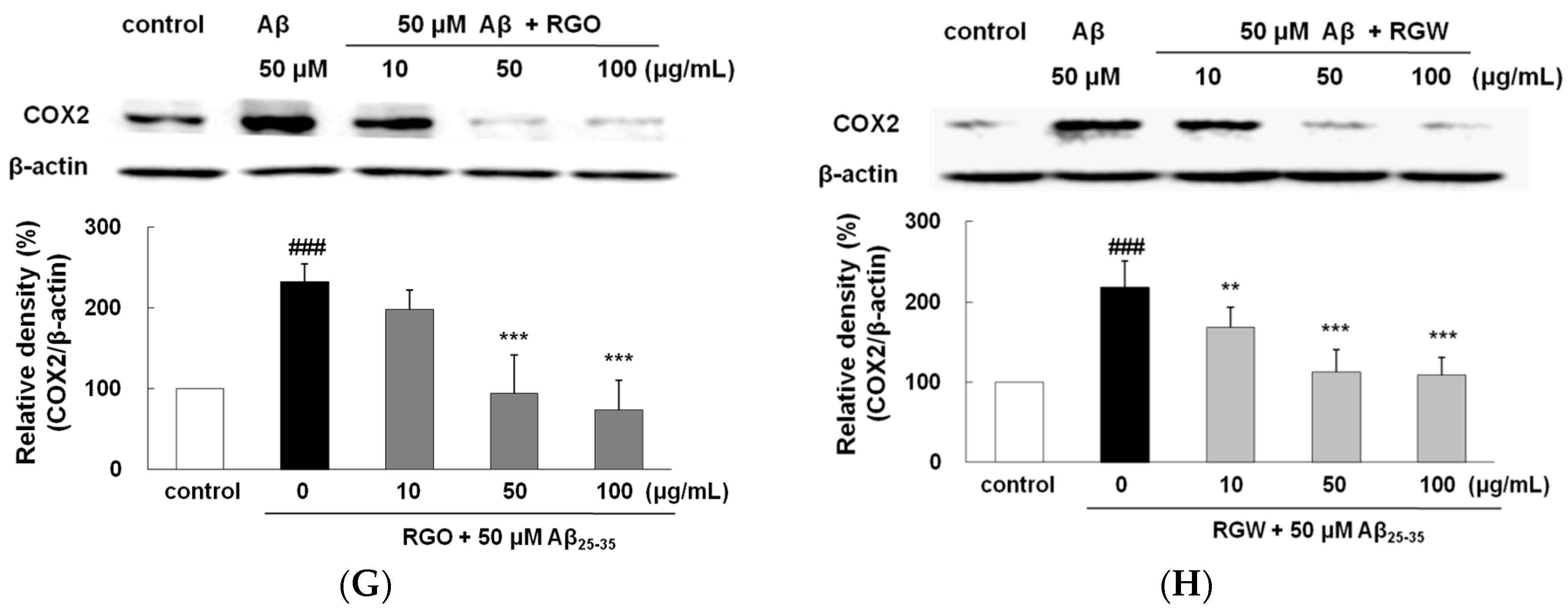
2.8. RGO on Aβ25–35-Stimulated NO/PGE2 Production and iNOS/COX-2 Expression
2.9. RGO on Aβ25–35-Stimulated NF-κB and IκB-α Phosphorylation
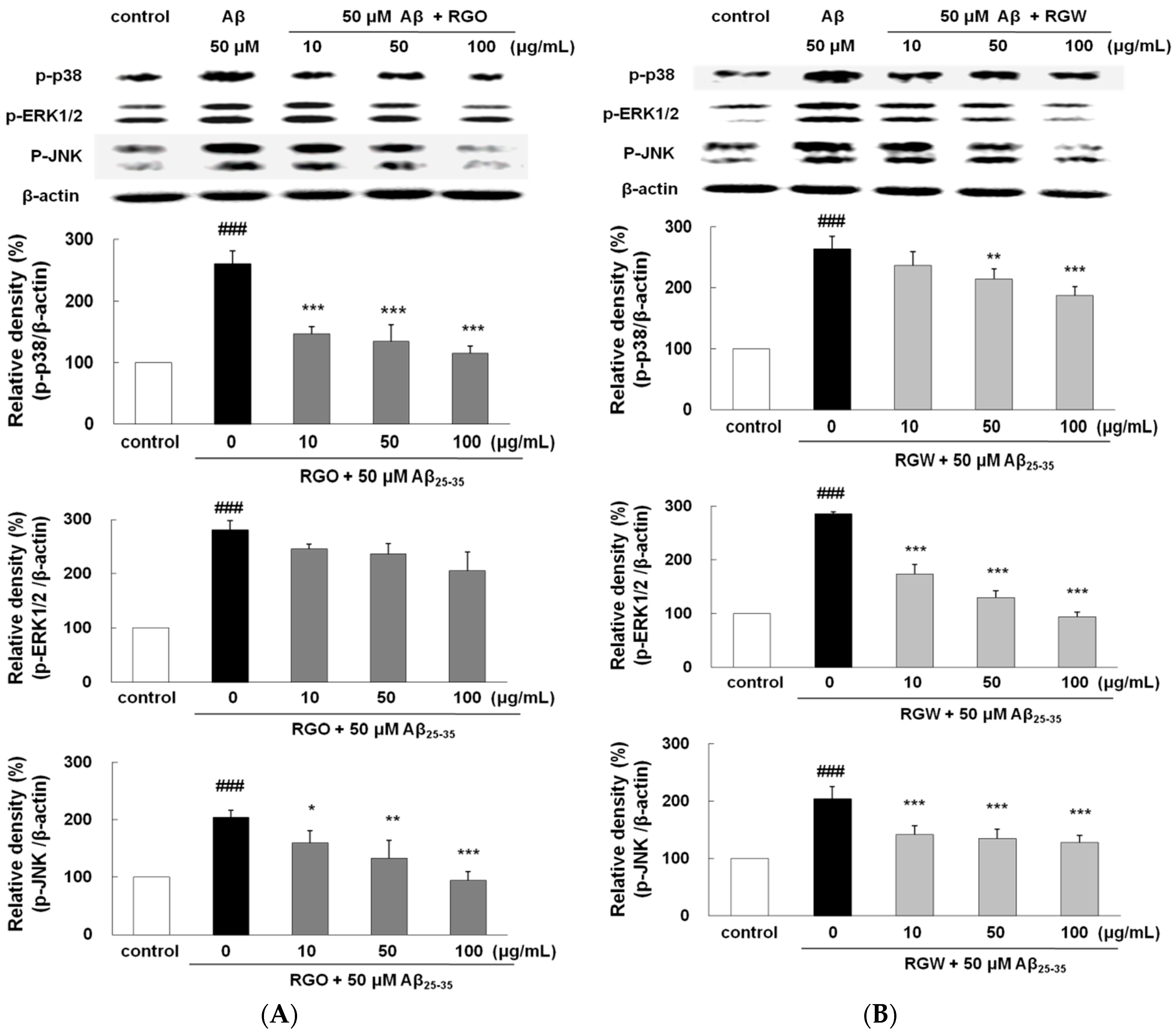
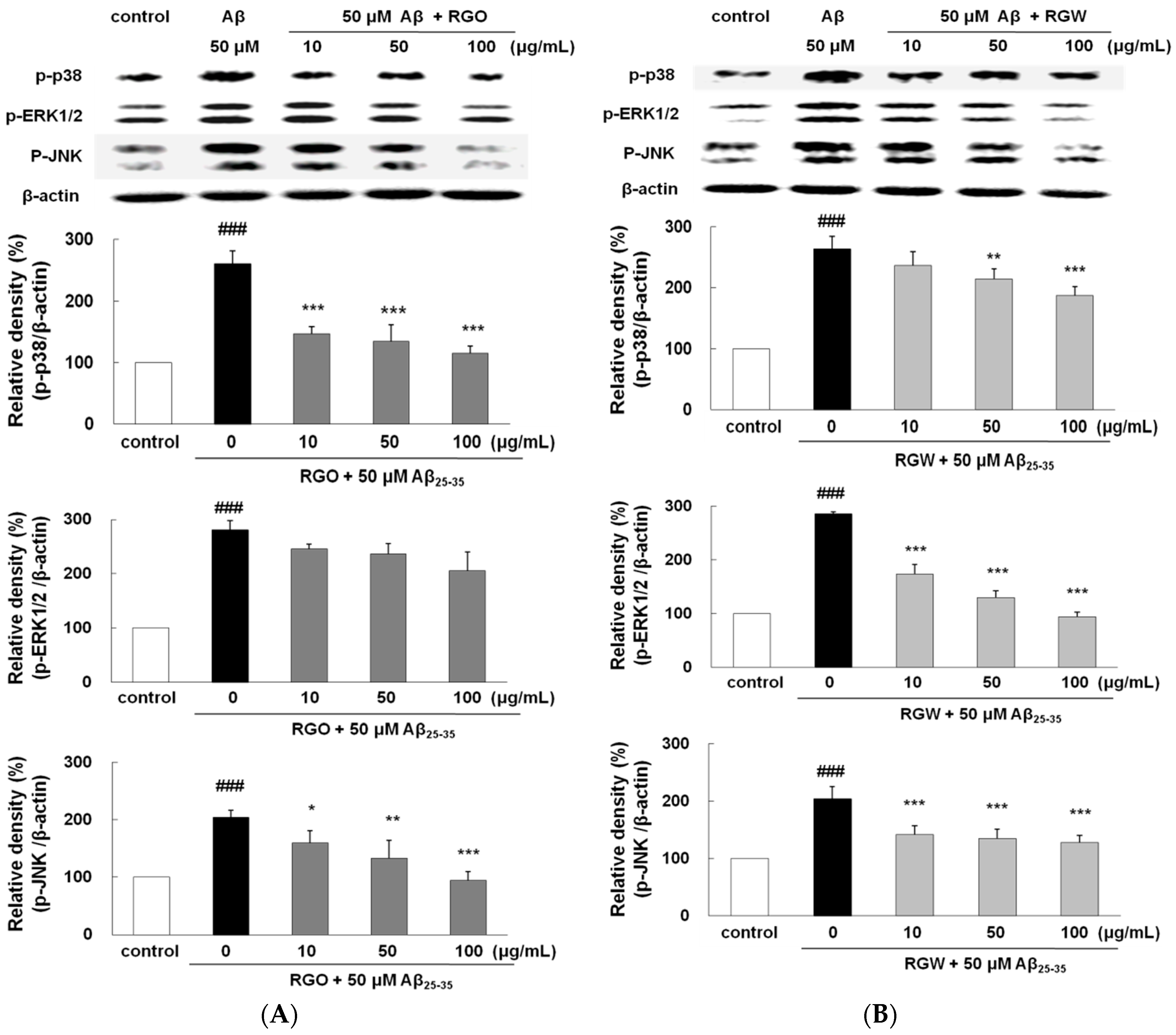
2.10. RGO on Aβ25–35-Induced MAPK Activation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Samples
4.3. Cell Culture and Peptides
4.4. Cell Viability
4.5. Intracellular ROS Accumulation
4.6. Fluorescent Staining of Cells with Hoechst 33342
4.7. MMP Assay
4.8. Intracellular Free Calcium
4.9. Nitrite Determination
4.10. PGE2 Enzyme-Linked Immunoassay
4.11. Flow Cytometry Analysis and Western Blot Analysis
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gandy, S. Molecular basis for anti-amyloid therapy in the prevention and treatment of Alzheimer’s disease. Neurobiol. Aging 2002, 23, 1009–1016. [Google Scholar] [CrossRef]
- Marcello, E.; Epis, R.; Di Luca, M. Amyloid flirting with synaptic failure: Towards a comprehensive view of Alzheimer’s disease pathogenesis. Eur. J. Pharmacol. 2008, 585, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, Y.; Liu, G.; Zhang, J. Dactylorhin B reduces toxic effects of β-amyloid fragment (25–35) on neuron cells and isolated rat brain mitochondria. Naunyn-Schmiedberg’s Arch. Pharmacol. 2006, 374, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.-K.; Yang, H.-Q.; Wang, Z.-Q.; Pan, J.; Hong, Z.; Chen, S.-D. Erythropoietin prevents PC12 cells from beta-amyloid-induced apoptosis via PI3K/Akt pathway. Transl. Neurodegener. 2012, 2012, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cheung, Z.H.; Leung, M.C.; Yip, H.K.; Wu, W.; Siu, F.K.; So, K.F. A neuroprotective herbal mixture inhibits caspase-3-independent apoptosis in retinal ganglion cells. Cell Mol. Neurobiol. 2008, 28, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Berger, N.A. Poly (ADP-ribose) in the cellular response to DNA damage. Radiat. Res. 1985, 101, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Terai, K.; Matsuo, A.; McGeer, P.L. Enhancement of immunoreactivity for NF-κB in the hippocampal formation and cerebral cortex of Alzheimer’s disease. Brain Res. 1996, 735, 159–168. [Google Scholar] [CrossRef]
- Khan, A.; Vaibhav, K.; Javed, H.; Tabassum, R.; Ahmed, M.E.; Khan, M.M.; Khan, M.B.; Shrivastava, P.; Islam, F.; Siddiqui, M.S.; et al. 1,8-Cineole (Eucalyptol) mitigates inflammation in amyloid beta toxicated PC12 cells: Relevance to Alzheimer’s disease. Neurochem. Res. 2014, 39, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shepherd, E.G.; Nelin, L.D. MAPK phosphatases regulating the immune response. Nat. Rev. Immunol. 2007, 7, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Jung, N.P.; Jin, S.H. Studies on the physiological and biochemical effects of Korean ginseng. J. Ginseng Res. 1996, 20, 431–471. [Google Scholar]
- Tu, L.H.; Ma, J.; Liu, H.P.; Wang, R.R.; Luo, J. The neuroprotective effects of ginsenosides on calcineurin activity and tau phosphorylation in SY5Y cells. Cell Mol. Neurobiol. 2009, 29, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Lee, M.J.; Kim, I.H.; Pyo, S.; Choi, K.T.; Rhee, D.K. Anti-apoptotic effects of red ginseng on oxidative stress induced by hydrogen peroxide in SK-N-SH cells. J. Ginseng Res. 2010, 34, 138–144. [Google Scholar] [CrossRef]
- Han, J.Y.; Ahn, S.Y.; Oh, E.H.; Nam, S.Y.; Hong, J.T.; Oh, K.W.; Lee, M.K. Red ginseng extract attenuates kainate-induced excitotoxicity a by antioxidative effects. Evid. Based Complement. Altern. Med. 2012, 2012, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Shim, J.; Lee, S.; Cho, W.H.; Hong, E.; Lee, J.H.; Han, J.H.; Lee, H.J.; Lee, K.W. Rg3-enriched ginseng extract ameliorates scopolamine-induced learning deficits in mice. BMC Complement. Altern. Med. 2016, 16, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.D.; Yoo, G.; Chae, H.J.; In, M.J.; Oh, N.S.; Hwang, Y.K.; Hwang, W.I.; Kim, D.C. Lipid-soluble extracts as the main source of anticancer activity in ginseng and ginseng marc. J. Am. Oil Chem. Soc. 2009, 86, 1065–1071. [Google Scholar] [CrossRef]
- Bak, M.J.; Jun, M.; Jeong, W.S. Antioxidant and hepatoprotective effects of the red ginseng essential oil in H2O2-treated HepG2 cells and CCl4-treated mice. Int. J. Mol. Sci. 2012, 13, 2314–2330. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.J.; Truong, V.L.; Ko, S.Y.; Nguyen, X.M.G.; Jun, M.; Hong, S.G.; Lee, J.W.; Jeong, W.S. Induction of Nrf2/ARE-mediated cytoprotective genes by red ginseng oil through ASK1eMKK4/7eJNK and p38 MAPK signaling pathways in HepG2 cells. J. Ginseng Res. 2016, 40, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Youn, K.; Yun, E.Y.; Lee, J.; Kim, J.Y.; Hwang, J.S.; Jeong, W.S.; Jun, M. Oleic acid and linoleic acid from Tenebrio molitor larvae inhibit BACE1 activity in vitro: Molecular docking studies. J. Med. Food 2014, 17, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.J.; Kim, K.B.; Jun, M.; Jeong, W.S. Safety of red ginseng oil for single oral administration in Sprague Dawley rats. J. Ginseng Res. 2014, 38, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Y.; Liao, J.; Kim, K.; Yurkow, E.J.; Yang, C.S. Inhibition of growth and induction of apoptosis in human cancer cell lines by tea polyphenols. Carcinogenesis 1998, 19, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Martίn, D.; Salinas, M.; Lόpez-Valdaliso, R.; Serrano, E.; Recuero, M.; Cuadrado, A. Effect of the Alzheimer amyloid fragment Aβ(25–35) on Akt/PKB kinase and survival of PC12 cells. J. Neurochem. 2001, 78, 1000–1008. [Google Scholar] [CrossRef]
- Cao, L.; Wang, H.; Wang, F.; Xu, D.; Liu, F.; Liu, C. Aβ-induced senescent retinal pigment epithelial cells create a proinflammatory microenvironment in AMD. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3738–3750. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.R.; Kim, H.M.; Kang, J.S.; Lee, K.; Lee, S.D.; Hyun, D.H.; In, M.J.; Park, S.K.; Kim, D.C. Lipid-soluble ginseng extract induces apoptosis and G0/G1 cell cycle arrest in NCI-H460 human lung cancer cells. Plant Foods Hum. Nutr. 2011, 6, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.; Samali, A.; Orrenius, S. Triggering and modulation of apoptosis by oxidative stress. Free Radic. Biol. Med. 2000, 29, 323–333. [Google Scholar] [CrossRef]
- Ullah, I.; Ullah, N.; Naseer, M.I.; Lee, H.Y.; Kim, M.O. Neuroprotection with metformin and thymoquinone against ethanol-induced apoptotic neurodegeneration in prenatal rat cortical neurons. BMC Neurosci. 2012, 13, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.; Yin, X.M.; Wang, K.; Wei, M.C.; Jockel, J.; Milliman, C. Caspase cleaved BID targets mitochondria and is required for cytochrome c release, while BCL-XL prevents this release but not tumor necrosis factor-R1/Fas death. J. Biol. Chem. 1999, 274, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, Y.; Cho, J. Korean red ginseng extract exhibits neuroprotective effects through inhibition of apoptotic cell death. Biol. Pharm. Bull. 2014, 37, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Hseu, Y.C.; Wu, F.Y.; Wu, J.J.; Chen, J.Y.; Chang, W.H.; Lu, F.J.; Lai, Y.C.; Yang, H.L. Anti-inflammatory potential of Antrodia Camphorata through inhibition of iNOS, COX-2 and cytokines via the NF-κB pathway. Int. Immunopharmacol. 2005, 5, 1914–1925. [Google Scholar] [CrossRef] [PubMed]
- Seibert, K.; Zhang, Y.; Leahy, K.; Hauser, S.; Masferrer, J.; Perkins, W. Pharmacological and biochemical demonstration of the role of cyclooxygenase-2 in inflammation and pain. Proc. Natl. Acad. Sci. USA 1997, 91, 12013–12017. [Google Scholar] [CrossRef]
- Jin, Y.; Kotakadiy, V.S.; Ying, L.; Hofseth, A.B.; Cui, X.; Wood, P.A.; Windust, A.; Matesic, L.E.; Pena, E.A.; Chiuzan, C.; et al. American ginseng suppresses inflammation and DNA damage associated with mouse colitis. Carcinogenesis 2008, 29, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Li, J.; Nagarkatti, P.; Nagarkatti, M.; Hofseth, L.J.; Windust, A.; Cui, T. American ginseng preferentially suppresses STAT/iNOS signaling in activated macrophages. J. Ethnopharmacol. 2009, 125, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.J.; Hong, S.G.; Lee, J.W.; Jeong, W.S. Red ginseng marc oil inhibits iNOS and COX-2 via NFκB and p38 pathways in LPS-stimulated RAW 264.7 macrophages. Molecules 2012, 17, 13769–13786. [Google Scholar] [CrossRef] [PubMed]
- Ortet, R.; Regalado, E.L.; Thomas, O.P.; Pino, J.A.; Fernandez, M.D. Chemical composition and antioxidant activities of the essential oil from Tornabenea bischoffii (Apiaceae). Nat. Prod. Commun. 2011, 6, 1179–1182. [Google Scholar] [PubMed]
- Sharififar, F.; Derakhshanfar, A.; Dehghan-Nudeh, G.; Abbasi, N.; Abbasi, R.; Gharaei, R.R.; Koohpayeh, A.; Daneshpajouh, M. In vivo antioxidant activity of Zataria multiflora Boiss essential oil. Pak. J. Pharm. Sci. 2011, 24, 221–225. [Google Scholar] [PubMed]
- Muhammad, A.; Abdul, S.; Muhammad, J.; Farhat, U.; Fazal, S.; Jawad, A. Neuroprotective and Anti-Aging Potentials of Essential Oils from Aromatic and Medicinal Plants. Front. Aging Neurosci. 2017, 9, 168–183. [Google Scholar]
- Miroddi, M.; Navarra, M.; Quattropani, M.C.; Calapai, F.; Gangemi, S.; Calapai, G. Systematic review of clinical trials assessing pharmacological properties of Salvia species on memory, cognitive impairment and Alzheimer’s disease. CNS Neurosci. Ther. 2014, 2, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Tildesley, N.T.; Kennedy, D.O.; Perry, E.K.; Ballard, C.G.; Savelev, S.; Wesnes, K.A.; Scholey, A.B. Salvia lavandulaefolia (Spanish Sage) enhances memory in healthy young volunteers. Pharmacol. Biochem. Behav. 2003, 75, 669–674. [Google Scholar] [CrossRef]
- Perry, N.S.; Bollen, C.; Perry, E.K.; Ballard, C. Salvia for dementia therapy: Review of pharmacological activity and pilot tolerability clinical trial. Pharmacol. Biochem. Behav. 2003, 75, 651–659. [Google Scholar] [CrossRef]
- Khazdair, M.R. The protective effects of Nigella sativa and its constituents on induced neurotoxicity. J. Toxicol. 2015, 2015, 841823. [Google Scholar] [CrossRef] [PubMed]
- Farr, S.A.; Price, T.O.; Dominguez, L.J.; Motisi, A.; Saiano, F.; Niehoff, M.L.; Morley, J.E.; Banks, W.A.; Ercal, N.; Barbagallo, M. Extra virgin olive oil improves learning and memory in SAMP8 mice. J. Alzheimers Dis. 2012, 28, 81–92. [Google Scholar] [PubMed]
- Qosa, H.; Mohamed, L.A.; Bataresh, Y.S.; Alqahtani, S.; Ibrahim, B.; LeVine, H.; Keller, J.N.; Kaddoumi, A. Extra virginolive oil attenuates amyloid-b and tau pathologies in the brains of TgSwDI mice. J. Nutr. Biochem. 2015, 26, 1479–1490. [Google Scholar] [CrossRef] [PubMed]
- Elisabett, L.; Luigi, I.; Domenico, P. Extra-virgin olive oil ameliorates cognition and neuropathology of the 3xTg mice: Role of autophagy. Ann. Clin. Transl. Neurol. 2017, 4, 564–574. [Google Scholar]
- Kim, H.J.; Kang, H.J.; Seo, J.Y.; Lee, C.H.; Kim, Y.S.; Kim, J.S. Antiobesity effect of oil extract of ginseng. J. Med. Food 2011, 14, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.D.; Park, S.K.; Lee, E.S.; Kim, H.M.; Lee, C.W.; Lee, K.; Lee, K.H.; Kang, M.R.; Lee, K.S.; Lee, J.; et al. A lipid-soluble red ginseng extract inhibits the growth of human lung tumor xenografts in nude mice. J. Med. Food 2010, 13, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, J.H.; Song, Y.B.; Park, K.H. Effects of dietary supplementation of lipophilic fraction from Panax ginseng on cGMP and cAMP in rat platelets and on blood coagulation. Biol. Pharm. Bull. 1996, 19, 1434–1439. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Kim, S.S.; Cho, C.W.; Choi, S.Y.; In, G.; Kim, K.T. Quality and characteristics of ginseng seed oil treated using different extraction methods. J. Ginseng Res. 2013, 37, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Youn, K.; Yun, E.-Y.; Hwang, J.S.; Jeong, W.S.; Ho, C.T.; Jun, M. Oleic acid ameliorates Aβ-induced inflammation by downregulation of COX-2 and iNOS via NFκB signaling pathway. J. Funct. Foods 2015, 14, 1–11. [Google Scholar] [CrossRef]
- Brimson, J.M.; Brimson, S.J.; Brimson, C.A.; Rakkhitawatthana, V.; Tencomnao, T. Rhinacanthus nasutus extracts prevent glutamate and amyloid-β neurotoxicity in HT-22 mouse hippocampal cells: Possible active compounds Include lupeol, stigmasterol and β-sitosterol. Int. J. Mol. Sci. 2012, 13, 5074–5097. [Google Scholar] [CrossRef] [PubMed]
- Drever, B.D.; Anderson, W.G.; Riedel, G.; Kim, D.H.; Ryu, J.H.; Choi, D.Y.; Platt, B. The seed extract of Cassia obtusifolia offers neuroprotection to mouse hippocampal cultures. J. Pharmacol. Sci. 2008, 107, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, T.; Kim, S.W.; Hwang, S.Y.; Sohn, S.H.; Yoo, S.K.; Kim, S.K. Panax ginseng reduces oxidative stress and restores antioxidant capacity in aged rats. Nutr. Res. 2012, 32, 718–726. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Youn, K.; Jeong, W.-S.; Ho, C.-T.; Jun, M. Protective Effects of Red Ginseng Oil against Aβ25–35-Induced Neuronal Apoptosis and Inflammation in PC12 Cells. Int. J. Mol. Sci. 2017, 18, 2218. https://doi.org/10.3390/ijms18102218
Lee S, Youn K, Jeong W-S, Ho C-T, Jun M. Protective Effects of Red Ginseng Oil against Aβ25–35-Induced Neuronal Apoptosis and Inflammation in PC12 Cells. International Journal of Molecular Sciences. 2017; 18(10):2218. https://doi.org/10.3390/ijms18102218
Chicago/Turabian StyleLee, Seonah, Kumju Youn, Woo-Sik Jeong, Chi-Tang Ho, and Mira Jun. 2017. "Protective Effects of Red Ginseng Oil against Aβ25–35-Induced Neuronal Apoptosis and Inflammation in PC12 Cells" International Journal of Molecular Sciences 18, no. 10: 2218. https://doi.org/10.3390/ijms18102218
APA StyleLee, S., Youn, K., Jeong, W.-S., Ho, C.-T., & Jun, M. (2017). Protective Effects of Red Ginseng Oil against Aβ25–35-Induced Neuronal Apoptosis and Inflammation in PC12 Cells. International Journal of Molecular Sciences, 18(10), 2218. https://doi.org/10.3390/ijms18102218