
Exogenous Nitric Oxide Suppresses in Vivo X-ray-Induced Targeted and Non-Targeted Effects in Zebrafish Embryos

_Kwan_Ngok_Yu.png)
Abstract
:1. Introduction
2. Results
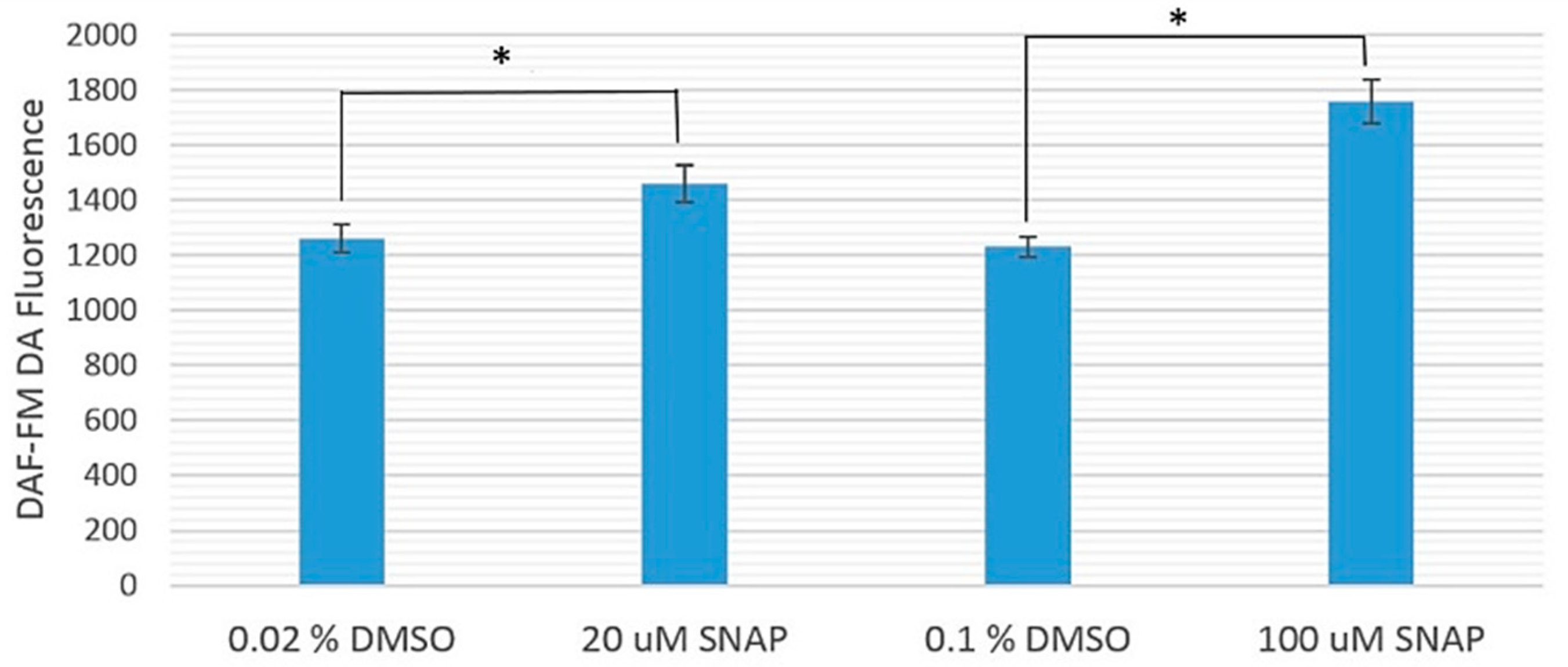
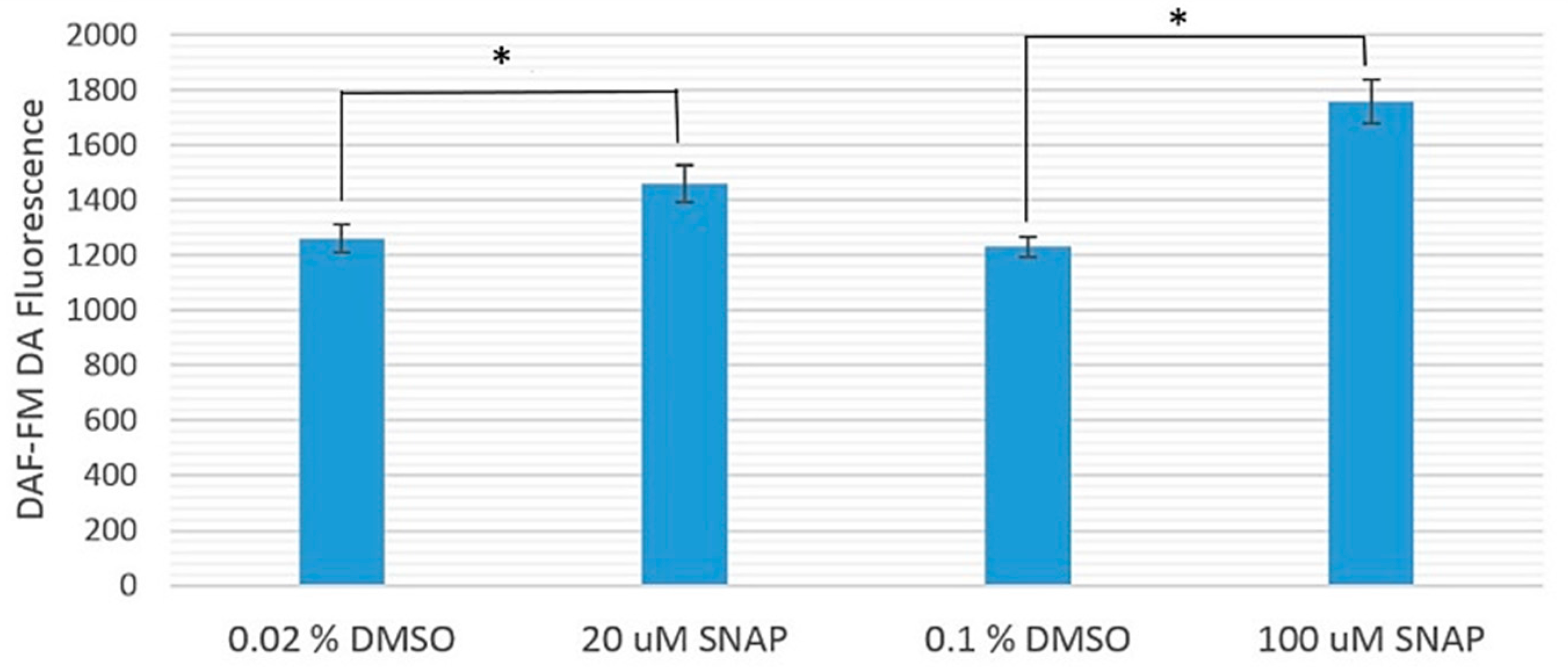
2.1. Cytotoxicity of 20 and 100 μM SNAP on Zebrafish Embryos
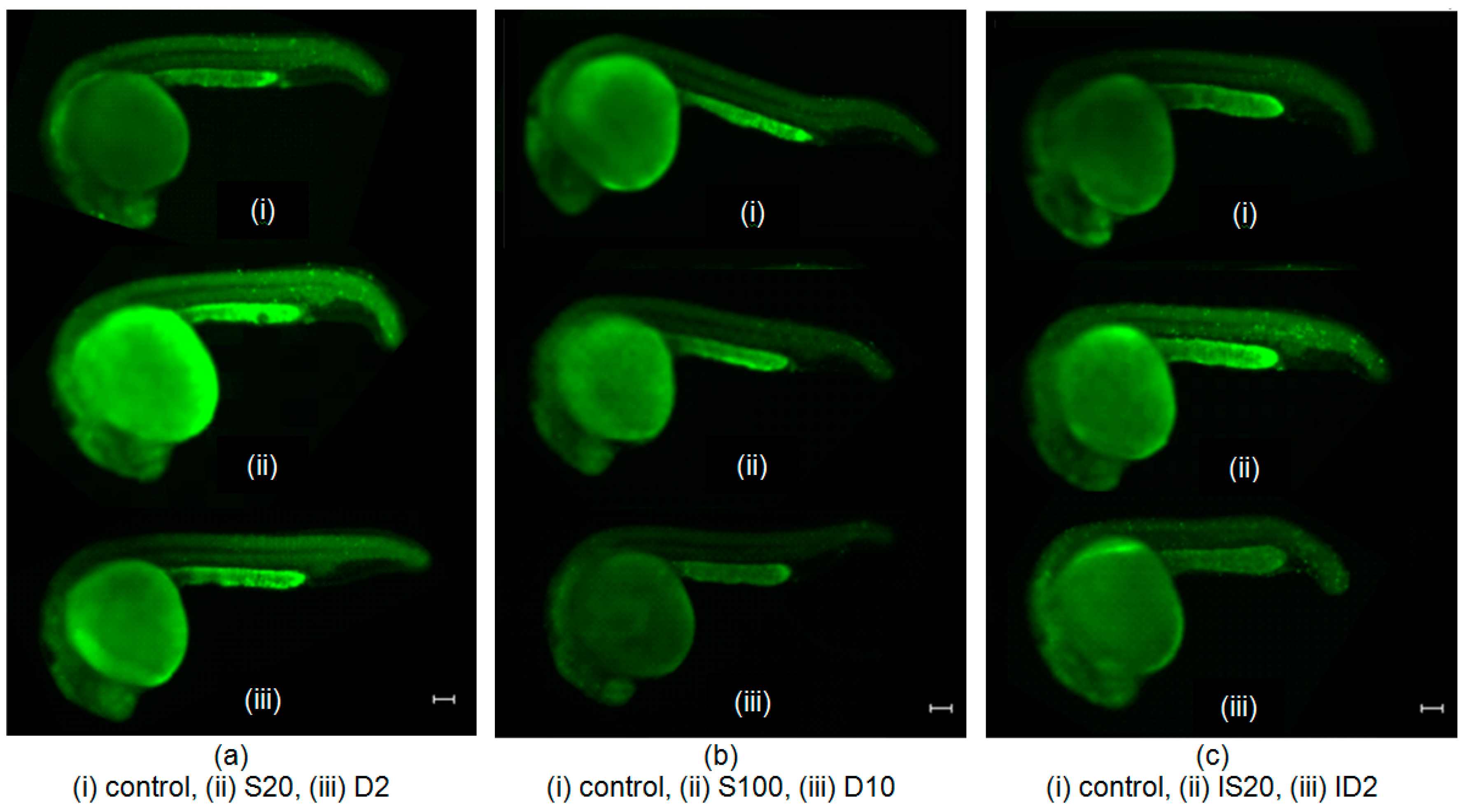
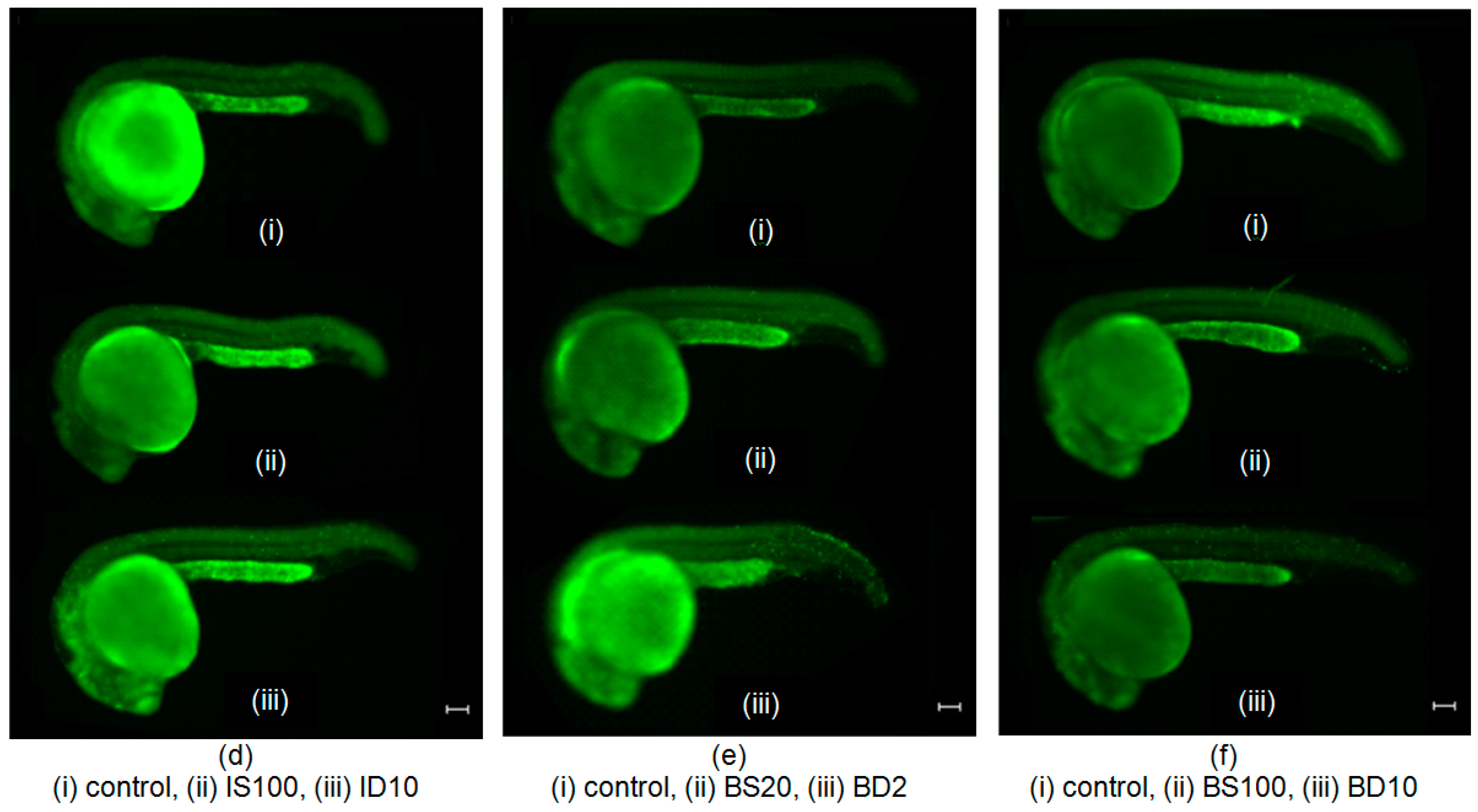
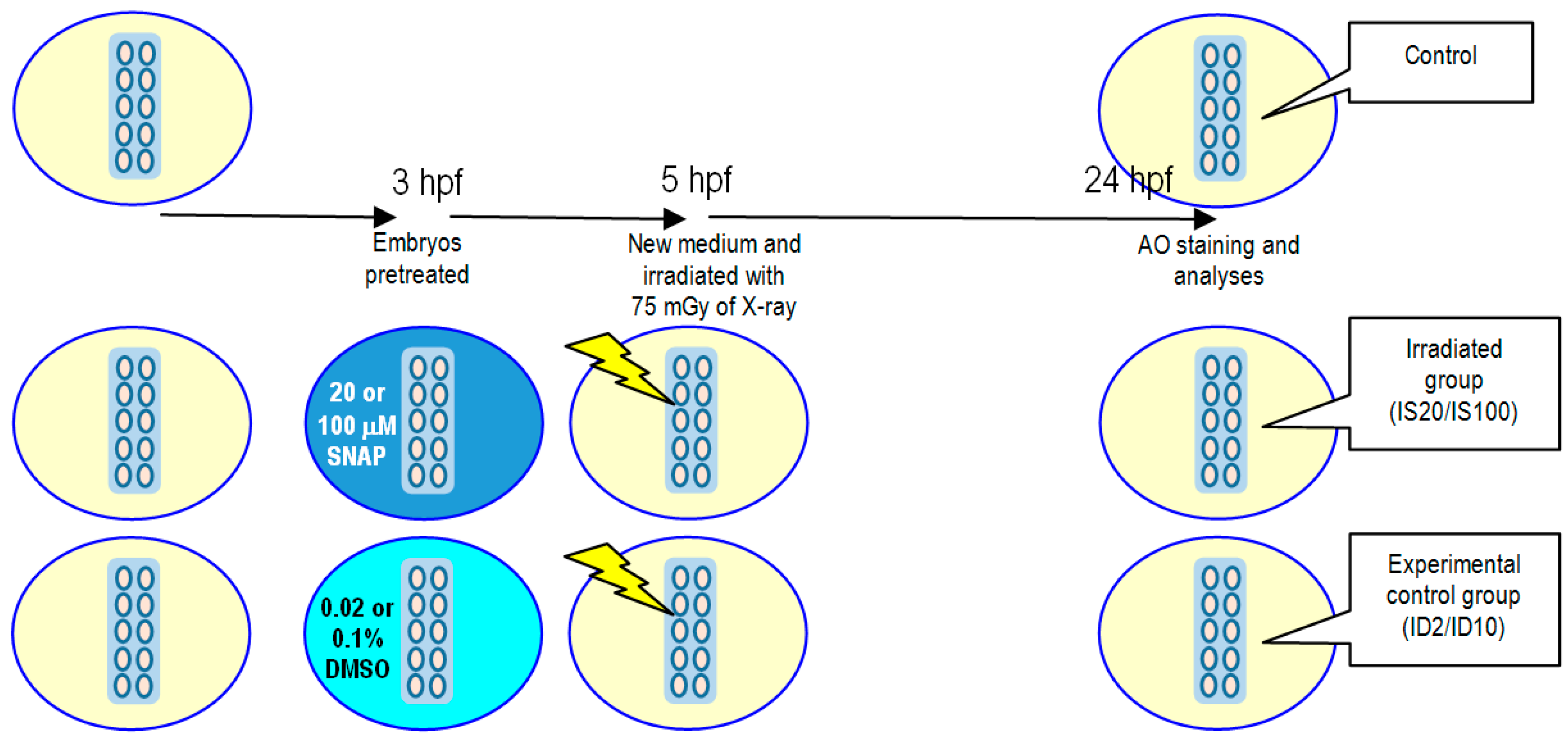
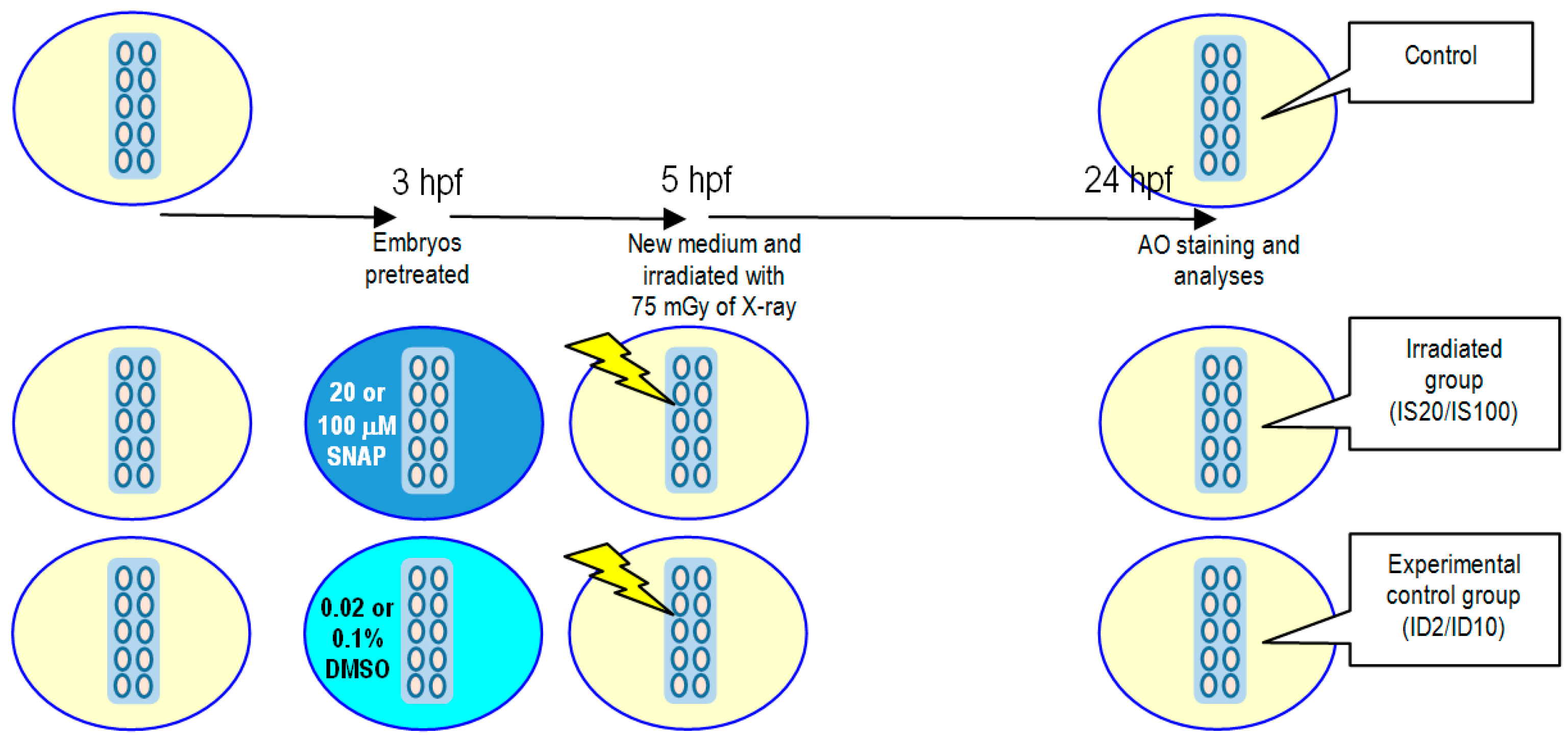
2.2. Effects of SNAP on Zebrafish Embryos Irradiated with 75 mGy X-ray
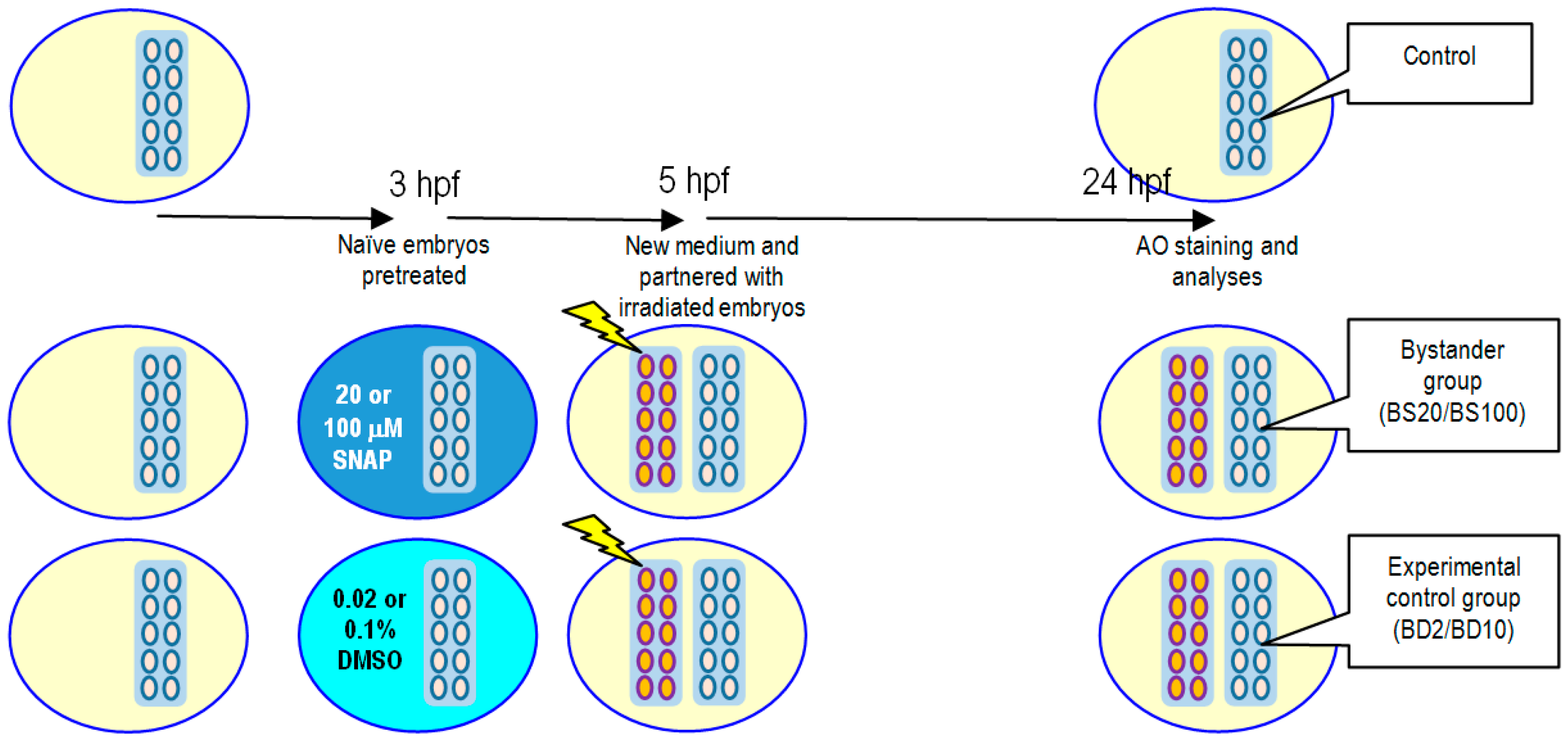
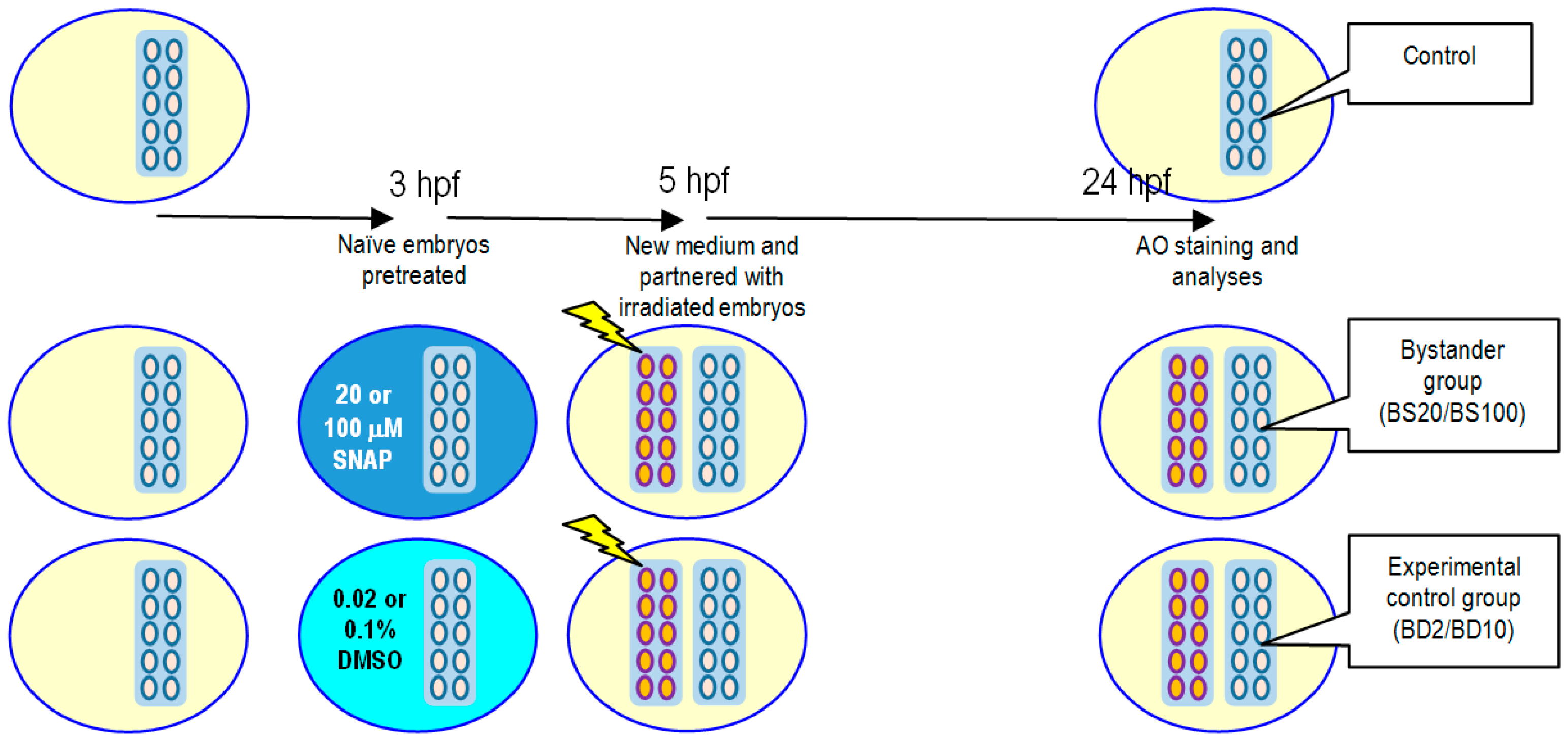
2.3. Effects of SNAP on Radiation-Induced Bystander Effect in Zebrafish Embryos
3. Discussion
4. Materials and Methods
4.1. Zebrafish Maintenance
4.2. Treatment with SNAP
4.3. X-ray Irradiation
4.4. Cytotoxicity of SNAP
4.5. X-ray-Induced Targeted Effects and Non-Targeted Bystander Effects
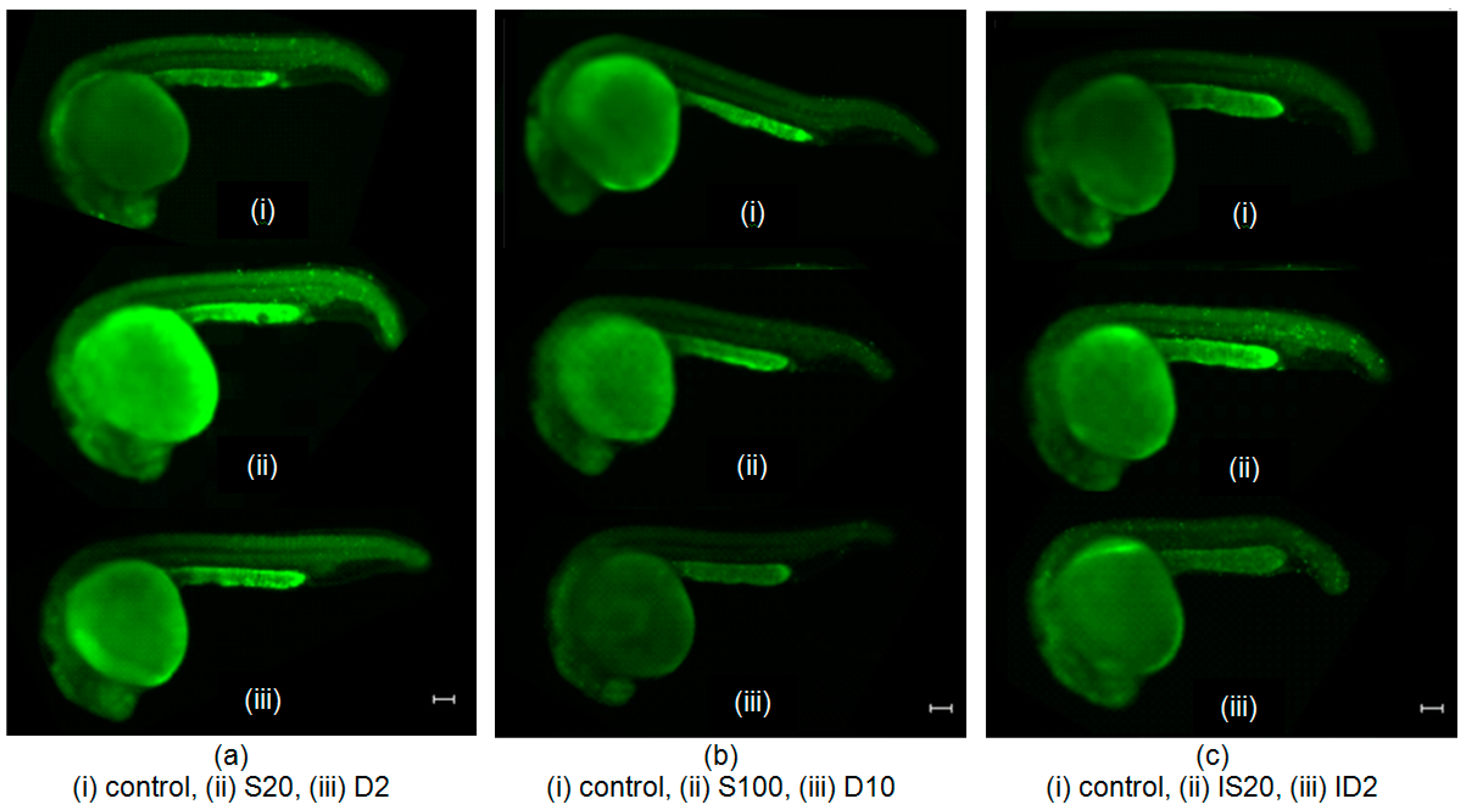
4.6. Vital Dye Acridine Orange (AO) Staining
4.7. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Moncada, S.; Palmer, R.M.; Higgs, E.A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar] [PubMed]
- Brennan, P.A.; Conroy, B.; Spedding, A.V. Expression of inducible nitric oxide synthase and p53 in oral epithelial dysplasia. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2000, 90, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.S.; Koppenol, W.H. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol. 1996, 271, C1424–C1437. [Google Scholar] [PubMed]
- Zhou, H.; Ivanov, V.N.; Lien, Y.C.; Davidson, M.; Hei, T.K. Mitochondrial function and nuclear factor-κb-mediated signaling in radiation-induced bystander effects. Cancer Res. 2008, 68, 2233–2240. [Google Scholar] [CrossRef] [PubMed]
- Leach, J.K.; Black, S.M.; Schmidt-Ullrich, R.K.; Mikkelsen, R.B. Activation of constitutive nitric oxide synthase activity is an early signaling event induced by ionizing radiation. J. Biol. Chem. 2002, 277, 15400–15406. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Ivanov, V.N.; Gillespie, J.; Geard, C.R.; Amundson, S.A.; Brenner, D.J.; Yu, Z.; Lieberman, H.B.; Hei, T.K. Mechanism of radiation-induced bystander effect: Role of the cyclooxygenase-2 signaling pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 14641–14646. [Google Scholar] [CrossRef] [PubMed]
- Watkins, D.N.; Garlepp, M.; Thompson, P.J. Regulation of the inducible cyclo-oxygenase pathway in human cultured airway epithelial (A549) cells by nitric oxide. Br. J. Pharmacol. 1997, 121, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Chun, K.S.; Cha, H.H. Nitric oxide induces expression of cyclooxygenase-2 in mouse skin through activation of NF-κB. Carcinogenesis 2004, 25, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Hei, T.K.; Zhou, H.; Ivanov, V.N.; Hong, M.; Lieberman, H.B.; Brenner, D.J.; Amundson, S.A.; Geard, C.R. Mechanism of radiation-induced bystander effects: A unifying model. J. Pharm. Pharmacol. 2008, 60, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Brunson, D.; Crespi, C.L.; Penman, B.W.; Wishnok, J.S.; Tannenbaum, S.R. DNA damage and mutation in human cells exposed to nitric oxide in vitro. Proc. Natl. Acad. Sci. USA 1992, 89, 3030–3034. [Google Scholar] [CrossRef] [PubMed]
- Bonavida, B.; Khineche, S.; Huerta-Yepez, S.; Garbán, H. Therapeutic potential of nitric oxide in cancer. Drug Resist. Updates 2006, 9, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Choi, V.W.Y.; Ng, C.Y.P.; Kobayashi, A.; Konishi, T.; Suya, N.; Ishikawa, T.; Cheng, S.H.; Yu, K.N. Bystander effect between zebrafish embryos in vivo induced by high-dose X-rays. Environ. Sci. Technol. 2013, 47, 6368–6376. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Stewart, V.; Folkard, M.; Michael, B.D.; Prise, K.M. Nitric oxide-mediated signaling in the bystander response of individually targeted glioma cells. Cancer Res. 2003, 63, 8437–8442. [Google Scholar] [PubMed]
- Han, W.; Wu, L.; Chen, S.; Bao, L.; Zhang, L.; Jiang, E.; Zhao, Y.; Xu, A.; Hei, T.K.; Yu, Z. Constitutive nitric oxide acting as a possible intercellular signaling molecule in the initiation of radiation-induced DNA double strand breaks in non-irradiated bystander cells. Oncogene 2007, 26, 2330–2339. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Folkard, M.; Michael, B.D.; Prise, K.M. Targeted cytoplasmic irradiation induces bystander responses. Proc. Natl. Acad. Sci. USA 2004, 101, 13495–13500. [Google Scholar] [CrossRef] [PubMed]
- Choi, V.W.Y.; Ng, C.Y.P.; Kobayashi, A.; Konishi, T.; Oikawa, M.; Cheng, S.H.; Yu, P.K.N. Roles of nitric oxide in adaptive response induced in zebrafish embryos in vivo by microbeam protons. J. Radiat. Res. 2014, 55, i114. [Google Scholar] [CrossRef]
- Dhariwala, F.; Narang, H.; Krishna, M. Differential response of three cell types to dual stress of nitric oxide and radiation. Cell Biol. Toxicol. 2012, 28, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Bladen, C.L.; Lam, W.K.; Dynan, W.S.; Kozlowski, D.J. DNA damage response and Ku80 function in the vertebrate embryo. Nucleic Acids Res. 2005, 33, 3002–3010. [Google Scholar] [CrossRef] [PubMed]
- Choi, V.W.Y.; Lam, R.K.K.; Chong, E.Y.W.; Cheng, S.H.; Yu, K.N. Designing experimental setup and procedures for studying alpha-particle-induced adaptive response in zebrafish embryos in vivo. Nucl. Instrum. Methods B 2010, 268, 651–656. [Google Scholar] [CrossRef]
- Choi, V.W.Y.; Yu, K.N. Embryos of the zebrafish Danio rerio in studies of non-targeted effects of ionizing radiation. Cancer Lett. 2015, 356, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Daroczi, B.; Kari, G.; Mcaleer, M.F.; Wolf, J.C.; Rodeck, U.; Dicker, A.P. In vivo radioprotection by the fullerene nanoparticle DF-1 as assessed in a zebrafish model. Clin. Cancer Res. 2006, 12, 7086–7091. [Google Scholar] [CrossRef] [PubMed]
- Geiger, G.A.; Parker, S.E.; Beothy, A.P.; Tucker, J.A.; Mullins, M.C.; Kao, G.D. Zebrafish as a “Biosensor”? Effects of ionzing radiation and amifostine on embryonic viability and development. Cancer Res. 2006, 66, 8172–8181. [Google Scholar] [CrossRef] [PubMed]
- McAleer, M.F.; Davidson, C. Novel use of zebrafish as a vertebrate model to screen radiation protectors and sensitizers. Int. J. Radiat. Oncol. Biol. Phys. 2005, 61, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; Mclaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Barbazuk, W.B.; Korf, I.; Kadavi, C.; Heyen, J.; Tate, S.; Wun, E.; Bedell, J.A.; Mcpherson, J.D.; Johnson, S.L. The syntenic relationship of the zebrafish and human genomes. Genome Res. 2000, 10, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Liebmann, J.; DeLuca, A.M.; Coffin, D.; Keefer, L.K.; Venzon, D.; Wink, D.A.; Mitchell, J.B. In vivo radiation protection by nitric oxide modulation. Cancer Res. 1994, 54, 3365–3368. [Google Scholar] [PubMed]
- Tokumizu, S.; Hori, M.; Monobe, M.; Hosoi, Y.; Kojima, S. Effect of nitric oxide on γ-ray-induced micronucleus frequency in RAW264.7 cells. Radiat. Res. 2005, 164, 723–732. [Google Scholar]
- Suschek, C.; Krischel, V.; Bruch-Gerharz, D.; Berendji, D.; Krutmann, J.; Kröncke, K.D.; Kolb-Bachofen, V. Nitric oxide fully protects against UVA-induced apoptosis in tight correlation with Bcl-2 up-regulation. J. Biol. Chem. 1999, 274, 6130–6137. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zalcenstein, A.; Oren, M. Nitric oxide promotes p53 nuclear retention and sensitizes neuroblastoma cells to apoptosis by ionizing radiation. Cell Death Differ. 2003, 10, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Jourd’heuil, D.; Kang, D.; Grisham, M.B. Interactions between superoxide and nitric oxide: Implications in DNA damage and mutagenesis. Front. Biosci. 1997, 2, 189–196. [Google Scholar]
- Chen, Y.; Stanford, A.; Simmons, R.L.; Ford, H.R.; Hoffman, R.A. Nitric oxide protects thymocytes from γ-irradiation-induced apoptosis in correlation with inhibition of p53 upregulation and mitochondrial damage. Cell. Immunol. 2001, 214, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Takahashi, A.; Guo, G.; Mori, E.; Okamoto, N.; Ohnishi, K.; Iwasaki, T.; Ohnishi, T. Biphasic effects of nitric oxide radicals on radiation-induced lethality and chromosome aberrations in human lung cancer cells carrying different p53 gene status. Int. J. Radiat. Oncol. Biol. Phys. 2010, 77, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Liu, L.; Smith, G.C.M.; Charles, L.G. Nitric oxide upregulates expression of DNA-PKcs to protect cells from DNA-damaging anti-tumour agents. Nat. Cell Biol. 2000, 2, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Hayashi, S.; Hatashita, M.; Shioura, H.; Ohtsubo, T.; Kitai, R.; Ohnishi, T.; Kano, E. Intercellular signaling initiated by nitric oxide produced in heat-shocked human glioblastoma cells. Nitric Oxide 1999, 3, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Hayashi, S.; Hatashita, M.; Ohnishi, K.; Ohtsubo, T.; Kitai, R.; Shioura, H.; Ohnishi, T.; Kano, E. Nitric oxide is an initiator of intercellular signal transduction for stress response after hyperthermia in mutant p53 cells of human glioblastoma. Cancer Res. 1999, 59, 3239–3244. [Google Scholar] [PubMed]
- Gansauge, S.; Nussler, A.K.; Beger, H.G.; Gansauge, F. Nitric oxide-induced apoptosis in human pancreatic carcinoma cell lines is associated with a G1-arrest and an increase of the cyclin-dependent kinase inhibitor p21WAF1/CIP1. Cell Growth Differ. 1998, 9, 611–617. [Google Scholar] [PubMed]
- Mitchell, J.B.; Wink, D.A.; DeGraff, W.; Gamson, J.; Keefer, L.K.; Krishna, M.C. Hypoxic mammalian cell radiosensitization by nitric oxide. Cancer Res. 1993, 53, 5845–5848. [Google Scholar] [PubMed]
- Cook, T.; Wang, Z.; Alber, S.; Liu, K.; Watkins, S.C.; Vodovotz, Y.; Billiar, T.R.; Blumberg, D. Nitric oxide and ionizing radiation synergistically promote apoptosis and growth inhibition of cancer by activating p53. Cancer Res. 2004, 64, 8015–8021. [Google Scholar] [CrossRef] [PubMed]
- Saleem, W.; Suzuki, Y.; Mobaraki, A.; Yoshida, Y.; Noda, S.; Saitoh, J.; Nakano, T. Reduction of nitric oxide level enhances the radiosensitivity of hypoxic non-small cell lung cancer. Cancer Sci. 2011, 102, 2150–2156. [Google Scholar] [CrossRef] [PubMed]
- Kong, E.Y.; Cheng, S.H.; Yu, K.N. Biphasic and triphasic dose responses in zebrafish embryos to low-dose 150 kV X-rays with different hardness. J. Radiat. Res. 2016, 57, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Tucker, B.; Lardelli, M.A. A rapid apoptosis assay measuring relative acridine orange fluorescence in zebrafish embryos. Zebrafish 2007, 4, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Yoshimoto, M.; Maeda, K.; Matsumoto, A.; Maruyama, K.; Ishikawa, Y. Rapid and simple method for quantitative evaluation of neurocytotoxic effects of radiation on developing Medaka brain. J. Radiat. Res. 2008, 49, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Zhang, Q.Y.; Li, Z.; Lin, S.; Gui, J.F. C1q-like inhibits p53-mediated apoptosis and controls normal hematopoiesis during zebrafish embryogenesis. Dev. Biol. 2008, 319, 273–284. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Set | Control | S20 | D2 | p # |
|---|---|---|---|---|
| 1 | 92.5 ± 13.6 (n = 10) | 86.1 ± 11.8 (n = 8) | 126 ± 19 (n = 8) | 0.164 |
| 2 | 83.4 ± 5.4 (n = 7) | 95.2 ± 10.4 (n = 6) | 89.4 ± 7.2 (n = 7) | 0.566 |
| 3 | 173 ± 11 (n = 11) | 156 ± 11 (n = 10) | 173 ± 18 (n = 10) | 0.619 |
| Set | Control | S100 | D10 | p # |
|---|---|---|---|---|
| 1 | 96.0 ± 7.4 (n = 7) | 108 ± 10 (n = 9) | 110 ± 15 (n = 9) | 0.689 |
| 2 | 65.5 ± 5.1 (n = 12) | 64.5 ± 6.0 (n = 12) | 62.7 ± 6.2 (n = 12) | 0.941 |
| 3 | 87.5 ± 5.4 (n = 12) | 84.5 ± 6.1 (n = 11) | 86.6 ± 5.0 (n = 10) | 0.922 |
| Set | Control | IS20 | ID2 | p # |
|---|---|---|---|---|
| 1 | 87.9 ± 6.4 (n = 7) | 186 ± 13 (n = 9) | 202 ± 19 (n = 9) | 0.517 |
| 2 | 108 ± 5 (n = 10) | 196 ± 12 (n = 11) | 198 ± 14 (n = 11) | 0.898 |
| 3 | 83.4 ± 5.4 (n = 7) | 136 ± 13 (n = 8) | 157 ± 11 (n = 7) | 0.236 |
| Set | Control | IS100 | ID10 | p # |
|---|---|---|---|---|
| 1 | 74.3 ± 4.3 (n = 10) | 157 ± 28 (n = 10) | 250 ± 37 (n = 9) | 4.65 × 10−3 * |
| 2 | 108 ± 5 (n = 10) | 177 ± 8 (n = 10) | 210 ± 10 (n = 11) | 9.69 × 10−3 * |
| 3 | 110 ± 9 (n = 7) | 131 ± 9 (n = 10) | 204 ± 13 (n = 10) | 1.99 × 10−4 * |
| Set | Control | BS20 | BD2 | p # |
|---|---|---|---|---|
| 1 | 82.7 ± 10.4 (n = 10) | 72.4 ± 7.2 (n = 9) | 154 ± 15 (n = 9) | 1.65 × 10−4 * |
| 2 | 102 ± 11 (n = 10) | 91.4 ± 7.1 (n = 8) | 137 ± 12 (n = 9) | 2.78 × 10−3 * |
| 3 | 63.0 ± 2.9 (n = 9) | 67.3 ± 4.3 (n = 9) | 95.3 ± 7.6 (n = 8) | 3.46 × 10−3 * |
| Set | Control | BS100 | BD10 | p # |
|---|---|---|---|---|
| 1 | 129 ± 15 (n = 10) | 113 ± 12 (n = 9) | 239 ± 21 (n = 9) | 7.14 × 10−5 * |
| 2 | 103 ± 14 (n = 8) | 104 ± 14 (n = 9) | 156 ± 15 (n = 9) | 9.77 × 10−3 * |
| 3 | 96.0 ± 7.4 (n = 7) | 87.6 ± 6.8 (n = 8) | 129 ± 15 (n = 8) | 1.67 × 10−2 * |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, E.Y.; Yeung, W.K.; Chan, T.K.Y.; Cheng, S.H.; Yu, K.N. Exogenous Nitric Oxide Suppresses in Vivo X-ray-Induced Targeted and Non-Targeted Effects in Zebrafish Embryos. Int. J. Mol. Sci. 2016, 17, 1321. https://doi.org/10.3390/ijms17081321
Kong EY, Yeung WK, Chan TKY, Cheng SH, Yu KN. Exogenous Nitric Oxide Suppresses in Vivo X-ray-Induced Targeted and Non-Targeted Effects in Zebrafish Embryos. International Journal of Molecular Sciences. 2016; 17(8):1321. https://doi.org/10.3390/ijms17081321
Chicago/Turabian StyleKong, E.Y., W.K. Yeung, T.K.Y. Chan, S.H. Cheng, and K.N. Yu. 2016. "Exogenous Nitric Oxide Suppresses in Vivo X-ray-Induced Targeted and Non-Targeted Effects in Zebrafish Embryos" International Journal of Molecular Sciences 17, no. 8: 1321. https://doi.org/10.3390/ijms17081321
APA StyleKong, E. Y., Yeung, W. K., Chan, T. K. Y., Cheng, S. H., & Yu, K. N. (2016). Exogenous Nitric Oxide Suppresses in Vivo X-ray-Induced Targeted and Non-Targeted Effects in Zebrafish Embryos. International Journal of Molecular Sciences, 17(8), 1321. https://doi.org/10.3390/ijms17081321