
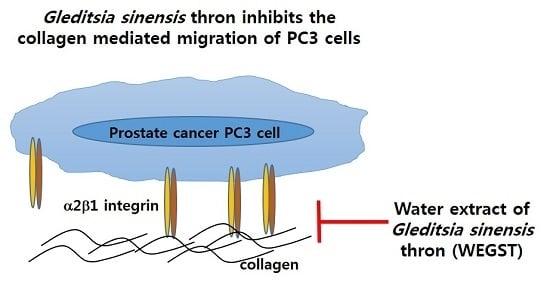
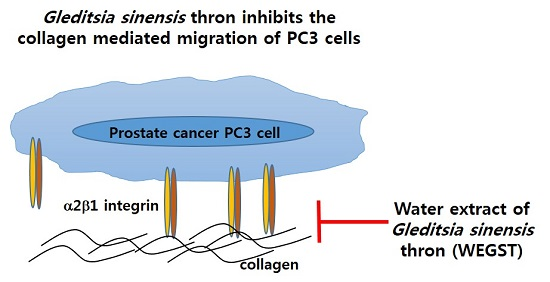
Gleditsia sinensis Thorn Attenuates the Collagen-Based Migration of PC3 Prostate Cancer Cells through the Suppression of α2β1 Integrin Expression



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
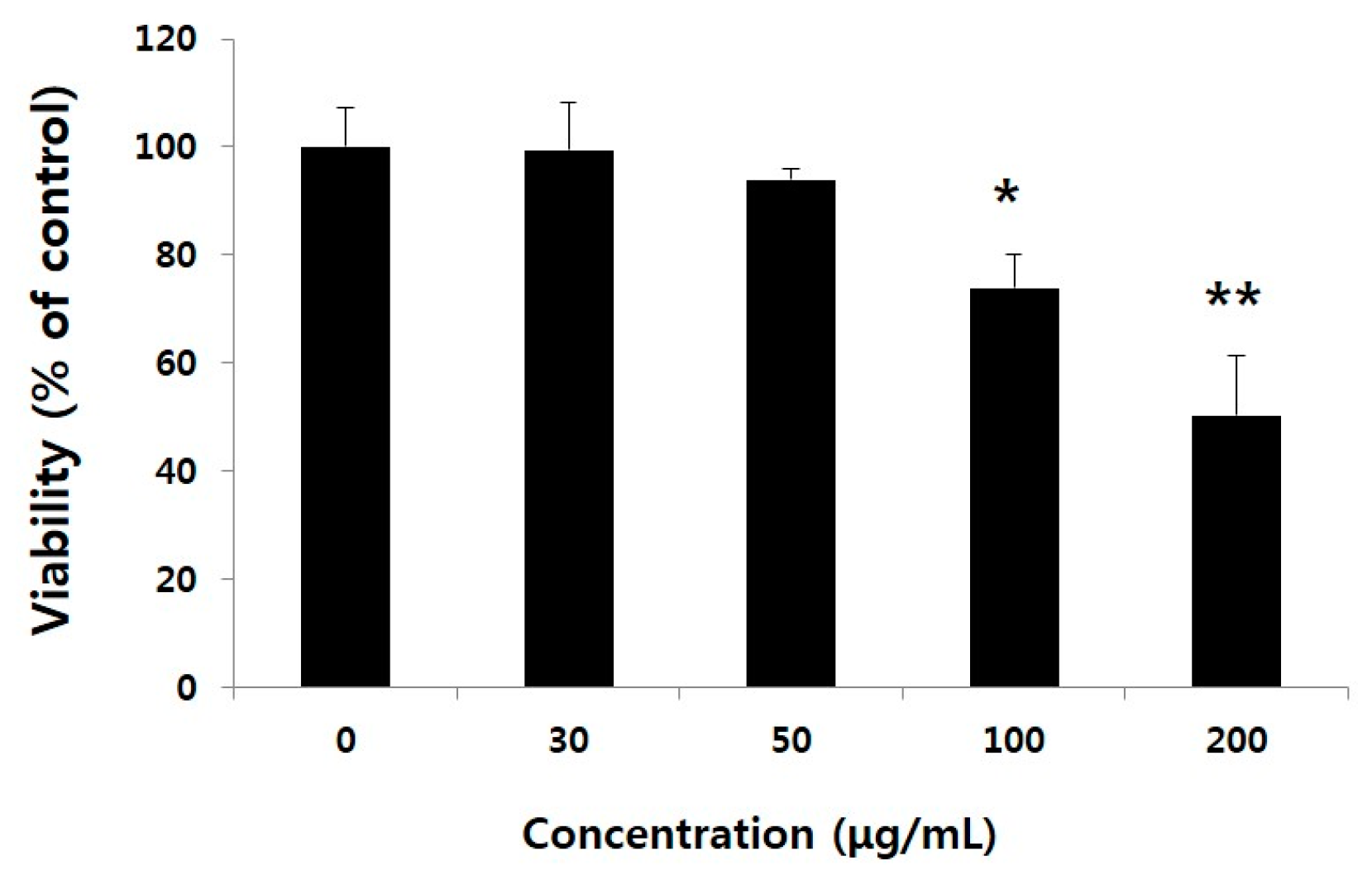
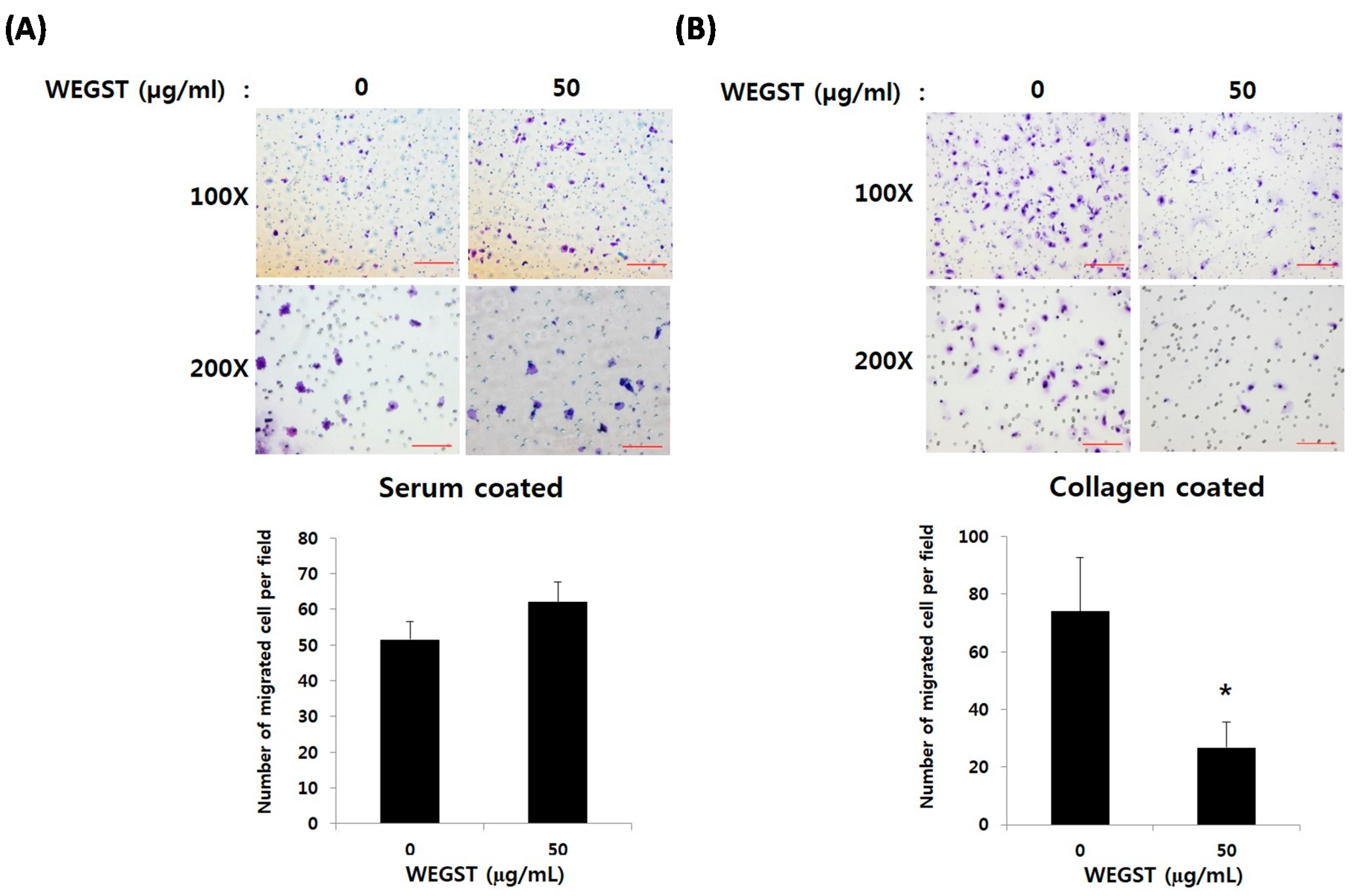
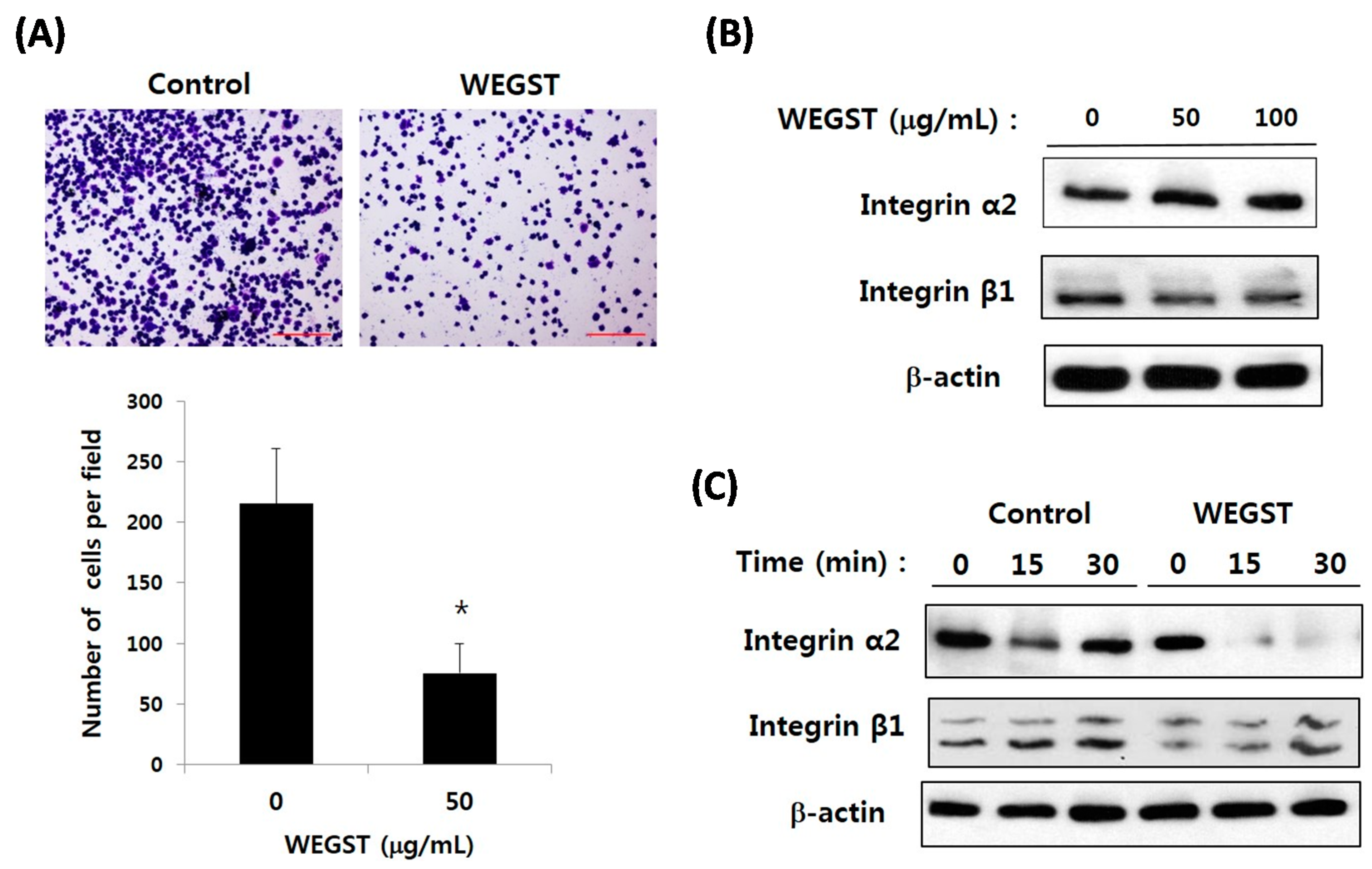
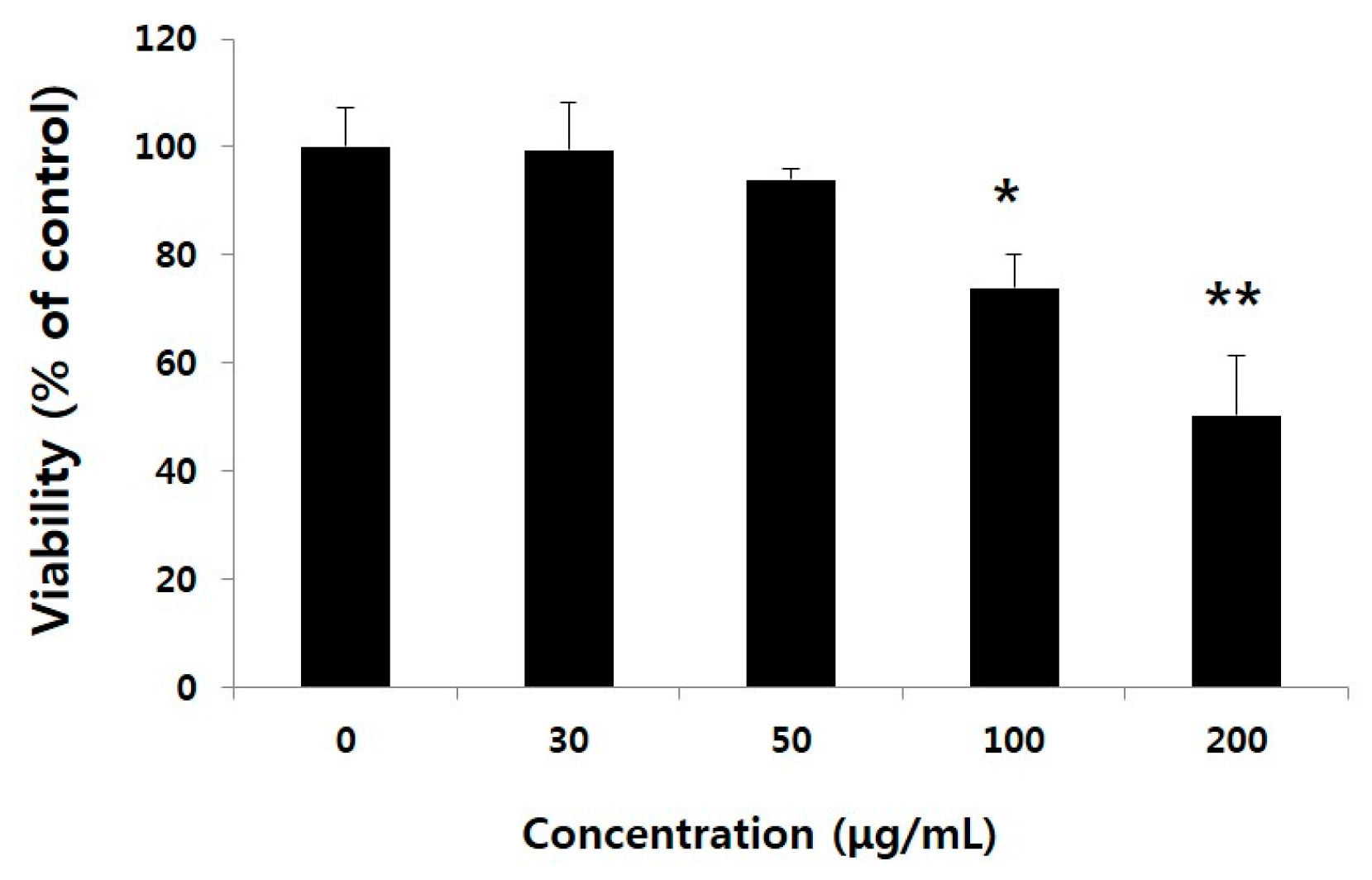
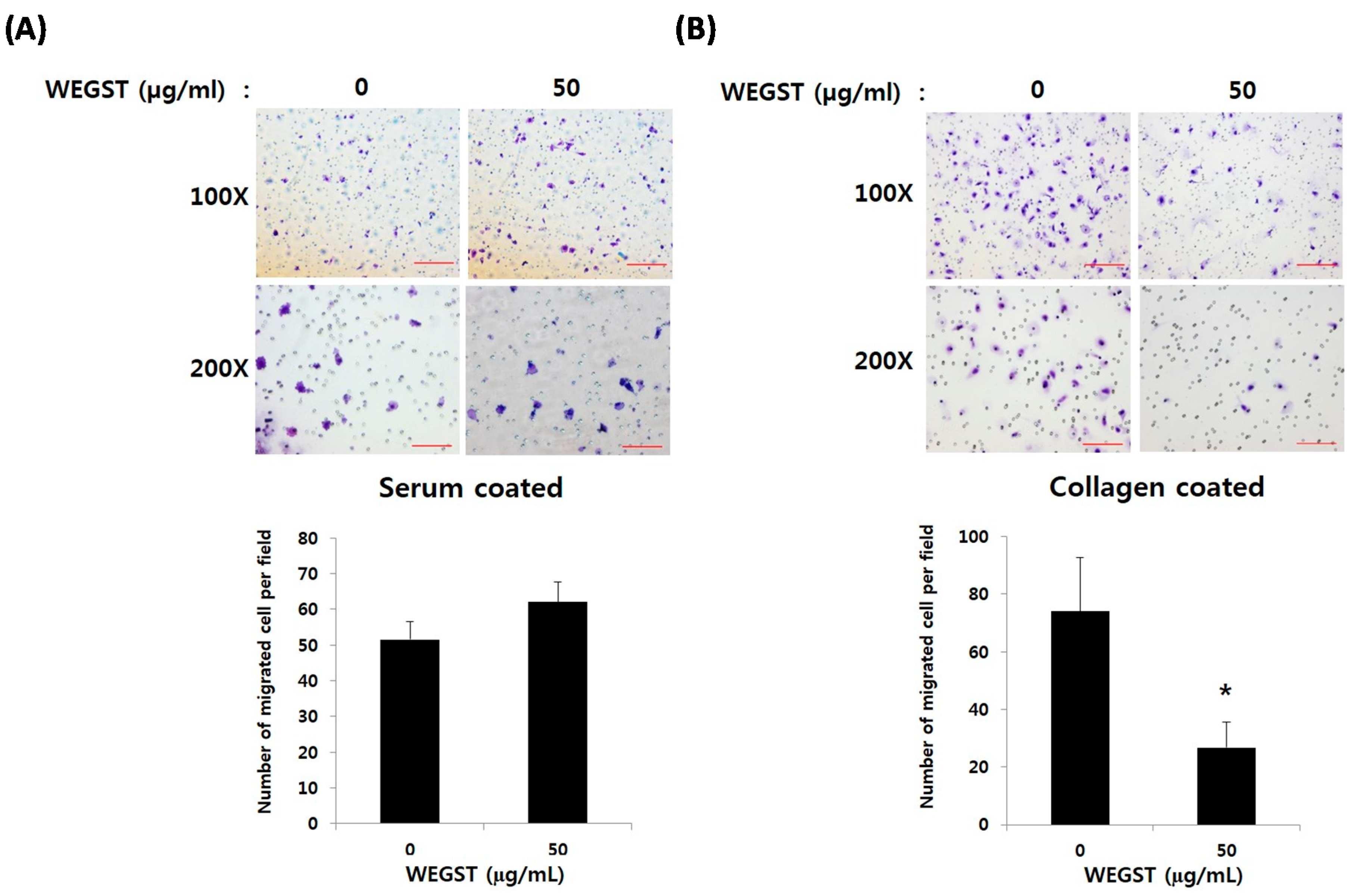
2.1. Non-Toxic Levels of Water Extract of GST (WEGST) Are Effectively Attenuated by the Collagen-Mediated Migration of PC3 Prostate Cancer Cells
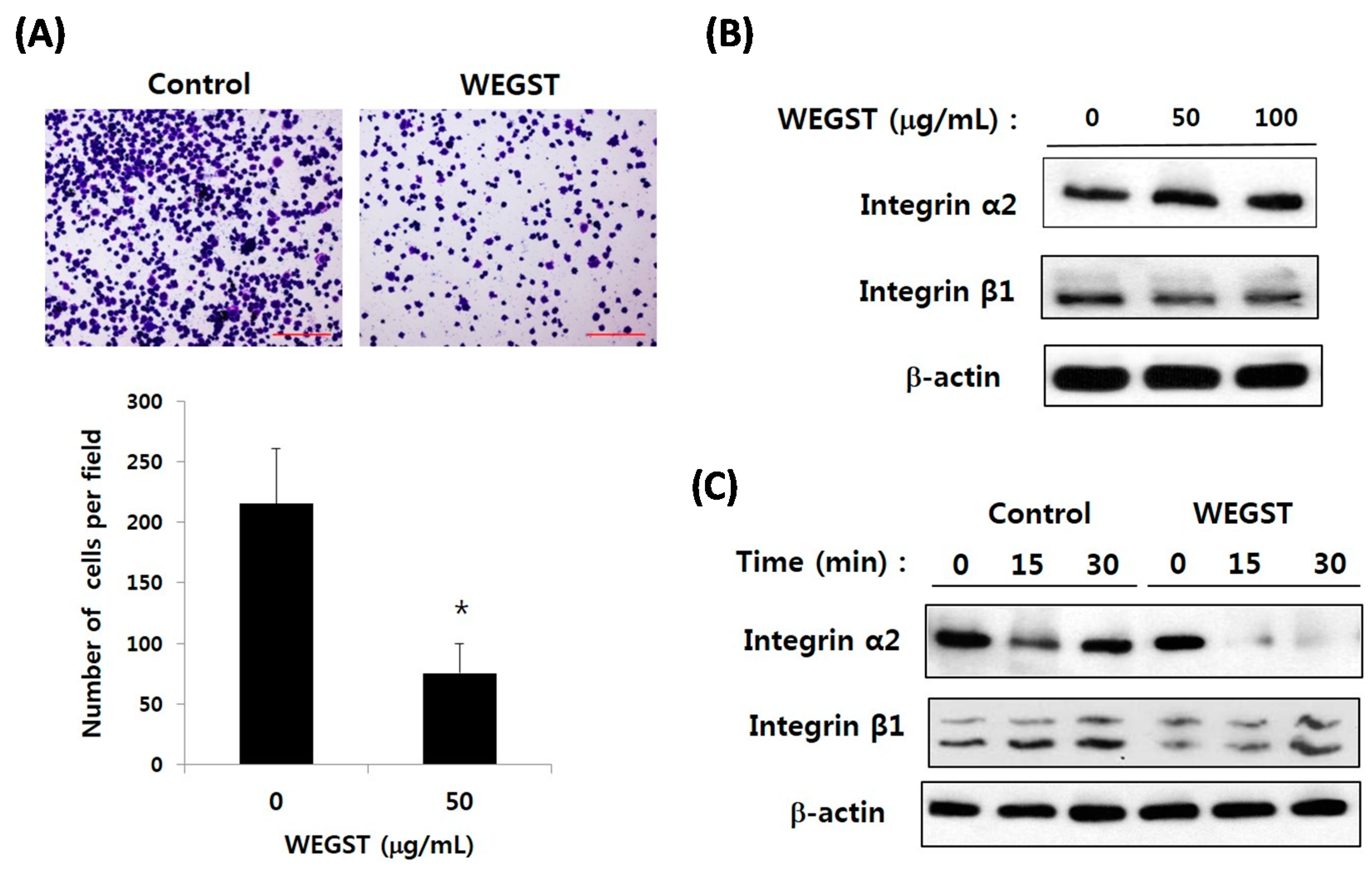
2.2. Collagen-Mediated Expression of α2 Integrin in PC3 Prostate Cancer Cells Is Attenuated by the Administration of WEGST
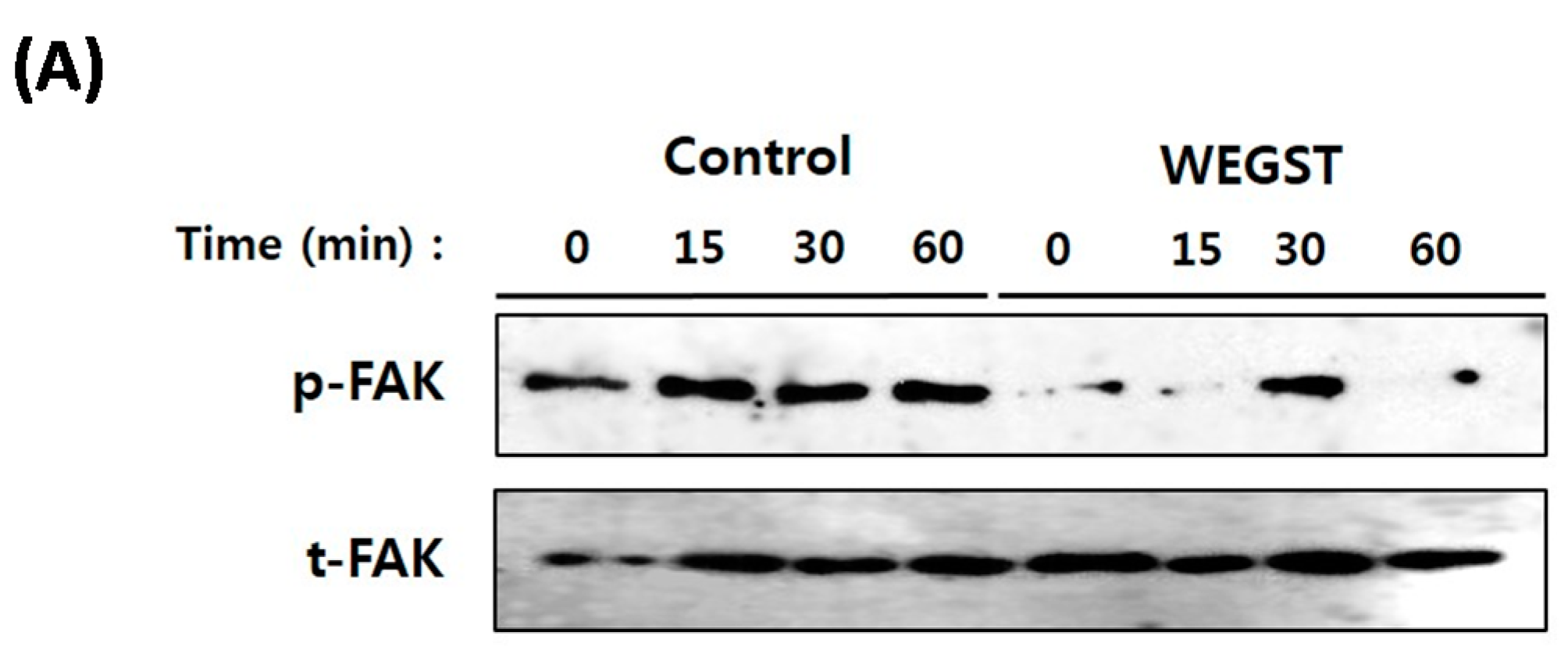
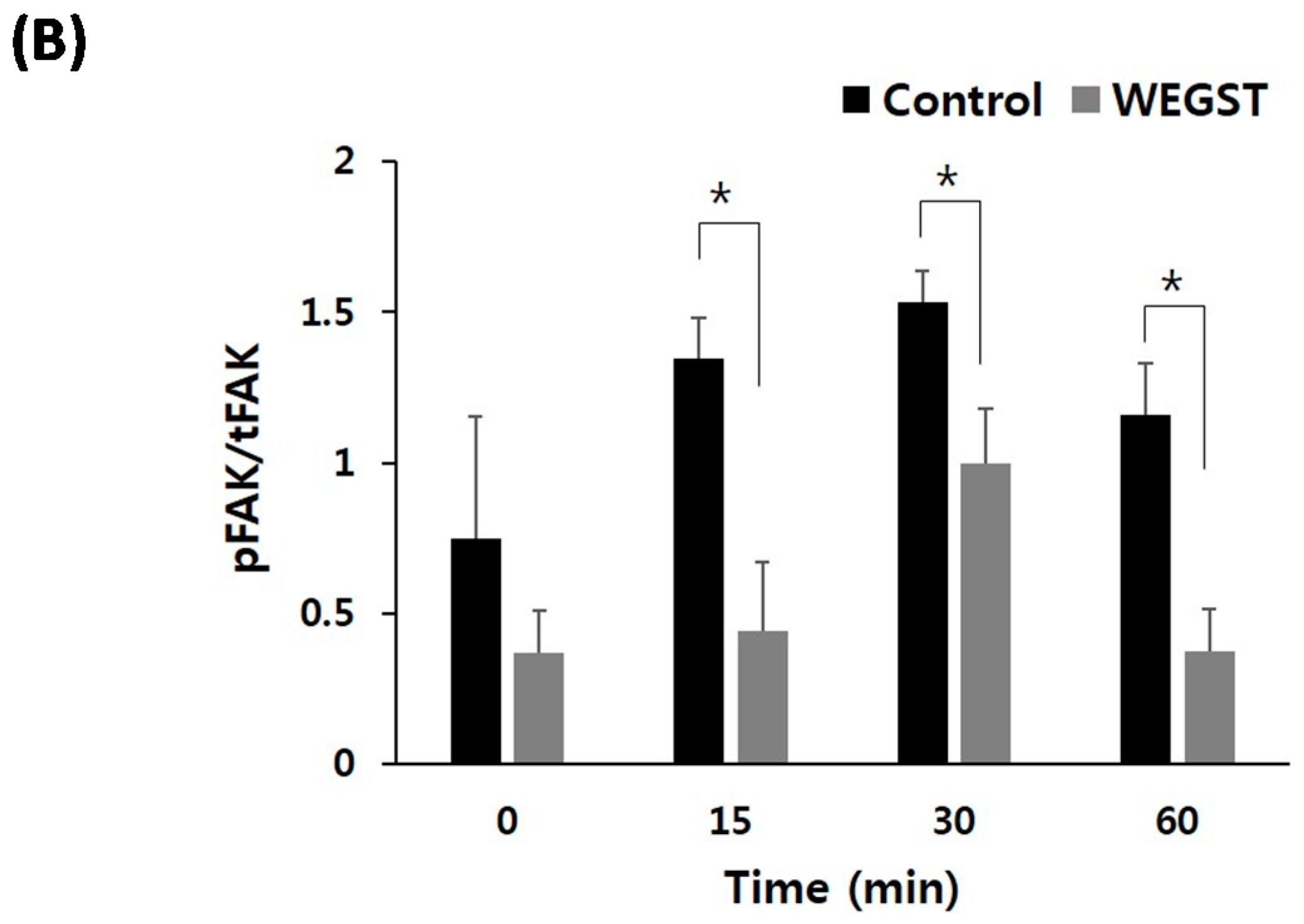
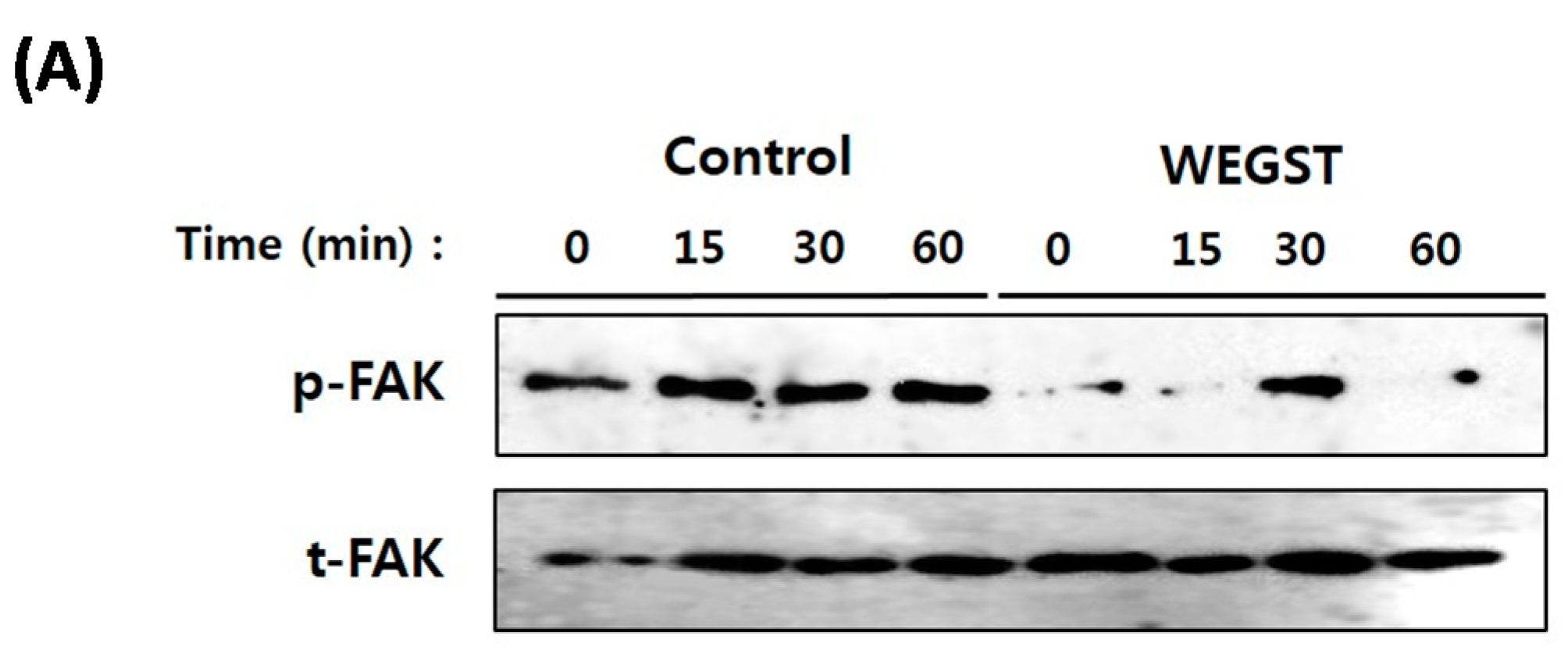
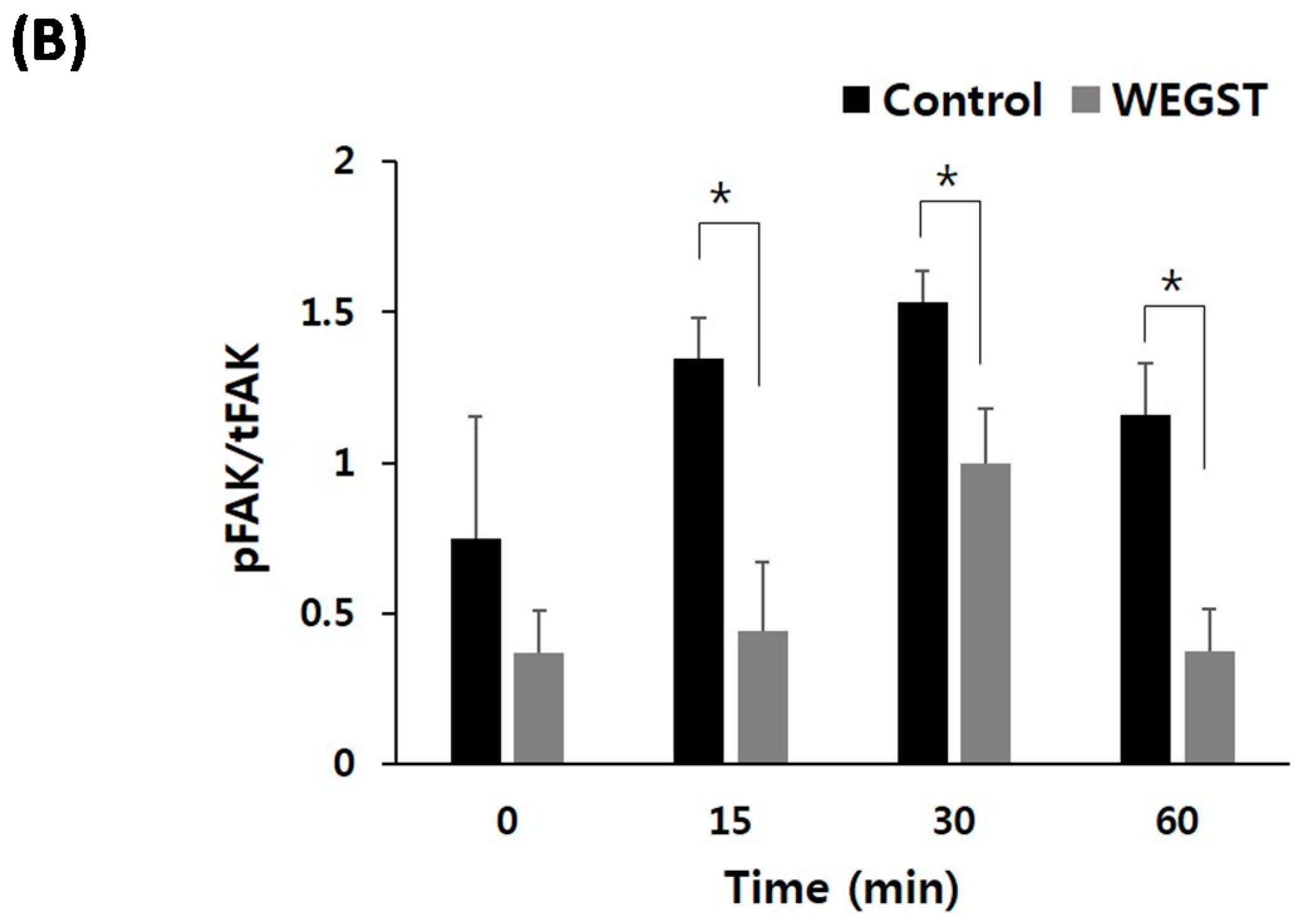
2.3. Collagen-Mediated Intracellular Focal Adhesion Kinase (FAK) Signaling of PC3 Prostate Cancer Cells Is Inhibited by Administration of WEGST
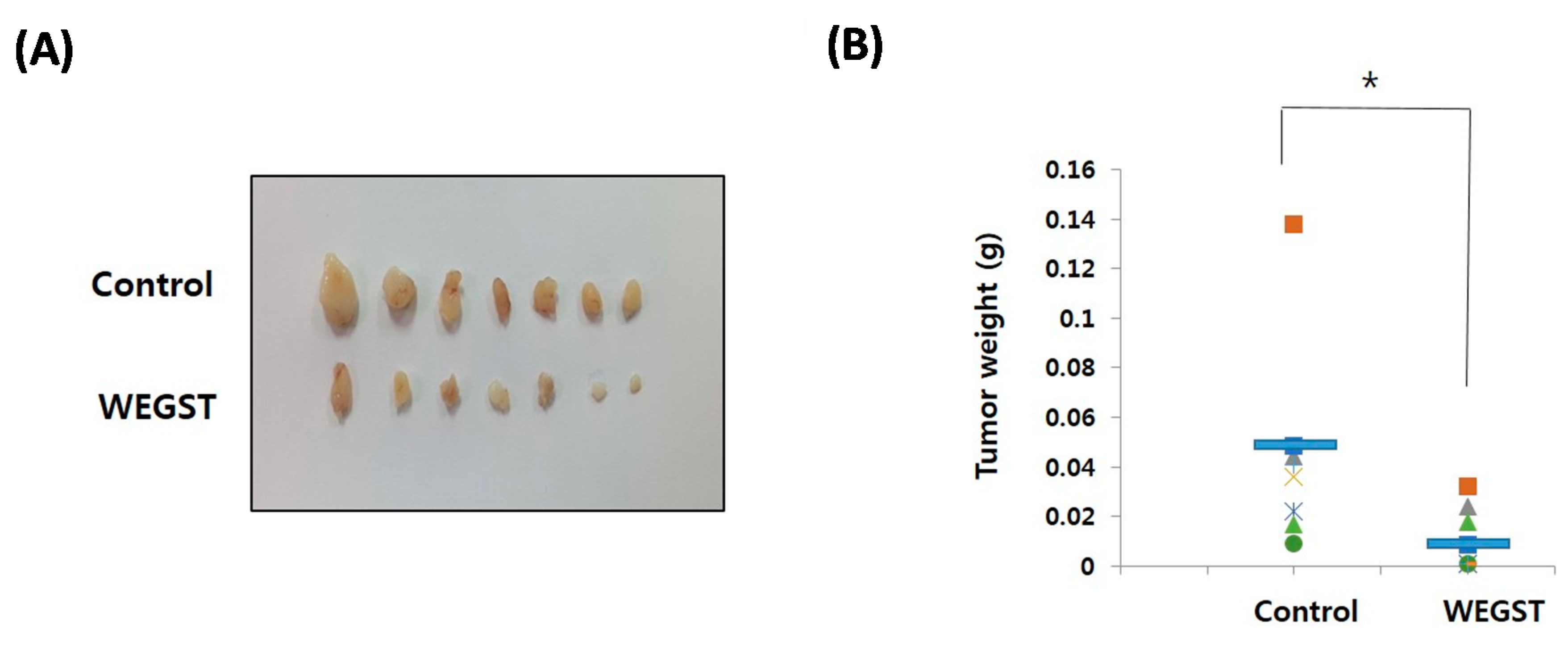
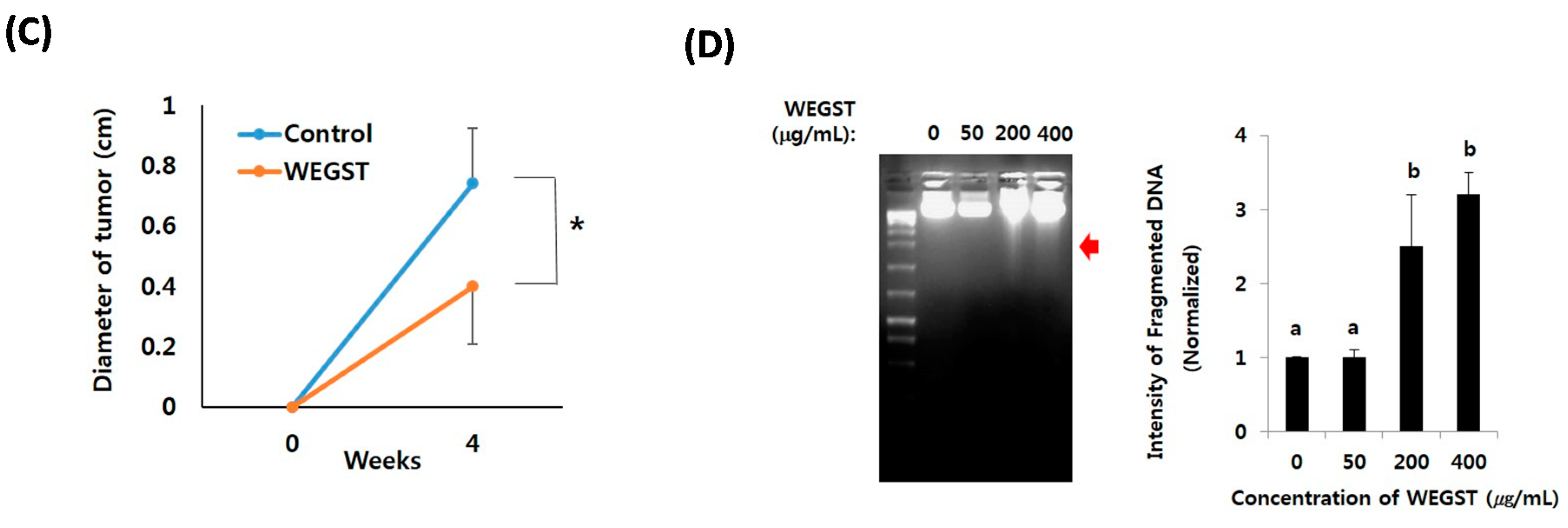
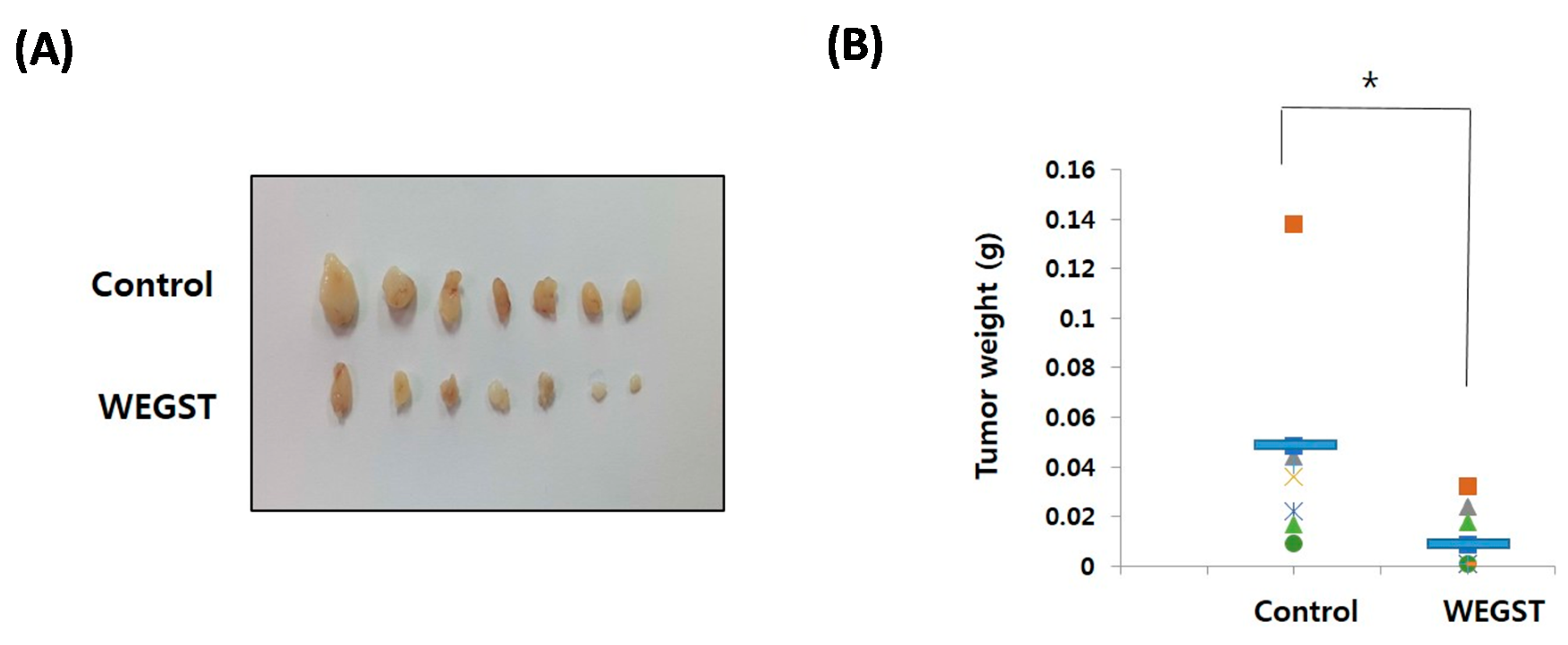
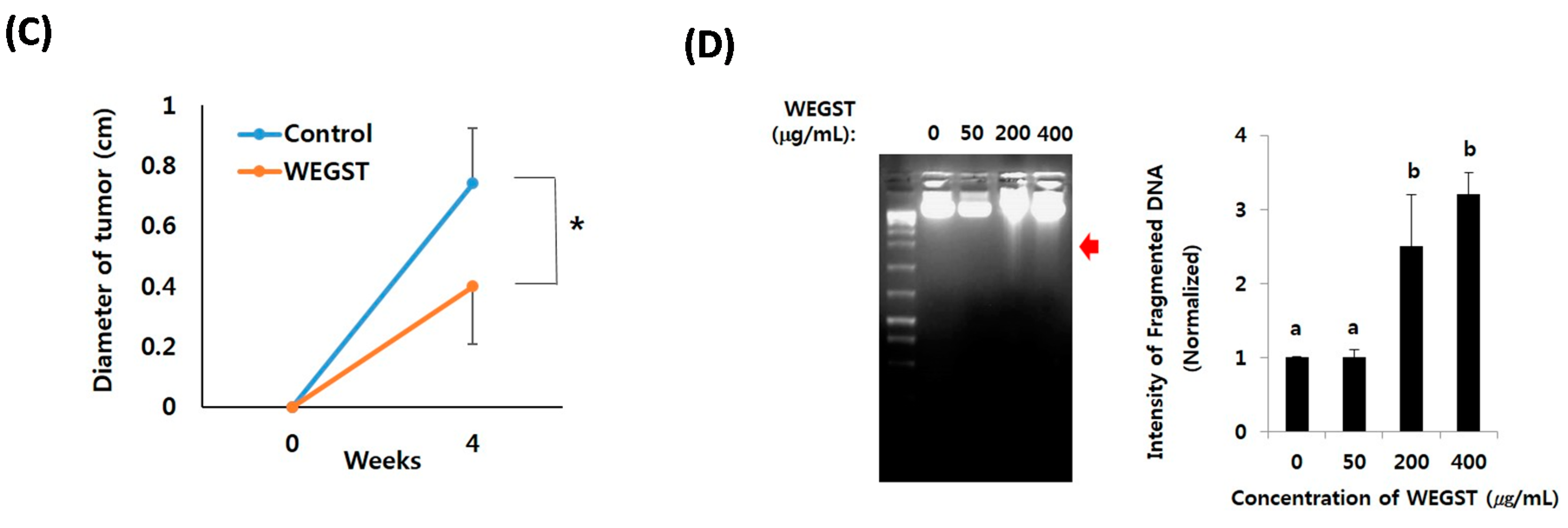
2.4. Oral Administration of WEGST Inhibits in Vivo Tumor Formation
3. Discussion
4. Materials and Methods
4.1. Preparation of WEGST
4.2. Cell Culture and Cytotoxicity Assay
4.3. Collagen against Migration Assay
4.4. Cell Adhesion Assay
4.5. Analysis of Collagen against Expression of α2β1 Integrin
4.6. Analysis of FAK Phosphorylation
4.7. DNA Fragmentation Assay
4.8. Xenograft Tumor Assay
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer statistics. CA Cancer J. Clin. 2010, 60, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Wegiel, B.; Evans, S.; Hellsten, R.; Otterbein, L.E.; Bjartell, A.; Persson, J.L. Molecular pathways in the progression of hormone-independent and metastatic prostate cancer. Curr. Cancer Drug Targets 2010, 10, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer. 2010, 10, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Morgan, M.R.; Humphries, M.J.; Bass, M.D. Synergistic control of cell adhesion by integrins and syndecans. Nat. Rev. Mol. Cell Biol. 2007, 8, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Emsley, J.; Knight, C.G.; Farndale, R.W.; Barnes, M.J.; Liddington, R.C. Structural basis of collagen recognition by integrin α2β1. Cell 2000, 101, 47–56. [Google Scholar] [CrossRef]
- Lee, S.H.; Hatakeyama, S.; Yu, S.Y.; Bao, X.; Ohyama, C.; Khoo, K.H.; Fukuda, M.N.; Fukuda, M. Core3 O-glycan synthase suppresses tumor formation and metastasis of prostate carcinoma PC3 and LNCaP cells through down-regulation of α2β1 integrin complex. J. Biol. Chem. 2009, 284, 17157–17169. [Google Scholar] [CrossRef] [PubMed]
- Ahn, D.K. Illustrated Book of Korean Medicinal Herbs; Kyohak Publishing Co.: Seoul, Korea, 2003; p. 628. [Google Scholar]
- Dai, Y.; Chan, Y.P.; Chu, L.M.; Bu, P.P. Antiallergic and anti-inflammatory properties of the ethanolic extract from Gleditsia sinensis. Biol. Pharm. Bull. 2002, 25, 1179–1182. [Google Scholar] [CrossRef] [PubMed]
- Lai, P.; Du, J.R.; Zhang, M.X.; Kuang, X.; Li, Y.J.; Chen, Y.S.; He, Y. Aqueous extract of Gleditsia sinensis Lam. fruits improves serum and liver lipid profiles and attenuates atherosclerosis in rabbits fed a high-fat diet. J. Ethnopharm. 2011, 137, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.Y.; Zhang, Z. Quantitive analysis of Gleditsia saponins in the fruits of Gleditsia sinensis Lam. by high performance liquid chromatography. J. Pharm. Biomed. Anal. 2013, 75, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Yi, J.M.; Kim, H.; Lee, Y.J.; Park, J.S.; Bang, O.S.; Kim, N.S. Cytochalasin H, an active anti-angiogenic constituent of the ethanol extract of Gleditsia sinensis thorns. Biol. Pharm. Bull. 2014, 37, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.M.; Park, J.S.; Oh, S.M.; Lee, J.; Kim, J.; Oh, D.S.; Bang, O.S.; Kim, N.S. Ethanol extract of Gleditsia sinensis thorn suppresses angiogenesis in vitro and in vivo. BMC Complement. Altern. Med. 2012, 12. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Cho, Y.H.; Kim, H.; Park, K.; Park, S.K.; Ha, S.D.; Kim, W.J.; Moon, S.K. Inhibitory effects of the ethanol extract of Gleditsia sinensis thorns on human colon cancer HCT116 cells in vitro and in vivo. Oncol. Rep. 2009, 22, 1505–1512. [Google Scholar] [PubMed]
- Lee, S.J.; Park, K.; Ha, S.D.; Kim, W.J.; Moon, S.K. Gleditsia sinensis thorn extract inhibits human colon cancer cells: The role of ERK1/2, G2/M-phase cell cycle arrest and p53 expression. Phytother. Res. PTR 2010, 24, 1870–1876. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.M.; Kim, J.; Park, J.S.; Lee, J.; Lee, Y.J.; Hong, J.T.; Bang, O.S.; Kim, N.S. In vivo anti-tumor effects of the ethanol extract of Gleditsia sinensis thorns and its active constituent, cytochalasin H. Biol. Pharm. Bull. 2015, 38, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Bonaccorsi, L.; Carloni, V.; Muratori, M.; Salvadori, A.; Giannini, A.; Carini, M.; Serio, M.; Forti, G.; Baldi, E. Androgen receptor expression in prostate carcinoma cells suppresses α6β4 integrin-mediated invasive phenotype. Endocrinology 2000, 141, 3172–3182. [Google Scholar] [PubMed]
- Li, W.H.; Zhang, X.M.; Tian, R.R.; Zheng, Y.T.; Zhao, W.M.; Qiu, M.H. A new anti-HIV lupane acid from Gleditsia sinensis Lam. J. Asian Nat. Prod. Res. 2007, 9, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, D.; Wang, J.; Liu, Y.; Wu, J. Antibacterial phenolic compounds from the spines of Gleditsia sinensis Lam. Nat. Prod. Res. 2007, 21, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.C.; Park, J.H.; Budesinsky, M.; Kasal, A.; Han, Y.H.; Koo, B.S.; Lee, S.I.; Lee, D.U. Antimutagenic constituents from the thorns of Gleditsia sinensis. Chem. Pharm. Bull. 2005, 53, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Kinouchi, Y.; Wada, S.; Tokuda, H. Potential anti-tumor promoting activity of lupane-type triterpenoids from the stem bark of Glochidion zeylanicum and Phyllanthus flexuosus. Planta Med. 2004, 70, 1234–1236. [Google Scholar] [CrossRef] [PubMed]
- Mutai, C.; Abatis, D.; Vagias, C.; Moreau, D.; Roussakis, C.; Roussis, V. Cytotoxic lupane-type triterpenoids from Acacia mellifera. Phytochemistry 2004, 65, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.X.; Wang, M.; Yuan, J.X.; Liu, J.C. Effect of ethyl gallate on invasion abilities and its mechanism of breast cancer MDA-MB-231 cells. Yao Xue Xue Bao 2015, 50, 45–49. [Google Scholar] [PubMed]
- Ali, H.; Dixit, S.; Ali, D.; Alqahtani, S.M.; Alkahtani, S.; Alarifi, S. Isolation and evaluation of anticancer efficacy of stigmasterol in a mouse model of DMBA-induced skin carcinoma. Drug Des. Dev. Ther. 2015, 28, 2793–2800. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Ryu, D.H.; Jang, L.C.; Cho, S.C.; Kim, W.J.; Moon, S.K. Suppressive effects of an ethanol extract of Gleditsia sinensis thorns on human SNU-5 gastric cancer cells. Oncol. Rep. 2013, 29, 1609–1616. [Google Scholar] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, S.; Park, K.M.; Lee, S.H. Gleditsia sinensis Thorn Attenuates the Collagen-Based Migration of PC3 Prostate Cancer Cells through the Suppression of α2β1 Integrin Expression. Int. J. Mol. Sci. 2016, 17, 328. https://doi.org/10.3390/ijms17030328
Ryu S, Park KM, Lee SH. Gleditsia sinensis Thorn Attenuates the Collagen-Based Migration of PC3 Prostate Cancer Cells through the Suppression of α2β1 Integrin Expression. International Journal of Molecular Sciences. 2016; 17(3):328. https://doi.org/10.3390/ijms17030328
Chicago/Turabian StyleRyu, Sujin, Ki Moon Park, and Seung Ho Lee. 2016. "Gleditsia sinensis Thorn Attenuates the Collagen-Based Migration of PC3 Prostate Cancer Cells through the Suppression of α2β1 Integrin Expression" International Journal of Molecular Sciences 17, no. 3: 328. https://doi.org/10.3390/ijms17030328
APA StyleRyu, S., Park, K. M., & Lee, S. H. (2016). Gleditsia sinensis Thorn Attenuates the Collagen-Based Migration of PC3 Prostate Cancer Cells through the Suppression of α2β1 Integrin Expression. International Journal of Molecular Sciences, 17(3), 328. https://doi.org/10.3390/ijms17030328