
Can Clethra barbinervis Distinguish Nickel and Cobalt in Uptake and Translocation?

Abstract
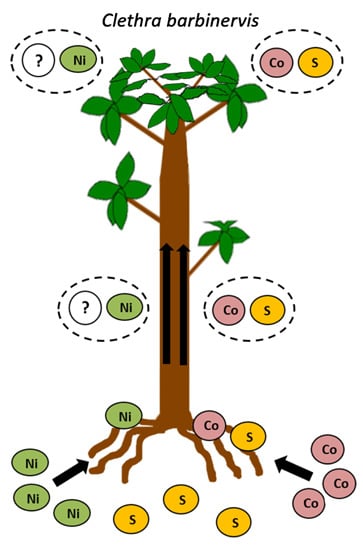
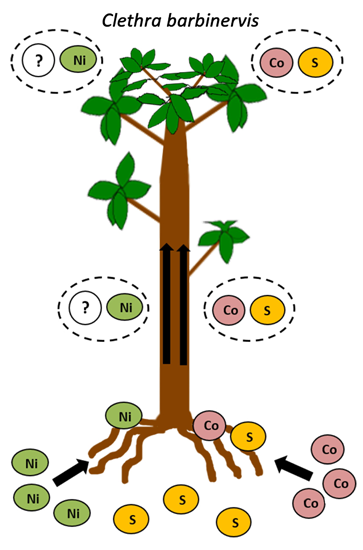
:
1. Introduction
2. Results
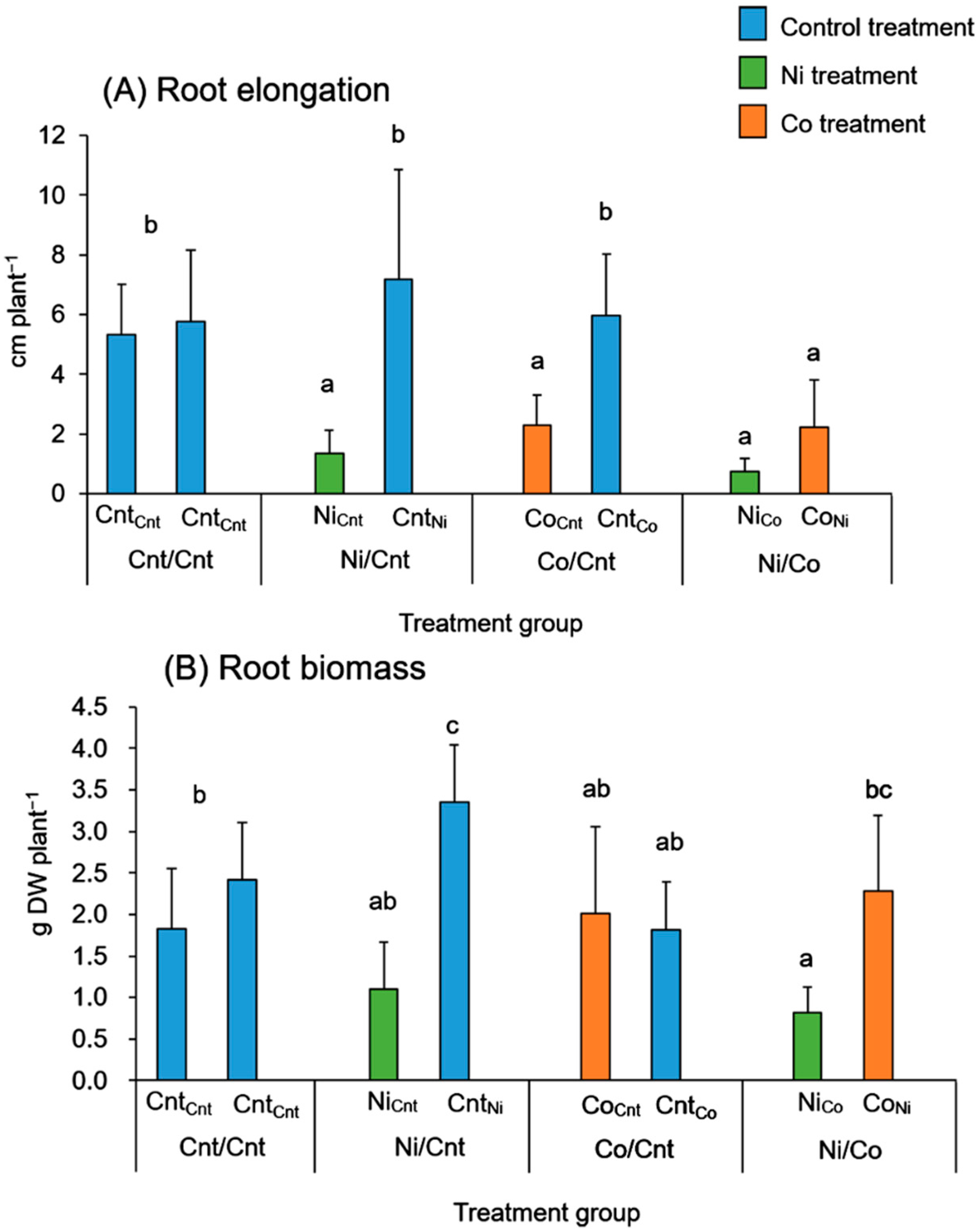
2.1. Plant Growth and Photosynthesis
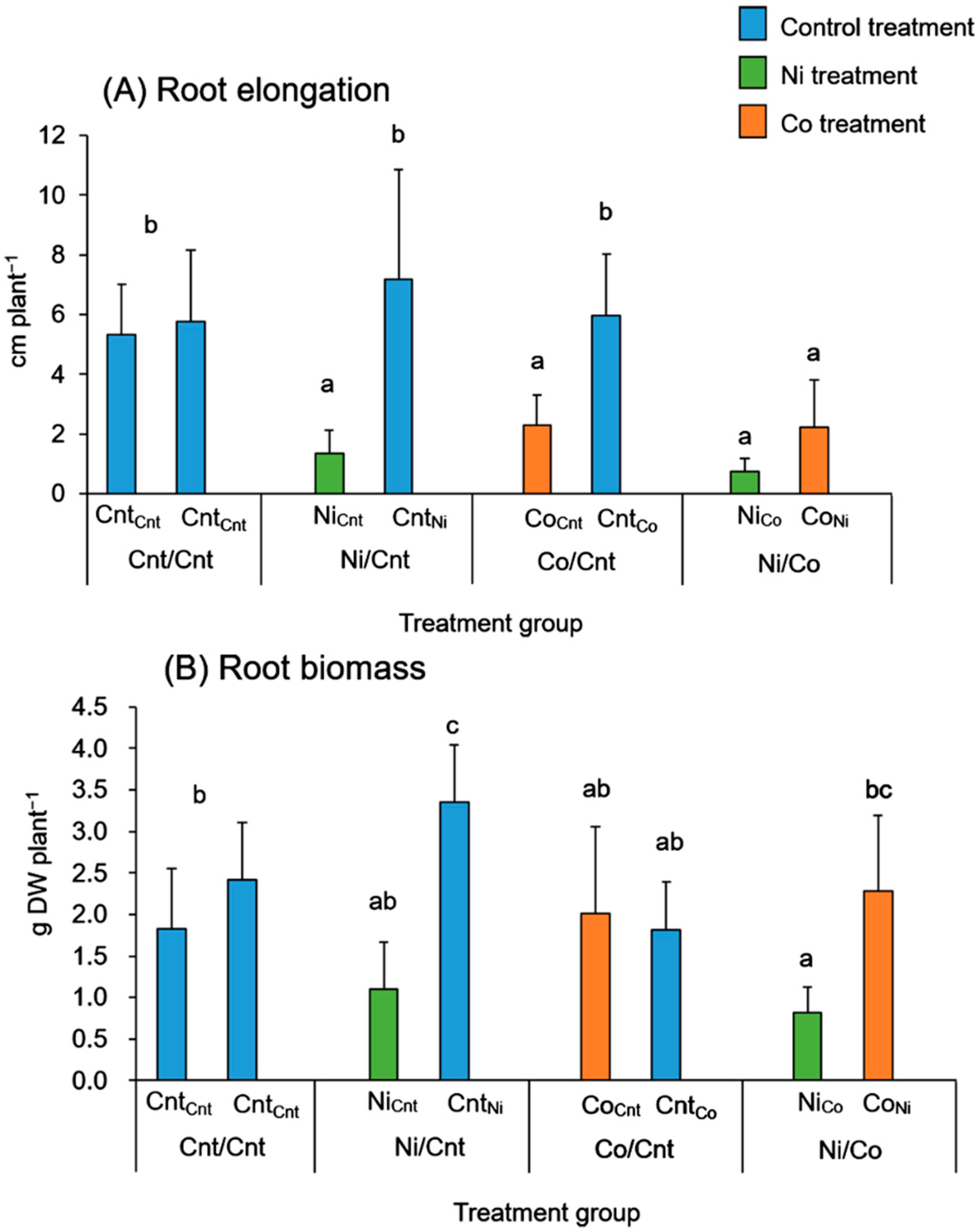

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Leaves Biomass | Net Assimilation Rates |
|---|---|---|
| g Plant−1 DW | µmol m−2 s−1 | |
| Cnt/Cnt | 8.11 ± 2.08 | 7.38 ± 2.6 |
| Ni/Cnt | 6.38 ± 1.18 | 6.47 ± 2.4 |
| Co/Cnt | 7.72 ± 0.59 | 7.55 ± 1.8 |
| Ni/Co | 6.13 ± 2.26 | 7.22 ± 2.2 |
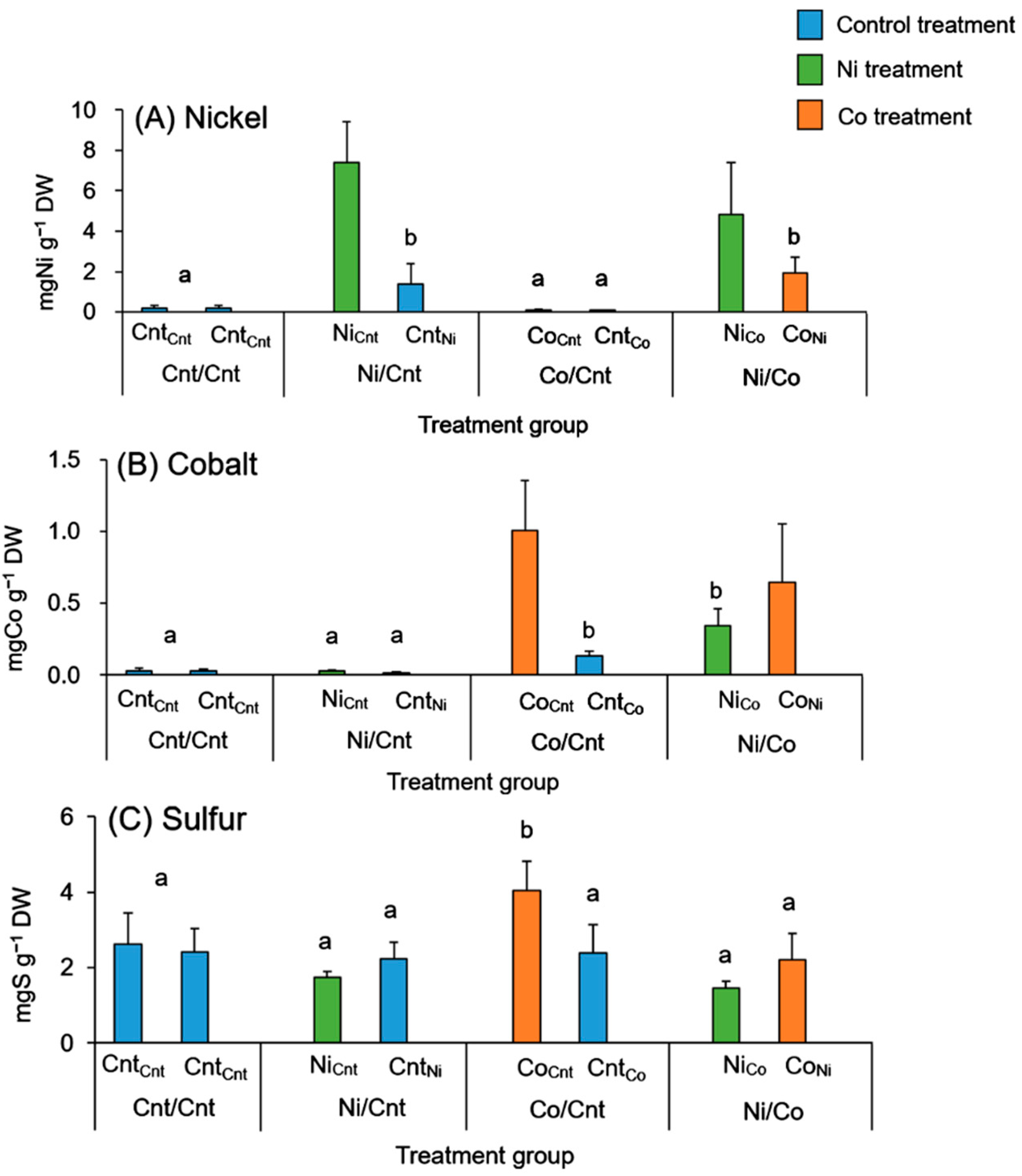
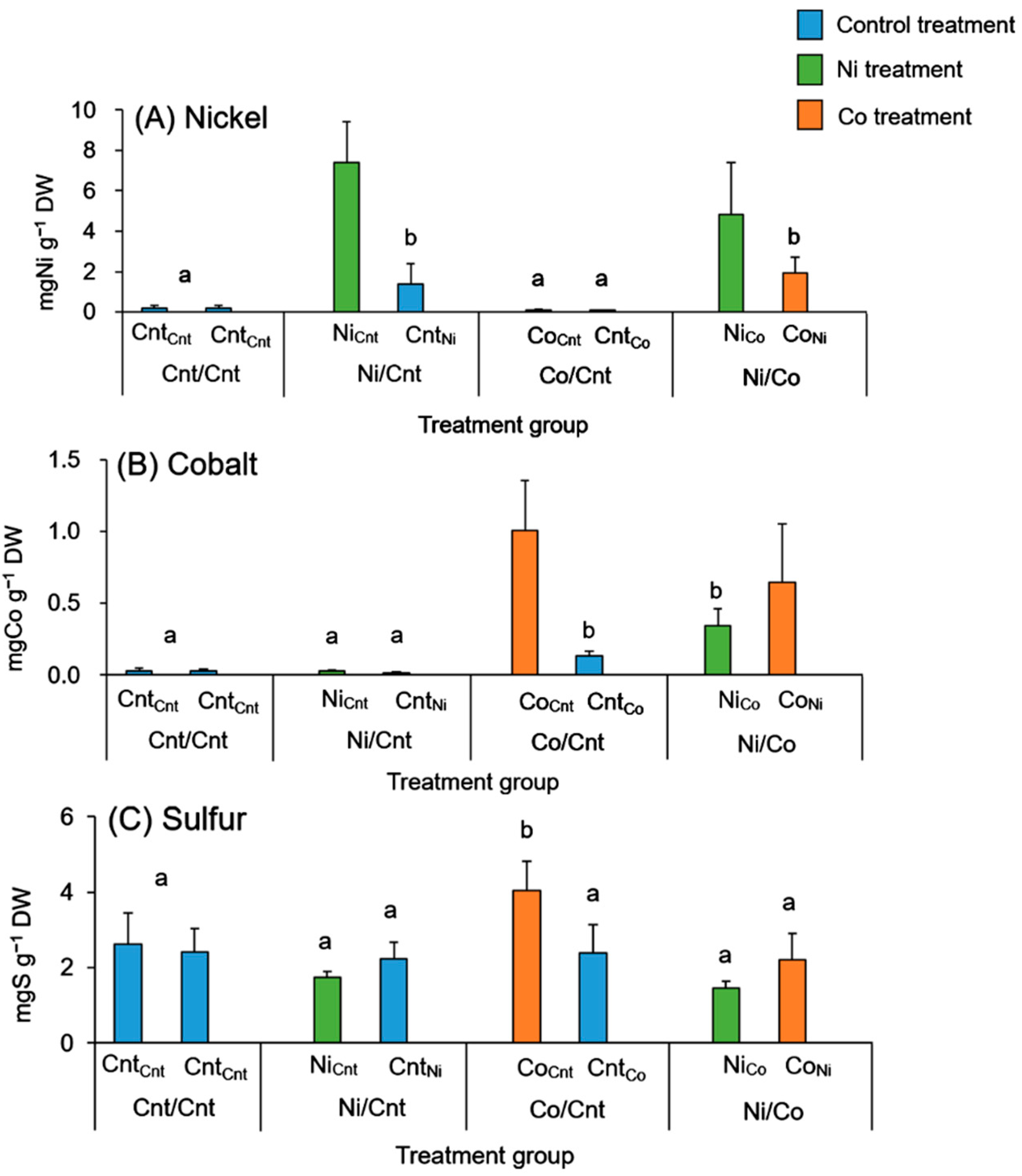
2.2. Ni, Co, and S Accumulation in Roots and Leaves
| Plant Tissues | Treatment | Ni | Co | S |
|---|---|---|---|---|
| µg g−1 DW | µg g−1 DW | µg g−1 DW | ||
| Leaves | Cnt/Cnt | 0.81 ± 0.07 a | 0.16 ± 0.12 a | 2240 ± 632 a |
| Ni/Cnt | 257 ± 134 b | 0.83 ± 0.69 a | 1890 ± 357 a | |
| Co/Cnt | 1.30 ± 0.74 a | 697 ± 324 b | 4040 ± 1510 b | |
| Ni/Co | 411 ± 263 b | 635 ± 257 b | 2820 ± 883 ab | |
| Barks | Cnt/Cnt | 3.62 ± 2.98 a | 2.15 ± 0.52 a | 904 ± 241 a |
| Ni/Cnt | 117 ± 65.0 b | 0.58 ± 0.43 a | 686 ± 82.4 a | |
| Co/Cnt | 3.16 ± 0.91 a | 50.6 ± 7.07 b | 792 ± 113 a | |
| Ni/Co | 151 ± 92.6 b | 81.5 ± 32.9 c | 844 ± 136 a | |
| Woods | Cnt/Cnt | 2.70 ± 0.68 a | 0.53 ± 0.34 a | 752 ± 208 a |
| Ni/Cnt | 42.3 ± 17.2 b | 0.13 ± 0.22 a | 704 ± 114 a | |
| Co/Cnt | 2.41 ± 1.23 a | 11.8 ± 2.95 b | 751 ± 275 a | |
| Ni/Co | 47.3 ± 15.1 b | 19.5 ± 6.06 c | 850 ± 200 a |
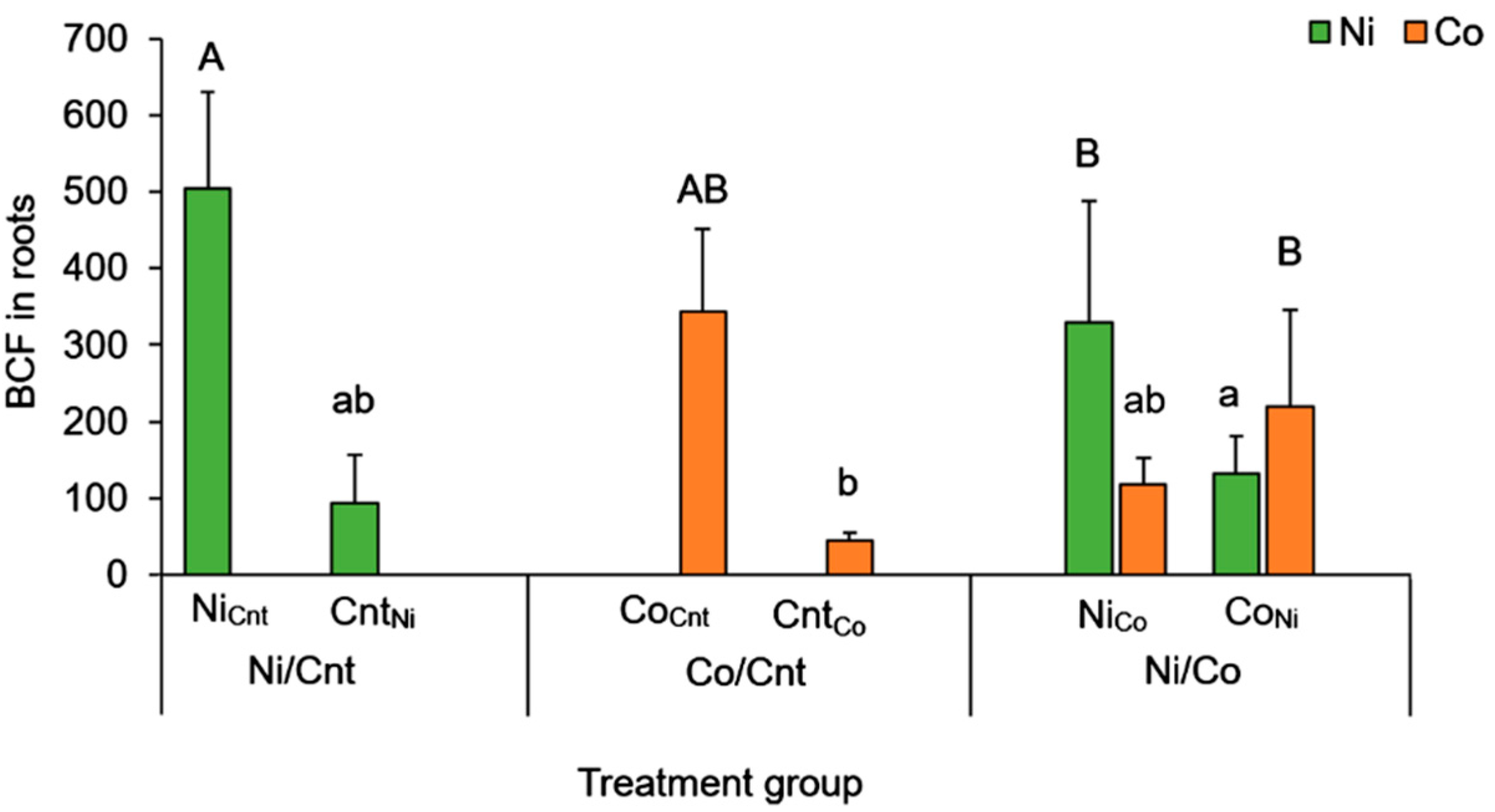
2.3. Comparison between Ni and Co Accumulation Using BCF
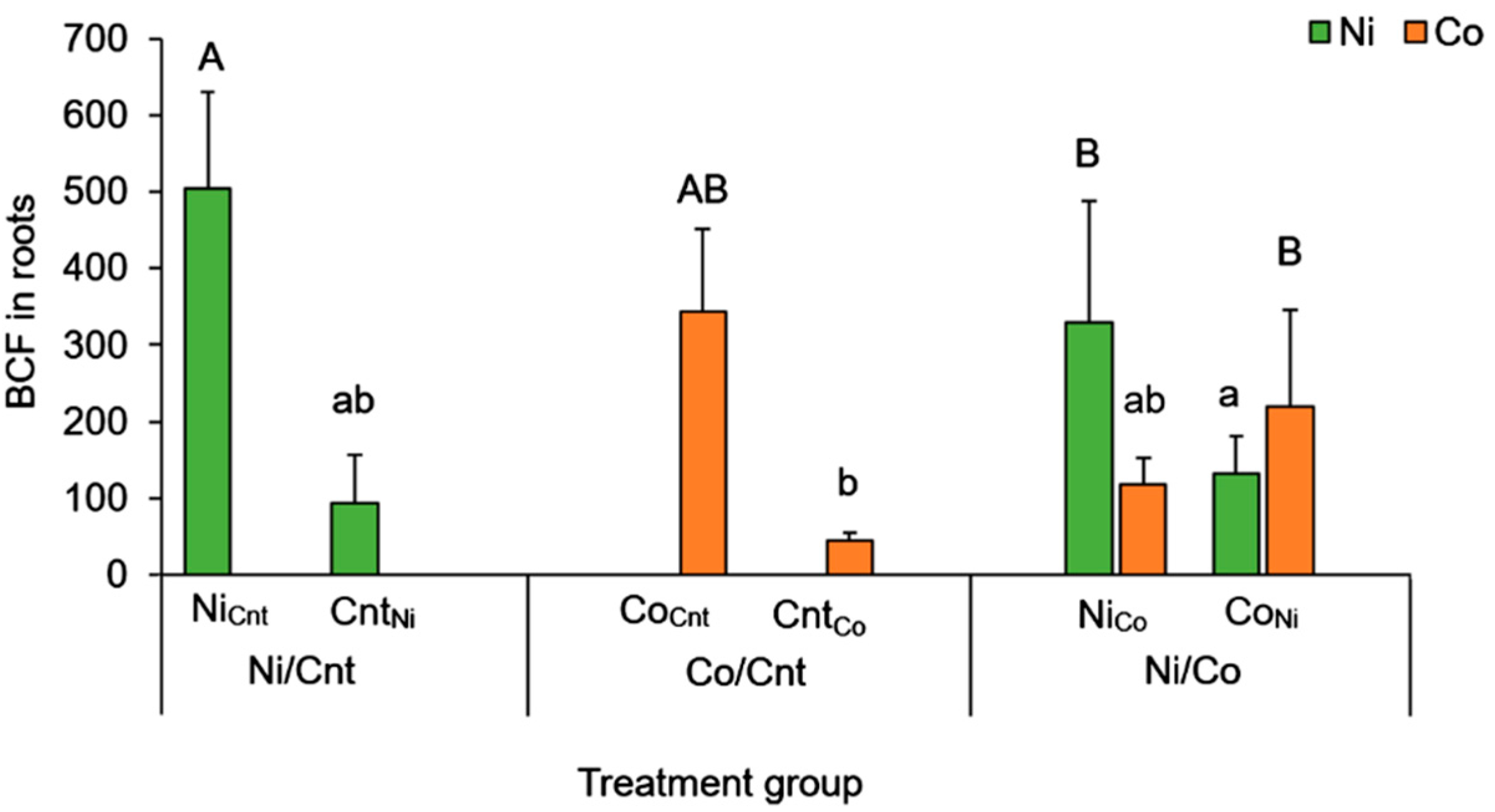
| Element | Treatment | BCF | ||
|---|---|---|---|---|
| Leaves | Barks | Wood | ||
| Ni | Ni/Cnt | 17.5 ± 8.36 a | 8.01 ± 4.05 a | 2.88 ± 1.05 a |
| Co/Cnt | - | - | - | |
| Ni/Co | 28.0 ± 16.4 a | 10.3 ± 5.76 a | 3.22 ± 0.94 a | |
| Co | Ni/Cnt | - | - | - |
| Co/Cnt | 236 ± 100 b | 17.2 ± 2.19 ab | 4.00 ± 0.91 a | |
| Ni/Co | 216 ± 79.5 b | 27.6 ± 10.2 b | 6.62 ± 1.88 b | |
3. Discussion
3.1. Effects of Ni and Co on Root Growth
3.2. Difference between Ni and Co Accumulation and Translocation
3.3. Relationship between S and Co
3.4. Interaction between Ni and Co
4. Experimental Section
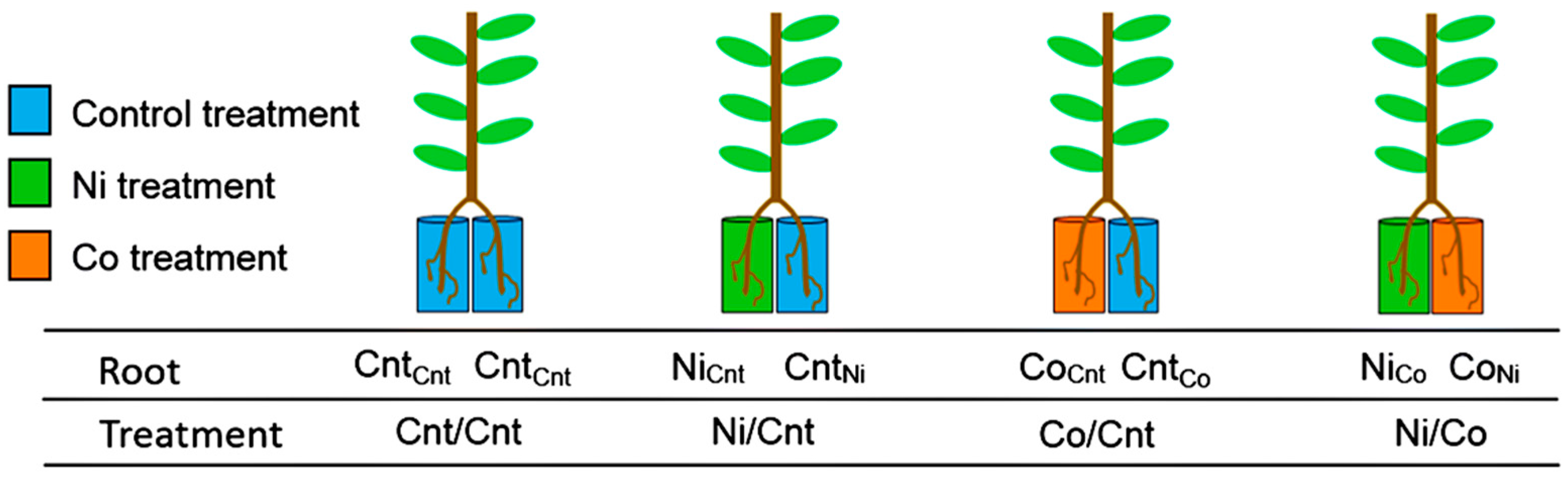
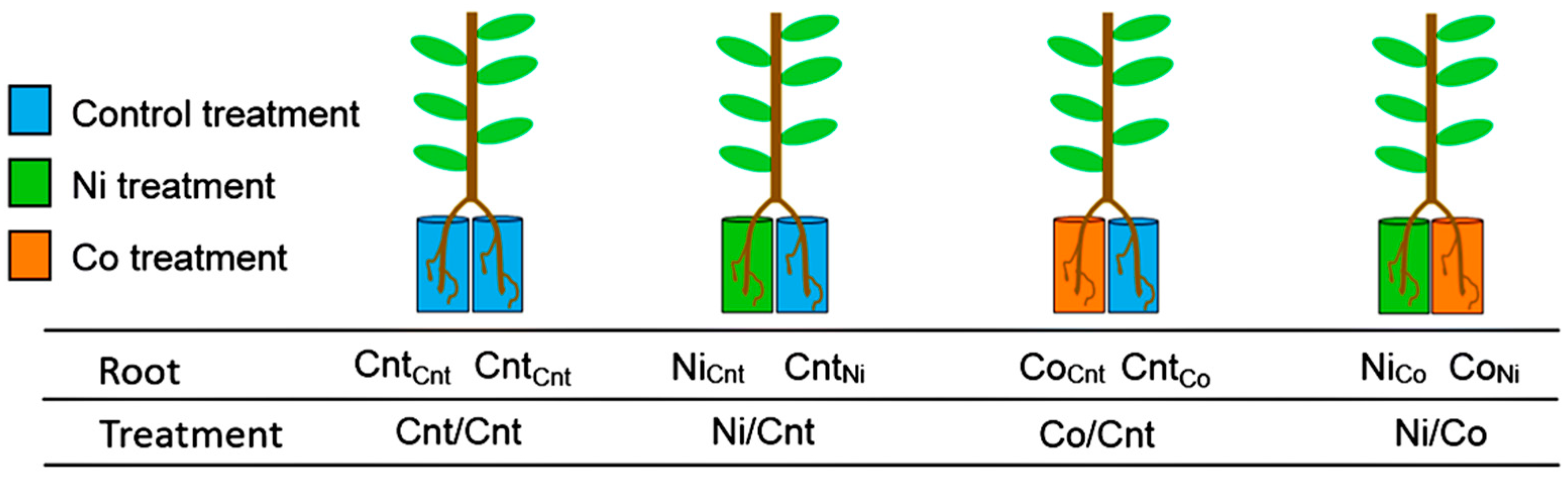
4.1. Experimental Design and Plant Growth
4.2. Plant Tissue Analyses
4.3. Calculation of BCF
4.4. Measurement of Photosynthesis
4.5. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kubota, M.; McGonigle, T.P.; Hyakumachi, M. Clethra barbinervis, a member of the order Ericales, forms arbuscular mycorrhizae. Can. J. Bot. 2001, 79, 300–306. [Google Scholar]
- Okamoto, K.; Yamamoto, Y.; Fuwa, K. Accumulation of manganese, zinc, cobalt, nickel and cadmium by Clethra barbinervis. Agric. Biol. Chem. 1978, 42, 663–664. [Google Scholar] [CrossRef]
- Azuma, A.K.; Tomioka, R.; Takenaka, C. Evaluation of microelement contents in Clethra barbinervis as food for human and animals in contrasting geological areas. Environ. Geochem. Health 2015. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of nutrients: Micronutrients. In Marschers’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier: London, UK, 2012; pp. 191–248. [Google Scholar]
- Broadley, M.; Brown, P.; Cakmak, I.; Ma, J.F.; Rengel, Z.; Zhao, F. Beneficial Elements. In Marschers’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier: London, UK, 2012; pp. 249–269. [Google Scholar]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press: Boca Raton, FL, USA, 2011; pp. 227–251. [Google Scholar]
- Palit, S.; Sharma, A.; Talukder, G. Effects of cobalt on plants. Bot. Rev. 1994, 60, 149–181. [Google Scholar] [CrossRef]
- Gabbrielli, R.; Mattioni, C.; Vergnano, O. Accumulation mechanisms and heavy metal tolerance of a nickel hyperaccumulator. J. Plant Nutr. 1991, 14, 1067–1080. [Google Scholar] [CrossRef]
- Keeling, S.M.; Stewart, R.B.; Anderson, C.W.N.; Robinson, B.H. Nickel and Cobalt Phytoextraction by the Hyperaccumulator Berkheya coddii : Implications for Polymetallic Phytomining and Phytoremediation. Int. J. Phytoremediation 2003, 5, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Welch, R.M.; Hart, J.J.; Norvell, W.A.; Sullivan, L.A.; Kochian, L.V. Effects of nutrient solution zinc activity on net uptake, translocation, and root export of cadmium and zinc by separated sections of intact durum wheat (Triticum turgidum L. var durum) seedling roots. Plant Soil 1999, 208, 243–250. [Google Scholar] [CrossRef]
- Schmidt, W. Influence of chromium(lll) on root-associated Fe(lll) reductase in Plantago lanceolata L. J. Exp. Bot. 1996, 47, 805–810. [Google Scholar] [CrossRef]
- Larrainzar, E.; Gil-Quintana, E.; Arrese-Igor, C.; González, E.M.; Marino, D. Split-root systems applied to the study of the legume-rhizobial symbiosis: What have we learned? J. Integr. Plant Biol. 2014, 56, 1118–1124. [Google Scholar] [CrossRef] [PubMed]
- Rouached, H.; Wirtz, M.; Alary, R.; Hell, R.; Arpat, A.B.; Davidian, J.-C.; Fourcroy, P.; Berthomieu, P. Differential regulation of the expression of two high-affinity sulfate transporters, SULTR1.1 and SULTR1.2, in Arabidopsis. Plant Physiol. 2008, 147, 897–911. [Google Scholar] [CrossRef] [PubMed]
- Na, G.; Salt, D.E. The role of sulfur assimilation and sulfur-containing compounds in trace element homeostasis in plants. Environ. Exp. Bot. 2011, 72, 18–25. [Google Scholar] [CrossRef]
- Küpper, H.; Lombi, E.; Zhao, F.; Wieshammer, G.; Mcgrath, S.P. Cellular compartmentation of nickel in the hyperaccumulators Alyssum lesbiacum, Alyssum bertolonii and Thlaspi goesingense. J. Exp. Bot. 2001, 52, 2291–2300. [Google Scholar] [CrossRef] [PubMed]
- Broadhurst, C.L.; Chaney, R.L.; Angle, J.S.; Erbe, E.F.; Maugel, T.K. Nickel localization and response to increasing Ni soil levels in leaves of the Ni hyperaccumulator Alyssum murale. Plant Soil 2004, 265, 225–242. [Google Scholar] [CrossRef]
- Tappero, R.; Peltier, E.; Gräfe, M.; Heidel, K.; Ginder-Vogel, M.; Livi, K.J.T.; Rivers, M.L.; Marcus, M.A.; Chaney, R.L.; Sparks, D.L. Hyperaccumulator Alyssum murale relies on a different metal storage mechanism for cobalt than for nickel. New Phytol. 2007, 175, 641–654. [Google Scholar] [CrossRef] [PubMed]
- He, S.; He, Z.; Yang, X.; Baligar, V.C. Mechanisms of nickel uptake and hyperaccumulation by plants and implications for soil remediation. Adv. Agron. 2012, 117, 117–189. [Google Scholar]
- Page, V.; Weisskopf, L.; Feller, U. Heavy metals in white lupin: Uptake, root-to-shoot transfer and redistribution within the plant. New Phytol. 2006, 171, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Page, V.; Feller, U. Selective transport of zinc, manganese, nickel, cobalt and cadmium in the root system and transfer to the leaves in young wheat plants. Ann. Bot. 2005, 96, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Riesen, O.; Feller, U. Redistribution of nickel, cobalt, manganese, zinc, and cadmium via the phloem in young and maturing wheat. J. Plant Nutr. 2005, 28, 421–430. [Google Scholar] [CrossRef]
- Wei, S.; Anders, I.; Feller, U. Selective uptake, distribution, and redistribution of 109Cd, 57Co, 65Zn, 63Ni, and 134Cs via xylem and phloem in the heavy metal hyperaccumulator Solanum nigrum L. Environ. Sci. Pollut. Res. 2014, 21, 7624–7630. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, M.; Fariduddin, Q.; Hayat, S.; Ahmad, A. Nickel: An overview of uptake, essentiality and toxicity in plants. Bull. Environ. Contam. Toxicol. 2011, 86, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Karuppanapandian, T.; Kim, W. Cobalt-induced oxidative stress causes growth inhibition associated with enhanced lipid peroxidation and activates antioxidant responses in Indian mustard (Brassica juncea L.) leaves. Acta Physiol. Plant. 2013, 35, 2429–2443. [Google Scholar] [CrossRef]
- Brooks, R.R.; Lee, J.; Reeves, R.D.; Jaffre, T. Detection of nickeliferous rocks by analysis of herbarium specimens of indicator plants. J. Geochem. Explor. 1977, 7, 49–57. [Google Scholar] [CrossRef]
- Baker, A.J.M. Accumulators and excluders—Strategies in the response of plants to heavy metals. J. Plant Nutr. 1981, 3, 643–654. [Google Scholar] [CrossRef]
- Küpper, H.; Kroneck, P.M.H. Heavy metal uptake by plants and cyanobacteria. Met. Ions Biol. Syst. 2005, 44, 97–144. [Google Scholar] [PubMed]
- Küpper, H.; Lombi, E.; Zhao, F.J.; McGrath, S.P. Cellular compartmentation of cadmium and zinc in relation to other elements in the hyperaccumulator Arabidopsis halleri. Planta 2000, 212, 75–84. [Google Scholar] [PubMed]
- Haslett, B.S.; Reid, R.J.; Rengel, Z. Zinc mobility in wheat: Uptake and distribution of zinc applied to leaves or roots. Ann. Bot. 2001, 87, 379–386. [Google Scholar] [CrossRef]
- McNear, D.H.; Chaney, R.L.; Sparks, D.L. The hyperaccumulator Alyssum murale uses complexation with nitrogen and oxygen donor ligands for Ni transport and storage. Phytochemistry 2010, 71, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of Macronutrients. In Marschers’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier: London, UK, 2012; pp. 135–189. [Google Scholar]
- Freeman, J.L.; Persans, M.W.; Nieman, K.; Albrecht, C.; Peer, W.; Pickering, I.J.; Salt, D.E. Increased glutathione biosynthesis plays a role in nickel tolerance in Thlaspi nickel hyperaccumulators. Plant Cell 2004, 16, 2176–2191. [Google Scholar] [CrossRef] [PubMed]
- Oven, M.; Grill, E.; Golan-Goldhirsh, A.; Kutchan, T.M.; Zenk, M.H. Increase of free cysteine and citric acid in plant cells exposed to cobalt ions. Phytochemistry 2002, 60, 467–474. [Google Scholar] [CrossRef]
- Assunção, A.G.L.; Bleeker, P.; ten Bookum, W.M.; Vooijs, R.; Schat, H. Intraspecific variation of metal preference patterns for hyperaccumulation in Thlaspi caerulescens: Evidence from binary metal exposures. Plant Soil 2008, 303, 289–299. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamaguchi, T.; Tomioka, R.; Takenaka, C. Can Clethra barbinervis Distinguish Nickel and Cobalt in Uptake and Translocation? Int. J. Mol. Sci. 2015, 16, 21378-21391. https://doi.org/10.3390/ijms160921378
Yamaguchi T, Tomioka R, Takenaka C. Can Clethra barbinervis Distinguish Nickel and Cobalt in Uptake and Translocation? International Journal of Molecular Sciences. 2015; 16(9):21378-21391. https://doi.org/10.3390/ijms160921378
Chicago/Turabian StyleYamaguchi, Tsuyoshi, Rie Tomioka, and Chisato Takenaka. 2015. "Can Clethra barbinervis Distinguish Nickel and Cobalt in Uptake and Translocation?" International Journal of Molecular Sciences 16, no. 9: 21378-21391. https://doi.org/10.3390/ijms160921378
APA StyleYamaguchi, T., Tomioka, R., & Takenaka, C. (2015). Can Clethra barbinervis Distinguish Nickel and Cobalt in Uptake and Translocation? International Journal of Molecular Sciences, 16(9), 21378-21391. https://doi.org/10.3390/ijms160921378