
A Review of Cell Adhesion Studies for Biomedical and Biological Applications

Abstract
:
1. Introduction
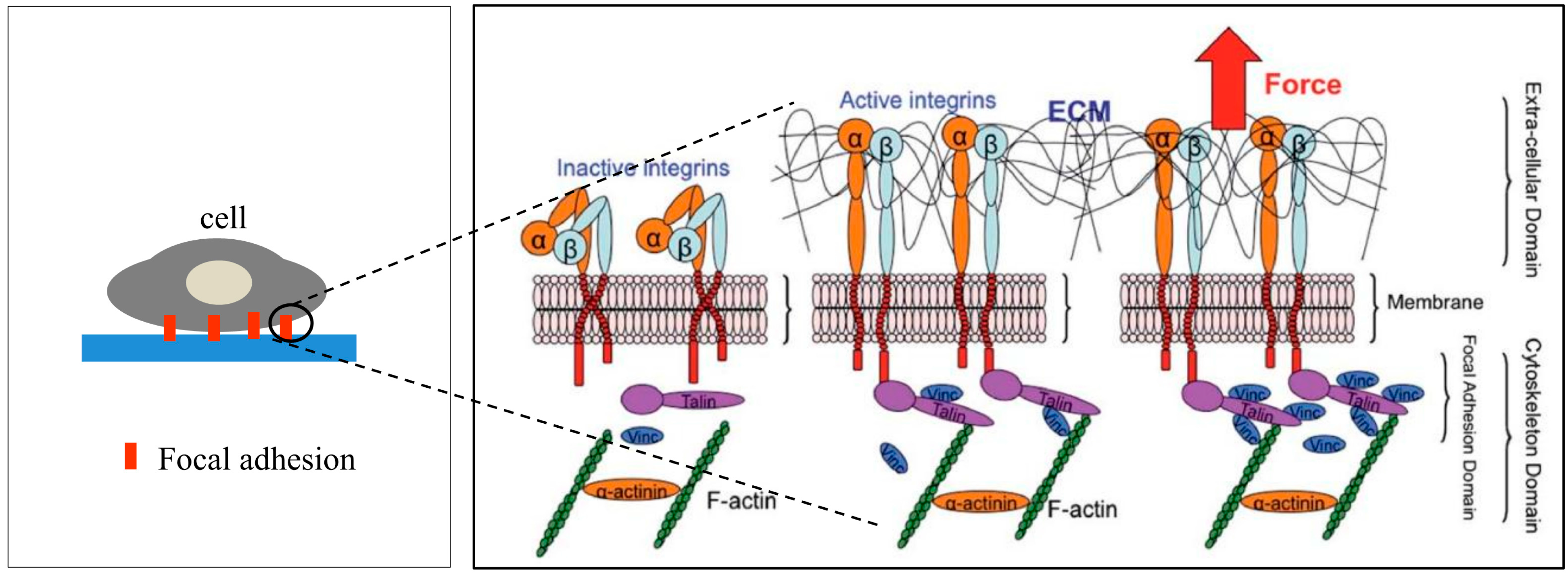
1.1. Focal Adhesion
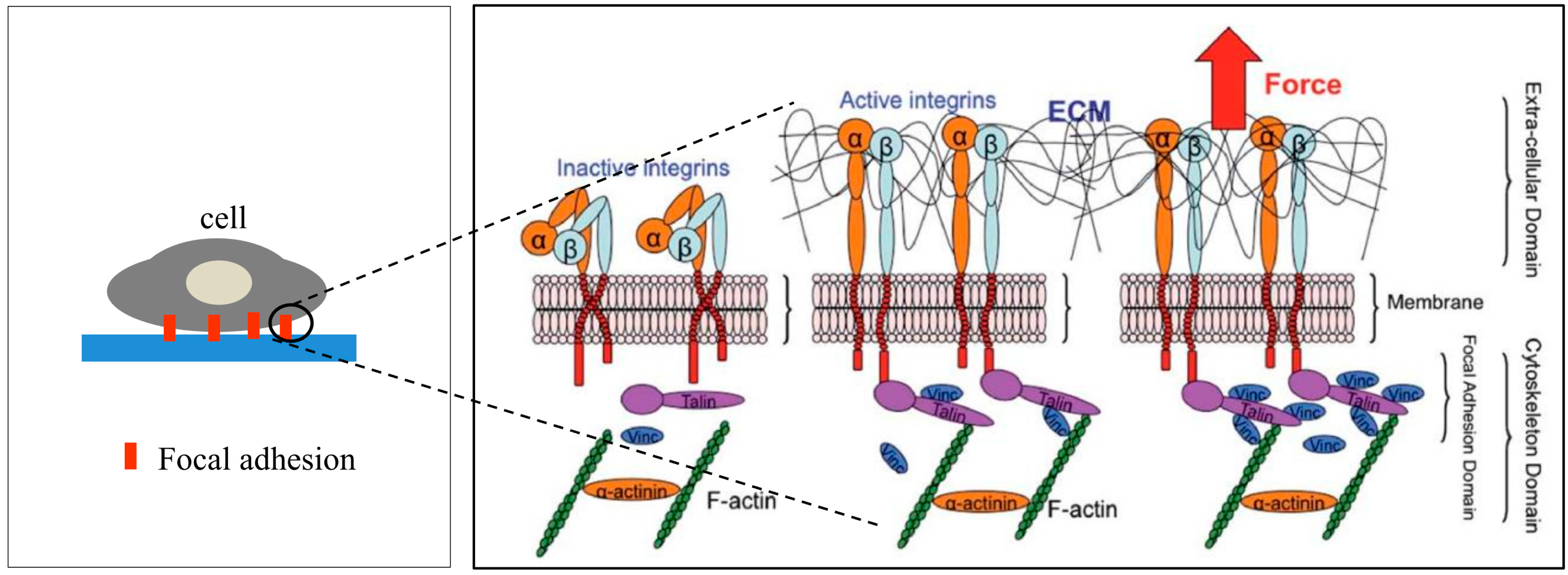
1.2. Phases of Cell Adhesion and Spreading
1.2.1. Passive in Vitro Cell Adhesion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Adhesion Phases | Phase I | Phase II | Phase III |
|---|---|---|---|
| Schematic diagram of cell adhesion |  | 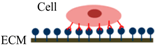 | 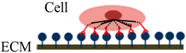 |
| Schematic diagram of the transformation of cell shape |  Initial attachment |  Flattening |  Fully spreading and structural organization |
| Cell adhesion intervension | Electrostatic interaction | Integrin bonding | Focal adhesion |
| Adhesion stages | Sedimentation | Cell attachment | Cell spreading and stable adhesion |
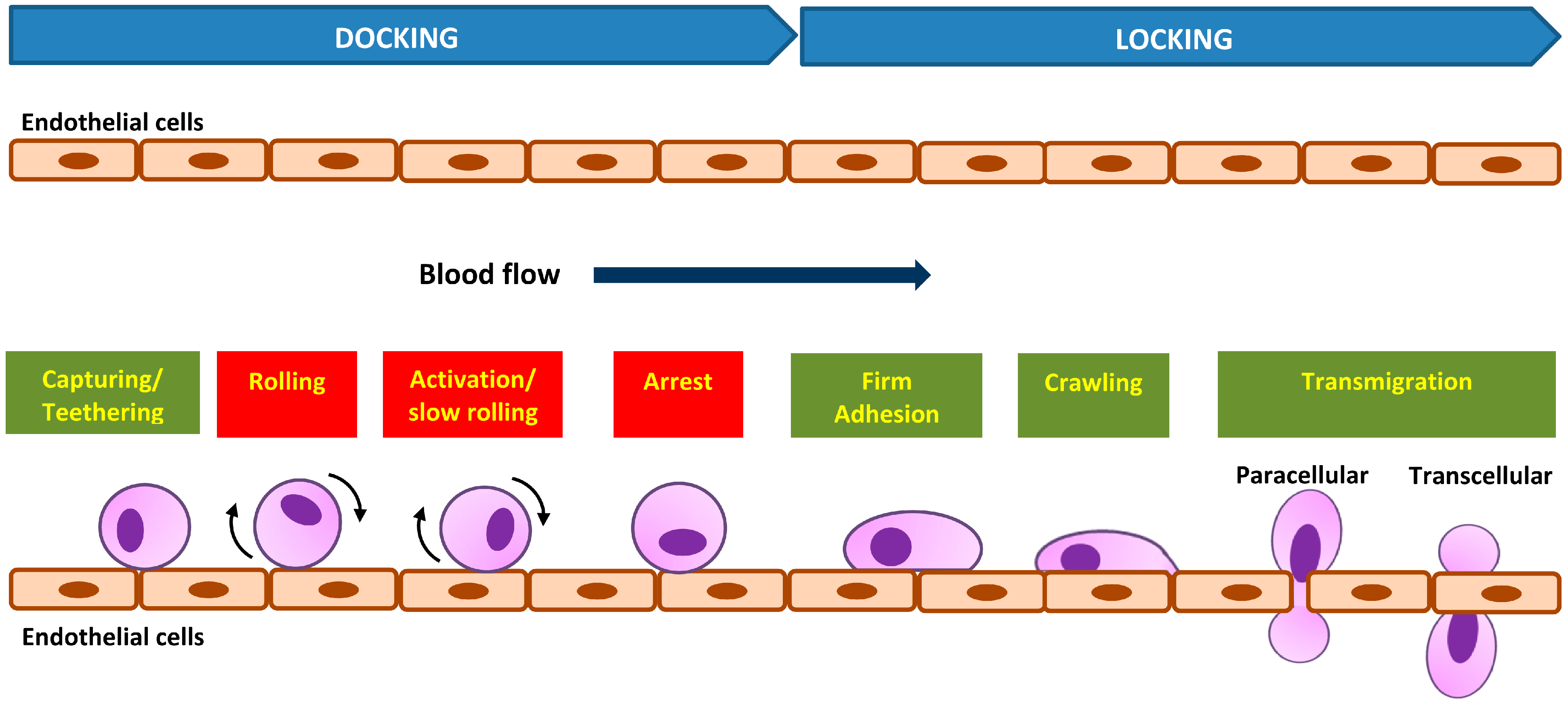
1.2.2. Dynamic in Vivo Cell Adhesion

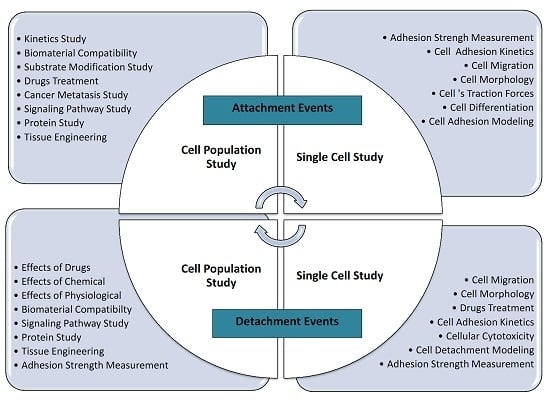
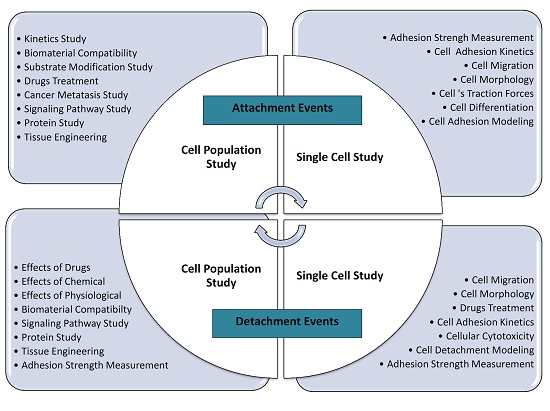
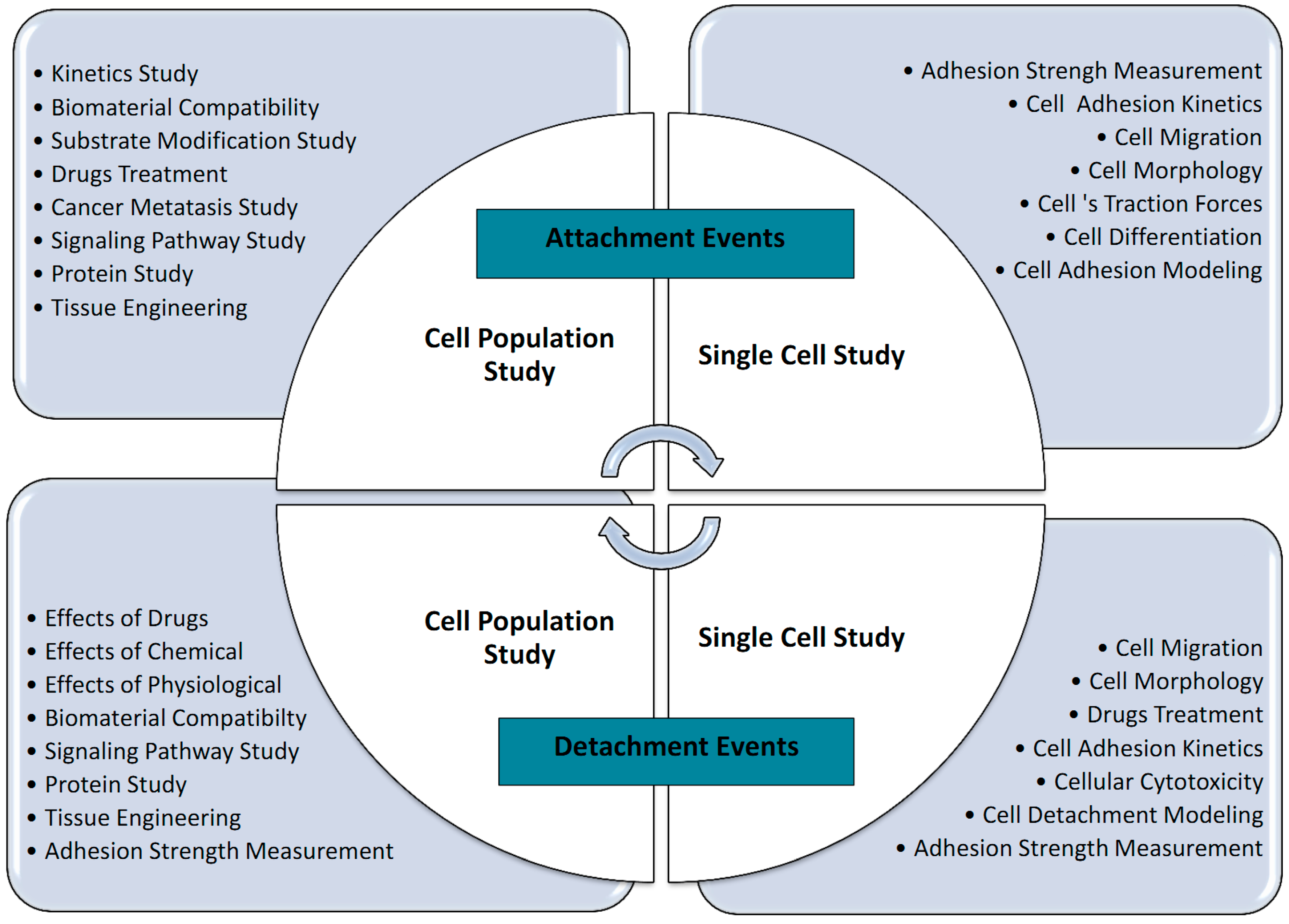
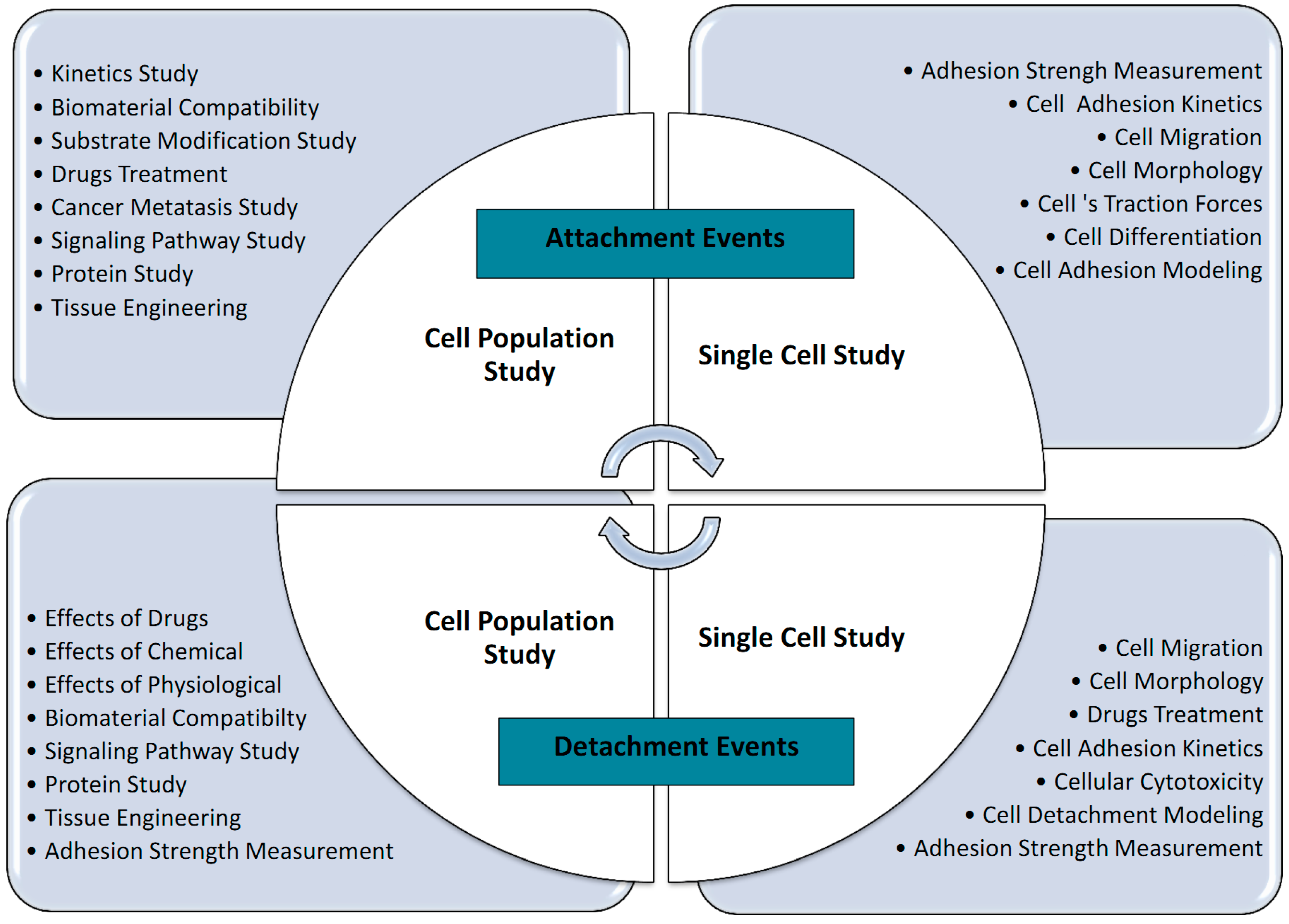
2. Types of Adhesion Studies
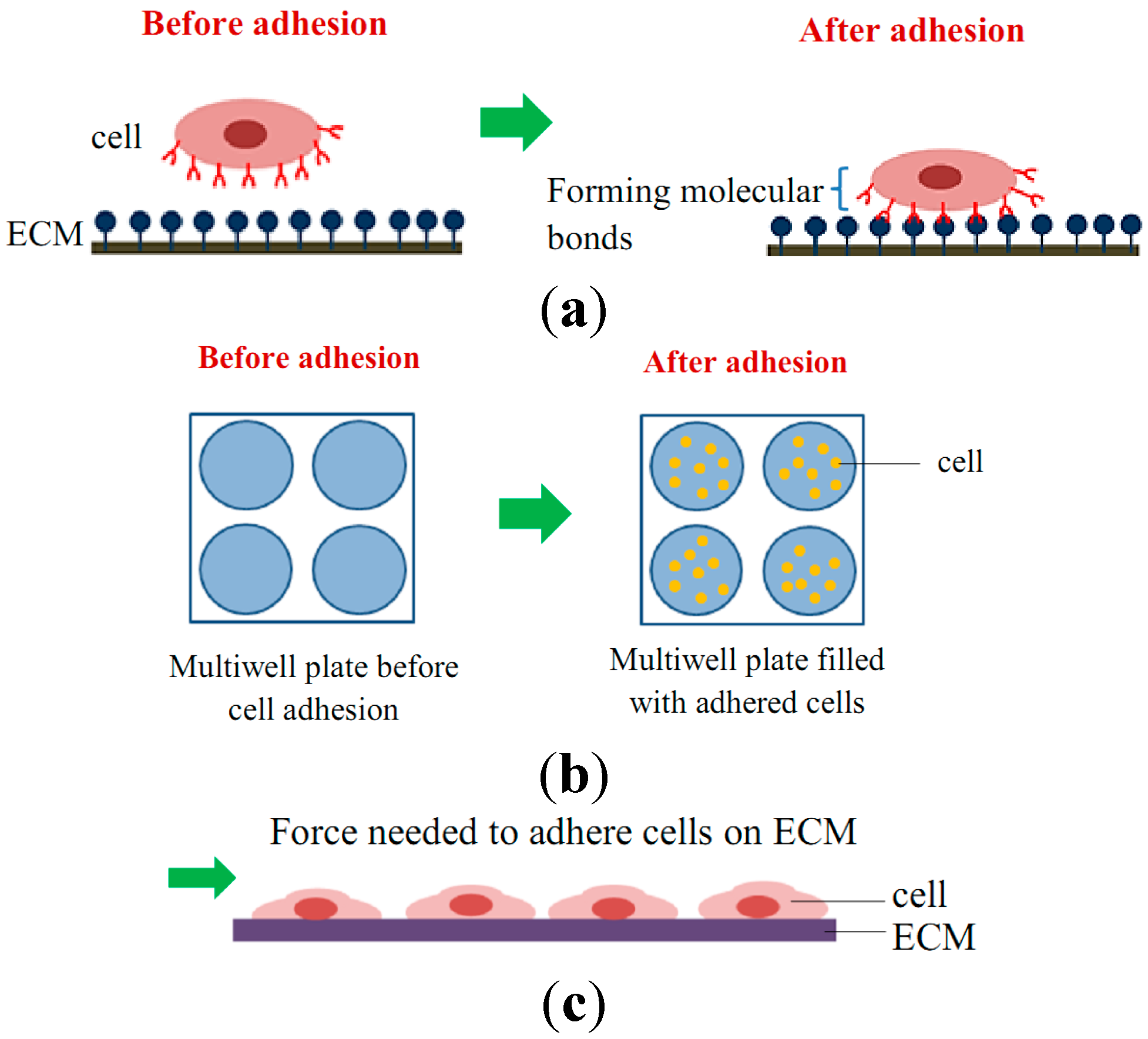
2.1. Cell Adhesion Attachment Events
2.1.1. Techniques to Study Cell Attachment Events
Attachment Events: Single Cell Approach
Attachment Events: Population Approach
2.2. Cell Adhesion Detachment Events
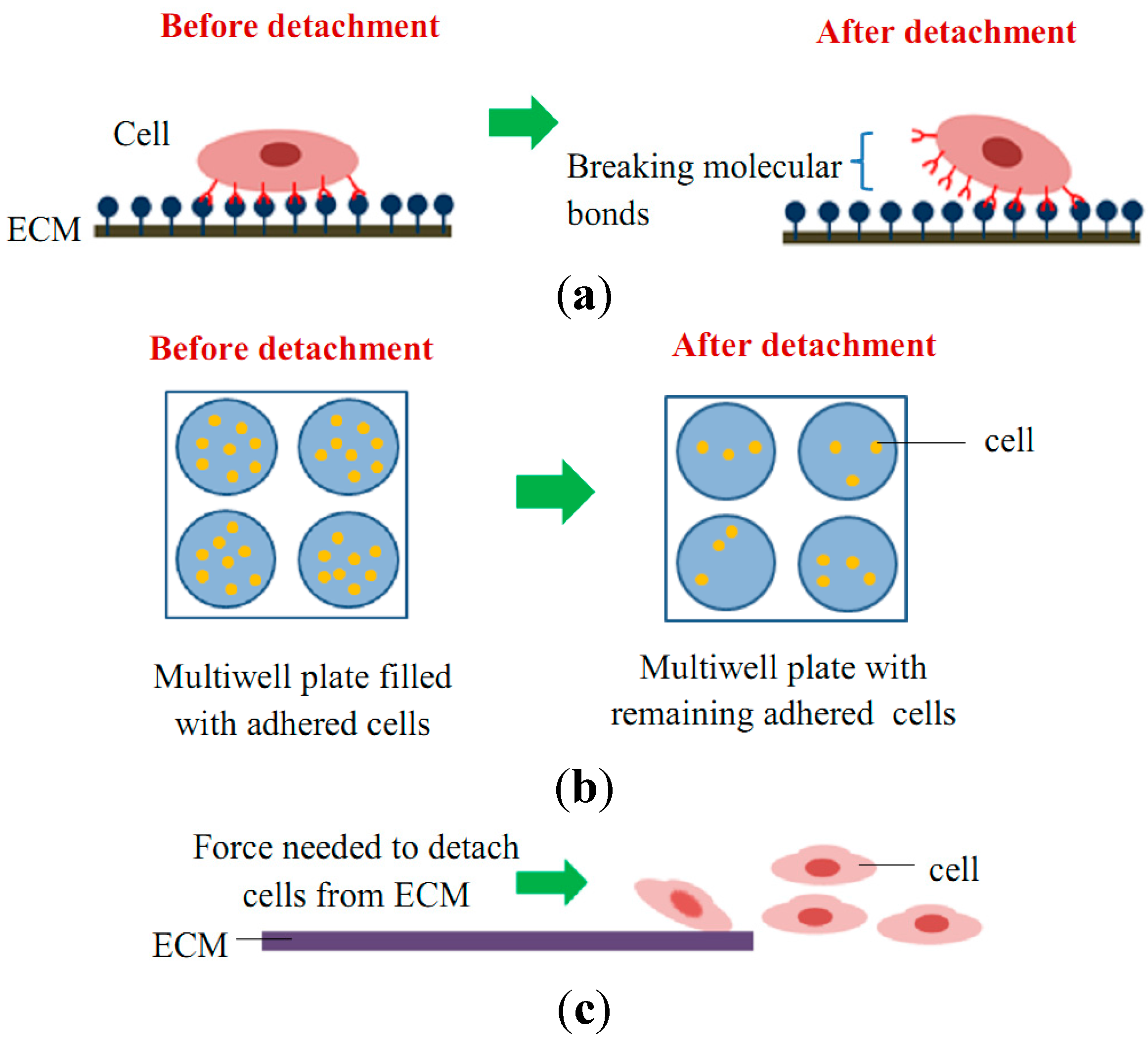
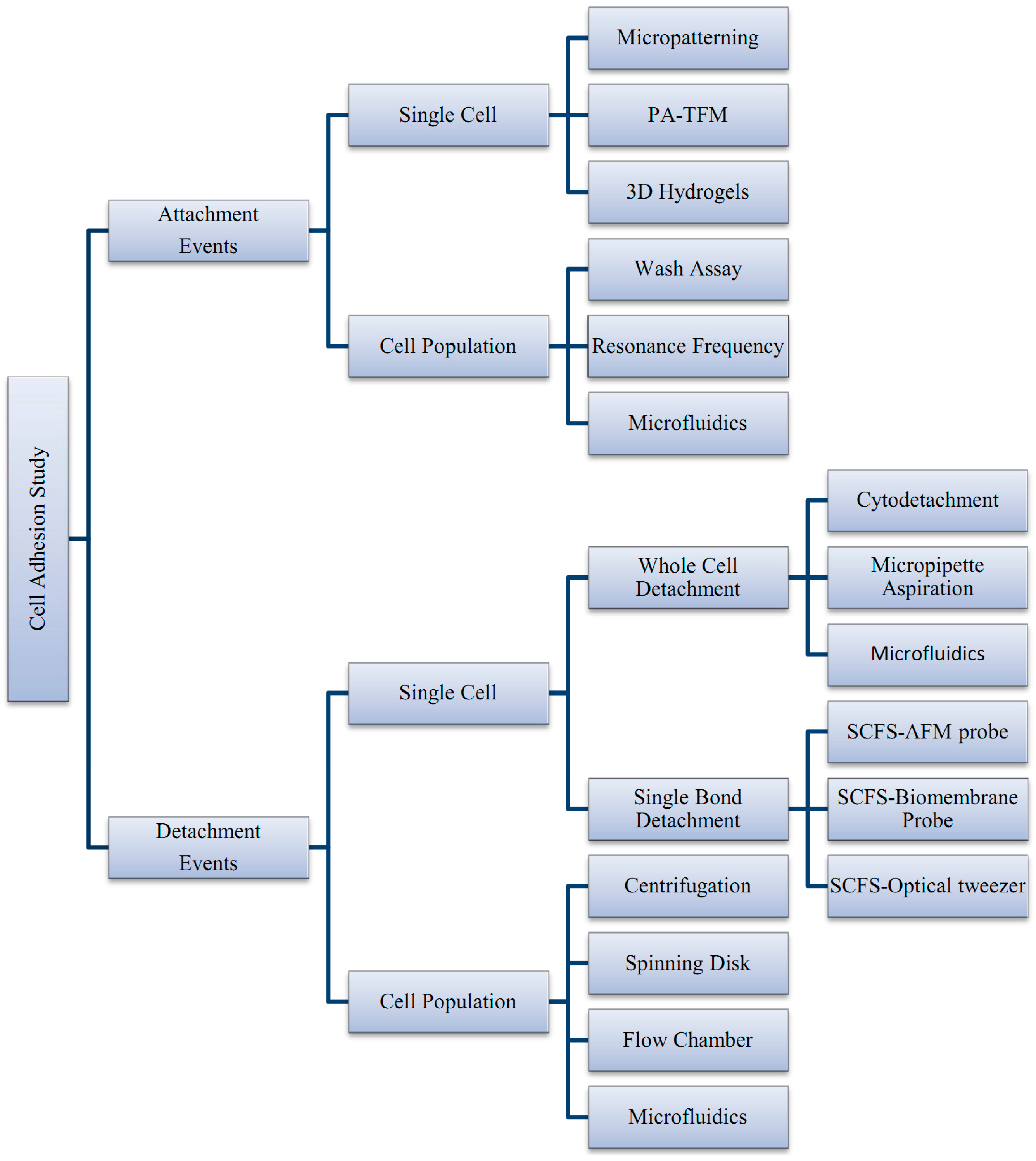
2.2.1. Techniques to Study Cell Detachment Events
Single Cell Approach
Detachment Events: Population Approach
3. Advantages and Limitations of the Techniques Used in Cell Adhesion Studies
| Method | Strength | Weaknesses | References |
|---|---|---|---|
| Polyacylamide-traction Force Microscopy (PA-TFM) | Real time observation; No special and expensive equipment needed for fabrication Inexpensive; Flexible to chemical and mechanical adjustment; Adaptable to a large variety of cells | Needs to record both unstressed and stressed state of the substrate; Suffers from uncertainties in tracking beads’ position | [47,48,49,50,51,52,53,54] |
| Micropatterning (Micropost array/micropillar) | Real-time observation; Force quantification easier and more reliable than PA-TFM; The micropillar stiffness is manipulated by its geometry; Gives good precision over surface chemical properties on micrometer scale | Substrate can alter cell’s behavior; Requires sophisticated equipment to fabricate; Needs skilled operator; Sensitivity of the microposts to the particular cell type needs to be optimized | [26,55,56,57,58,59,60] |
| Three Dimensional Traction Force Quantification (3D-TFM) | Real-time observation; Flexible to chemical and mechanical adjustment; Adaptable to a large variety of cells; Flexible to chemical and mechanical adjustment; Adaptable to a large variety of cells | Needs high-end confocal microscope; Needs high computational processing; Needs to record both unstressed and stressed state of the substrate; Suffers from uncertainties in tracking beads position | [61,62,63,64,65,66,67,68,69,70] |
| Wash Assay | Simple | Not a quantitative data, needs further analysis to obtain quantitative data; Poor reproducibility; Insensitive | [71,72,73,74] |
| Resonance Frequency | Real-time observation; Real-time measurement | Poor reproducibility | [36,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95] |
| Microfluidics | Straightforward construction and operation; Real-time observation and measurement; Convenience in size (compatible with cell sizes); Fast and simple to operate; Non-invasive to cell | Low detachment force; Restricted to short-term adhesion | Attachment events [96,97,98,99,100,101,102,103,107]; Detachment Events [19,24,107]; [197,198,199,200] |
| Cytodetachment | Real-time observation; Quick detachment of cell; Range of force produced is high and applicable to long-term adhesion | Alignment of probe and cell; Time-consuming; Needs highly skilled (experienced) operator; Operator variable; Cell damage (hard contact); Expensive equipment; Not real-time measurement | [104,105,106] |
| Micropipette Aspiration | Real-time observation and measurement; Common lab equipments | Alignment of probe and cell; High skilled (experienced) operator; Operator variable; Cell damage (hard contact) | [18,108,109] |
| SCFS-AFM probe | Real-time observation Precise data for short term adhesion studies | Alignment of probe and cell require micromanipulator; Time consuming; Need skilled operator; Operator variable; Cell damage (hard contact); Expensive equipments; Not real-time measurement | [105]; [116,117,118,119,120,121,122,123,124] |
| SCFS-Biomembrane Probe | Real-time observation; Precise data for short term adhesion studies | Low maximum force (pN); Restricted to short term adhesion; High skilled (experienced) operator; Operator variable; Probe variable (fluctuation of probe due to thermal excitation) | [125,126,127,128] |
| SCFS-Optical Tweezer | Real-time observation; Precise data for short term adhesion studies; Compatible with microfluidic device | Low maximum force (pN); Restricted to short term adhesion; High skilled (experienced) operator; Operator variable; Cell damage | [129,130,131,132,133,134,135,136,137,138] |
| Centrifugation | Many analysis can be examined in parallel; Common lab equipments | Low maximum force (uncomplete detachment); Only a single force can be applied per experiment; Nota real-time analysis | [139,140,141,142,143,144,145,146,147] |
| Spinning Disk | A range of stresses able to be applied in single experiment; High stresses | Not a real-time analysis; Custom-made apparatuses | [148,149,150,151,152,153,154,155,156,157,158,159,160] |
| Flow chamber: Radial flow; Parallel flow | Radial flow: Ranges of stresses applicable in single experiment; Real-time cell detachment observation; Paralel flow: Simple fabrication; Straightforward operation; Real-time cell detachment observation | Radial flow: Low detachment force; Restricted to short term adhesion; Paralel flow: Low detachment force; Restricted to short term adhesion | [163,164,165,166,167,168,169,170,171,172,173,174,175]; [176,177,178,179,180,181,182,183,184,185,186,187,188,189,190,191,192,193,194,195] |
4. Summary
5. Conclusions and Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sagvolden, G.; Giaever, I.; Pettersen, E.O.; Feder, J. Cell adhesion force microscopy. Proc. Natl. Acad. Sci. USA 1999, 96, 471–476. [Google Scholar] [PubMed]
- Dembo, M.; Torney, D.; Saxman, K.; Hammer, D. The kinetics of membrane-to-surface adhesion and detachment. Proc. R. Soc. 1988, 234, 55–83. [Google Scholar]
- Shen, Y.; Nakajima, M.; Kojima, S.; Homma, M.; Kojima, M.; Fukuda, T. Single cell adhesion force measurement for cell viability identification using an AFM cantilever-based micro putter. Meas. Sci. Technol. 2011, 22, 115802. [Google Scholar]
- Huang, S.; Ingber, D.E. The structural and mechanical complexity of cell-growth control. Nat. Cell Biol. 1999, 1, E131. [Google Scholar] [PubMed]
- Lasky, L.A.; Singer, M.S.; Dowbenko, D.; Imai, Y.; Henzel, W.J.; Grimley, C.; Fennie, C.; Gillett, N.; Watson, S.R.; Rosent, S.D. An endothelial ligand for L-Selectin is a novel mucin-like molecule. Cell 1992, 69, 927–938. [Google Scholar] [PubMed]
- Szekanecz, Z.; Koch, A.E. Cell-cell interactions in synovitis: Endothelial cells and immune cell migration. Arthritis Res. 2000, 2, 368–373. [Google Scholar] [PubMed]
- Okegawa, T.; Pong, R.-C.; Li, Y.; Hsieh, J.-T. The role of cell adhesion molecule in cancer progression and its application in cancer therapy. Acta Biochim. Pol. 2004, 51, 445–457. [Google Scholar] [PubMed]
- Hirohashi, S.; Kanai, Y. Cell adhesion system and human cancer morphogenesis. Cancer Sci. 2003, 94, 575–581. [Google Scholar] [PubMed]
- Perinpanayagam, H.; Zaharias, R.; Stanford, C.; Keller, J.; Schneider, G.; Brand, R. Early cell adhesion events differ between osteoporotic and non-osteoporotic osteoblasts. J. Orthop. Res. 2001, 19, 993–1000. [Google Scholar] [PubMed]
- Cho, P.; Schneider, G.B.; Kellogg, B.; Zaharias, R.; Keller, J.C. Effect of glucocorticoid-induced osteoporotic-like conditions on osteoblast cell attachment to implant surface microtopographies. Implant Dent. 2006, 15, 377–385. [Google Scholar] [PubMed]
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Green, C.E. Molecular mechanics and dynamics of leukocyte recruitment during inflammation. Annu. Rev. Biol. 2005, 7, 151–185. [Google Scholar] [CrossRef] [PubMed]
- Spangenberg, C.; Lausch, E.U.; Trost, T.M.; Prawitt, D.; May, A.; Keppler, R.; Fees, S.A.; Reutzel, D.; Bell, C.; Schmitt, S.; et al. ERBB2-mediated transcriptional up-regulation of the α5β1 integrin fibronectin receptor promotes tumor cell survival under adverse conditions. Cancer Res. 2006, 66, 3715–3725. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.X.; Liu, Y.; Pasquale, E.B.; Ruoslahti, E. Activated Src oncogene phosphorylates R-ras and suppresses integrin activity. J. Biol. Chem. 2002, 277, 1824–1827. [Google Scholar] [CrossRef] [PubMed]
- Mierke, C.T. Cancer cells regulate biomechanical properties of human microvascular endothelial cells. J. Biol. Chem. 2011, 286, 40025–40037. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Anvari, B.; Torres, J.H.; LeBaron, R.G.; Athanasiou, K.A. Temporal effects of cell adhesion on mechanical characteristics of the single chondrocyte. J. Orthop. Res. 2003, 21, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-C.; Hsu, Y.C.; Su, F.C.; Lu, S.C.; Lee, T.M. Effects of passivation treatments on titanium alloy with nanometric scale roughness and induced changes in fibroblast initial adhesion evaluated by a cytodetacher. J. Biomed. Mater. Res. 2009, 88, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.P.; Mycielska, M.E.; Burcu, H.; Osman, K.; Collins, T.; Beckerman, R.; Perrett, R.; Johnson, H.; Aydar, E.; Djamgoz, M.B.A. Single cell adhesion measuring apparatus (SCAMA): Application to cancer cell lines of different metastatic potential and voltage-gated Na+ channel expression. Eur. Biophys. J. 2008, 37, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.W.; Choi, S.S.; Lee, S.H.; Kim, B.; Lee, S.N.; Park, M.C.; Kim, P.; Hwang, S.Y.; Suh, K.Y. Label-free, microfluidic separation and enrichment of human breast cancer cells by adhesion difference. Lab Chip 2007, 7, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Saif, M.A.T.; Sager, C.R.; Coyer, S. Functionalized biomicroelectromechanical systems sensors for force response study at local adhesion sites of single living cells on substrates. Ann. Biomed. Eng. 2003, 31, 950–961. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, A.F. Integrins and health. Sci. Am. 1997, 276, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.S. Geometric control of cell life and death. Science 1997, 276, 1425–1428. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.C.; Waugh, R.E. Energy of dissociation of lipid bilayer from the membrane skeleton of red blood cells. Biophys. J. 1997, 72, 2669–2678. [Google Scholar] [CrossRef]
- Honarmandi, P.; Lee, H.; Lang, M.J.; Kamm, R.D. A microfluidic system with optical laser tweezers to study mechanotransduction and focal adhesion recruitment. Lab Chip 2011, 11, 684–694. [Google Scholar] [CrossRef] [PubMed]
- Beningo, K.A.; Dembo, M.; Kaverina, I.; Small, J.V.; Wang, Y. Nascent focal adhesions are responsible for the generation of strong propulsive forces in migrating fibroblasts. J. Cell Biol. 2001, 153, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.L.; Tien, J.; Pirone, D.M.; Gray, D.S.; Bhadriraju, K.; Chen, C.S. Cells lying on a bed of microneedles: An approach to isolate mechanical force. Proc. Natl. Acad. Sci. USA 2003, 100, 1484–1489. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; He, S.; Ji, B. Mechanics in mechanosensitivity of cell adhesion and its roles in cell migration. Int. J. Comp. Mater. Sci. Eng. 2012, 01, 1250032. [Google Scholar] [CrossRef]
- Dumbauld, D.W.; Lee, T.T.; Singh, A.; Scrimgeour, J.; Gersbach, C.A.; Zamir, E.A.; Fu, J.; Chen, C.S.; Curtis, J.E.; Craig, S.W.; et al. How vinculin regulates force transmission. Proc. Natl. Acad. Sci. USA 2013, 110, 9788–9793. [Google Scholar] [CrossRef] [PubMed]
- Geiger, B.; Bershadsky, A.; Pankov, R.; Yamada, K.M. Transmembrane crosstalk between the extracellular matrix and the cytoskeleton. Nat. Rev. Mol. Cell Biol. 2001, 2, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. Integrins: A family of cell surface receptors. Cell 1987, 48, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Van der Flier, A.; Sonnenberg, A. Function and interactions of integrins. Cell Tissue Res. 2001, 305, 285–298. [Google Scholar] [CrossRef] [PubMed]
- DiMilla, P.A.; Barbee, K.; Douglas, A.; Lauffenburger, D.A. Mathematical model for the effects of adhesion and mechanics on cell migration speed. Biophys. J. 1991, 60, 15–37. [Google Scholar] [CrossRef] [PubMed]
- Burridge, K.; Wennerberg, K.; Hill, C.; Carolina, N. Rho and Rac take center stage. Cell 2004, 116, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Hall, A. Rho GTPases and the actin cytoskeleton. Science 1998, 279, 509–514. [Google Scholar] [CrossRef] [PubMed]
- LeBaron, R.G.; Athanasiou, K.A. Ex vivo synthesis of articular cartilage. Biomaterials 2000, 21, 2575–2587. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Ergezen, E.; Lec, R.; Barbee, K.A. Real-time analysis of cell-surface adhesive interactions using thickness shear mode resonator. Biomaterials 2006, 27, 5813–5820. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, A.; Cisneros, D.; Friedrichs, J.; Puech, P.-H.; Muller, D.J.; Franz, C.M. Revealing early steps of alpha2beta1 integrin-mediated adhesion to collagen type I by using single-cell force spectroscopy. Mol. Biol. Cell 2007, 18, 1634–1644. [Google Scholar] [CrossRef] [PubMed]
- Helenius, J.; Heisenberg, C.-P.; Gaub, H.E.; Muller, D.J. Single-cell force spectroscopy. J. Cell Sci. 2008, 121, 1785–1791. [Google Scholar] [CrossRef] [PubMed]
- McEver, R.P.; Zhu, C. Rolling cell adhesion. Ann. Rev. Cell Dev. Biol. 2010, 26, 363–396. [Google Scholar] [CrossRef] [PubMed]
- Green, C.E.; Pearson, D.N.; Camphausen, R.T.; Staunton, D.E.; Simon, S.I. Shear-dependent capping of L-selectin and P-selectin glycoprotein ligand 1 by E-selectin signals activation of high-avidity β2-integrin on neutrophils. J. Immunol. 2004, 172, 7780–7790. [Google Scholar] [CrossRef] [PubMed]
- Belloni, P.; Tressler, R. Microvascular endothelial cell heterogeneity: Interactions with leukocytes and tumor cells. Cancer Metastasis Rev. 1990, 8, 353–389. [Google Scholar] [CrossRef] [PubMed]
- Honn, K.; Tang, D. Adhesion molecules and tumor cell interaction with endothelium and subendothelial matrix. Cancer Metastasis Rev. 1992, 11, 353–375. [Google Scholar] [CrossRef] [PubMed]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Petri, B.; Phillipson, M.; Kubes, P. The physiology of leukocyte recruitment: An in vivo perspective. J. Immunol. 2008, 180, 6439–6446. [Google Scholar] [CrossRef] [PubMed]
- Imhof, B.A.; Aurrand-Lions, M. Adhesion mechanisms regulating the migration of monocytes. Nat. Rev. Immunol. 2004, 4, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Muller, W.A. Leukocyte–endothelial-cell interactions in leukocyte transmigration and the inflammatory response. Trends Immunol. 2015, 24, 326–333. [Google Scholar] [CrossRef]
- Dembo, M.; Wang, Y. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophys. J. 1999, 76, 2307–2316. [Google Scholar] [CrossRef]
- Kraning-Rush, C.M.; Califano, J.P.; Reinhart-King, C.A. Cellular traction stresses increase with increasing metastatic potential. PLoS ONE 2012, 7, e32572. [Google Scholar] [CrossRef] [PubMed]
- Huynh, J.; Bordeleau, F.; Kraning-Rush, C.M.; Reinhart-King, C.A. Substrate stiffness regulates PDGF-induced circular dorsal ruffle formation through MLCK. Cell. Mol. Bioeng. 2014, 6, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Reinhart-King, C.A.; Dembo, M.; Hammer, D.A. Endothelial cell traction forces on RGD-derivatized polyacrylamide substrata. Langmuir 2003, 19, 1573–1579. [Google Scholar] [CrossRef]
- Sabass, B.; Gardel, M.L.; Waterman, C.M.; Schwarz, U.S. High resolution traction force microscopy based on experimental and computational advances. Biophys. J. 2008, 94, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.R.; Besser, A.; Danuser, G.; Brugge, J.S. Substrate stiffness regulates cadherin-dependent collective migration through myosin-II contractility. J. Cell Biol. 2012, 199, 545–563. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.H.; Vincent, L.G.; Fuhrmann, A.; Choi, Y.S.; Hribar, K.C.; Taylor-weiner, H.; Chen, S.; Engler, A.J. Interplay of matrix stiffness and protein tethering in stem cell differentiation. Nat. Mater. 2014, 13, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Califano, J.P.; Reinhart-King, C.A. Substrate stiffness and cell area predict cellular traction stresses in single cells and cells in contact. Cell. Mol. Bioeng. 2011, 3, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.W.; García, A.J. Measurement systems for cell adhesive forces. J. Biomech. Eng. 2015, 137, 020908. [Google Scholar] [CrossRef] [PubMed]
- Théry, M. Micropatterning as a tool to decipher cell morphogenesis and functions. J. Cell Sci. 2010, 123, 4201–4213. [Google Scholar] [CrossRef] [PubMed]
- D’Arcangelo, E.; McGuigan, A.P. Micropatterning strategies to engineer controlled cell and tissue architecture in vitro. Biotechniques 2014, 58, 13–23. [Google Scholar]
- Mandal, K.; Balland, M.; Bureau, L. Thermoresponsive micropatterned substrates for single cell studies. PLoS ONE 2012, 7, e37548. [Google Scholar] [CrossRef] [PubMed]
- Polio, S.R.; Rothenberg, K.E.; Stamenović, D.; Smith, M.L. A micropatterning and image processing approach to simplify measurement of cellular traction forces. Acta Biomater. 2012, 8, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Ting, L.H.; Jahn, J.R.; Jung, J.I.; Shuman, B.R.; Feghhi, S.; Han, S.J.; Rodriguez, M.L.; Sniadecki, N.J. Flow mechanotransduction regulates traction forces, intercellular forces, and adherens junctions. Am. J. Physiol. 2012, 302, H2220–H2229. [Google Scholar]
- Santos, E.; Hernández, R.M.; Pedraz, J.L.; Orive, G. Novel advances in the design of three-dimensional bio-scaffolds to control cell Fate: Translation from 2D to 3D. Trends Biotechnol. 2012, 30, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.M.; Chen, C.S. Deconstructing the third dimension: How 3D culture microenvironments alter cellular cues. J. Cell Sci. 2012, 125, 3015–3024. [Google Scholar] [CrossRef] [PubMed]
- Legant, W.R.; Miller, J.S.; Blakely, B.L.; Cohen, D.M.; Genin, G.M.; Chen, C.S. Measurement of mechanical tractions exerted by cells in three-dimensional matrices. Nat. Methods 2010, 7, 969–971. [Google Scholar] [CrossRef] [PubMed]
- Khetan, S.; Guvendiren, M.; Legant, W.R.; Cohen, D.M.; Chen, C.S.; Burdick, J.A. Degradation-mediated cellular traction directs stem cell fate in covalently crosslinked three-dimensional hydrogels. Nat. Mater. 2013, 12, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Meseke, M.; Förster, E. A 3D-matrigel/microbead assay for the visualization of mechanical tractive forces at the neurite-substrate interface of cultured neurons. J. Biomed. Mater. Res. A 2013, 101A, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Fraley, S.I.; Feng, Y.; Krishnamurthy, R.; Kim, D.-H.; Celedon, A.; Longmore, G.D.; Wirtz, D. A distinctive role for focal adhesion proteins in three-dimensional cell motility. Nat. Cell Biol. 2010, 12, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Kraning-Rush, C.M.; Carey, S.P.; Califano, J.P.; Smith, B.N.; Reinhart-King, C.A. The role of the cytoskeleton in cellular force generation in 2D and 3D environments. Phys. Biol. 2011, 8, 015009. [Google Scholar] [CrossRef] [PubMed]
- Zaman, M.H.; Trapani, L.M.; Sieminski, A.L.; MacKellar, D.; Gong, H.; Kamm, R.D.; Wells, A.; Lauffenburger, D.A.; Matsudaira, P. Migration of tumor cells in 3D matrices is governed by matrix stiffness along with cell-matrix adhesion and proteolysis. Proc. Natl. Acad. Sci. USA 2006, 103, 10889–10894. [Google Scholar] [CrossRef] [PubMed]
- Bonakdar, N.; Butler, J.P.; Fabry, B.; Koch, T.M.; Mu, S. 3D traction forces in cancer cell invasion. PLoS ONE 2012, 7, e33476. [Google Scholar]
- Kutys, M.L.; Yamada, K.M. An extracellular matrix-specific GEF-GAP interaction regulates Rho GTPase crosstalk for 3D collagen migration. Nat. Cell Biol. 2015, 16, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Arnaout, M.A.; Gupta, V. A simple, no-wash cell adhesion-based high-throughput assay for the discovery of small-molecule regulators of the integrin CD11b/CD18. J. Biomol. Screen. 2011, 12, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.J.; Gallant, N.D. Stick and grip: Measurement systems and quantitative analyses of integrin-mediated cell adhesion strength. Cell Biochem. Biophys. 2003, 39, 61–73. [Google Scholar] [CrossRef]
- Chen, Y.; Lu, B.; Yang, Q.; Fearns, C.; Yates, J.R.; Lee, J.D. Combined integrin phosphoproteomic analyses and siRNA-based functional screening identified key regulators for cancer cell adhesion and migration. Cancer Res. 2010, 69, 3713–3720. [Google Scholar] [CrossRef] [PubMed]
- Mianabadi, M.; Yazdanparast, R. Inhibition of substrate-tumor cell adhesion under the effect of gnidilatimonoein purified from daphne mucronata. Am. J. Chin. Med. 2004, 32, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Christ, K.V.; Turner, K.T. Methods to measure the strength of cell adhesion to substrates. J. Adhes. Sci. Technol. 2010, 24, 37–41. [Google Scholar] [CrossRef]
- Ferreira, G.N.M.; da-Silva, A.-C.; Tomé, B. Acoustic wave biosensors: Physical models and biological applications of quartz crystal microbalance. Trends Biotechnol. 2009, 27, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Wegener, J.; Janshoff, A. Cell adhesion monitoring using a quartz crystal microbalance: Comparative analysis of different mammalian cell lines. Eur. Biophys. J. 1998, 28, 26–37. [Google Scholar] [CrossRef]
- Heitmann, V.; Wegener, J. Monitoring cell adhesion by piezoresonators: Impact of increasing oscillation amplitudes. Anal. Chem. 2007, 79, 3392–3400. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Chen, J.Y.; Garcia, M.P.; Penn, L.S. Quartz crystal microbalance in cell biology studies. J. Biochips Tissue Chipissue Chip. 2013, S5, 1–9. [Google Scholar]
- Li, F.; Wang, J.H.-C.; Wang, Q.-M. Monitoring cell adhesion by using thickness shear mode acoustic wave sensors. Biosens. Bioelectron. 2007, 23, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Fohlerová, Z.; Skládal, P.; Turánek, J. Adhesion of eukaryotic cell lines on the gold surface modified with extracellular matrix proteins monitored by the piezoelectric sensor. Biosens. Bioelectron. 2007, 22, 1896–1901. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, V.; Reiβ, B.; Wegener, J. The quartz crystal microbalance in cell biology: Basics and applications. Springer Ser. Chem. Sens. Biosens. 2007, 5, 303–338. [Google Scholar]
- Wegener, J.; Seebach, J.; Janshoff, A.; Galla, H.J. Analysis of the composite response of shear wave resonators to the attachment of mammalian cells. Biophys. J. 2000, 78, 2821–2833. [Google Scholar] [CrossRef]
- Modin, C.; Stranne, A.-L.; Foss, M.; Duch, M.; Justesen, J.; Chevallier, J.; Andersen, L.K.; Hemmersam, A.G.; Pedersen, F.S.; Besenbacher, F. QCM-D studies of attachment and differential spreading of pre-osteoblastic cells on Ta and Cr surfaces. Biomaterials 2006, 27, 1346–1354. [Google Scholar] [CrossRef] [PubMed]
- Lord, M.S.; Modin, C.; Foss, M.; Duch, M.; Simmons, A.; Pedersen, F.S.; Milthorpe, B.K.; Besenbacher, F. Monitoring cell adhesion on tantalum and oxidised polystyrene using a quartz crystal Microbalance with dissipation. Biomaterials 2006, 27, 4529–4537. [Google Scholar] [CrossRef] [PubMed]
- Le Guillou-Buffello, D.; Hélary, G.; Gindre, M.; Pavon-Djavid, G.; Laugier, P.; Migonney, V. Monitoring cell adhesion processes on bioactive polymers with the quartz crystal resonator technique. Biomaterials 2005, 26, 4197–4205. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Qiu, H.; Trzeciakowski, J.P.; Sun, Z.; Li, Z.; Hong, Z.; Hill, M.A; Hunter, W.C.; Vatner, D.E.; Vatner, S.F.; et al. Temporal analysis of vascular smooth muscle cell elasticity and adhesion reveals oscillation waveforms that differ with aging. Aging Cell 2012, 11, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Hayward, L.; Thompson, M. Acoustic waves and the study of biochemical macromolecules and cells at the sensor—Liquid interface critical review. Analyst 1999, 124, 1405–1420. [Google Scholar]
- Da-Silva, A.-C.; Soares, S.S.; Ferreira, G.N.M. Acoustic detection of cell Adhesion to a coated quartz crystal microbalance—Implications for studying the biocompatibility of polymers. Biotechnol. J. 2013, 8, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Marx, K.A.; Zhou, T.; Montrone, A.; McIntosh, D.; Braunhut, S.J. Quartz crystal microbalance biosensor study of endothelial cells and their extracellular matrix following cell removal: Evidence for transient cellular stress and viscoelastic changes during detachment and the elastic behavior of the pure matrix. Anal. Biochem. 2005, 343, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Marx, K.A.; Zhou, T.; Montrone, A.; McIntosh, D.; Braunhut, S.J. A comparative study of the cytoskeleton binding drugs nocodazole and taxol with a mammalian cell quartz crystal microbalance biosensor: Different dynamic responses and energy dissipation effects. Anal. Biochem. 2007, 361, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Marx, K.A.; Zhou, T.; Montrone, A.; Schulze, H.; Braunhut, S.J. A quartz crystal microbalance cell biosensor: Detection of microtubule alterations in living cells at nM nocodazole concentrations. Biosens. Bioelectron. 2001, 16, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Marx, K.A.; Warren, M.; Schulze, H.; Braunhut, S.J. The quartz crystal microbalance as a continuous monitoring tool for the study of endothelial cell surface attachment and growth. Biotechnol. Prog. 2000, 16, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Warrick, J.W.; Young, E.W.K.; Schmuck, E.G.; Saupe, K.W.; Beebe, D.J. High-content adhesion assay to address limited cell samples. Integr. Biol. 2013, 5, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, A.; Stamp, M.; Kmeth, R.; Buchegger, S.; Stritzker, B.; Saldamli, B.; Burgkart, R.; Schneider, M.F.; Wixforth, A. A novel tool for dynamic cell adhesion studies-the De-Adhesion number investigator DANI. Lab Chip 2014, 14, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Sraj, I.A.; Konstantopoulos, K.; Eggleton, C.D. Multi-scale simulation of l-selectin-PSGL-1-dependent homotypic leukocyte binding and rupture. Biomech. Model. Mechanobiol. 2010, 9, 613–627. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, P.; Golé, L.; Rieu, J.-P.; Vézy, C.; Ferrigno, R.; Mertani, H.C.; Rivière, C. A tapered channel microfluidic device for comprehensive cell adhesion analysis, using measurements of detachment kinetics and shear stress-dependent motion. Biomicrofluidics 2012, 6, 14107–1410712. [Google Scholar] [CrossRef] [PubMed]
- Alapan, Y.; Little, J.A.; Gurkan, U.A. Heterogeneous red blood cell adhesion and deformability in sickle cell disease. Sci. Rep. 2014, 4, 7173. [Google Scholar] [CrossRef] [PubMed]
- Chaw, K.C.; Manimaran, M.; Tay, E.H.; Swaminathan, S. Multi-step microfluidic device for studying cancer metastasis. Lab Chip 2007, 7, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Nalayanda, D.D.; Wang, Q.; Fulton, W.B.; Wang, T.-H.; Abdullah, F. Engineering an artificial alveolar-capillary membrane: A novel continuously perfused model within microchannels. J. Pediatr. Surg. 2010, 45, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Song, J.W.; Cavnar, S.P.; Walker, A.C.; Luker, K.E.; Gupta, M.; Tung, Y.-C.; Luker, G.D.; Takayama, S. Microfluidic endothelium for studying the intravascular adhesion of metastatic breast cancer cells. PLoS ONE 2009, 4, e5756. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Kunz, R.; Wu, J.; Dong, C. Study of local hydrodynamic environment in cell-substrate adhesion using side-view μPIV technology. PLoS ONE 2012, 7, e30721. [Google Scholar] [CrossRef] [PubMed]
- Riahi, R.; Yang, Y.L.; Kim, H.; Jiang, L.; Wong, P.K.; Zohar, Y. A microfluidic model for organ-specific extravasation of circulating tumor cells. Biomicrofluidics 2014, 8, 024103. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Mishima, S.; Maruyama, N.; Sumita, M. Quantitative evaluation of cell attachment to glass, polystyrene, and fibronectin- or collagen-coated polystyrene by measurement of cell adhesive shear force and cell detachment energy. J. Biomed. Mater. Res. 2000, 50, 114–124. [Google Scholar] [CrossRef]
- Lee, C. The technique for measurement of cell adhesion force. J. Med. Biol. Eng. 2004, 24, 51–56. [Google Scholar]
- Yang, S.-P.; Yang, C.-Y.; Lee, T.-M.; Lui, T.-S. Effects of calcium-phosphate topography on osteoblast mechanobiology determined using a cytodetacher. Mater. Sci. Eng. 2012, 32, 254–262. [Google Scholar] [CrossRef]
- Christ, K.V.; Williamson, K.B.; Masters, K.S.; Turner, K.T. Measurement of single-cell adhesion strength using a microfluidic assay. Biomed. Microdevices 2010, 12, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Hochmuth, R.M. Micropipette aspiration of living cells. J. Biomech. 2000, 33, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Wang, S.; Zhu, H.; Su, C.; Xu, G.; Lian, X. Using selected uniform cells in round shape with a micropipette to measure cell adhesion strength on silk fibroin-based materials. Mater. Sci. Eng. C 2008, 28, 1227–1235. [Google Scholar] [CrossRef]
- Rotsch, C.; Jacobson, K.; Radmacher, M. Dimensional and mechanical dynamics of active and stable edges in motile fibroblasts investigated by using atomic force microscopy. Proc. Natl. Acad. Sci. USA 1999, 96, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.J. AFM: A nanotool in membrane biology. Biochemistry 2008, 47, 7896–7898. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.; Gaub, H.E. Structure and mechanics of membrane proteins. Annu. Rev. Biochem. 2008, 77, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Hansma, H.G.; Kasuya, K.O.E. Atomic force microscopy imaging and pulling of nucleic acids. Curr. Opin. Struct. Biol. 2004, 14, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Hirano, Y.; Takahashi, H.; Kumeta, M.; Hizume, K.; Hirai, Y.; Otsuka, S.; Yoshimura, S.H.; Takeyasu, K. Nuclear architecture and chromatin dynamics revealed by atomic force microscopy in combination with biochemistry and cell biology. Pflügers Archiv 2008, 456, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Pesen, D.; Hoh, J.H. Micromechanical architecture of the endothelial cell cortex. Biophys. J. 2005, 88, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Collinsworth, A.M.; Zhang, S.; Kraus, W.E.; Truskey, G.A. Apparent elastic modulus and hysteresis of skeletal muscle cells throughout differentiation. Am. J. Physiol. Cell Physiol. 2002, 283, C1219–C1227. [Google Scholar] [CrossRef] [PubMed]
- Solon, J.; Levental, I.; Sengupta, K.; Georges, P.C.; Janmey, P.A. Fibroblast adaptation and stiffness matching to soft elastic substrates. Biophys. J. 2007, 93, 4453–4461. [Google Scholar] [CrossRef] [PubMed]
- Rotsch, C.; Braet, F.; Wisse, E.; Radmacher, M. AFM imaging and elasticity measurements on living rat liver macrophages. Cell Biol. Int. 1997, 21, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-C.; Su, H.-W.; Lee, C.-C.; Tang, M.-J.; Su, F.-C. Quantitative measurement of changes in adhesion force involving focal adhesion kinase during cell attachment, spread, and migration. Biochem. Biophys. Res. Commun. 2005, 329, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Kim, W. A Micro-Aspirator Chip Using Vacuum Expanded Microchannels for High-Throughput Mechanical Characterization of Biological Cells; The Royal Society of Chemistry Publishing: London, UK, 2010; pp. 1–86. [Google Scholar]
- Puech, P.-H.; Taubenberger, A.; Ulrich, F.; Krieg, M.; Muller, D.J.; Heisenberg, C.-P. Measuring cell adhesion forces of primary gastrulating cells from zebrafish using atomic force microscopy. J. Cell Sci. 2005, 118, 4199–4206. [Google Scholar] [CrossRef] [PubMed]
- Weder, G.; Vörös, J.; Giazzon, M.; Matthey, N.; Heinzelmann, H.; Liley, M. Measuring cell adhesion forces during the cell cycle by force spectroscopy. Biointerphases 2009, 4, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, S.C.; Cohnen, A.; Ludwig, T.; Watzl, C. 2B4 engagement mediates rapid LFA-1 and actin-dependent NK cell adhesion to tumor cells as measured by single cell force spectroscopy. J. Immunol. 2011, 186, 2757–2764. [Google Scholar] [CrossRef] [PubMed]
- Beaussart, A.; El-Kirat-Chatel, S.; Sullan, R.M.A; Alsteens, D.; Herman, P.; Derclaye, S.; Dufrêne, Y.F. Quantifying the forces guiding microbial cell adhesion using single-cell force spectroscopy. Nat. Protoc. 2014, 9, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Gourier, C.; Jegou, A.; Husson, J.; Pincet, F. A Nanospring named erythrocyte. The biomembrane force probe. Cell. Mol. Bioeng. 2008, 1, 263–275. [Google Scholar] [CrossRef]
- Evans, E.; Ritchie, K.; Merkel, R. Sensitive force technique to probe molecular adhesion and structural linkages at biological interfaces. Biophys. J. 1995, 68, 2580–2587. [Google Scholar] [CrossRef]
- Evans, E.; Berk, D.; Leung, A. Detachment of agglutinin-bonded red blood cells. I. Forces to rupture molecular-point attachments. Biophys. J. 1991, 59, 838–848. [Google Scholar] [CrossRef]
- Evans, E.; Heinrich, V.; Leung, A.; Kinoshita, K. Nano- to microscale dynamics of P-selectin detachment from leukocyte interfaces. I. Membrane separation from the cytoskeleton. Biophys. J. 2005, 88, 2288–2298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, K.-K. Optical tweezers for single cells. J. R. Soc. Interface 2008, 5, 671–690. [Google Scholar] [CrossRef] [PubMed]
- Ashkin, A. Optical trapping and manipulation of neutral particles using lasers. Proc. Natl. Acad. Sci. USA 1997, 94, 4853–4860. [Google Scholar] [CrossRef] [PubMed]
- Thoumine, O.; Kocian, P.; Kottelat, A; Meister, J.J. Short-term binding of fibroblasts to fibronectin: Optical tweezers experiments and probabilistic analysis. Eur. Biophys. J. 2000, 29, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.-W.; Liao, B.-Y.; Huang, C.-C.; Hwang, W.-L.; Wang, D.-W.; Chiou, A.E.T.; Lin, C.-H. Applications of optical tweezers and an integrated force measurement module for biomedical research. Int. Soc. Opt. Photonics 2000, 4082, 213–221. [Google Scholar]
- Curtis, J.E.; Spatz, J.P. Getting a grip: Hyaluronan-mediated cellular adhesion. Int. Soc. Opt. Photonics 2004, 5514, 455–466. [Google Scholar]
- Castelain, M.; Pignon, F.; Piau, J.-M.; Magnin, A.; Mercier-Bonin, M.; Schmitz, P. Removal forces and adhesion properties of saccharomyces cerevisiae on glass substrates probed by optical tweezer. J. Chem. Phys. 2007, 127, 135104. [Google Scholar] [CrossRef] [PubMed]
- Castelain, M.; Rouxhet, P.G.; Pignon, F.; Magnin, A.; Piau, J.-M. Single-cell adhesion probed in-situ using optical tweezers: A case study with saccharomyces cerevisiae. J. Appl. Phys. 2012, 111, 114701. [Google Scholar] [CrossRef]
- Schwingel, M.; Bastmeyer, M. Force mapping during the formation and maturation of cell adhesion sites with multiple optical tweezers. PLoS ONE 2013, 8, e54850. [Google Scholar] [CrossRef] [PubMed]
- Askenasy, N.; Farkas, D.L. Optical imaging of PKH-labeled hematopoietic cells in recipient bone marrow in vivo. Stem Cells 2002, 20, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Castelain, M.; Pignon, F.; Piau, J.-M.; Magnin, A. The initial single yeast cell adhesion on glass via optical trapping and Derjaguin–Landau–Verwey–Overbeek predictions. J. Chem. Phys. 2008, 128, 135101. [Google Scholar] [CrossRef] [PubMed]
- Koo, L.Y.; Irvine, D.J.; Mayes, A.M.; Lauffenburger, D.A.; Griffith, L.G. Co-regulation of cell adhesion by nanoscale RGD organization and mechanical stimulus. J. Cell Sci. 2002, 115, 1423–1433. [Google Scholar] [PubMed]
- Giacomello, E.; Neumayer, J.; Colombatti, A.P.R. Centrifugal assay for fluorescence-based cell adhesion adapted to the analysis of ex vivo cells and capable of determining relative binding strengths. Biotechniques 1999, 26, 764–766. [Google Scholar]
- Channavajjala, L.S.; Eidsath, A.; Saxinger, W.C.A. Simple method for measurement of cell-substrate attachment forces: Application to HIV-1 Tat. J. Cell Sci. 1997, 110, 249–256. [Google Scholar] [PubMed]
- Capadona, J.R.; Collard, D.M.; García, A.J. Fibronectin adsorption and cell adhesion to mixed monolayers of tri(ethylene glycol)- and methyl-terminated alkanethiols. Langmuir 2002, 19, 1847–1852. [Google Scholar] [CrossRef]
- Keselowsky, B.G.; García, A.J. Quantitative methods for analysis of integrin binding and focal adhesion formation on biomaterial surfaces. Biomaterials 2005, 26, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Reyes, C.D.; García, A.J. A centrifugation cell adhesion assay for high-throughput screening of biomaterial surfaces. J. Biomed. Mater. Res. A 2003, 67, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Reyes, C.D.; Petrie, T.A.; García, A.J. Mixed extracellular matrix ligands synergistically modulate integrin adhesion and signaling. J. Cell. Physiol. 2008, 217, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Harbers, G.M.; Healy, K.E. The effect of ligand type and density on osteoblast adhesion, proliferation, and matrix mineralization. J. Biomed. Mater. Res. A 2005, 75, 855–869. [Google Scholar] [CrossRef] [PubMed]
- Harbers, G.M.; Gamble, L.J.; Irwin, E.F.; Castner, D.G.; Healy, K.E. Development and characterization of a high-throughput system for assessing cell-surface receptor-ligand engagement. Langmuir 2005, 21, 8374–8384. [Google Scholar] [CrossRef] [PubMed]
- Reutelingsperger, C.P.M.; van Gool, R.G.J.; Heijnen, V.; Frederik, P.; Lindhout, T. The rotating disc as a device to study the adhesive properties of endothelial cells under differential shear stresses. J. Mater. Sci. Mater. Med. 1994, 5, 361–367. [Google Scholar] [CrossRef]
- Gallant, N.D.; Michael, K.E.; Garcı, J. Cell adhesion strengthening: Contributions of adhesive area , integrin binding , and focal adhesion assembly. Mol. Biol. Cell 2005, 16, 4329–4340. [Google Scholar] [CrossRef] [PubMed]
- García, A.J.; Ducheyne, P.; Boettiger, D. Quantification of cell adhesion using a spinning disc device and application to surface-reactive materials. Biomaterials 1997, 18, 1091–1098. [Google Scholar] [CrossRef]
- García, A.J.; Ducheyne, P.; Boettiger, D. Effect of surface reaction stage on fibronectin-mediated adhesion of osteoblast-like cells to bioactive glass. J. Biomed. Mater. Res. 1998, 40, 48–56. [Google Scholar] [CrossRef]
- Deligianni, D.D.; Katsala, N.D.; Koutsoukos, P.G.; Missirlis, Y.F. Effect of surface roughness of hydroxyapatite on human bone marrow cell adhesion, proliferation, differentiation and detachment strength. Biomaterials 2000, 22, 87–96. [Google Scholar] [CrossRef]
- Deligianni, D.; Korovessis, P.; Porte-Derrieu, M.C.; Amedee, J. Fibronectin preadsorbed on hydroxyapatite together with rough surface structure increases osteoblasts’ adhesion “in vitro”: The theoretical usefulness of fibronectin preadsorption on hydroxyapatite to increase permanent stability and longevity in spine implant. J. Spinal Disord. Tech. 2005, 18, 257–262. [Google Scholar] [PubMed]
- Lee, M.H.; Adams, C.S.; Boettiger, D.; DeGrado, W.F.; Shapiro, I.M.; Composto, R.J.; Ducheyne, P. Adhesion of MC3T3-E1 cells to RGD peptides of different flanking residues: Detachment strength and correlation with long-term cellular function. J. Biomed. Mater. Res. A 2007, 81A, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Michael, K.E.; Dumbauld, D.W.; Burns, K.L.; Hanks, S.K.; García, A.J. Focal adhesion kinase modulates cell adhesion strengthening via integrin activation. Mol. Biol. Cell 2009, 20, 2508–2519. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Boettiger, D. A novel mode for integrin-mediated signaling: Tethering is required for phosphorylation of FAK Y397. Mol. Biol. Cell 2003, 14, 4306–4315. [Google Scholar] [CrossRef] [PubMed]
- Datta, A.; Huber, F.; Boettiger, D. Phosphorylation of B3 integrin controls ligand binding strength. J. Biol. Chem. 2002, 277, 3943–3949. [Google Scholar] [CrossRef] [PubMed]
- Lynch, L.; Vodyanik, P.I.; Boettiger, D.; Guvakova, M.A. Insulin-like growth factor I controls adhesion strength mediated by α5β1 integrins in motile carcinoma cells. Mol. Biol. Cell 2005, 16, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.; Boettiger, D. Control of intracellular signaling by modulation of fibronectin conformation at the cell-materials interface. Langmuir 2002, 19, 1723–1729. [Google Scholar] [CrossRef]
- Garcı́a, A.J.; Takagi, J.; Boettiger, D. Two-stage activation for α5β1 integrin binding to surface-adsorbed fibronectin. J. Biol. Chem. 1998, 273, 34710–34715. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.S.; DiMilla, P.A. Application of fluid mechanic and kinetic models to characterize mammalian cell detachment in a radial-flow chamber. Biotechnol. Bioeng. 1997, 55, 616–629. [Google Scholar] [CrossRef]
- Detry, J.G.; Deroanne, C.; Sindic, M. Hydrodynamic systems for assessing surface fouling, soil adherence and cleaning in laboratory installations. Biotechnol. Agron. Soc. Environ. 2009, 13, 427–439. [Google Scholar]
- Cozens-roberts, C.; Quinn, J.A.; Lauffenburger, D.A. Receptor-mediated adhesion phenomena model studies with the radial-flow detachment assay. Biophys. J. 1990, 58, 107–125. [Google Scholar] [CrossRef]
- Dimilla, P.A.; Stone, J.A.; Albelda, S.M.; Biology, S. Maximal migration of human smooth muscle cells on fibronectin and type IV collagen occurs at an intermediate attachment strength. J. Cell Biol. 1993, 122, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.S.; DiMilla, P.A. Examination of membrane rupture as a mechanism for mammalian cell detachment from fibronectin-coated biomaterials. J. Biomed. Mater. Res. A 2003, 67A, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.S.; DiMilla, P.A. Effect of adsorbed fibronectin concentration on cell adhesion and deformation under shear on hydrophobic surfaces. J. Biomed. Mater. Res. 2002, 59, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.S.; DiMilla, P.A. Comparison of converging and diverging radial flow for measuring cell adhesion. AIChE J. 1998, 44, 465–473. [Google Scholar] [CrossRef]
- Kreke, M.R.; Badami, A.S.; Brady, J.B.; Michael Akers, R.; Goldstein, A.S. Modulation of protein adsorption and cell adhesion by poly(allylamine hydrochloride) heparin films. Biomaterials 2005, 26, 2975–2981. [Google Scholar] [CrossRef] [PubMed]
- Rezania, A.; Healy, K.E. Biomimetic peptide surfaces that regulate adhesion, spreading, cytoskeletal organization, and mineralization of the matrix deposited by osteoblast-like cells. Biotechnol. Prog. 1999, 15, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Rezania, A.; Healy, K.E. Integrin subunits responsible for adhesion of human osteoblast-like cells to biomimetic peptide surfaces. J. Orthop. Res. 1999, 17, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Rezania, A.; Thomas, C.H.; Branger, A.B.; Waters, C.M.; Healy, K.E. The detachment strength and morphology of bone cells contacting materials modified with a peptide sequence found within bone sialoprotein. J. Biomed. Mater. Res. 1997, 37, 9–19. [Google Scholar] [CrossRef]
- Rezania, A.; Thomas, C.; Healy, K. A probabilistic approach to measure the strength of bone cell adhesion to chemically modified surfaces. Ann. Biomed. Eng. 1997, 25, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Bearinger, J.P.; Castner, D.G.; Golledge, S.L.; Rezania, A.; Hubchak, S.; Healy, K.E. P(AAm-co-EG) interpenetrating polymer networks grafted to oxide surfaces: Surface characterization, protein adsorption, and cell detachment studies. Langmuir 1997, 13, 5175–5183. [Google Scholar] [CrossRef]
- Brown, M.A.; Wallace, C.S.; Anamelechi, C.C.; Clermont, E.; Reichert, W.M.; Truskey, G.A. The use of mild trypsinization conditions in the detachment of endothelial cells to promote subsequent endothelialization on synthetic surfaces. Biomaterials 2007, 28, 3928–3935. [Google Scholar] [CrossRef] [PubMed]
- Sordel, T.; Kermarec-Marcel, F.; Garnier-Raveaud, S.; Glade, N.; Sauter-Starace, F.; Pudda, C.; Borella, M.; Plissonnier, M.; Chatelain, F.; Bruckert, F.; et al. Influence of glass and polymer coatings on CHO cell morphology and adhesion. Biomaterials 2007, 28, 1572–1584. [Google Scholar] [CrossRef] [PubMed]
- Gerszten, R.E.; Lim, Y.-C.; Ding, H.T.; Snapp, K.; Kansas, G.; Dichek, D.A.; Cabañas, C.; Sánchez-Madrid, F.; Gimbrone, M.A.; Rosenzweig, A.; et al. Adhesion of monocytes to vascular cell adhesion molecule-1–transduced human endothelial cells: Implications for atherogenesis. Circ. Res. 1998, 82, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Truskey, G.A.; Pirone, J.S. The effect of fluid shear stress upon cell adhesion to fibronectin-treated surfaces. J. Biomed. Mater. Res. 1990, 24, 1333–1353. [Google Scholar] [CrossRef] [PubMed]
- Van Kooten, T.G.; Schakenraad, J.M.; van der Mei, H.C.; Busscher, H.J. Development and use of a parallel-plate flow chamber for studying cellular adhesion to solid surfaces. J. Biomed. Mater. Res. 1992, 26, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Lane, W.O.; Jantzen, A.E.; Carlon, T.A; Jamiolkowski, R.M.; Grenet, J.E.; Ley, M.M.; Haseltine, J.M.; Galinat, L.J.; Lin, F.-H.; Allen, J.D.; et al. Parallel-plate flow chamber and continuous flow circuit to evaluate endothelial progenitor cells under laminar flow shear stress. J. Vis. Exp. 2012, 1–12. [Google Scholar]
- Xiao, Y.; Truskey, G.A. Effect of receptor-ligand affinity on the strength of endothelial cell adhesion. Biophys. J. 1996, 71, 2869–2884. [Google Scholar] [CrossRef]
- Bhat, V.D.; Truskey, G.A.; Reichert, W.M. Using avidin-mediated binding to enhance initial endothelial cell attachment and spreading. J. Biomed. Mater. Res. 1998, 40, 57–65. [Google Scholar] [CrossRef]
- Chan, B.P.; Bhat, V.D.; Yegnasubramanian, S.; Reichert, W.M.; Truskey, G.A. An equilibrium model of endothelial cell adhesion via integrin-dependent and integrin-independent ligands. Biomaterials 1999, 20, 2395–2403. [Google Scholar] [CrossRef]
- Bhat, V.D.; Truskey, G.A.; Reichert, W.M. Fibronectin and avidin–biotin as a heterogeneous ligand system for enhanced endothelial cell adhesion. J. Biomed. Mater. Res. 1998, 41, 377–385. [Google Scholar] [CrossRef]
- Usami, S.; Chen, H.-H.; Zhao, Y.; Chien, S.; Skalak, R. Design and construction of a linear shear stress flow chamber. Ann. Biomed. Eng. 1993, 21, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Usami, S.; Dong, C. Development of a side-view chamber for studying cell-surface adhesion under flow conditions. Ann. Biomed. Eng. 1997, 25, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Heilshorn, S.C.; DiZio, K.A.; Welsh, E.R.; Tirrell, D.A. Endothelial cell adhesion to the fibronectin CS5 domain in artificial extracellular matrix proteins. Biomaterials 2003, 24, 4245–4252. [Google Scholar] [CrossRef]
- Wan, Y.; Yang, J.; Yang, J.; Bei, J.; Wang, S. Cell adhesion on gaseous plasma modified poly-(l-lactide) surface under shear stress field. Biomaterials 2003, 24, 3757–3764. [Google Scholar] [CrossRef]
- Renshaw, K.M.; Orr, D.E.; Burg, K.J.L. Design and evaluation of a novel flow chamber for measuring cell adhesion to absorbable polymer films. Biotechnol. Prog. 2005, 21, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wan, Y.; Yang, J.; Bei, J.; Wang, S. Plasma-treated, collagen-anchored polylactone: Its cell affinity evaluation under shear or shear-free conditions. J. Biomed. Mater. Res. A 2003, 67A, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Boura, C.; Muller, S.; Vautier, D.; Dumas, D.; Schaaf, P.; Claude Voegel, J.; François Stoltz, J.; Menu, P. Endothelial cell—Interactions with polyelectrolyte multilayer films. Biomaterials 2005, 26, 4568–4575. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, S.J.; Khang, G.; Lee, H.B. The effect of fluid shear stress on endothelial cell adhesiveness to polymer surfaces with wettability gradient. J. Colloid Interf. Sci. 2000, 230, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Van Kooten, T.G.; Schakenraad , J.M.; van der Mei, H.C.; Busscher, H.J. Influence of Substratum Wettability on the Strength of Adhesion of Human Fibroblasts. Biomaterials 1992, 13, 897–904. [Google Scholar] [CrossRef]
- Truskey, G.A.; Proulx, T.L. Relationship between 3T3 cell spreading and the strength of adhesion on glass and silane surfaces. Biomaterials 1993, 14, 243–254. [Google Scholar] [CrossRef]
- Vogel, J.; Bendas, G.; Bakowsky, U.; Hummel, G.; Schmidt, R.R.; Kettmann, U.; Rothe, U. The role of glycolipids in mediating cell adhesion: A flow chamber Study 1. Biochim. Biophys. Acta 1998, 1372, 205–215. [Google Scholar] [CrossRef]
- Palange, A.L.; di Mascolo, D.; Singh, J.; de Franceschi, M.S.; Carallo, C.; Gnasso, A.; Decuzzi, P. Modulating the vascular behavior of Mmetastatic breast cancer cells by curcumin treatment. Front. Oncol. 2012, 2, 161. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Whitesides, G.M. Soft lithography. Angew. Chem. Int. Ed. 1998, 37, 550–575. [Google Scholar] [CrossRef]
- Young, E.W.K.; Wheeler, A.R.; Simmons, C.A. Matrix-dependent adhesion of vascular and valvular endothelial cells in microfluidic channels. Lab Chip 2007, 7, 1759–1766. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Koo, L.Y.; Wang, W.M.; Lauffenburger, D.A; Griffith, L.G.; Jensen, K.F. Microfluidic shear devices for quantitative analysis of cell adhesion. Anal. Chem. 2004, 76, 5257–5264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jones, P.; Haswell, S. Attachment and detachment of living cells on modified microchannel surfaces in a microfluidic-based lab-on-a-chip system. Chem. Eng. J. 2008, 135, S82–S88. [Google Scholar] [CrossRef]
- Tang, Z.; Akiyama, Y.; Itoga, K.; Kobayashi, J.; Yamato, M.; Okano, T. Shear stress-dependent cell detachment from temperature-responsive cell culture surfaces in a microfluidic device. Biomaterials 2012, 33, 7405–7411. [Google Scholar] [CrossRef] [PubMed]
- Merkel, R.; Nassoy, P.; Leung, A.; Ritchie, K.; Evans, E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature 1999, 397, 50–53. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Mohandas, N. Disorders of red cell membrane. Br. J. Haematol. 2008, 141, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, N.; Gallagher, P.G. Red cell membrane: Past, present, and future. Blood 2008, 112, 3939–3949. [Google Scholar] [CrossRef] [PubMed]
- Correlates, M. Sickle erythrocyte adherence to vascular endothelium morphologic correlates and the requirement for divalent cations and collagen-binding plasma proteins. J. Clin. Investig. 1985, 76, 1605–1612. [Google Scholar]
- Hebbel, R.P.; Boogaerts, M.A.B.; Eaton, J.W.; Steinberg, M.H. Erythrocyte adherence to endothelium in sickle-cell anemia. N. Engl. J. Med. 1980, 302, 992–995. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.Y.H.; Lim, C.T. Biomechanics approaches to studying human diseases. Trends Biotechnol. 2007, 25, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Girdhar, G.; Shao, J.-Y. Simultaneous tether extraction from endothelial cells and leukocytes: Observation, mechanics, and significance. Biophys. J. 2007, 93, 4041–4052. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, D.K.; Ramis-Conde, I.; Chaplain, M.A.J. Multi-scale modelling of the dynamics of cell colonies: Insights into cell-adhesion forces and cancer invasion from in silico simulations. J. R. Soc. Interface 2014, 12, 20141080. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, V.; Mrksich, M.; Mcmeeking, R.; Evans, A. A bio-mechanical model for coupling cell contractility with focal adhesion formation. J. Mech. Phys. Solids 2008, 56, 1484–1510. [Google Scholar] [CrossRef]
- Cheung, L.S.-L.; Zheng, X.; Wang, L.; Baygents, J.C.; Guzman, R.; Schroeder, J.A.; Heimark, R.L.; Zohar, Y. Adhesion dynamics of circulating tumor cells under shear flow in a bio-functionalized microchannel. J. Micromech. Microeng. 2011, 21, 054033. [Google Scholar] [CrossRef]
- Kong, D.; Ji, B.; Dai, L. Nonlinear mechanical modeling of cell adhesion. J. Theor. Biol. 2008, 250, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Hodges, S.R.; Jensen, O.E. Spreading and peeling dynamics in a model of cell adhesion. J. Fluid Mech. 2002, 460, 381–409. [Google Scholar] [CrossRef]
- Kong, D.; Ji, B.; Dai, L. Stability of adhesion clusters and cell reorientation under lateral cyclic tension. Biophys. J. 2008, 95, 4034–4044. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, A.; Besser, A.; Safran, S.A. Dynamics of cellular focal adhesions on deformable substrates: Consequences for cell force microscopy. Biophys. J. 2008, 95, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Gallant, N.D.; García, A.J. Model of integrin-mediated cell adhesion strengthening. J. Biomech. 2007, 40, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Ronan, W.; Deshpande, V.S.; McMeeking, R.M.; McGarry, J.P. Numerical investigation of the active role of the actin cytoskeleton in the compression resistance of cells. J. Mech. Behav. Biomed. Mater. 2012, 14, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Ji, B.; Dai, L. Stabilizing to disruptive transition of focal adhesion response to mechanical forces. J. Biomech. 2010, 43, 2524–2529. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalili, A.A.; Ahmad, M.R. A Review of Cell Adhesion Studies for Biomedical and Biological Applications. Int. J. Mol. Sci. 2015, 16, 18149-18184. https://doi.org/10.3390/ijms160818149
Khalili AA, Ahmad MR. A Review of Cell Adhesion Studies for Biomedical and Biological Applications. International Journal of Molecular Sciences. 2015; 16(8):18149-18184. https://doi.org/10.3390/ijms160818149
Chicago/Turabian StyleKhalili, Amelia Ahmad, and Mohd Ridzuan Ahmad. 2015. "A Review of Cell Adhesion Studies for Biomedical and Biological Applications" International Journal of Molecular Sciences 16, no. 8: 18149-18184. https://doi.org/10.3390/ijms160818149
APA StyleKhalili, A. A., & Ahmad, M. R. (2015). A Review of Cell Adhesion Studies for Biomedical and Biological Applications. International Journal of Molecular Sciences, 16(8), 18149-18184. https://doi.org/10.3390/ijms160818149