
Interleukin-17A Gene Expression in Morbidly Obese Women


Abstract
:
1. Introduction
2. Results
2.1. Baseline Characteristics of Subjects

{kind=link}
{kind=link}
{kind=link}
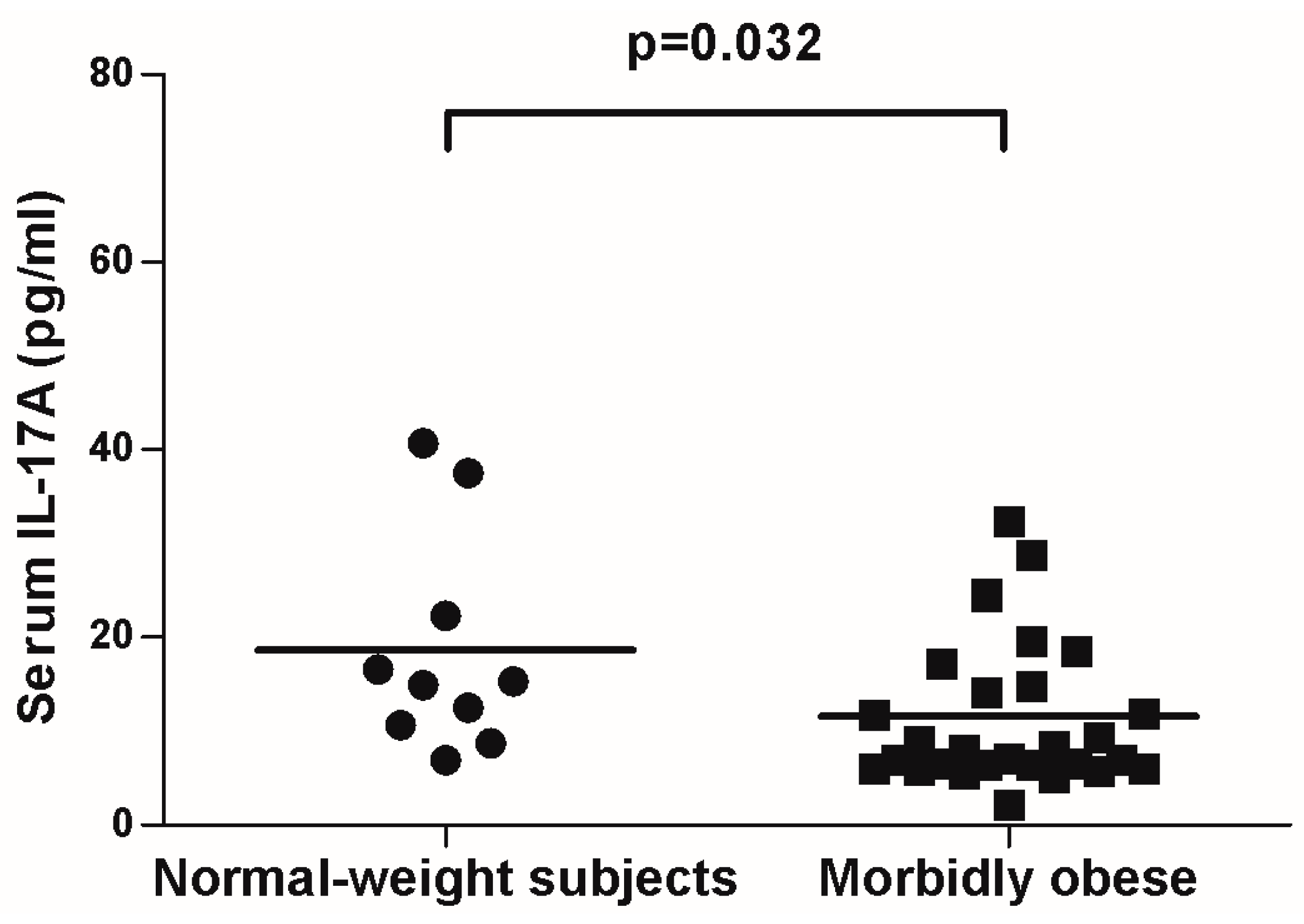
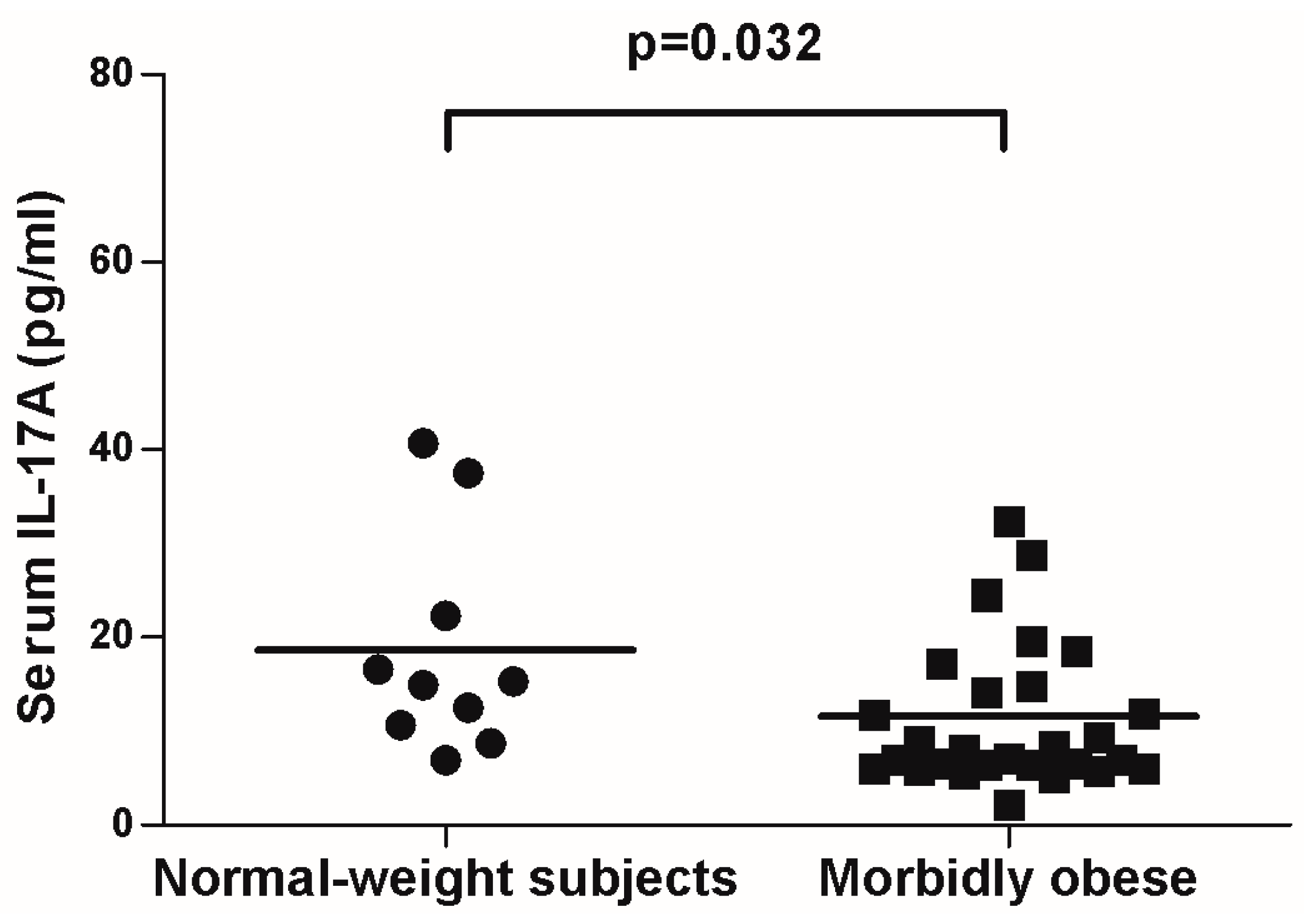{kind=link}
| Variables | Normal-Weight Control (n = 10) | Morbidly Obese (n = 30) |
|---|---|---|
| Mean ± SD | Mean ± SD | |
| Age (years) | 43.70 ± 12.35 | 47.00 ± 7.35 |
| Weight (kg) | 56.30 ± 8.64 | 123.69 ± 13.18 * |
| WC (cm) | 68.60 ± 10.5 | 135.16 ± 11.32 * |
| BMI (kg/m2) | 22.61 ± 1.91 | 47.21 ± 5.4 * |
| SBP (mmHg) | 119.88 ± 13.08 | 139.10 ± 14.78 * |
| DBP (mmHg) | 67.55 ± 7.57 | 80.26 ± 14.76 * |
| Glucose (mg/dL) | 88.11 ± 9.63 | 115.5 ± 26.12 * |
| Insulin (mU/L) | 7.93 ± 5.86 | 20.26 ± 13.99 * |
| HbA1c (%) | 4.55 ± 0.28 | 5.7 ± 1.22 * |
| HOMA2-IR | 1.07 ± 0.83 | 2.69 ± 1.77 * |
| Cholesterol (mg/dL) | 172.68 ± 25.91 | 179.39 ± 35.22 |
| HDL-C (mg/dL) | 54.66 ± 17.85 | 41.26 ± 7.52 |
| LDL-C (mg/dL) | 93.77 ± 26.74 | 106.13 ± 28.64 |
| Triglycerides (mg/dL) | 121 ± 79.32 | 159.6 ± 53.25 |
| Adipo/cytokine circulating levels | ||
| Adiponectin (µg/mL) | 11.48 ± 6.13 | 6.77 ± 2.84 * |
| IL-6 (pg/mL) | 1.78 ± 1.55 | 2.95 ± 1.56 |
| Lipocalin-2 (ng/mL) | 63.53 ± 28.33 | 82.26 ± 29.85 |
| Resistin (ng/mL) | 3.61 ± 1.31 | 4.52 ± 1.63 |
| TNFRII (ng/mL) | 3.09 ± 1.51 | 5.19 ± 2.37 * |
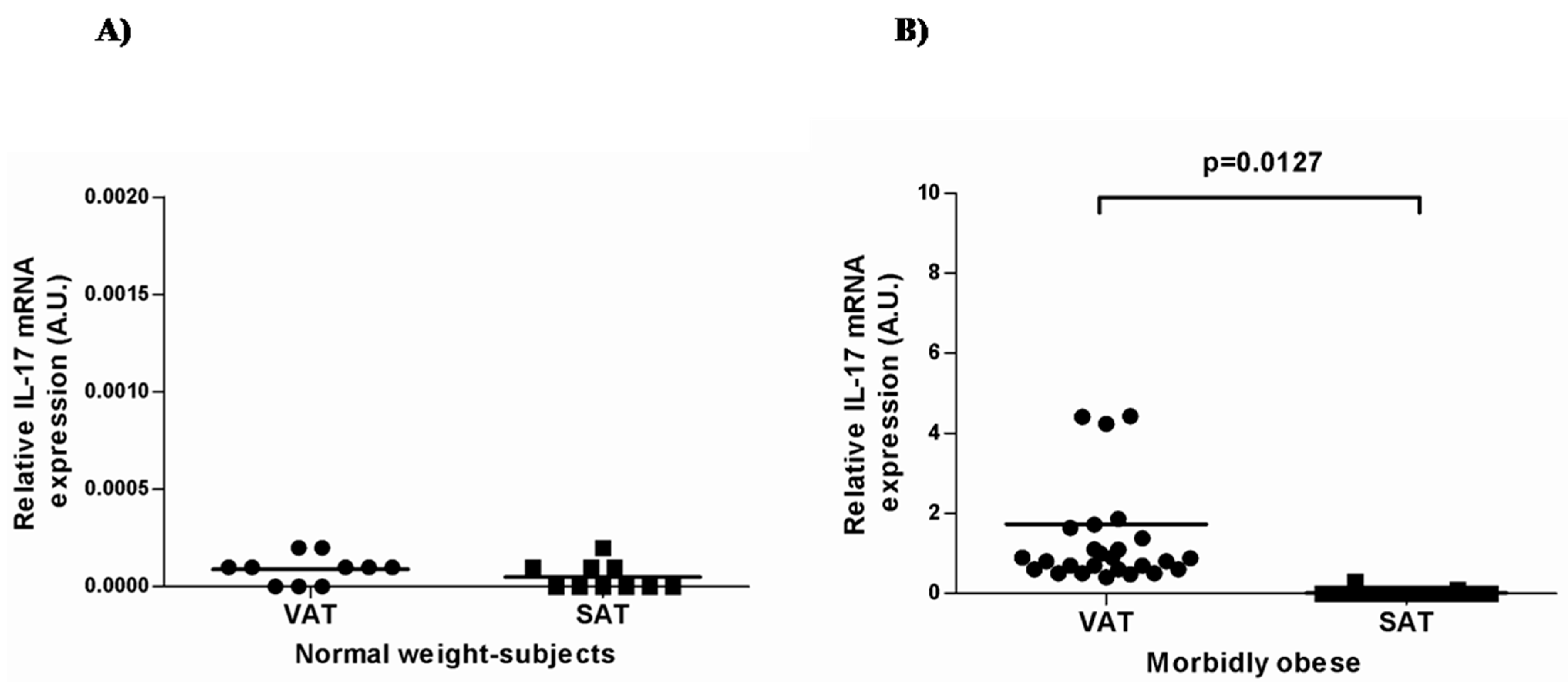
2.2. IL-17A and Adipo/Cytokine mRNA Expression in Adipose Tissue
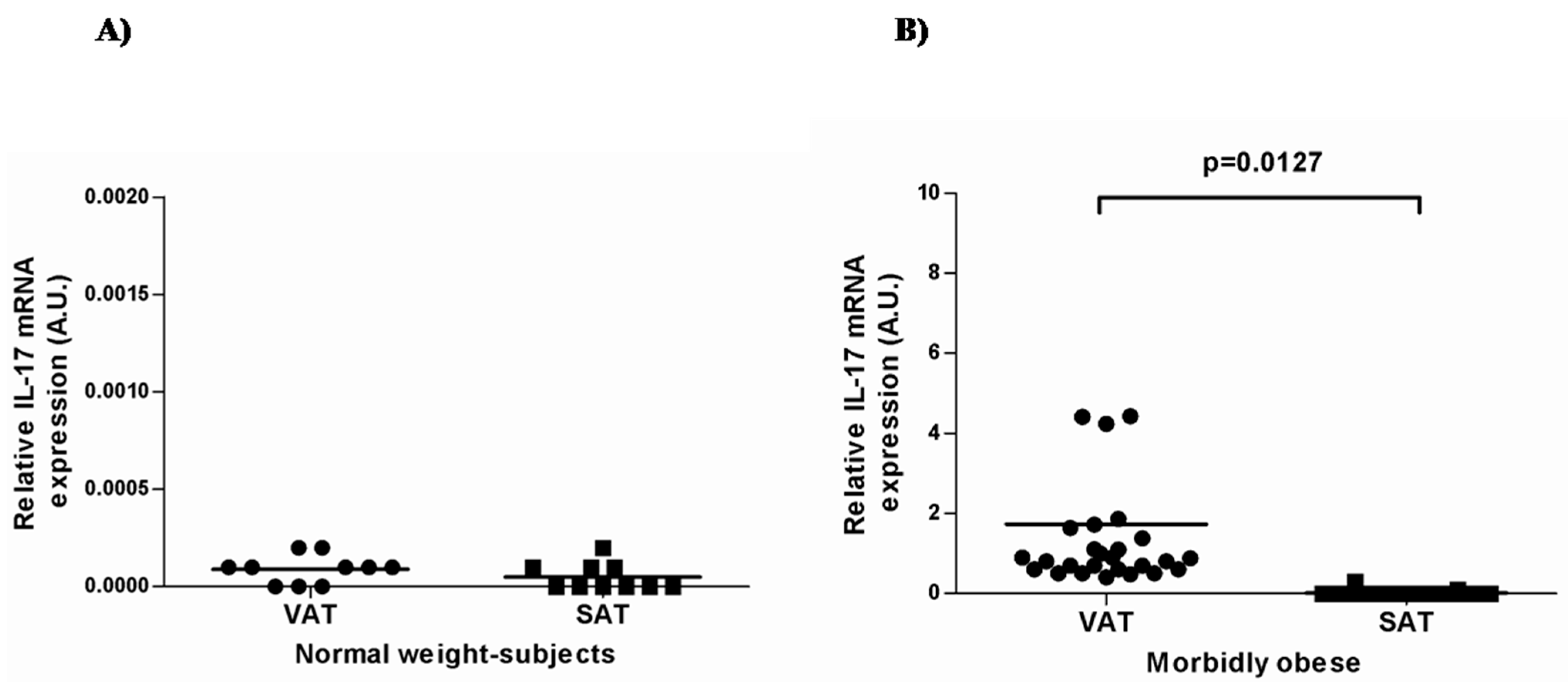
| Gene Expression | Normal-Weight Control (n = 10) | Morbidly Obese (n = 30) | ||
|---|---|---|---|---|
| VAT | SAT | VAT | SAT | |
| Mean ± SD | Mean ± SD | Mean ± SD | Mean ± SD | |
| IL-6 | 2.38 ± 1.52 | 17. 07 ± 11.66 | 61.09 ± 14.13 * | 86.40 ± 28.71 * |
| Adiponectin | 0.77 ± 0.32 | 0.58 ± 0.30 | 0.53 ± 0.27 | 0.29 ± 0.17 * |
| Lipocalin-2 | 0.006 ± 0.003 | 0.029 ± 0.02 | 0.02 ± 0.01 | 0.064 ± 0.03 |
| TNFα | 0.039 ± 0.02 | 0.076 ± 0.06 | 0.26 ± 0.20 * | 0.043 ± 0.02 |
| Resistin | 0.008 ± 0.004 | 0.016 ± 0.005 | 0.030 ± 0.02 * | 0.030 ± 0.02 |
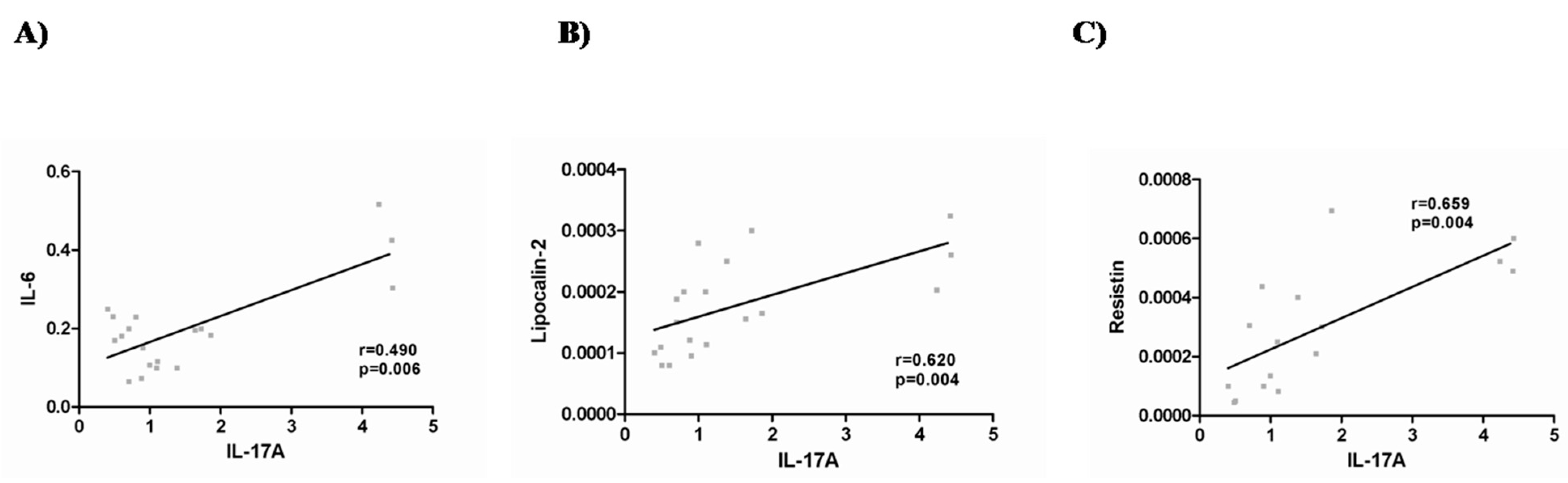
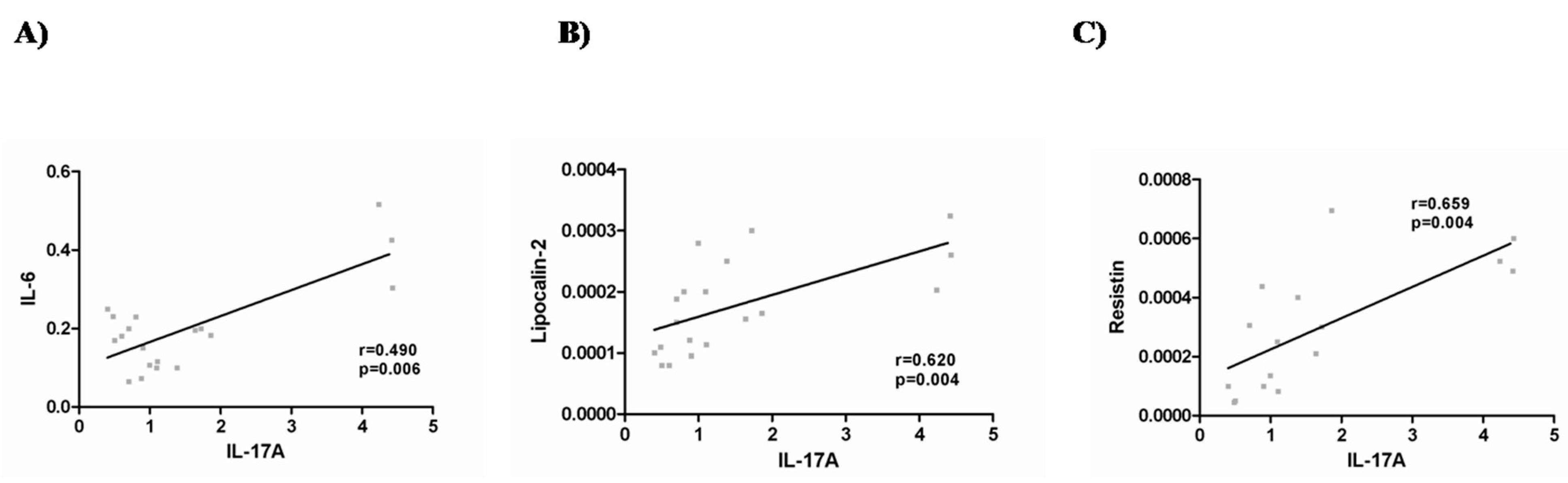
2.3. IL-17A and Adipo/Cytokine Circulating Levels
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Biochemical Analyses
4.3. RNA Isolation and Real-Time PCR
4.4. IL-17A and Adipo/Cytokine Serum Levels
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| BMI | body mass index |
| VAT | visceral adipose tissue |
| SAT | subcutaneous adipose tissue |
| MO | morbidly obese |
| IL-17A | interleukin-17A |
| IL-6 | interleukin 6 |
| HbA1c | glycosylated hemoglobin |
| HOMA2-IR | homeostasis model assessment of insulin resistance |
| HDL-C | high density lipoprotein |
| LDL-C | low density lipoprotein |
References
- Knight, J.A. Diseases and disorders associated with excess body weight. Ann. Clin. Lab. Sci. 2011, 41, 107–121. [Google Scholar] [PubMed]
- Sartipy, P.; Loskutoff, D.J. Expression profiling identifies genes that continue to respond to insulin in adipocytes made insulin-resistant by treatment with tumor necrosis factor-alpha. J. Biol. Chem. 2003, 278, 52298–52306. [Google Scholar] [CrossRef] [PubMed]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The hormone resistin links obesity to diabetes. Nature 2001, 409, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Cramer, T.; Johnson, R.S. A novel role for the hypoxia inducible transcription factor HIF-1alpha: Critical regulation of inflammatory cell function. Cell Cycle 2003, 3, 92–93. [Google Scholar] [CrossRef]
- Ozcan, U.; Cao, Q.; Yilmaz, E.; Lee, A.H.; Iwakoshi, N.N.; Ozdelen, E.; Tuncman, G.; Görgün, C.; Glimcher, L.H.; Hotamisligil, G.S. Endoplasmic reticulum stress links obesity, insulin action, and type 2 diabetes. Science 2004, 306, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.M.; Olefsky, J.M. The origins and drivers of insulin resistance. Cell 2013, 152, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Ozcan, U. Unfolded protein response signaling and metabolic diseases. J. Biol. Chem. 2014, 289, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor–alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and insulin resistance. J. Clin. Investig. 2006, 116, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Olefsky, J.M.; Glass, C.K. Macrophages, inflammation, and insulin resistance. Annu. Rev. Physiol. 2010, 72, 219–246. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Auguet, T.; Guiu-Jurado, E.; Berlanga, A.; Terra, X.; Martinez, S.; Porras, J.A.; Ceausu, A.; Sabench, F.; Hernandez, M.; Aguilar, C.; et al. Downregulation of lipogenesis and fatty acid oxidation in the subcutaneous adipose tissue of morbidly obese women. Obesity 2014, 22, 2032–2038. [Google Scholar] [CrossRef] [PubMed]
- Ortega Martinez de Victoria, E.; Xu, X.; Koska, J.; Francisco, A.M.; Scalise, M.; Ferrante, A.W., Jr.; Krakoff, J. Macrophage content in subcutaneous adipose tissue: Associations with adiposity, age, inflammatory markers, and whole-body insulin action in healthy Pima Indians. Diabetes 2009, 58, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Hummasti, S.; Hotamisligil, G.S. Endoplasmic reticulum stress and inflammation in obesity and diabetes. Circ. Res. 2010, 107, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Kintscher, U.; Hartge, M.; Hess, K.; Foryst-Ludwig, A.; Clemenz, M.; Wabitsch, M.; Fischer-Posovszky, P.; Barth, T.F.; Dragun, D.; Skurk, T.; et al. T-lymphocyte infiltration in visceral adipose tissue: A primary event in adipose tissue inflammation and the development of obesity-mediated insulin resistance. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Divoux, A.; Sun, J.; Zhang, J.; Clement, K.; Glickman, J.N.; Sukhova, G.K.; Wolters, P.J.; Du, J.; Gorgun, C.Z.; et al. Genetic deficiency and pharmacological stabilization of mast cells reduce diet-induced obesity and diabetes in mice. Nat. Med. 2009, 15, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, K.; Ishimori, N.; Ohmura, Y.; Tokuhara, S.; Nozawa, A.; Horii, S.; Andoh, Y.; Fujii, S.; Iwabuchi, K.; Onoe, K.; et al. Natural killer T–cells areinvolved in adipose tissues inflammation and glucose intolerance in diet induced obese mice. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.; Strodthoff, D.; Robertson, A.K.; Paulsson-Berne, G.; Fauconnier, J.; Parini, P.; Ryden, M.; Thierry-Mieg, N.; Johansson, M.E.; Chibalin, A.V.; et al. T cell-mediated inflammation inadipose tissue does not cause insulin resistance in hyperlipidemic mice. Circ. Res. 2009, 104, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Afzali, B.; Lombardi, G.; Lechler, R.I.; Lord, G.M. The role of T helper 17 (Th17) and regulatory T cells (Treg) in human organ transplantation and autoimmune disease. Clin. Exp. Immunol. 2007, 148, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga, L.A.; Shen, W.J.; Joyce-Shaikh, B.; Pyatnova, E.A.; Richards, A.G.; Thom, C.; Andrade, S.M.; Cua, D.J.; Kraemer, F.B.; Butcher, E.C. IL-17A regulates adipogenesis, glucose homeostasis, and obesity. J. Immunol. 2010, 185, 6947–6959. [Google Scholar] [CrossRef] [PubMed]
- Sumarac-Dumanovic, M.; Stevanovic, D.; Ljubic, A.; Jorga, J.; Simic, M.; Stamenkovic-Pejkovic, D.; Starcevic, V.; Trajkovic, V.; Micic, D. Increased activity of interleukin-23/interleukin-17 proinflammatory axis in obese women. Int. J. Obes. 2009, 33, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Pini, M.; Fantuzzi, G. Enhanced production of IL-17A during zymosan-induced peritonitis in obese mice. J. Leukoc. Biol. 2010, 87, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, I.; Pingris, K.; Poitou, C.; Bessoles, S.; Venteclef, N.; Kiaf, B.; Beaudoin, L.; da Silva, J.; Allatif, O.; Rossjohn, J.; et al. Mucosal-associated invariant T cell alterations in obese and type 2 diabetic patients. J. Clin. Investig. 2015, 125, 1752–1762. [Google Scholar] [CrossRef] [PubMed]
- Dalmas, E.; Venteclef, N.; Caer, C.; Poitou, C.; Cremer, I.; Aron-Wisnewsky, J.; Lacroix-Desmazes, S.; Bayry, J.; Kaveri, S.V.; Clément, K.; et al. T cell-derived IL-22 amplifies IL-1β-driven inflammation in human adipose tissue: Relevance to obesity and type 2 diabetes. Diabetes 2014, 63, 1966–1977. [Google Scholar] [CrossRef] [PubMed]
- Fabbrini, E.; Cella, M.; McCartney, S.A.; Fuchs, A.; Abumrad, N.A.; Pietka, T.A.; Chen, Z.; Finck, B.N.; Han, D.H.; Magkos, F.; et al. Association between specific adipose tissue CD4+ T-cell populations and insulin resistance in obese individuals. Gastroenterology 2013, 145, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Carolan, E.; Tobin, L.M.; Mangan, B.A.; Corrigan, M.; Gaoatswe, G.; Byrne, G.; Geoghegan, J.; Cody, D.; O’Connell, J.; Winter, D.C.; et al. Altered distribution and increased IL-17 production by mucosal-associated invariant T cells in adult and childhood obesity. J. Immunol. 2015, 194, 5775–5780. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Aljada, A.; Ghanim, H.; Hofmeyer, D.; Chaudhuri, A. Increased plasma concentration of macrophage migration inhibitory factor (MIF) and MIF mRNA in mononuclear cells in the obese and the suppressive action of metformin. J. Clin. Endocrinol. Metab. 2004, 89, 5043–5047. [Google Scholar] [CrossRef] [PubMed]
- Setty, A.R.; Curhan, G.; Choi, H.K. Obesity, waist circumference, weight change, and the risk of psoriasis in women: Nurses’ Health Study II. Arch. Intern. Med. 2007, 167, 1670–1675. [Google Scholar] [CrossRef] [PubMed]
- Hass, D.J.; Brensinger, C.M.; Lewis, J.D.; Lichtenstein, G.R. The impact of increased body mass index on the clinical course of Crohn’s disease. Clin. Gastroenterol. Hepatol. 2006, 4, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Esser, N.; Legrand-Poels, S.; Piette, J.; Scheen, A.J.; Paquot, N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res. Clin. Pract. 2014, 105, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Bertola, A.; Ciucci, T.; Rousseau, D.; Bourlier, V.; Duffaut, C.; Bonnafous, S.; Blin-Wakkach, C.; Anty, R.; Iannelli, A.; Gugenheim, J.; et al. Identification of adipose tissue dendritic cells correlated with obesity-associated insulin-resistance and inducing Th17 responses in mice and patients. Diabetes 2012, 61, 2238–2247. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, F.; Santarlasci, V.; Maggi, L.; Cosmi, L.; Liotta, F.; Romagnani, S. Reasons for rarity of Th17 cells in inflammatory sites of human disorders. Semin. Immunol. 2013, 25, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Kissebah, A. Central obesity: Measurement and metabolic effects. Diabetes Rev. 1997, 5, 8–20. [Google Scholar]
- Després, J.P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Onishi, R.M.; Gaffen, S.L. Interleukin-17 and its target genes: Mechanisms of interleukin-17 function in disease. Immunology 2010, 129, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Gaffen, S.L. IL-17 in obesity and adipogenesis. Cytokine Growth Factor Rev. 2010, 21, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Gaffen, S.L. Structure and signalling in the IL-17 receptor family. Nat. Rev. Immunol. 2009, 9, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.E.; Hatton, R.D.; Mangan, P.R.; Murphy, K.M.; Weaver, C.T. Interleukin-17 producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immunol. 2005, 6, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Li, Z.; Yang, X.O.; Chang, S.H.; Nurieva, R.; Wang, Y.H.; Wang, Y.; Hood, L.; Zhu, Z.; Tian, Q.; et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat. Immunol. 2005, 6, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Ruddy, M.J.; Wong, G.C.; Liu, X.K.; Yamamoto, H.; Kasayama, S.; Kirkwood, K.L.; Gaffen, S.L. Functional cooperation between interleukin-17 and tumor necrosis factor-alpha is mediated by CCAAT/enhancer–binding protein family members. J. Biol. Chem. 2004, 279, 2559–2567. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Ruddy, M.J.; Plamondon, P.; Gaffen, S.L. Cytokines link osteoblasts and inflammation: Microarray analysis of interleukin-17and TNF-alpha-induced genes in bone cells. J. Leukoc. Biol. 2005, 77, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Auguet, T.; Quintero, Y.; Terra, X.; Martínez, S.; Lucas, A.; Pellitero, S.; Aguilar, C.; Hernández, M.; del Castillo, D.; Richart, C. Upregulation of lipocalin 2 in adipose tissues of severely obese women: Positive relationship with proinflammatory cytokines. Obesity 2011, 19, 2295–2300. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Jin, D.; Chen, X. Lipocalin 2 is a regulator of macrophage polarization and NF-κB/STAT3 pathway activation. Mol. Endocrinol. 2014, 28, 1616–1628. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, Y.; Zhang, Y.; Leroith, D.; Bernlohr, D.A.; Chen, X. The role of lipocalin 2 in the regulation of inflammation in adipocytes and macrophages. Mol. Endocrinol. 2008, 22, 1416–1426. [Google Scholar] [CrossRef] [PubMed]
- Terra, X.; Auguet, T.; Quesada, I.; Aguilar, C.; Luna, A.M.; Hernández, M.; Sabench, F.; Porras, J.A.; Martínez, S.; Lucas, A.; et al. Increased levels and adipose tissue expression of visfatin in morbidly obese women: The relationship with pro-inflammatory cytokines. Clin. Endocrinol. 2012, 77, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Al-Suhaimi, E.A.; Shehzad, A. Leptin, resistin and visfatin: The missing link between endocrine metabolic disorders and immunity. Eur. J. Med. Res. 2013, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Witowski, J.; Ksiazek, K.; Jörres, A. Interleukin-17: A mediator of inflammatory responses. Cell. Mol. Life Sci. 2004, 61, 567–579. [Google Scholar] [CrossRef] [PubMed]
- De Angulo, A.; Faris, R.; Daniel, B.; Jolly, C.; de Graffenried, L. Age-related increase in IL-17 activates pro-inflammatory signaling in prostate cells. Prostate 2015, 75, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Xuan, M.L.; Lu, C.J.; Han, L.; Xiang, Y. Circulating levels of inflammatory cytokines in patients with psoriasis vulgaris of different Chinese medicine syndromes. Chin. J. Integr. Med. 2015, 21, 108–114. [Google Scholar] [CrossRef] [PubMed]
- HOMA Calculator (Version 2.2.2). Available online: http://www.dtu.ox.ac.uk (accessed on 1 May 2010).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zapata-Gonzalez, F.; Auguet, T.; Aragonès, G.; Guiu-Jurado, E.; Berlanga, A.; Martinez, S.; Martí, A.; Sabench, F.; Hernandez, M.; Aguilar, C.; et al. Interleukin-17A Gene Expression in Morbidly Obese Women. Int. J. Mol. Sci. 2015, 16, 17469-17481. https://doi.org/10.3390/ijms160817469
Zapata-Gonzalez F, Auguet T, Aragonès G, Guiu-Jurado E, Berlanga A, Martinez S, Martí A, Sabench F, Hernandez M, Aguilar C, et al. Interleukin-17A Gene Expression in Morbidly Obese Women. International Journal of Molecular Sciences. 2015; 16(8):17469-17481. https://doi.org/10.3390/ijms160817469
Chicago/Turabian StyleZapata-Gonzalez, Fernando, Teresa Auguet, Gemma Aragonès, Esther Guiu-Jurado, Alba Berlanga, Salomé Martinez, Andreu Martí, Fátima Sabench, Mercé Hernandez, Carmen Aguilar, and et al. 2015. "Interleukin-17A Gene Expression in Morbidly Obese Women" International Journal of Molecular Sciences 16, no. 8: 17469-17481. https://doi.org/10.3390/ijms160817469